
Oxidative and Endoplasmic Reticulum Stress Represent Novel Therapeutic Targets for Choroideremia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry
2.2. Drugs
2.3. Zebrafish Dosing
2.4. Fibroblast Cell Culture and Dosing
2.5. RT-qPCR
2.6. Western Blot
2.7. SOD Activity Assay
2.8. Retinal Histology
2.9. Melanin Quantification
2.10. Statistical Analysis
3. Results
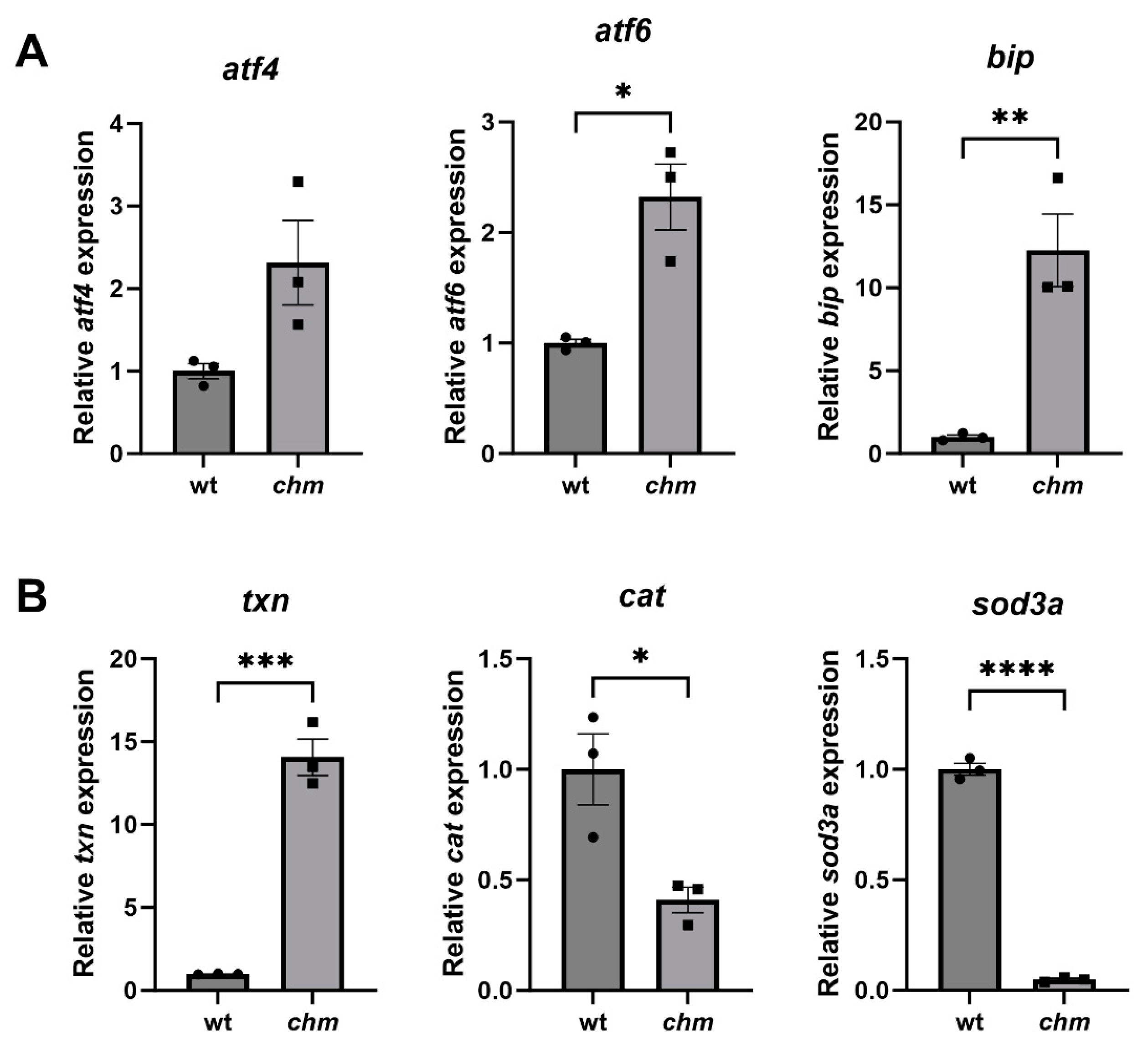
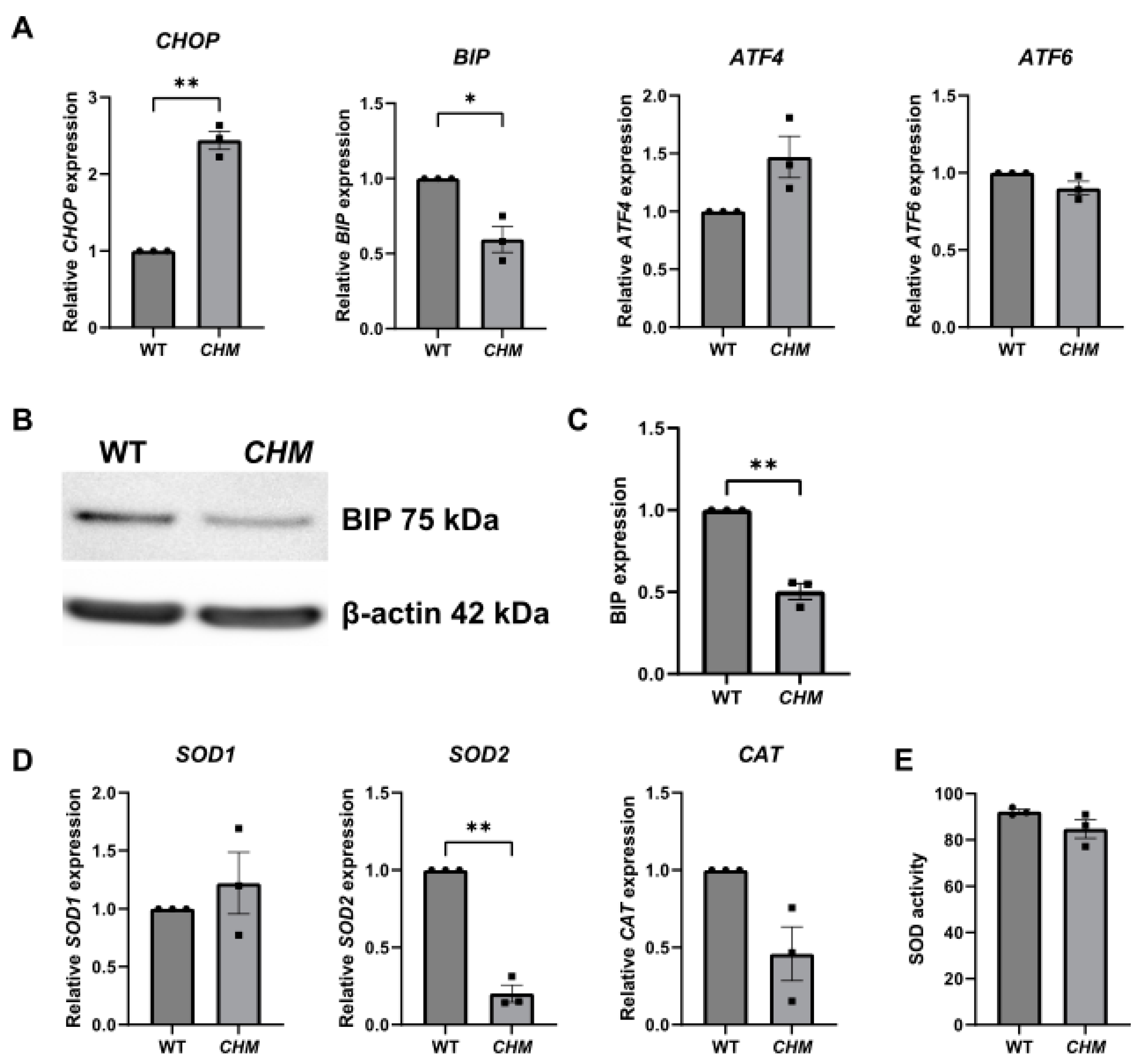
3.1. Increased ER and Oxidative Stress in chmru848 Zebrafish and CHMY42X Patient Fibroblasts
3.2. Drug Screening of Therapeutics Targeting ER and Oxidative Stress
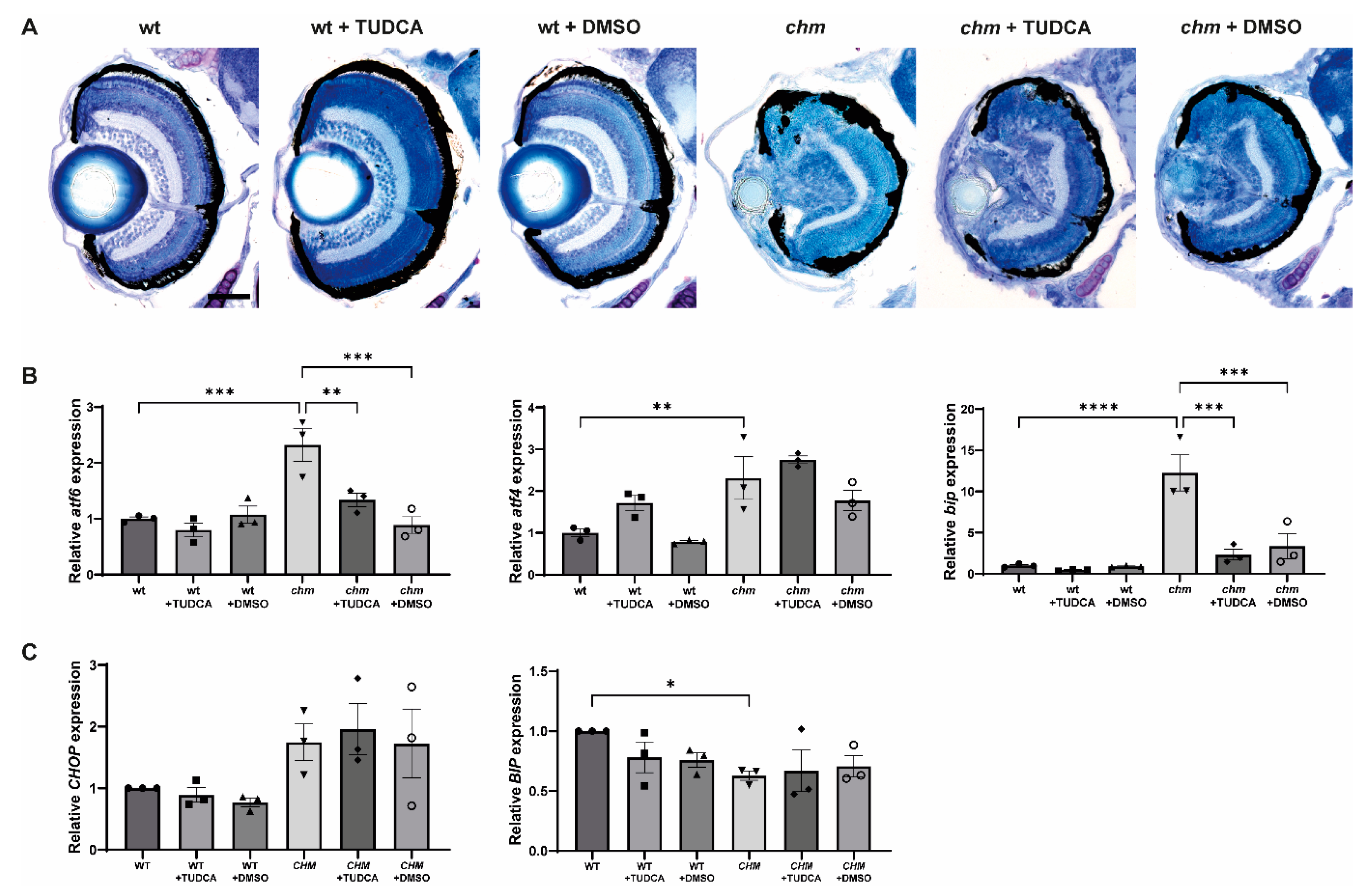
3.3. Tauroursodeoxycholic Acid (TUDCA)
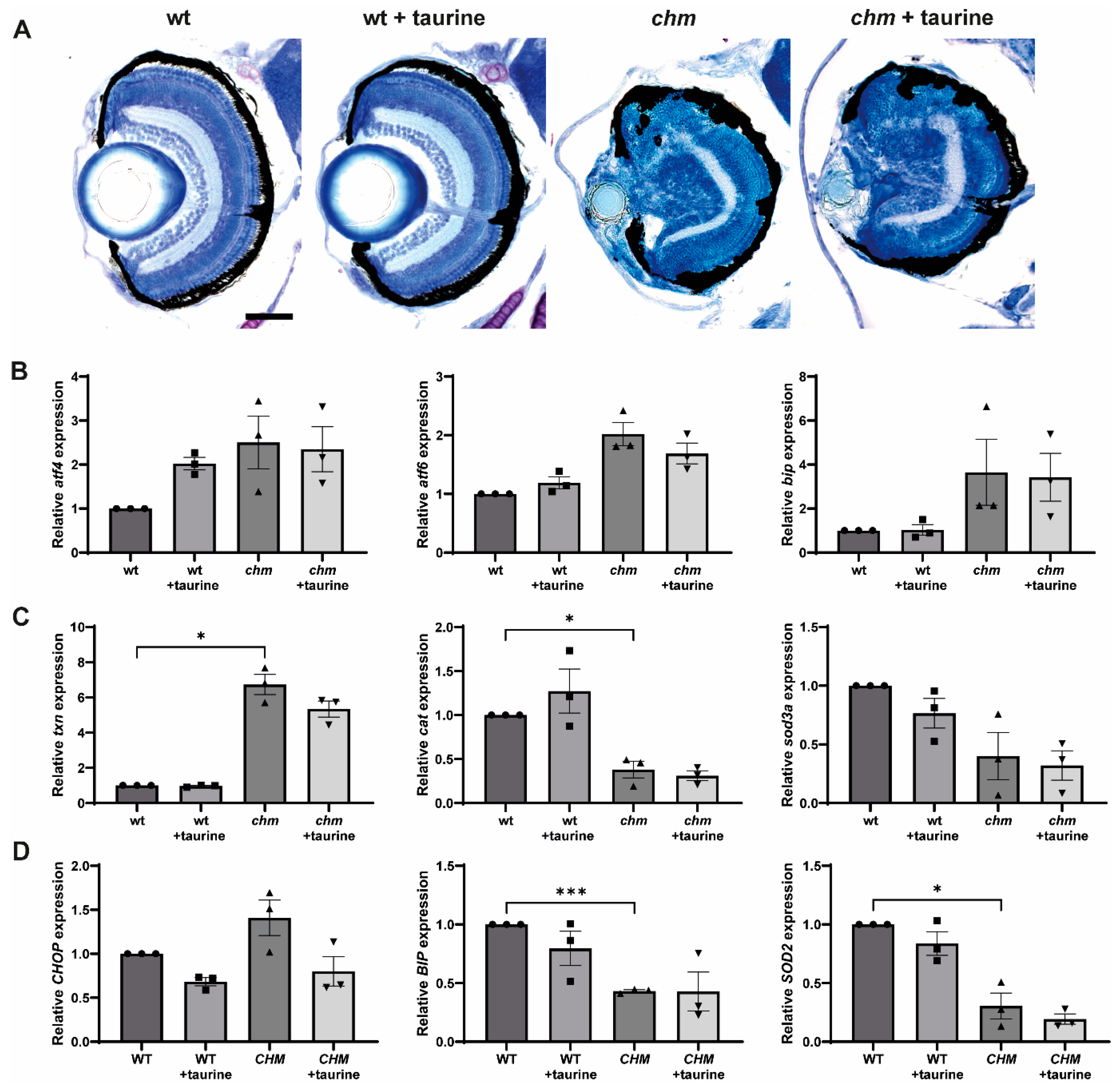
3.4. Taurine
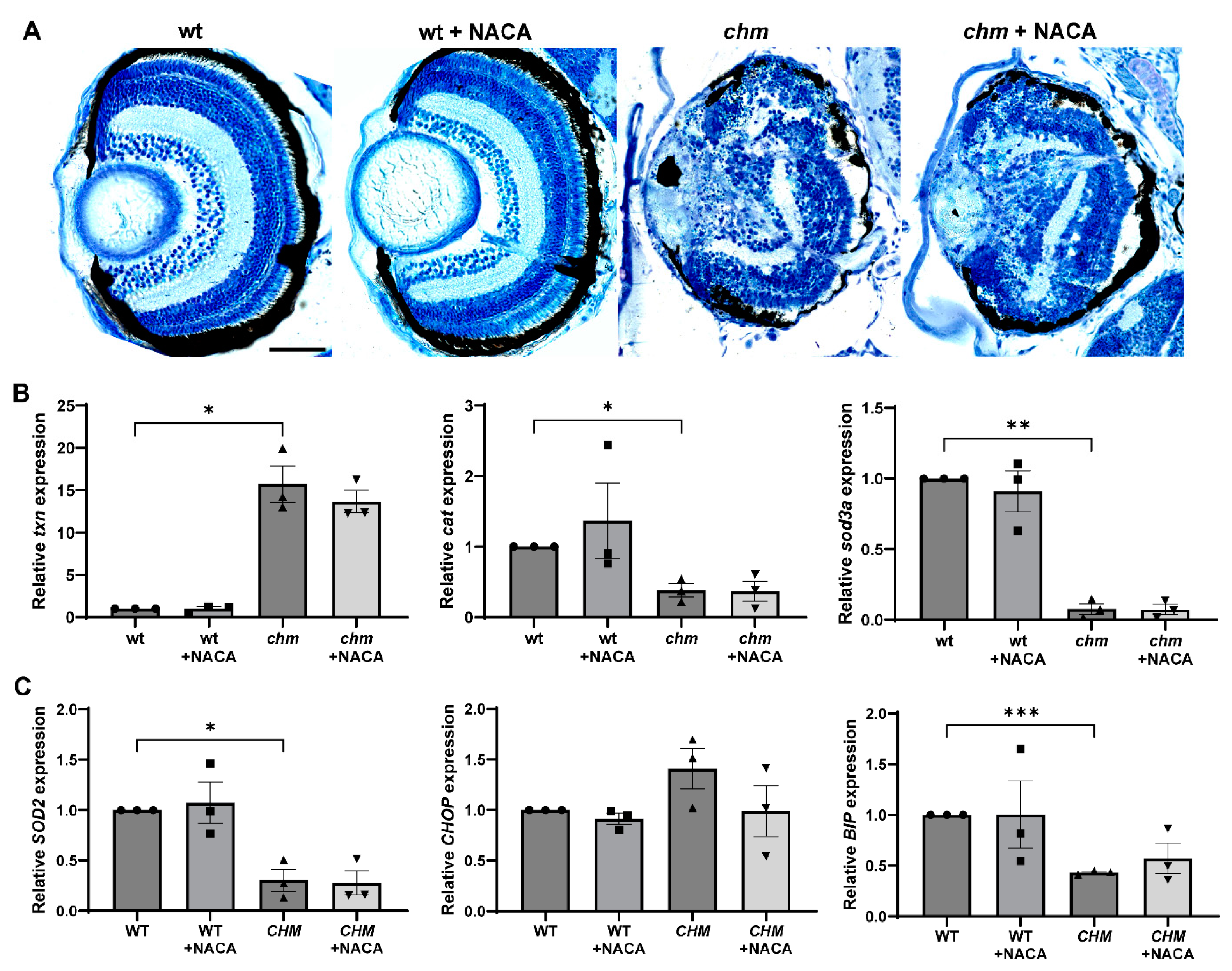
3.5. N-Acetylcysteine Amide (NACA)
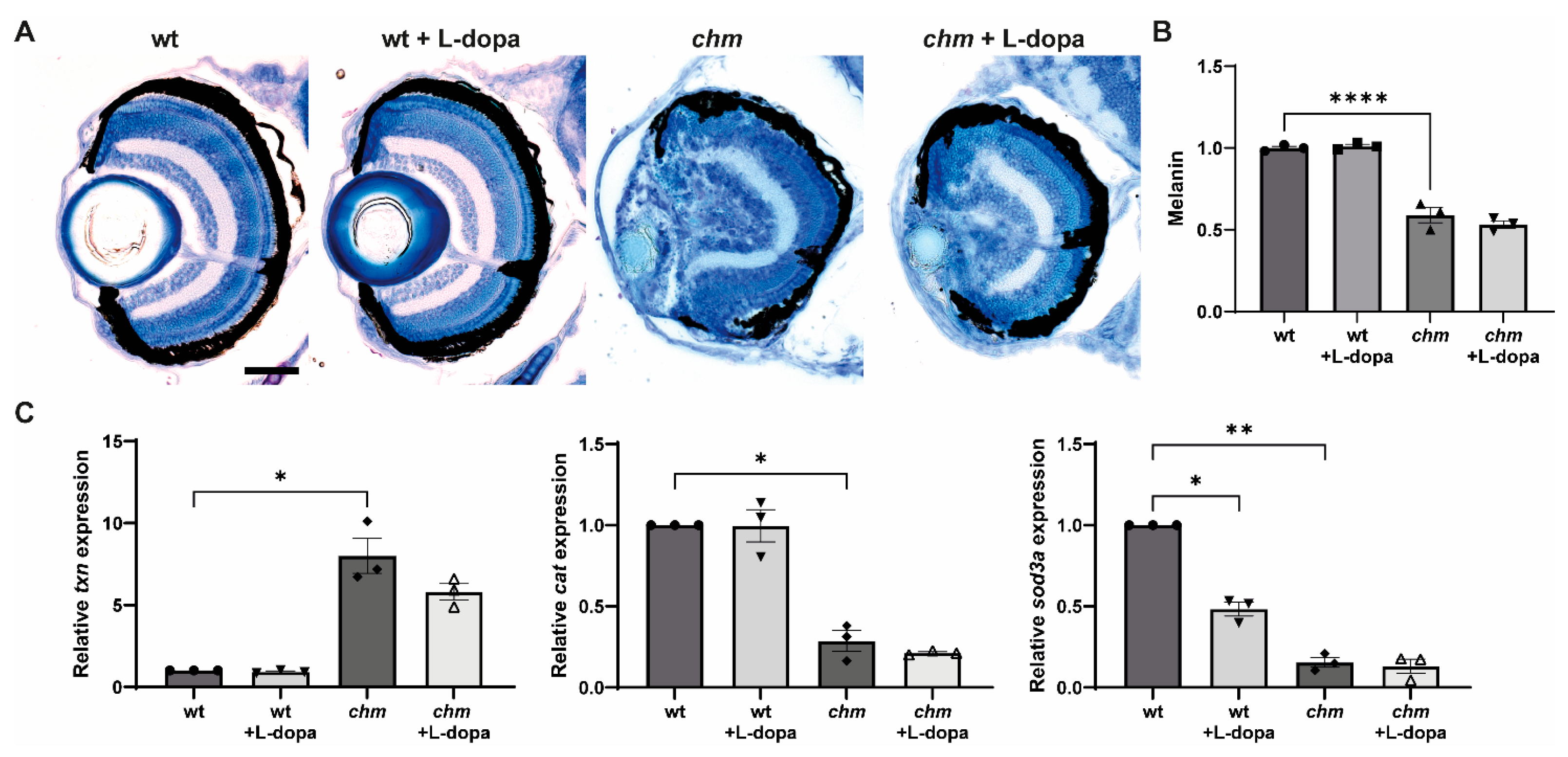
3.6. Levodopa (L-Dopa)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mitsios, A.; Dubis, A.M.; Moosajee, M. Choroideremia: From genetic and clinical phenotyping to gene therapy and future treatments. Ther. Adv. Ophthalmol. 2018, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, H.; Moosajee, M. Choroideremia: Molecular mechanisms and therapies. Trends Mol. Med. 2022, 28, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Gordiyenko, N.V.; Fariss, R.N.; Zhi, C.; MacDonald, I.M. Silencing of the CHM gene alters phagocytic and secretory pathways in the retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 2010, 51, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Strunnikova, N.V.; Barb, J.; Sergeev, Y.V.; Thiagarajasubramanian, A.; Silvin, C.; Munson, P.J.; MacDonald, I.M. Loss-of-Function Mutations in Rab Escort Protein 1 (REP-1) Affect Intracellular Transport in Fibroblasts and Monocytes of Choroideremia Patients. PLoS ONE 2009, 4, e8402. [Google Scholar] [CrossRef]
