
Genomic Identification and Expression Profiling of Lesion Simulating Disease Genes in Alfalfa (Medicago sativa) Elucidate Their Responsiveness to Seed Vigor


Abstract
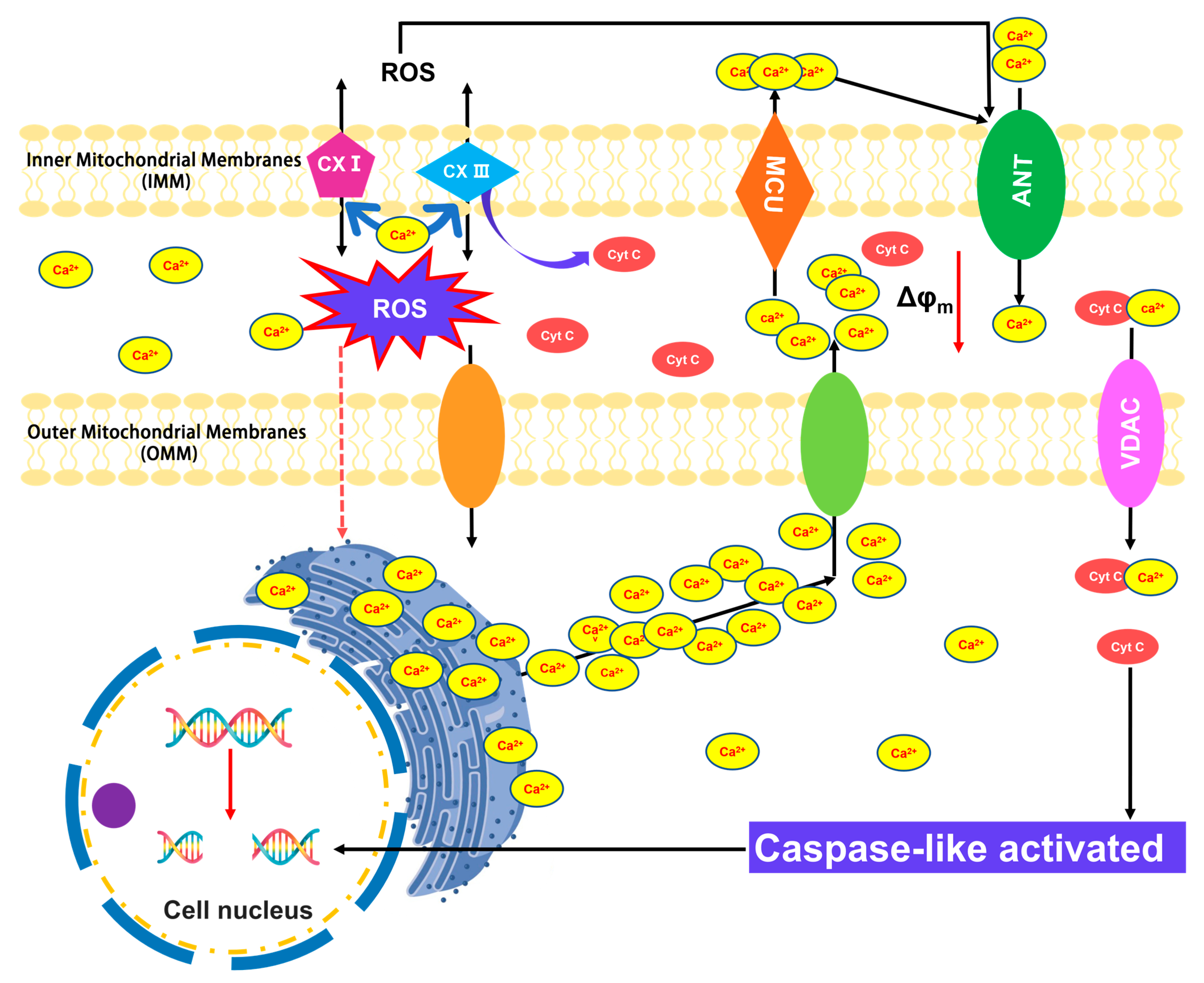
:1. Introduction
2. Materials and Methods
2.1. Identification of the MsLSD Gene Family in Alfalfa
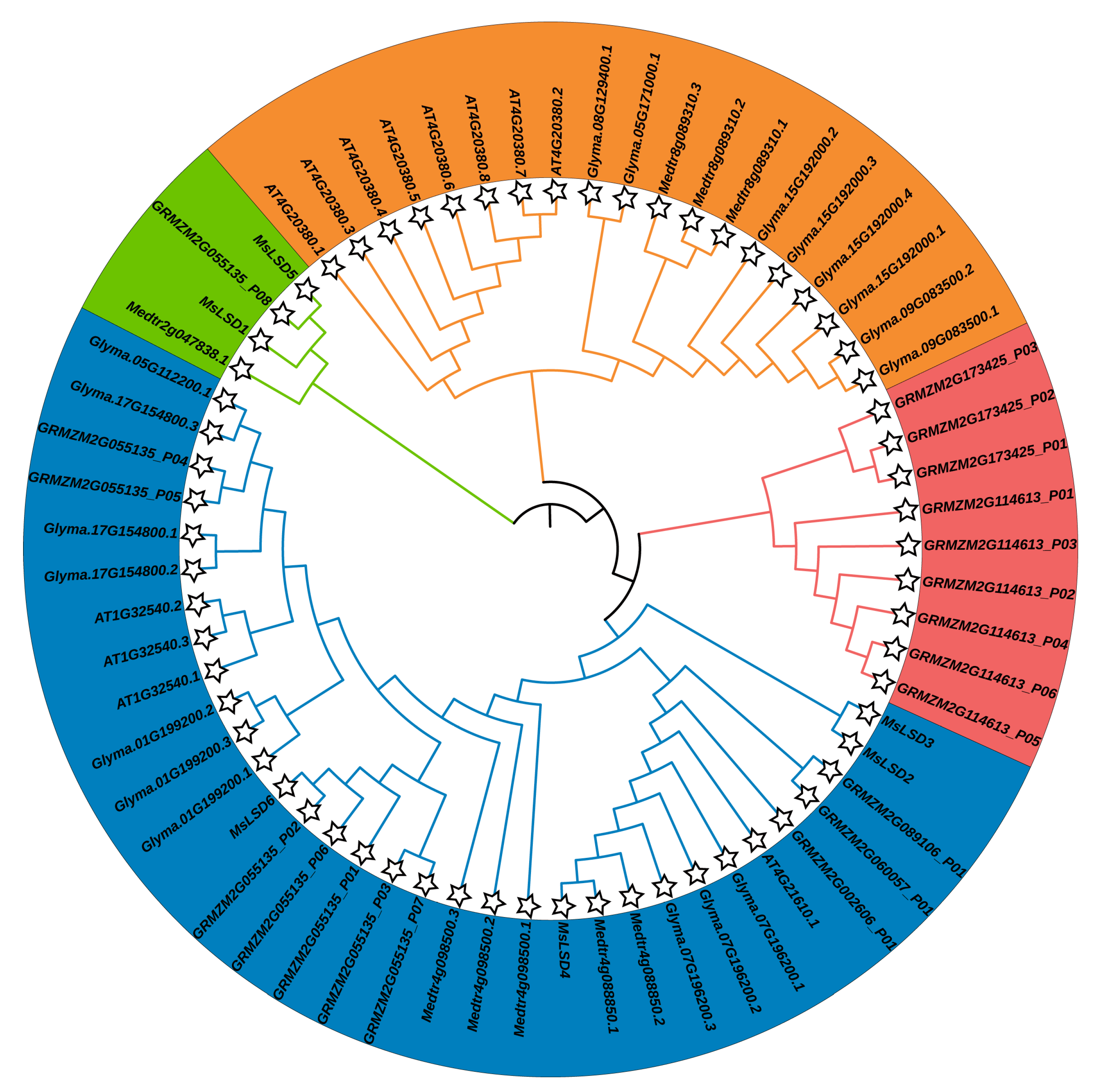
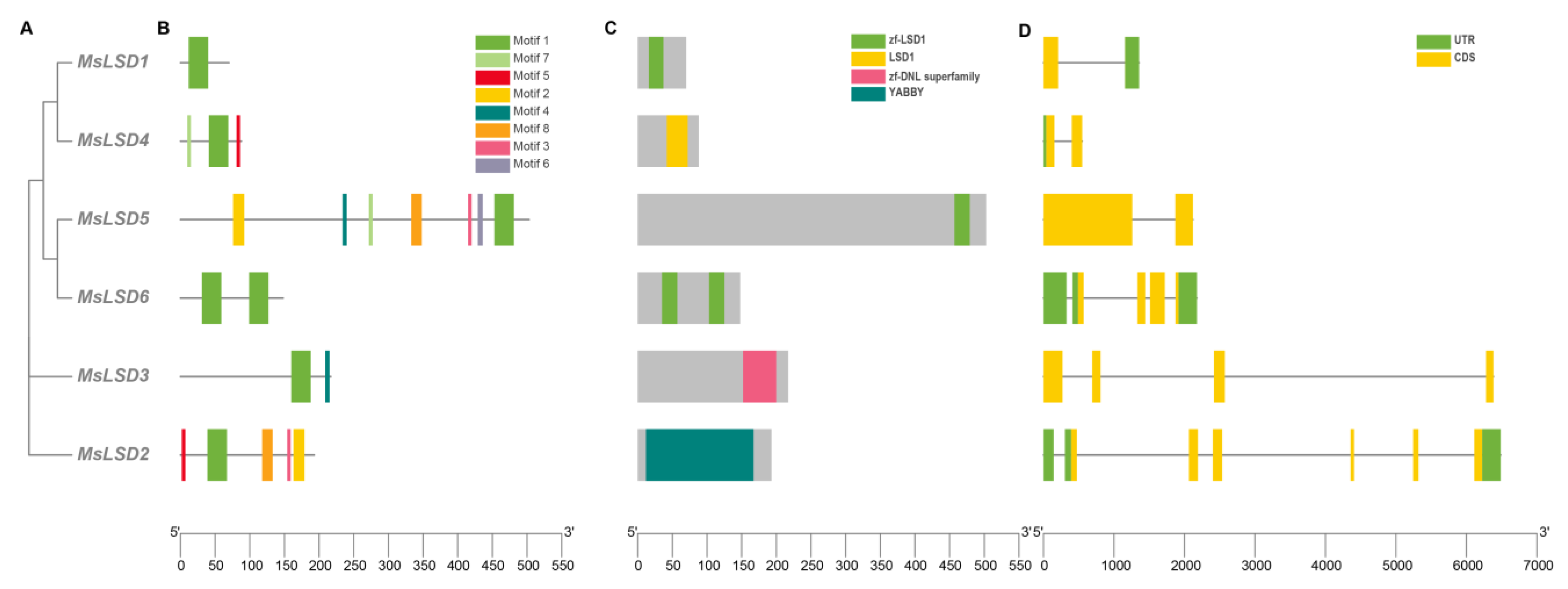
2.2. Phylogenetic, Gene Structure, and Conserved Motif Analyses of the MsLSD Members
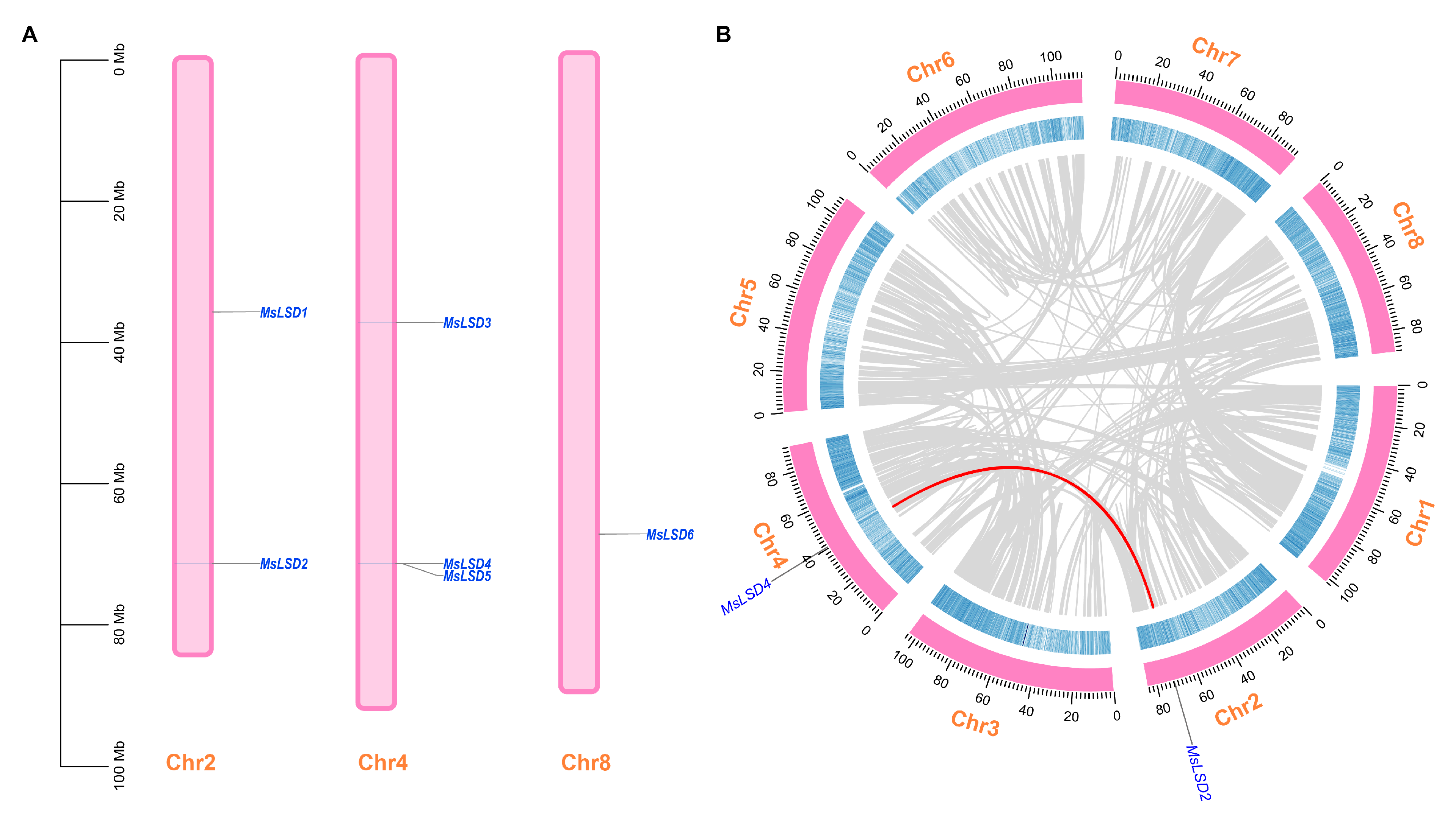
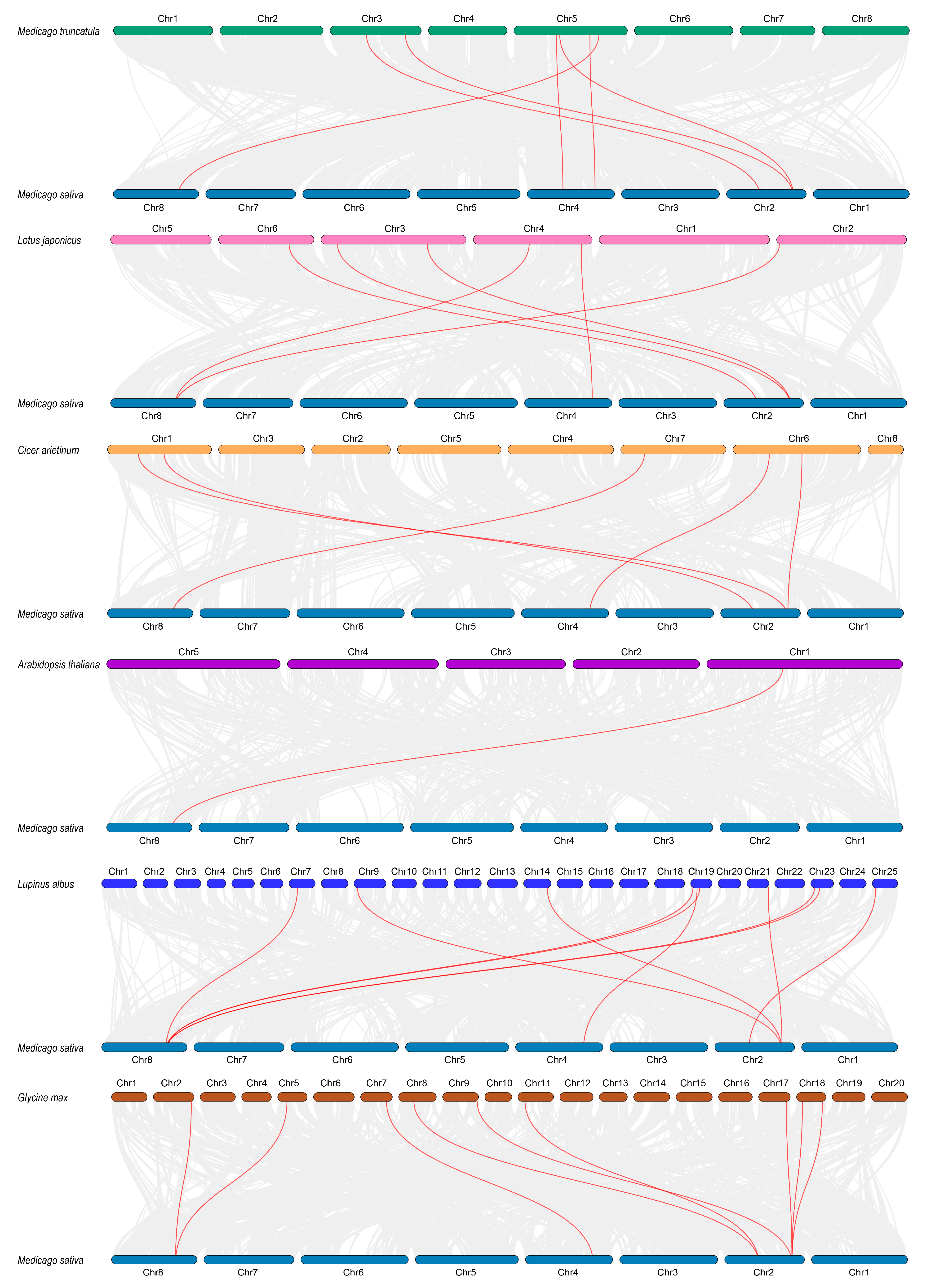
2.3. Gene Duplication and Chromosomal Mapping Analysis
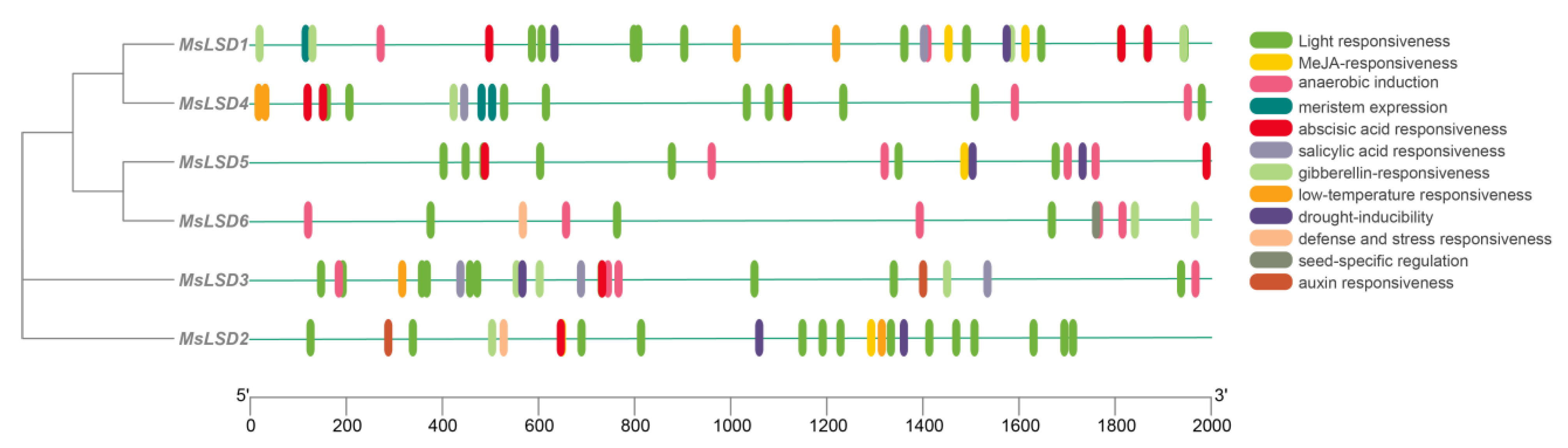
2.4. cis-Regulatory Element Analysis
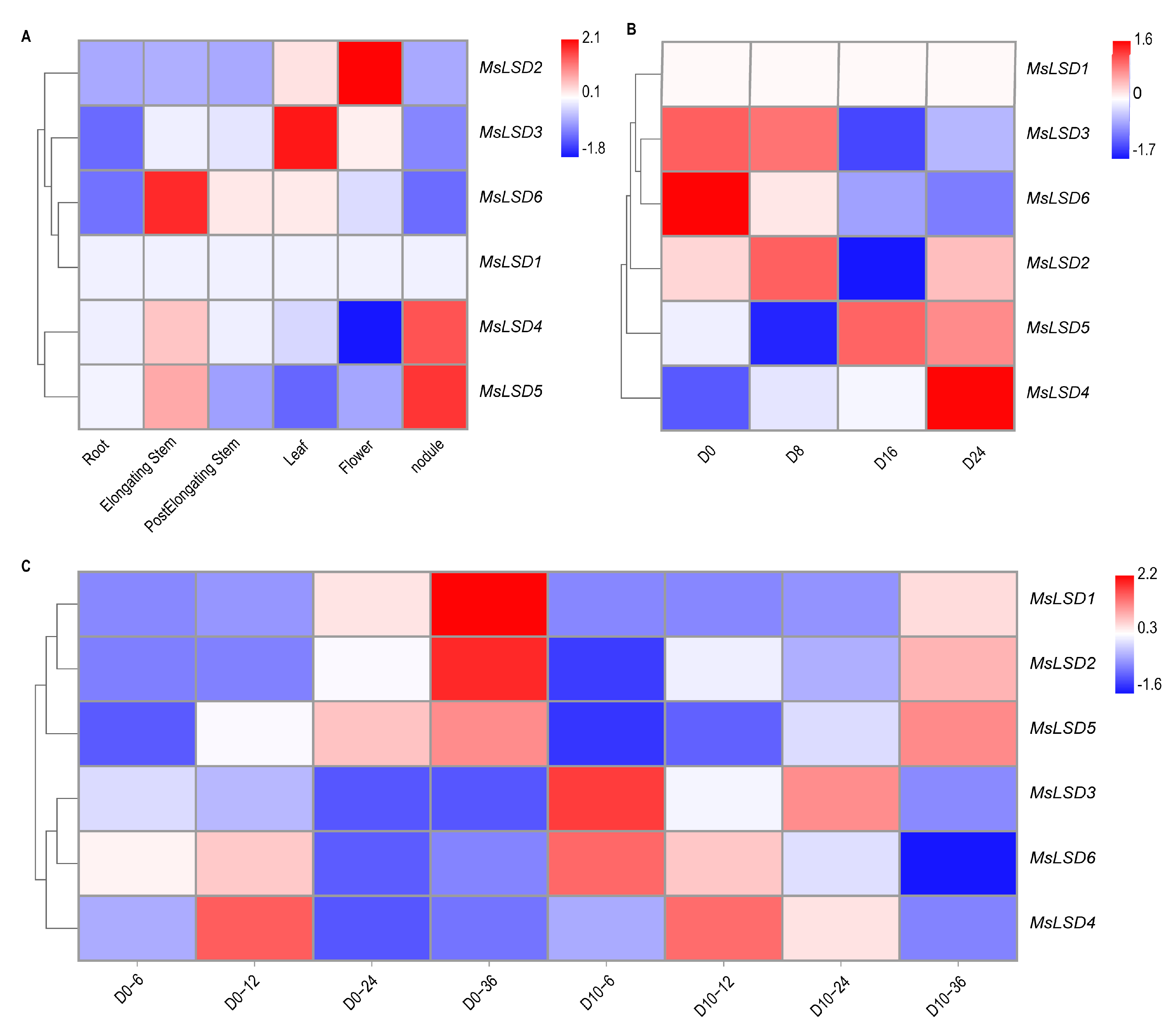
2.5. Tissue-Specific Expression Analysis of MsLSD Genes in Alfalfa
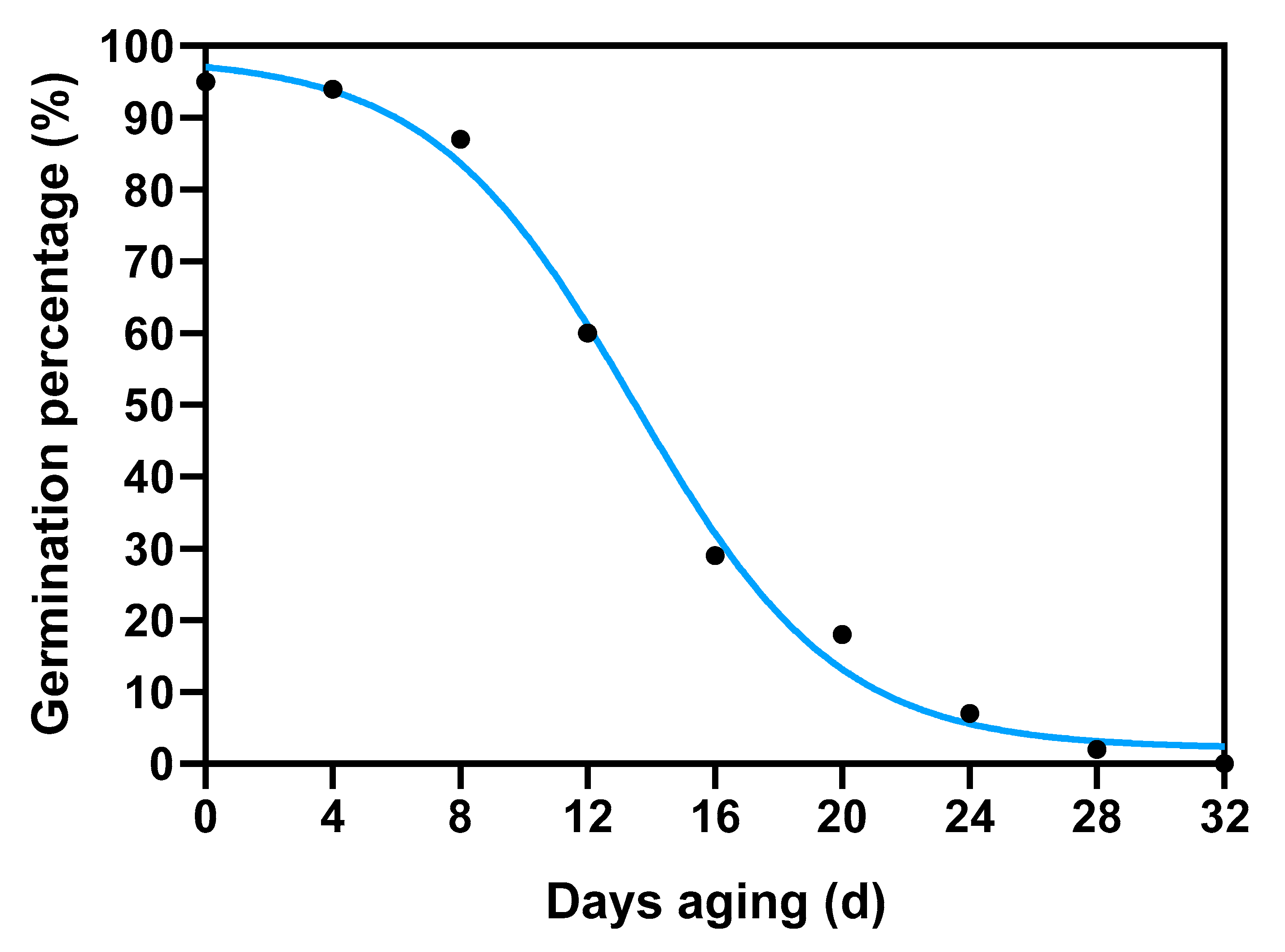
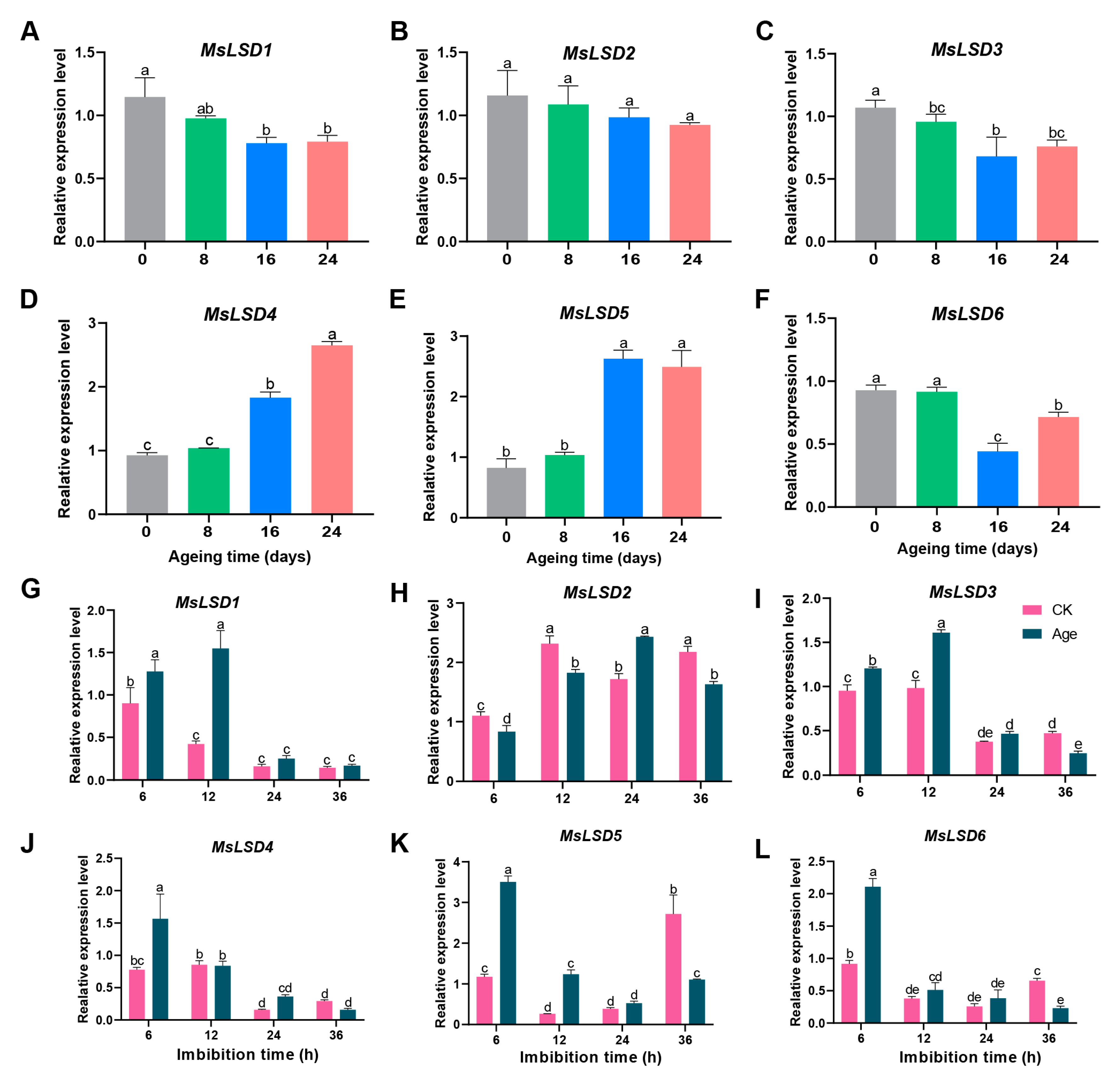
2.6. Plant Materials and Seed Aging Treatments
2.7. RT-qPCR Analysis
3. Results
3.1. Identification and Multiple Sequence Analysis of LSD Genes in Alfalfa
3.2. Phylogenetic Analysis, Gene Structures, and Motif Composition of MsLSD Genes
3.3. cis-Regulatory Element Analysis of MsLSD Gene Promoters
3.4. Secondary Structure Analysis of MsLSD Proteins
3.5. Expression Pattern Analysis of MsLSD Genes in Alfalfa Tissues
