
Phytochemical Constituents and Biological Activity of Wild and Cultivated Rosmarinus officinalis Hydroalcoholic Extracts
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Plant Material
2.3. Extract Preparation of R. officinalis L.
2.4. Chemical Analysis: LC-HRESIMS/MS Analysis
2.5. Chromatographic Purification of Compound 4
 |
2.5.1. Quantitative Analysis of Major Phenolic Compounds
2.5.2. Analysis of Total Phenol and Flavonoid Content
2.6. Antioxidant Activity
2.6.1. DPPH Assay
2.6.2. FRAP Assay
2.7. Anti-Enzymatic Activities
Anti-Acetylcholinesterase (AChE), Anti-Butyrylcholinesterase (BChE), and Anti-α-Amylase Activities
2.8. Cell Culture
2.9. MTT Assay
2.10. Quantitative Real-Time PCR
2.11. Intracellular ROS and Calcium Concentration Measurement
2.12. Quantification of Nitrite in Cell Culture Supernatants
2.13. Flow Cytometry
2.14. Western Blot Analysis
2.15. Prostaglandin E2 Assay
2.16. NF-kB Activity Assay
2.17. Statistical Analysis
3. Results
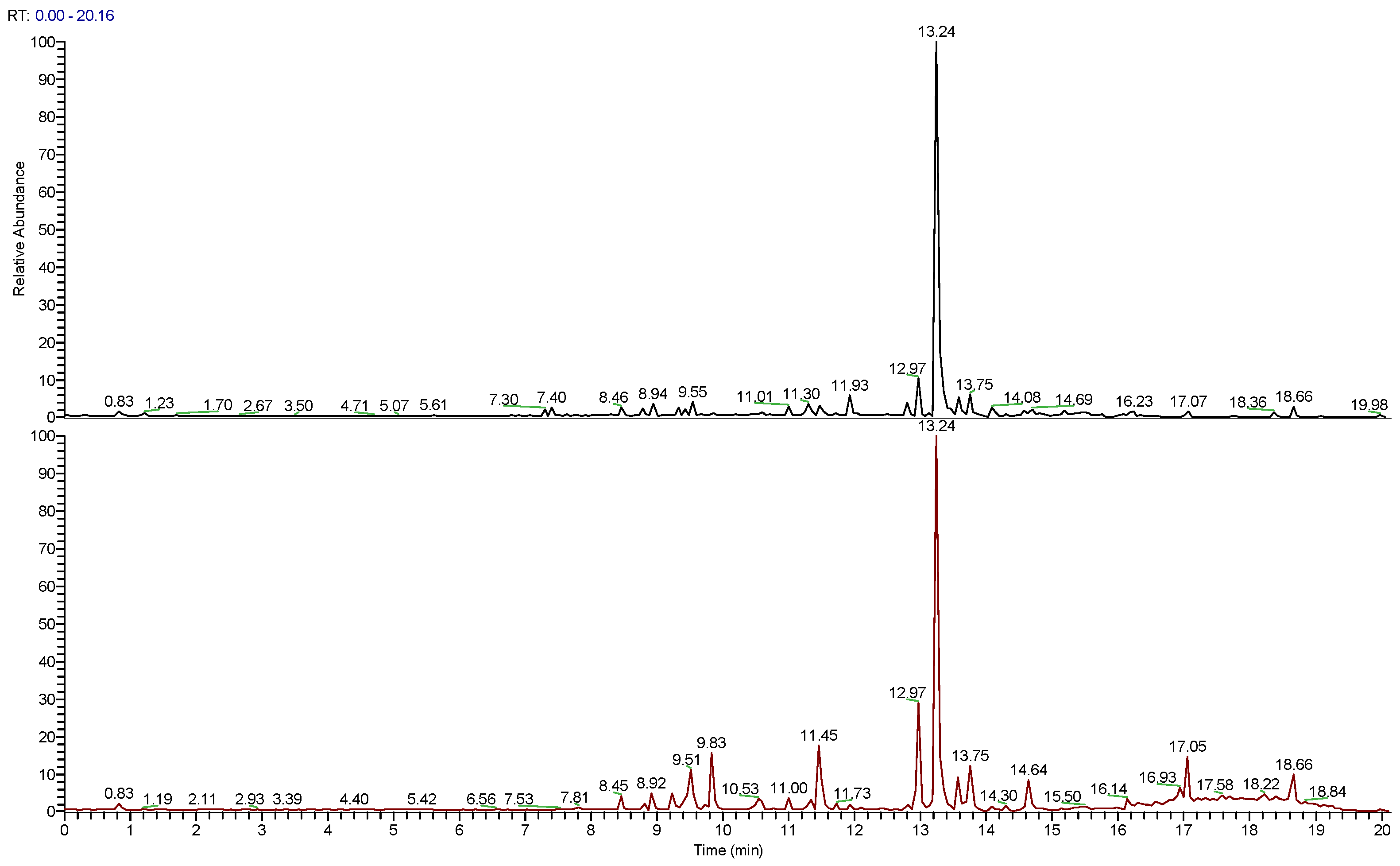
3.1. Chemical Analisys of Cultivated and Wild Extracts
3.1.1. Chemical Composition
3.1.2. Quantitative Analysis of Major Phenolic Compounds
3.1.3. Total Phenolic and Flavonoid Content
3.2. Antioxidant Activity: DPPH and FRAP Assay
3.3. Enzymatic Activities
Anti-Acetylcolinesterase (AChE), Anti-Butyrylcolinesterase (BChE), and Anti-α-Amylase Activities
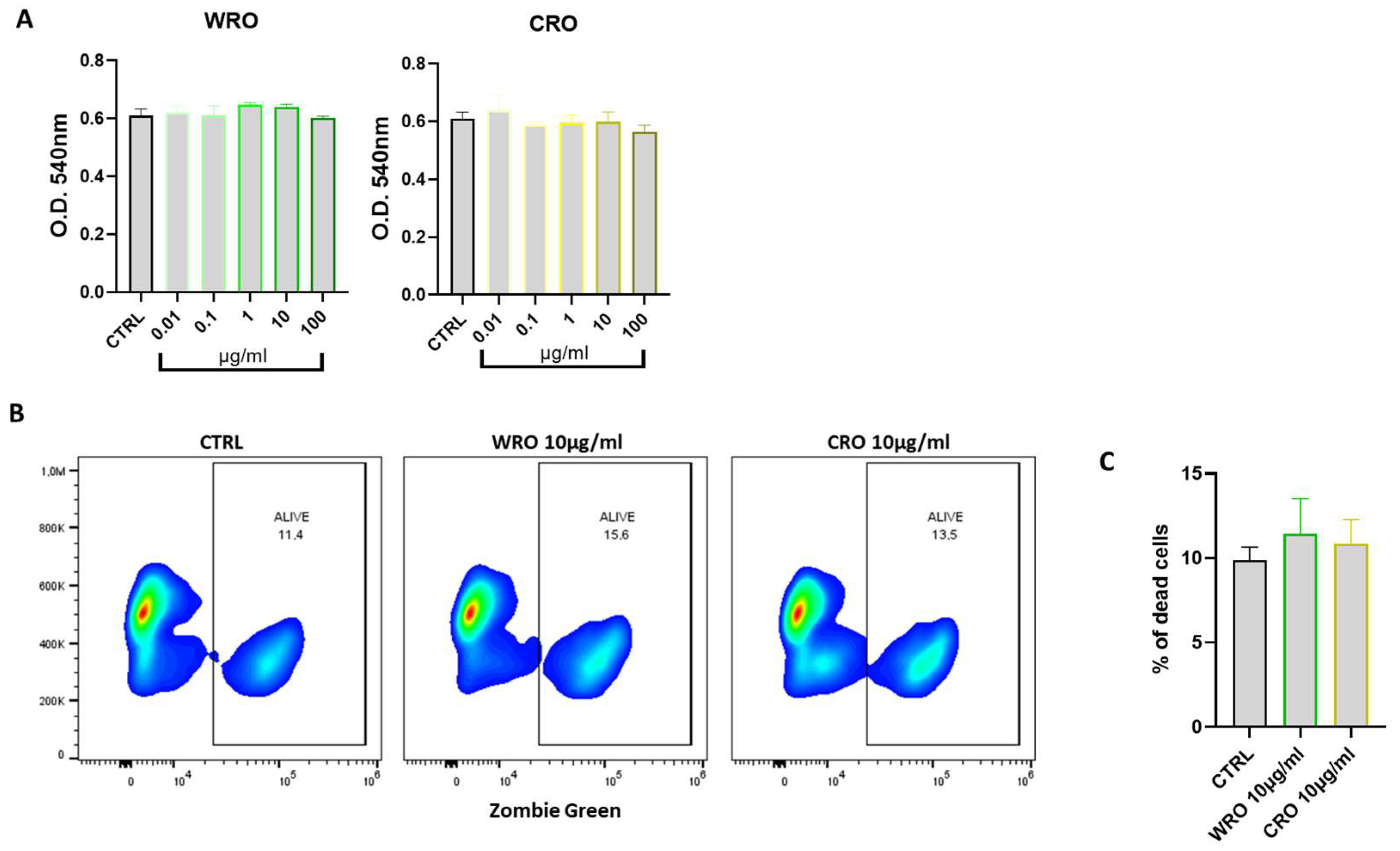
3.4. Effect of Rosmarinus Officinalis Extracts on Macrophage Cell Viability
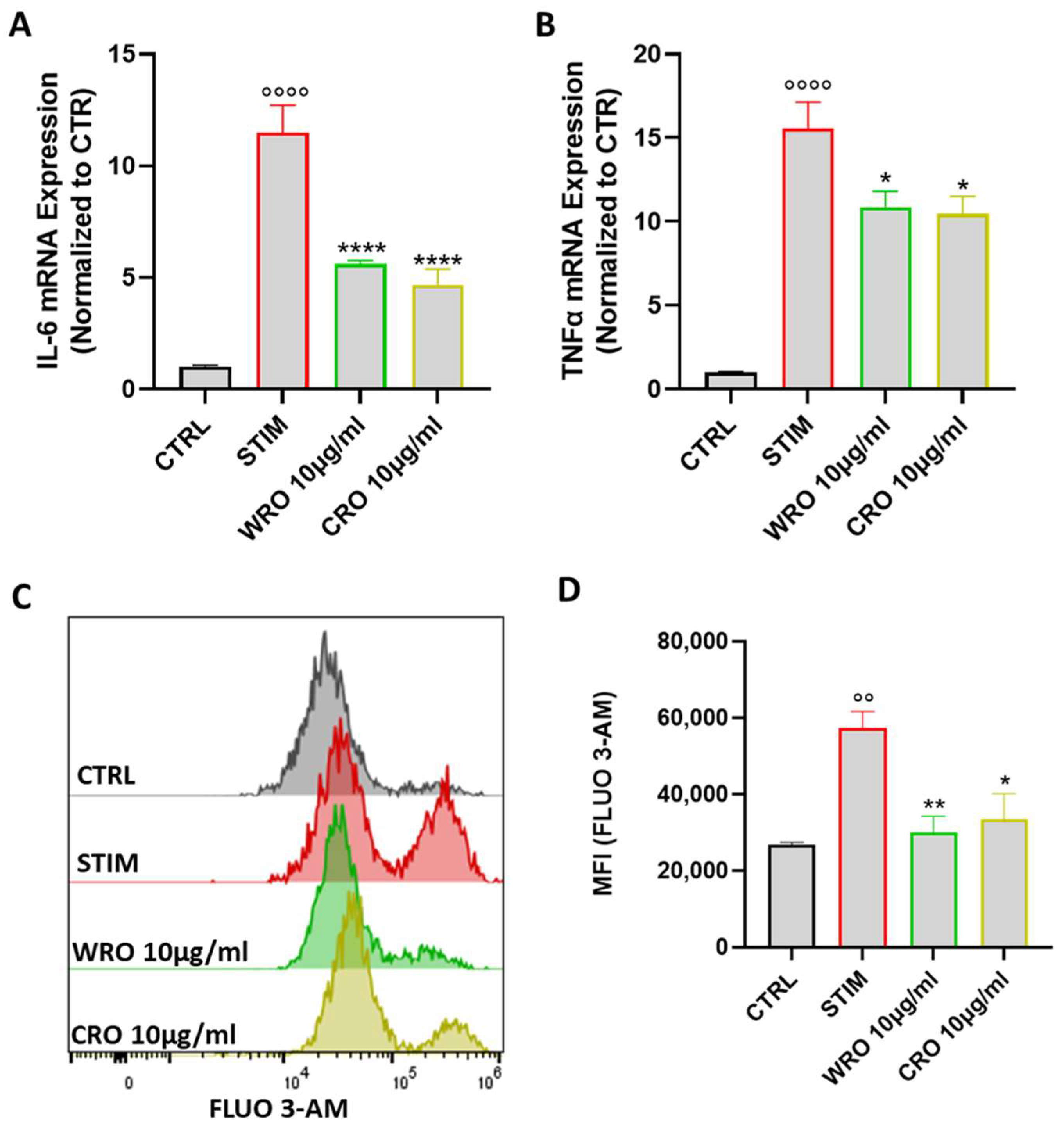
3.5. WRO and CRO Extracts Inhibited the Production of TNF- and IL-6 by Stimulated Macrophages
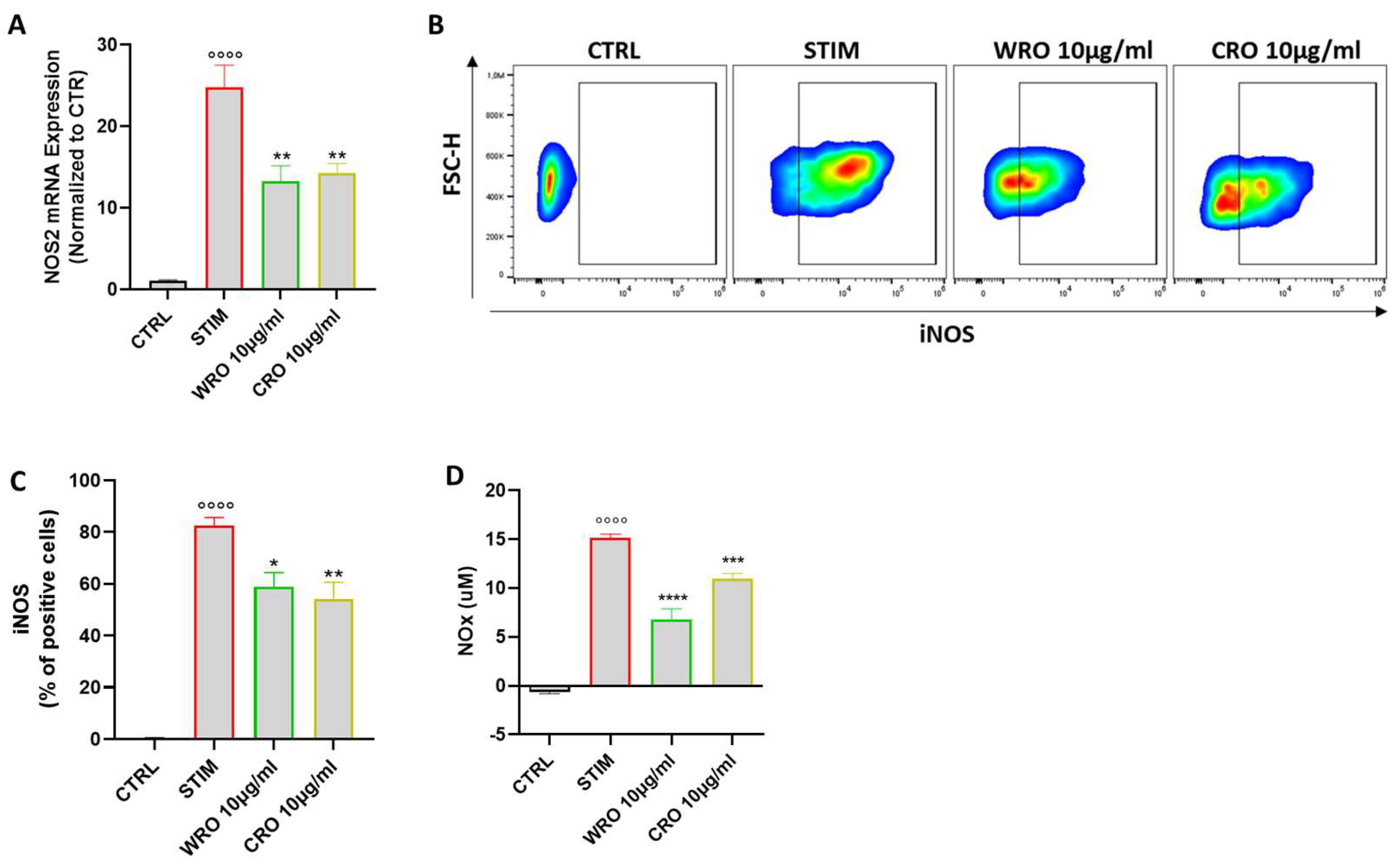
3.6. WRO and CRO Extracts Modulated the Nitric Oxide (NO) Pathway in Activated Macrophages
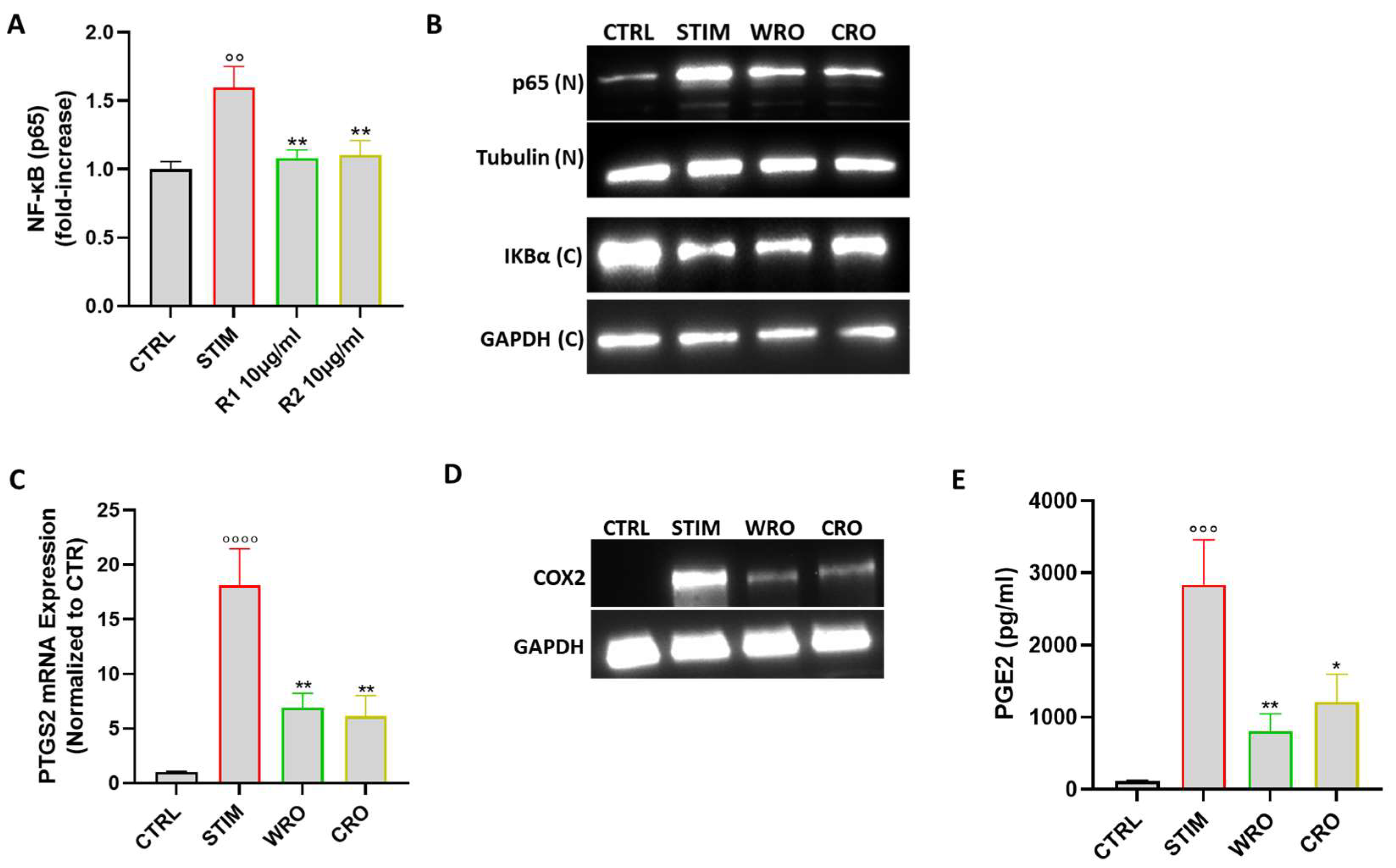
3.7. WRO and CRO Exert Antiinflammatory Effect by Inhibiting NF-κB Activation