- Tolmachova, T.; Wavre-Shapton, S.T.; Barnard, A.R.; MacLaren, R.E.; Futter, C.E.; Seabra, M.C. Retinal Pigment Epithelium Defects Accelerate Photoreceptor Degeneration in Cell Type–Specific Knockout Mouse Models of Choroideremia. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4913–4920. [Google Scholar] [CrossRef]
- Wavre-Shapton, S.T.; Tolmachova, T.; da Silva, M.L.; Futter, C.E.; Seabra, M.C. Conditional Ablation of the Choroideremia Gene Causes Age-Related Changes in Mouse Retinal Pigment Epithelium. PLoS ONE 2013, 8, e57769. [Google Scholar] [CrossRef]
- Cunha, D.L.; Richardson, R.; Tracey-White, D.; Abbouda, A.; Mitsios, A.; der Sluis, V.H.-V.; Takis, P.; Owen, N.; Skinner, J.; Welch, A.A.; et al. REP1 deficiency causes systemic dysfunction of lipid metabolism and oxidative stress in choroideremia. JCI Insight 2021, 6, e146934. [Google Scholar] [CrossRef]
- MacLaren, R.E.; Groppe, M.; Barnard, A.R.; Cottriall, C.L.; Tolmachova, T.; Seymour, L.; Clark, K.R.; During, M.J.; Cremers, F.P.; Black, G.C.; et al. Retinal gene therapy in patients with choroideremia: Initial findings from a phase 1/2 clinical trial. Lancet 2014, 383, 1129–1137. [Google Scholar] [CrossRef]
- Xue, K.; Jolly, J.K.; Barnard, A.R.; Rudenko, A.; Salvetti, A.P.; Patrício, M.I.; Edwards, T.L.; Groppe, M.; Orlans, H.O.; Tolmachova, T.; et al. Beneficial effects on vision in patients undergoing retinal gene therapy for choroideremia. Nat. Med. 2018, 24, 1507–1512. [Google Scholar] [CrossRef]
- Moosajee, M.; Tulloch, M.; Baron, R.A.; Gregory-Evans, C.Y.; Pereira-Leal, J.B.; Seabra, M.C. Single choroideremia gene in nonmammalian vertebrates explains early embryonic lethality of the zebrafish model of choroideremia. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3009–3016. [Google Scholar] [CrossRef]