3.6. RNA-Seq Data Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bishaw, Z.; Niane, A.A.; Gan, Y. Quality seed production. In Lentil: An Ancient Crop for Modern Times; Springer: Dordrecht, The Netherlands, 2007; pp. 349–383. [Google Scholar]
- Ahmed, Z.; Shah, Z.H.; Rehman, H.M.; Shahzad, K.; Daur, I.; Elfeel, A.; Elsafori, A.K.; Yang, S.H.; Chung, G. Genomics: A hallmark to monitor molecular and biochemical processes leading toward a better perceptive of seed aging and ex-situ conservation. Curr. Issues Mol. Biol. 2017, 22, 89–112. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. Comptes Rendus Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Donà, M.; Balestrazzi, A.; Mondoni, A.; Rossi, G.; Ventura, L.; Buttafava, A.; Macovei, A.; Sabatini, M.E.; Valassi, A.; Carbonera, D. DNA profiling, telomere analysis and antioxidant properties as tools for monitoring ex situ seed longevity. Ann. Bot. 2013, 111, 987–998. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.E.; Yuan, J.; Horvitz, H.R. Mechanisms and functions of cell death. Annu. Rev. Cell Biol. 1991, 7, 663–698. [Google Scholar] [CrossRef] [PubMed]
- Locato, V.; Gadaleta, C.; De Gara, L.; De Pinto, M.C. Production of reactive species and modulation of antioxidant network in response to heat shock: A critical balance for cell fate. Plant Cell Environ. 2008, 31, 1606–1619. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Xing, D.; Li, L.; Zhang, L. Implication of reactive oxygen species and mitochondrial dysfunction in the early stages of plant programmed cell death induced by ultraviolet-C overexposure. Planta 2008, 227, 755–767. [Google Scholar] [CrossRef]
- Hu, D.I.E.; Ma, G.; Wang, Q.; Yao, J.; Wang, Y.U.; Pritchard, H.W.; Wang, X. Spatial and temporal nature of reactive oxygen species production and programmed cell death in elm (Ulmus pumila L.) seeds during controlled deterioration. Plant Cell Environ. 2012, 35, 2045–2059. [Google Scholar] [CrossRef]
- Epple, P.; Mack, A.A.; Morris, V.R.F.; Dangl, J.L. Antagonistic control of oxidative stress-induced cell death in Arabidopsis by two related, plant-specific zinc finger proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 6831–6836. [Google Scholar] [CrossRef]
- Czarnocka, W.; Van Der Kelen, K.; Willems, P.; Szechyńska-Hebda, M.; Shahnejat-Bushehri, S.; Balazadeh, S.; Rusaczonek, A.; Mueller-Roeber, B.; Van Breusegem, F.; Karpiński, S. The dual role of LESION SIMULATING DISEASE 1 as a condition-dependent scaffold protein and transcription regulator. Plant Cell Environ. 2017, 40, 2644–2662. [Google Scholar] [CrossRef]