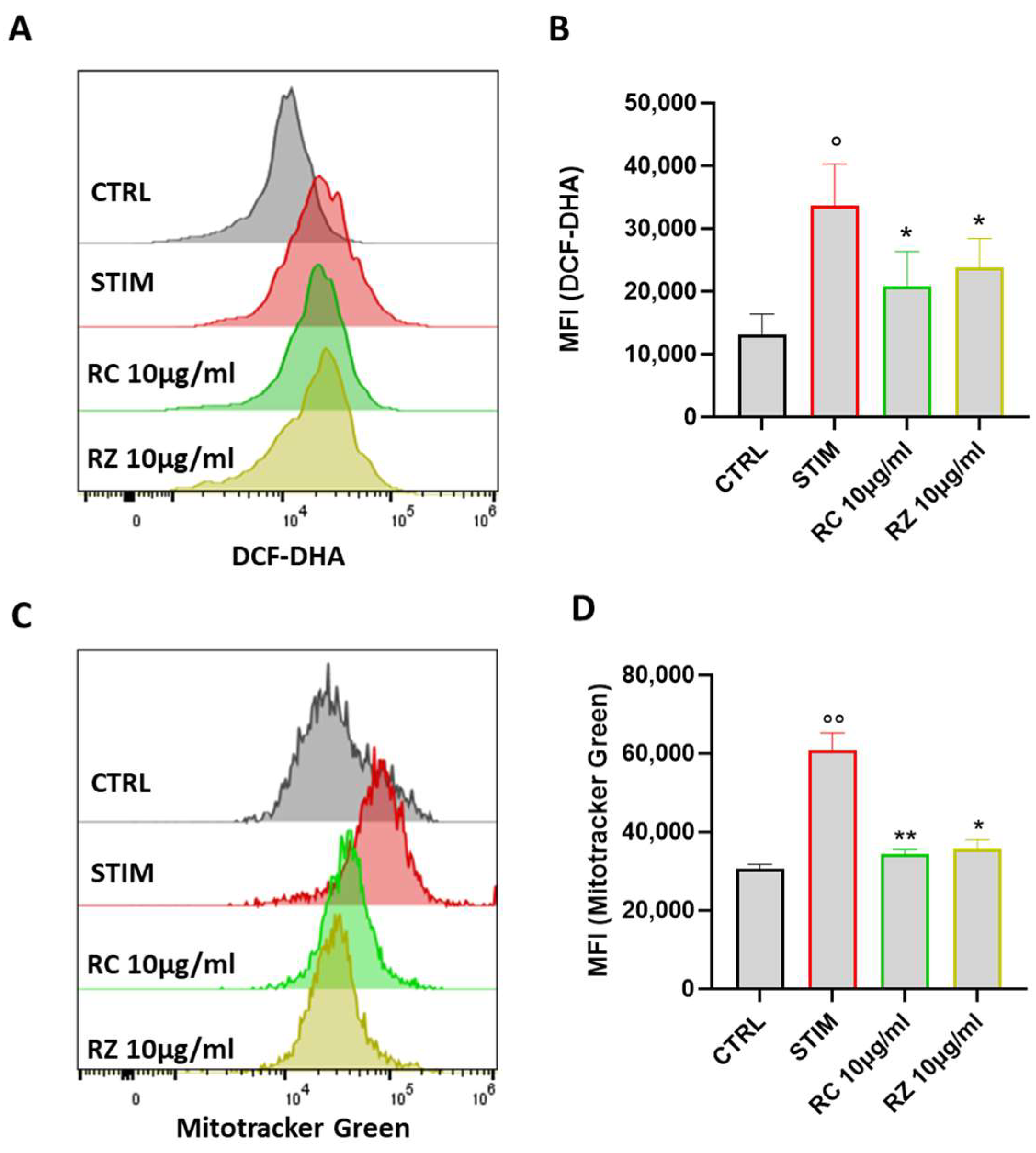
3.8. WRO and CRO Extracts Reduced ROS Production and Impaired Mitochondrial Function
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, L.; Ding, B.; Zhang, H.; Kong, B.; Xiong, Y.L. Textural and sensorial quality protection in frozen dumplings through the inhibition of lipid and protein oxidation with clove and rosemary extracts. J. Sci. Food Agric. 2019, 99, 4739–4747. [Google Scholar] [CrossRef] [PubMed]
- Bozin, B.; Mimica-Dukic, N.; Samojlik, I.; Jovin, E. Antimicrobial and antioxidant properties of rosemary and sage (Rosmarinus officinalis L. and Salvia officinalis L., Lamiaceae) essential oils. J. Agric. Food Chem. 2007, 55, 7879–7885. [Google Scholar] [CrossRef]
- Yamamoto, J.; Yamada, K.; Naemura, A.; Yamashita, T.; Arai, R. Testing various herbs for antithrombotic effect. Nutrition 2005, 21, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Erkan, N.; Ayranci, G.; Ayranci, E. Antioxidant activities of rosemary (Rosmarinus officinalis L.) extract, blackseed (Nigella sativa L.) essential oil, carnosic acid, rosmarinic acid and sesamol. Food Chem. 2008, 110, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Kontogianni, V.G.; Tomic, G.; Nikolic, I.; Nerantzaki, A.A.; Sayyad, N.; Stosic-Grujicic, S.; Stojanovic, I.; Gerothanassis, I.P.; Tzakos, A.G. Phytochemical profile of Rosmarinus officinalis and Salvia officinalis extracts and correlation to their antioxidant and anti-proliferative activity. Food Chem. 2013, 136, 120–129. [Google Scholar] [CrossRef]
- Pérez-Fons, L.; GarzÓn, M.T.; Micol, V. Relationship between the antioxidant capacity and effect of rosemary (Rosmarinus officinalis L.) polyphenols on membrane phospholipid order. J. Agric. Food Chem. 2010, 58, 161–171. [Google Scholar] [CrossRef]