- Moosajee, M.; Tracey-White, D.; Smart, M.; Weetall, M.; Torriano, S.; Kalatzis, V.; da Cruz, L.; Coffey, P.; Webster, A.R.; Welch, E. Functional rescue of REP1 following treatment with PTC124 and novel derivative PTC-414 in human choroideremia fibroblasts and the nonsense-mediated zebrafish model. Hum. Mol. Genet. 2016, 25, 3416–3431. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, H.; Toms, M.; Moosajee, M. Involvement of Oxidative and Endoplasmic Reticulum Stress in RDH12-Related Retinopathies. Int. J. Mol. Sci. 2021, 22, 8863. [Google Scholar] [CrossRef] [PubMed]
- Agalou, A.; Thrapsianiotis, M.; Angelis, A.; Papakyriakou, A.; Skaltsounis, A.-L.; Aligiannis, N.; Beis, D. Identification of Novel Melanin Synthesis Inhibitors from Crataegus pycnoloba Using an in Vivo Zebrafish Phenotypic Assay. Front. Pharmacol. 2018, 9, 265. [Google Scholar] [CrossRef] [PubMed]
- Drack, A.V.; Dumitrescu, A.V.; Bhattarai, S.; Gratie, D.; Stone, E.M.; Mullins, R.; Sheffield, V.C. TUDCA slows retinal degeneration in two different mouse models of retinitis pigmentosa and prevents obesity in Bardet-Biedl syndrome type 1 mice. Investig. Ophthalmol. Vis. Sci. 2012, 53, 100–106. [Google Scholar] [CrossRef]
- Fernández-Sánchez, L.; Lax, P.; Pinilla, I.; Martín-Nieto, J.; Cuenca, N. Tauroursodeoxycholic acid prevents retinal degeneration in transgenic P23H rats. Investig. Ophthalmol. Vis. Sci. 2011, 52, 4998–5008. [Google Scholar] [CrossRef]
- Phillips, M.J.; Walker, T.A.; Choi, H.Y.; Faulkner, A.E.; Kim, M.K.; Sidney, S.S.; Boyd, A.P.; Nickerson, J.M.; Boatright, J.H.; Pardue, M.T. Tauroursodeoxycholic acid preservation of photoreceptor structure and function in the rd10 mouse through postnatal day 30. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2148–2155. [Google Scholar] [CrossRef]