- Dietrich, R.A.; Richberg, M.H.; Schmidt, R.; Dean, C.; Dangl, J.L. A novel zinc finger protein is encoded by the Arabidopsis LSD1 gene and functions as a negative regulator of plant cell death. Cell 1997, 88, 685–694. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Rusaczonek, A.; Czarnocka, W.; Karpiński, S. Salicylic Acid Accumulation Controlled by LSD1 Is Essential in Triggering Cell Death in Response to Abiotic Stress. Cells 2021, 10, 962. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pei, Z.; Tian, Y.; He, C. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation. Mol. Plant-Microbe Interact. 2005, 18, 375–384. [Google Scholar] [CrossRef]
- Ren, L.; Bennett, J.A.; Coulman, B.; Liu, J.; Biligetu, B. Forage yield trend of alfalfa cultivars in the Canadian prairies and its relation to environmental factors and harvest management. Grass Forage Sci. 2021, 76, 390–399. [Google Scholar] [CrossRef]
- Jia, Q.; Kamran, M.; Ali, S.; Sun, L.; Zhang, P.; Ren, X.; Jia, Z. Deficit irrigation and fertilization strategies to improve soil quality and alfalfa yield in arid and semi-arid areas of northern China. PeerJ 2018, 6, e4410. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Du, H.; Chen, Z.; Lu, H.; Zhu, F.; Chen, H.; Meng, X.; Liu, Q.; Liu, P.; Zheng, L. The chromosome-level genome sequence of the autotetraploid alfalfa and resequencing of core germplasms provide genomic resources for alfalfa research. Mol. Plant 2020, 13, 1250–1261. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.; Cheng, H.; Chen, L.; Zhu, H.; Mao, P.; Wang, M. Influence of exogenous ascorbic acid and glutathione priming on mitochondrial structural and functional systems to alleviate aging damage in oat seeds. BMC Plant Biol. 2020, 20, 104. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Han, B.; Yin, X.; Mao, P.; Luo, D.; Zhou, Q.; Liu, Z. Genome-wide identification of the GRAS transcription factor family in autotetraploid cultivated alfalfa (Medicago sativa L.) and expression analysis under drought stress. Ind. Crops Prod. 2023, 194, 116379. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Czarnocka, W.; Rusaczonek, A.; Witoń, D.; Kęska, S.; Czyż, J.; Szechyńska-Hebda, M.; Karpiński, S. LSD1-, EDS1-and PAD4-dependent conditional correlation among salicylic acid, hydrogen peroxide, water use efficiency and seed yield in Arabidopsis thaliana. Physiol. Plant. 2019, 165, 369–382. [Google Scholar] [CrossRef]
- Bernacki, M.J.; Mielecki, J.; Antczak, A.; Drożdżek, M.; Witoń, D.; Dąbrowska-Bronk, J.; Gawroński, P.; Burdiak, P.; Marchwicka, M.; Rusaczonek, A. Biotechnological Potential of the Stress Response and Plant Cell Death Regulators Proteins in the Biofuel Industry. Cells 2023, 12, 2018. [Google Scholar] [CrossRef]
- Kliebenstein, D.J.; Dietrich, R.A.; Martin, A.C.; Last, R.L.; Dangl, J.L. LSD1 regulates salicylic acid induction of copper zinc superoxide dismutase in Arabidopsis thaliana. Mol. Plant-Microbe Interact. 1999, 12, 1022–1026. [Google Scholar] [CrossRef]
- Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucleic Acids Res. 2020, 48, D1104–D1113. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2016, 45, D1040–D1045. [Google Scholar] [CrossRef]
- Jin, J.; He, K.; Tang, X.; Li, Z.; Lv, L.; Zhao, Y.; Luo, J.; Gao, G. An Arabidopsis transcriptional regulatory map reveals distinct functional and evolutionary features of novel transcription factors. Mol. Biol. Evol. 2015, 32, 1767–1773. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhang, H.; Kong, L.; Gao, G.; Luo, J. PlantTFDB 3.0: A portal for the functional and evolutionary study of plant transcription factors. Nucleic Acids Res. 2014, 42, D1182–D1187. [Google Scholar] [CrossRef] [PubMed]
- Mao, P.; Jin, X.; Bao, Q.; Mei, C.; Zhou, Q.; Min, X.; Liu, Z. WRKY transcription factors in Medicago sativa L.: Genome-wide identification and expression analysis under abiotic stress. DNA Cell Biol. 2020, 39, 2212–2225. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Huang, K.; Zhang, X.; Yu, X.; Huang, P.; An, C. The LSD1-type zinc finger motifs of Pisum sativa LSD1 are a novel nuclear localization signal and interact with importin alpha. PLoS ONE 2011, 6, e22131. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Beiko, R.G.; Langille, M.G.I. Predicting the functional potential of the microbiome from marker genes using PICRUSt. In Microbiome Analysis: Methods and Protocols; Humana Press: New York, NY, USA, 2018; pp. 169–177. [Google Scholar]
- Bernacki, M.J.; Czarnocka, W.; Szechyńska-Hebda, M.; Mittler, R.; Karpiński, S. Biotechnological Potential of LSD1, EDS1, and PAD4 in the Improvement of Crops and Industrial Plants. Plants 2019, 8, 290. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Id in ‘Zhongmu No. 1’ Assembly | Isoelectric Point | Molecular Weight (Da) | Amino Acids | Subcellular Localization | Instability Index | Average of Hydropathicity |
|---|---|---|---|---|---|---|---|
| MsLSD1 | MsG0280008660.01. T 01 | 9.13 | 7588.24 | 69 | Cytoplasm | 56.26 | 1.139 |
| MsLSD2 | MsG0280010569.01. T 01 | 9.08 | 21,433.39 | 192 | Nucleus | 47.59 | −0.543 |
| MsLSD3 | MsG0480020269.01. T 01 | 9.22 | 24,524.33 | 216 | Nucleus | 50.53 | −0.223 |
| MsLSD4 | MsG0480022392.01. T 01 | 8.37 | 9638.29 | 87 | Chloroplast | 75.9 | −0.333 |
| MsLSD5 | MsG0480022393.01. T 01 | 5.33 | 57,730.72 | 502 | Cytoplasm | 46.51 | −0.237 |
| MsLSD6 | MsG0880046041.01. T 01 | 6.77 | 15,775.28 | 147 | Chloroplast | 39.12 | 0.211 |
| Gene Name | Gene Id in ‘Zhongmu No. 1’ Assembly | α-Helix (%) | Extended Strand (%) | β-Turn (%) | Random Coil (%) |
|---|---|---|---|---|---|