- Andrade, J.M.; Faustino, C.; Garcia, C.; Ladeiras, D.; Reis, C.P.; Rijo, P. Rosmarinus officinalis L.: An update review of its phytochemistry and biological activity. Future Sci. OA 2018, 4, FSO283. [Google Scholar] [CrossRef]
- Sun, L.; Chen, W.; Meng, Y.; Yang, X.; Yuan, L.; Guo, Y. Interactions between polyphenols in thinned young apples and porcine pancreatic α-amylase: Inhibition, detailed kinetics and fluorescence quenching. Food Chem. 2016, 208, 51–60. [Google Scholar] [CrossRef]
- Kamli, M.R.; Sharaf, A.A.M.; Sabir, J.S.; Rather, I.A. Phytochemical Screening of Rosmarinus officinalis L. as a Potential Anticholinesterase and Antioxidant–Medicinal Plant for Cognitive Decline Disorders. Plants 2022, 11, 514. [Google Scholar] [CrossRef]
- Das, U.N. Acetylcholinesterase and butyrylcholinesterase as possible markers of low-grade systemic inflammation. Med. Sci. Monit. 2007, 13, RA214–RA221. [Google Scholar]
- Rao, A.A.; Reddy, C.S.; Sridhar, G.R.; Annapurna, A.; Hanuman, T.; Prameela, M.; Suresh, K.; Prasannalaxmi, S.; Das, U.N. Enhanced butyrylcholinesterase activity may be the common link in triggering low-grade systemic inflammation and decrease in cognitive function in diabetes mellitus and Alzheimer’s disease. Curr. Nutr. Food Sci. 2008, 4, 213–216. [Google Scholar] [CrossRef]
- Matull, W.R.; Pereira, S.P.; O’Donohue, J.W. Biochemical markers of acute pancreatitis. J. Clin. Pathol. 2006, 59, 340–344. [Google Scholar] [CrossRef]
- Lamponi, S.; Baratto, M.C.; Miraldi, E.; Baini, G.; Biagi, M. Chemical Profile, Antioxidant, Anti-Proliferative, Anticoagulant and Mutagenic Effects of a Hydroalcoholic Extract of Tuscan Rosmarinus officinalis. Plants 2021, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yuan, J.; Liu, F.; Ye, J. Determination of active components in rosemary by capillary electrophoresis with electrochemical detection. J. Pharm. Biomed. Anal. 2005, 39, 431–437. [Google Scholar] [CrossRef] [PubMed]