- Jakaria, M.; Azam, S.; Haque, M.E.; Jo, S.-H.; Uddin, M.S.; Kim, I.-S.; Choi, D.-K. Taurine and its analogs in neurological disorders: Focus on therapeutic potential and molecular mechanisms. Redox Biol. 2019, 24, 101223. [Google Scholar] [CrossRef]
- Tao, Y.; He, M.; Yang, Q.; Ma, Z.; Qu, Y.; Chen, W.; Peng, G.; Teng, D. Systemic taurine treatment provides neuroprotection against retinal photoreceptor degeneration and visual function impairments. Drug Des. Dev. Ther. 2019, 13, 2689–2702. [Google Scholar] [CrossRef]
- Zhang, Y.; Ren, S.; Liu, Y.; Gao, K.; Liu, Z.; Zhang, Z. Inhibition of Starvation-Triggered Endoplasmic Reticulum Stress, Autophagy, and Apoptosis in ARPE-19 Cells by Taurine through Modulating the Expression of Calpain-1 and Calpain-2. Int. J. Mol. Sci. 2017, 18, 2146. [Google Scholar] [CrossRef]
- Lee, S.Y.; Usui, S.; Zafar, A.-B.; Oveson, B.C.; Jo, Y.-J.; Lu, L.; Masoudi, S.; Campochiaro, P.A. N-acetylcysteine promotes long-term survival of cones in a model of retinitis pigmentosa. J. Cell. Physiol. 2010, 226, 1843–1849. [Google Scholar] [CrossRef]
- Campochiaro, P.A.; Iftikhar, M.; Hafiz, G.; Akhlaq, A.; Tsai, G.; Wehling, D.; Lu, L.; Wall, G.M.; Singh, M.S.; Kong, X. Oral N-acetylcysteine improves cone function in retinitis pigmentosa patients in phase I trial. J. Clin. Investig. 2020, 130, 1527–1541. [Google Scholar] [CrossRef] [PubMed]
- Sunitha, K.; Hemshekhar, M.; Thushara, R.M.; Santhosh, M.S.; Yariswamy, M.; Kemparaju, K.; Girish, K.S. N-Acetylcysteine amide: A derivative to fulfill the promises of N-Acetylcysteine. Free Radic. Res. 2013, 47, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Scott, J.; Griffiths, H.; Self, J.E.; Lotery, A. Oral levodopa rescues retinal morphology and visual function in a murine model of human albinism. Pigment Cell Melanoma Res. 2019, 32, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Lopez, V.M.; Decatur, C.L.; Stamer, W.D.; Lynch, R.M.; McKay, B.S. L-DOPA is an endogenous ligand for OA1. PLoS Biol. 2008, 6, e236. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Duong, T.T.; Vasireddy, V.; Ramachandran, P.; Herrera, P.S.; Leo, L.; Merkel, C.; Bennett, J.; Mills, J.A. Use of induced pluripotent stem cell models to probe the pathogenesis of Choroideremia and to develop a potential treatment. Stem Cell Res. 2018, 27, 140–150. [Google Scholar] [CrossRef]
- Krock, B.L.; Bilotta, J.; Perkins, B.D. Noncell-autonomous photoreceptor degeneration in a zebrafish model of choroideremia. Proc. Natl. Acad. Sci. USA 2007, 104, 4600–4605. [Google Scholar] [CrossRef]
- Seabra, M.C.; Ho, Y.K.; Anant, J.S. Deficient Geranylgeranylation of Ram/Rab27 in Choroideremia (∗). J. Biol. Chem. 1995, 270, 24420–24427. [Google Scholar] [CrossRef]
- Futter, C.E.; Ramalho, J.S.; Jaissle, G.B.; Seeliger, M.W.; Seabra, M.C. The role of Rab27a in the regulation of melanosome distribution within retinal pigment epithelial cells. Mol. Biol. Cell 2004, 15, 2264–2275. [Google Scholar] [CrossRef]
- Lopes, V.S.; Ramalho, J.S.; Owen, D.M.; Karl, M.O.; Strauss, O.; Futter, C.E.; Seabra, M.C. The ternary Rab27a-Myrip-Myosin VIIa complex regulates melanosome motility in the retinal pigment epithelium. Traffic 2007, 8, 486–499. [Google Scholar] [CrossRef]
- Sarkar, H.; Tracey-White, D.; Hagag, A.M.; Burgoyne, T.; Jensen, L.D.; Edwards, M.; Moosajee, M. Loss of REP1 impacts choroidal melanogenesis in choroideremia. bioRxiv 2023. [Google Scholar] [CrossRef]
- Ibrahim, I.M.; Abdelmalek, D.H.; Elfiky, A.A. GRP78: A cell’s response to stress. Life Sci. 2019, 226, 156–163. [Google Scholar] [PubMed]