| MsLSD1 | MsG0280008660.01. T 01 | 20.29% | 42.03% | 10.14% | 27.54% |
| MsLSD2 | MsG0280010569.01. T 01 | 22.92% | 17.19% | 5.21% | 54.69% |
| MsLSD3 | MsG0480020269.01. T 01 | 24.54% | 22.69% | 6.94% | 45.83% |
| MsLSD4 | MsG0480022392.01. T 01 | 21.84% | 21.84% | 8.05% | 48.28% |
| MsLSD5 | MsG0480022393.01. T 01 | 36.06% | 17.93% | 3.78% | 42.23% |
| MsLSD6 | MsG0880046041.01. T 01 | 17.69% | 26.53% | 5.44% | 50.34% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, S.; Ma, W.; Jia, Z.; Ou, C.; Li, M.; Mao, P. Genomic Identification and Expression Profiling of Lesion Simulating Disease Genes in Alfalfa (Medicago sativa) Elucidate Their Responsiveness to Seed Vigor. Antioxidants 2023, 12, 1768. https://doi.org/10.3390/antiox12091768
Sun S, Ma W, Jia Z, Ou C, Li M, Mao P. Genomic Identification and Expression Profiling of Lesion Simulating Disease Genes in Alfalfa (Medicago sativa) Elucidate Their Responsiveness to Seed Vigor. Antioxidants. 2023; 12(9):1768. https://doi.org/10.3390/antiox12091768
Chicago/Turabian StyleSun, Shoujiang, Wen Ma, Zhicheng Jia, Chengming Ou, Manli Li, and Peisheng Mao. 2023. "Genomic Identification and Expression Profiling of Lesion Simulating Disease Genes in Alfalfa (Medicago sativa) Elucidate Their Responsiveness to Seed Vigor" Antioxidants 12, no. 9: 1768. https://doi.org/10.3390/antiox12091768