- El Bouzidi, L.; Jamali, C.A.; Bekkouche, K.; Hassani, L.; Wohlmuth, H.; Leach, D.; Abbad, A. Chemical composition, antioxidant and antimicrobial activities of essential oils obtained from wild and cultivated Moroccan Thymus species. Ind. Crops Prod. 2013, 43, 450–456. [Google Scholar] [CrossRef]
- Lubbe, A.; Verpoorte, R. Cultivation of medicinal and aromatic plants for specialty industrial materials. Ind. Crops Prod. 2011, 34, 785–801. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of total phenolics. Curr. Protoc. Food Anal. Chem. 2002, 6, I1-1. [Google Scholar]
- Baba, S.A.; Malik, S.A. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Med. Sci. 2015, 9, 449–454. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]
- De Martino, L.; Amato, G.; Caputo, L.; Nazzaro, F.; Scognamiglio, M.R.; De Feo, V. Variations in composition and bioactivity of Ocimum basilicum cv ‘Aroma 2’ essential oils. Ind. Crops Prod. 2021, 172, 114068. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Amamcharla, J.K.; Metzger, L.E. Modification of the ferric reducing antioxidant power (FRAP) assay to determine the susceptibility of raw milk to oxidation. Int. Dairy J. 2014, 34, 177–179. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres Jr, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Caputo, L.; Amato, G.; De Martino, L.; De Feo, V.; Nazzaro, F. Anti-Cholinesterase and Anti-α-Amylase Activities and Neuroprotective Effects of Carvacrol and p-Cymene and Their Effects on Hydrogen Peroxide Induced Stress in SH-SY5Y Cells. Int. J. Mol. Sci. 2023, 24, 6073. [Google Scholar] [CrossRef] [PubMed]
- Bernfeld, P. [17] Amylases, α and β. Methods Enzymol. 1955, 1, 149–158. [Google Scholar] [CrossRef]