- Casas, C. GRP78 at the Centre of the Stage in Cancer and Neuroprotection. Front. Neurosci. 2017, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Long, P.; He, M.; Yan, W.; Chen, W.; Wei, D.; Wang, S.; Zhang, Z.; Ge, W.; Chen, T. ALDH2 protects naturally aged mouse retina via inhibiting oxidative stress-related apoptosis and enhancing unfolded protein response in endoplasmic reticulum. Aging 2020, 13, 2750–2767. [Google Scholar] [CrossRef]
- Sandoval, C.O.; Simmen, T. Rab proteins of the endoplasmic reticulum: Functions and interactors. Biochem. Soc. Trans. 2012, 40, 1426–1432. [Google Scholar] [CrossRef]
- Zhang, T.; Baehr, W.; Fu, Y. Chemical chaperone TUDCA preserves cone photoreceptors in a mouse model of Leber congenital amaurosis. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3349–3356. [Google Scholar] [CrossRef]
- Simões-Correia, J.; Figueiredo, J.; Oliveira, C.; van Hengel, J.; Seruca, R.; van Roy, F.; Suriano, G. Endoplasmic reticulum quality control: A new mechanism of E-cadherin regulation and its implication in cancer. Hum. Mol. Genet. 2008, 17, 3566–3576. [Google Scholar] [CrossRef]
- Lee, S.-A.; Belyaeva, O.V.; Kedishvili, N.Y. Evidence that proteosome inhibitors and chemical chaperones can rescue the activity of retinol dehydrogenase 12 mutant T49M. Chem. Interact. 2011, 191, 55–59. [Google Scholar] [CrossRef]
- Martínez-Vacas, A.; Di Pierdomenico, J.; Gallego-Ortega, A.; Valiente-Soriano, F.J.; Vidal-Sanz, M.; Picaud, S.; Villegas-Pérez, M.P.; García-Ayuso, D. Systemic taurine treatment affords functional and morphological neuroprotection of photoreceptors and restores retinal pigment epithelium function in RCS rats. Redox Biol. 2022, 57, 102506. [Google Scholar] [CrossRef]
- Schimel, A.M.; Abraham, L.; Cox, D.; Sene, A.; Kraus, C.; Dace, D.S.; Ercal, N.; Apte, R.S. N-Acetylcysteine Amide (NACA) Prevents Retinal Degeneration by Up-Regulating Reduced Glutathione Production and Reversing Lipid Peroxidation. Am. J. Pathol. 2011, 178, 2032–2043. [Google Scholar] [CrossRef]