- De Cicco, P.; Busà, R.; Ercolano, G.; Formisano, C.; Allegra, M.; Taglialatela-Scafati, O.; Ianaro, A. Inhibitory effects of cynaropicrin on human melanoma progression by targeting MAPK, NF-κB, and Nrf-2 signaling pathways In vitro. Phytother. Res. 2021, 35, 1432–1442. [Google Scholar] [CrossRef]
- Hossain, M.B.; Rai, D.K.; Brunton, N.P.; Martin-Diana, A.B.; Barry-Ryan, C. Characterization of phenolic composition in Lamiaceae spices by LC-ESI-MS/MS. J. Agric. Food Chem. 2010, 58, 10576–10581. [Google Scholar] [CrossRef] [PubMed]
- Linares, I.B.; Arráez-Román, D.; Herrero, M.; Ibáñez, E.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Comparison of different extraction procedures for the comprehensive characterization of bioactive phenolic compounds in Rosmarinus officinalis by reversed-phase high-performance liquid chromatography with diode array detection coupled to electrospray time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 7682–7690. [Google Scholar] [CrossRef]
- Cuvelier, M.E.; Richard, H.; Berset, C. Antioxidative activity and phenolic composition of pilot-plant and commercial extracts of sage and rosemary. J. Am. Oil Chem. Soc. 1996, 73, 645–652. [Google Scholar] [CrossRef]
- Borrás-Linares, I.; Stojanović, Z.; Quirantes-Piné, R.; Arráez-Román, D.; Švarc-Gajić, J.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Rosmarinus officinalis leaves as a natural source of bioactive compounds. Int. J. Mol. Sci. 2014, 15, 20585–20606. [Google Scholar] [CrossRef]
- Okamura, N.; Haraguchi, H.; Hashimoto, K.; Yagi, A. Flavonoids in Rosmarinus officinalis leaves. Phytochemistry 1994, 37, 1463–1466. [Google Scholar] [CrossRef] [PubMed]