- Summers, C.G.; E Connett, J.; Holleschau, A.M.; Anderson, J.L.; De Becker, I.; McKay, B.S.; Brilliant, M.H. Does levodopa improve vision in albinism? Results of a randomized, controlled clinical trial. Clin. Exp. Ophthalmol. 2014, 42, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Falk, T.; Congrove, N.R.; Zhang, S.; McCourt, A.D.; Sherman, S.J.; McKay, B.S. PEDF and VEGF-A output from human retinal pigment epithelial cells grown on novel microcarriers. J. Biomed. Biotechnol. 2012, 2012, 278932. [Google Scholar] [CrossRef] [PubMed]
- Brilliant, M.H.; Vaziri, K.; Connor, T.B.; Schwartz, S.G.; Carroll, J.J.; McCarty, C.A.; Schrodi, S.J.; Hebbring, S.J.; Kishor, K.S.; Flynn, H.W.; et al. Mining Retrospective Data for Virtual Prospective Drug Repurposing: L-DOPA and Age-related Macular Degeneration. Am. J. Med. 2015, 129, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, A.G.; Boyd, B.M.; Christensen, C.A.; Javid, C.G.; McKay, B.S.; Fagan, T.C.; Snyder, R.W. Levodopa Positively Affects Neovascular Age-Related Macular Degeneration. Am. J. Med. 2021, 134, 122–128.e3. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, C.; Fu, J.; Lilley, B.; Berlinicke, C.; Hansen, B.; Ding, D.; Wang, G.; Wang, T.; Shou, D.; et al. Large-scale phenotypic drug screen identifies neuroprotectants in zebrafish and mouse models of retinitis pigmentosa. eLife 2021, 10, e57245. [Google Scholar] [PubMed]
- Ganzen, L.; Venkatraman, P.; Pang, C.P.; Leung, Y.F.; Zhang, M. Utilizing Zebrafish Visual Behaviors in Drug Screening for Retinal Degeneration. Int. J. Mol. Sci. 2017, 18, 1185. [Google Scholar] [CrossRef] [PubMed]