- Grayer, R.J.; Bryan, S.E.; Veitch, N.C.; Goldstone, F.J.; Paton, A.; Wollenweber, E. External flavones in sweet basil, Ocimum basilicum, and related taxa. Phytochemistry 1996, 43, 1041–1047. [Google Scholar] [CrossRef]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage polarization: Different gene signatures in M1 (LPS+) vs. classically and M2 (LPS–) vs. alternatively activated macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Suzuki, J.; Kobayashi, Y. Role of calcium in tumor necrosis factor-α production by activated macrophages. J. Biochem. 1996, 120, 1190–1195. [Google Scholar] [CrossRef]
- Gronski, M.A.; Kinchen, J.M.; Juncadella, I.J.; Franc, N.C.; Ravichandran, K.S. An essential role for calcium flux in phagocytes for apoptotic cell engulfment and the anti-inflammatory response. Cell Death Differ. 2009, 16, 1323–1331. [Google Scholar] [CrossRef]
- Palmieri, E.M.; McGinity, C.; Wink, D.A.; McVicar, D.W. Nitric oxide in macrophage immunometabolism: Hiding in plain sight. Metabolites 2020, 10, 429. [Google Scholar] [CrossRef]
- Palmieri, E.M.; Gonzalez-Cotto, M.; Baseler, W.A.; Davies, L.C.; Ghesquière, B.; Maio, N.; Rice, C.M.; Rouault, T.A.; Cassel, T.; Higashi, R.M.; et al. Nitric oxide orchestrates metabolic rewiring in M1 macrophages by targeting aconitase 2 and pyruvate dehydrogenase. Nat. Commun. 2020, 11, 698. [Google Scholar] [CrossRef]
- Aktan, F. NOS2-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef]
- Kang, Y.J.; Mbonye, U.R.; DeLong, C.J.; Wada, M.; Smith, W.L. Regulation of intracellular cyclooxygenase levels by gene transcription and protein degradation. Prog. Lipid Res. 2007, 46, 108–125. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Veleva, R.; Moskova-Doumanova, V.; Doumanov, J.; Kapchina-Toteva, V.; Topouzova-Hristova, T. Comparative analysis of biological activity of extracts from cultivated and wild plants Lamium album L. Sci. Technol. 2014, IV, 122–126. [Google Scholar]