- Tolmachova, T.; Anders, R.; Abrink, M.; Bugeon, L.; Dallman, M.J.; Futter, C.E.; Ramalho, J.S.; Tonagel, F.; Tanimoto, N.; Seeliger, M.W.; et al. Independent degeneration of photoreceptors and retinal pigment epithelium in conditional knockout mouse models of choroideremia. J. Clin. Investig. 2006, 116, 386–394. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| Human | ||
| ATF4 | TCAAACCTCATGGGTTCTCC | GTGTCATCCAACGTGGTCAG |
| ATF6 | ACCCGTATTCTTCAGGGTGC | TCACTCCCTGAGTTCCTGCT |
| CHOP | GACCTGCAAGAGGTCCTGTC | TGTGACCTCTGCTGGTTCTG |
| BIP | GCCTGTATTTCTAGACCTGCC | TTCATCTTGCCAGCCAGTTG |
| SOD1 | TAGCGAGTTATGGCGACGAAG | TGGTCCATTACTTTCCTTCTGCT |
| SOD2 | GCTGGAAGCCATCAAACGTG | GCAGTGGAATAAGGCCTGTTG |
| CAT | CTCCGGAACAACAGCCTTCT | GAATGCCCGCACCTGAGTAA |
| GAPDH | ACAGTTGCCATGTAGACC | TTTTTGGTTGAGCACAGG |
| Zebrafish | ||
| atf4 | TGAGCACACTGAGGTTCCAG | GTCTTCACTCGGCCTTTGAG |
| atf6 | TGATGAGGCACTGTCTCCAG | ATGGGTCTTTTTGCTGGTTG |
| bip | CAAGAAGAAGACGGGCAAAG | CTCCTCAAACTTGGCTCTGG |
| txn | GACCATCGGGCCGTACTTTA | CATAAAGCGGCCACATCCTGT |
| cat | ACGATGACAACGTGACCCAA | CCATCAGGTTTTGCACCATGC |
| sod3a | TCAAGTGCGTGCCATCCATA | CCGCCGGATAAGTCCTTGTT |
| actin | CGAGCTGTCTTCCCATCCA | TCACCAACGTAGCTGTCTTTCTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarkar, H.; Lahne, M.; Nair, N.; Moosajee, M. Oxidative and Endoplasmic Reticulum Stress Represent Novel Therapeutic Targets for Choroideremia. Antioxidants 2023, 12, 1694. https://doi.org/10.3390/antiox12091694
Sarkar H, Lahne M, Nair N, Moosajee M. Oxidative and Endoplasmic Reticulum Stress Represent Novel Therapeutic Targets for Choroideremia. Antioxidants. 2023; 12(9):1694. https://doi.org/10.3390/antiox12091694
Chicago/Turabian StyleSarkar, Hajrah, Manuela Lahne, Neelima Nair, and Mariya Moosajee. 2023. "Oxidative and Endoplasmic Reticulum Stress Represent Novel Therapeutic Targets for Choroideremia" Antioxidants 12, no. 9: 1694. https://doi.org/10.3390/antiox12091694