- Wang, Y.Z.; Fu, S.G.; Wang, S.Y.; Yang, D.J.; Wu, Y.H.S.; Chen, Y.C. Effects of a natural antioxidant, polyphenol-rich rosemary (Rosmarinus officinalis L.) extract, on lipid stability of plant-derived omega-3 fatty-acid rich oil. LWT 2018, 89, 210–216. [Google Scholar] [CrossRef]
- Bourhia, M.; Laasri, F.E.; Aourik, H.; Boukhris, A.; Ullah, R.; Bari, A.; Ali, S.S.; El Mzibri, M.; Benbacer, L.; Gmouh, S. Antioxidant and antiproliferative activities of bioactive compounds contained in Rosmarinus officinalis used in the Mediterranean diet. Evid. Based Complement. Altern. Med. 2019, 2019, 7623830. [Google Scholar] [CrossRef]
- Marrelli, M.; Cristaldi, B.; Menichini, F.; Conforti, F. Inhibitory effects of wild dietary plants on lipid peroxidation and on the proliferation of human cancer cells. FCT 2015, 86, 16–24. [Google Scholar] [CrossRef]
- Conforti, F.; Marrelli, M.; Carmela, C.; Menichini, F.; Valentina, P.; Uzunov, D.; Statti, G.A.; Duez, P.; Menichini, F. Bioactive phytonutrients (omega fatty acids, tocopherols, polyphenols), In vitro inhibition of nitric oxide production and free radical scavenging activity of non-cultivated Mediterranean vegetables. Food Chem. 2011, 129, 1413–1419. [Google Scholar] [CrossRef]
- Ali, M.B.; Mnafgui, K.; Feki, A.; Damak, M.; Allouche, N. In vitro Antidiabetic, anti-obesity and antioxidant properties of Rosemary extracts. J. Adv. Chem. 2014, 10, 2305–2316. [Google Scholar] [CrossRef]
- Ho, C.T.; Wang, M.; Wei, G.J.; Huang, T.C.; Huang, M.T. Chemistry and antioxidative factors in rosemary and sage. Biofactors 2000, 13, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Lešnik, S.; Furlan, V.; Bren, U. Rosemary (Rosmarinus officinalis L.): Extraction techniques, analytical methods and health-promoting biological effects. Phytochem. Rev. 2021, 20, 1273–1328. [Google Scholar] [CrossRef]
- Carvalho Jr, R.N.; Moura, L.S.; Rosa, P.T.; Meireles, M.A.A. Supercritical fluid extraction from rosemary (Rosmarinus officinalis): Kinetic data, extract’s global yield, composition, and antioxidant activity. J. Supercrit. Fluids 2005, 35, 197–204. [Google Scholar] [CrossRef]
- Jordán, M.J.; Lax, V.; Rota, M.C.; Lorán, S.; Sotomayor, J.A. Effect of the phenological stage on the chemical composition, and antimicrobial and antioxidant properties of Rosmarinus officinalis L. essential oil and its polyphenolic extract. Ind. Crops Prod. 2013, 48, 144–152. [Google Scholar] [CrossRef]
- Kabubii, Z.N.; Mbaria, J.M.; Mathiu, M.P.; Wanjohi, J.M.; Nyaboga, E.N. Evaluation of seasonal variation, effect of extraction solvent on phytochemicals and antioxidant activity on Rosmarinus officinalis grown in different agro-ecological zones of Kiambu County, Kenya. CABI Agric. Biosci. 2023, 4, 1. [Google Scholar] [CrossRef]
- Saini, A.; Pandey, A.; Sharma, S.; Suradkar, U.S.; Ambedkar, Y.R.; Meena, P.; Raman, M.; Gurjar, A.S. Assessment of antioxidant activity of rosemary (Rosmarinus officinalis) leaves extract. J. Pharmacogn. Phytochem 2020, 9, 14–17. [Google Scholar]
- Santos, R.D.; Shetty, K.; da Silva Miglioranza, L.H. Oxidative stability of butter with added phenolics from Lamiaceae herbs and In vitro evaluation of potential cytotoxicity of rosemary (Rosmarinus officinalis L.) extract. Int. J. Food Sci. 2014, 49, 768–775. [Google Scholar] [CrossRef]
- Massoulié, J. The origin of the molecular diversity and functional anchoring of cholinesterases. Neurosignals 2002, 11, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Kolak, U.; Hacibekiroğlu, I.; Öztürk, M.; Özgökçe, F.; Topcu, G.; Ulubelen, A. Antioxidant and anticholinesterase constituents of Salvia poculata. Turk. J. Chem. 2009, 33, 813–823. [Google Scholar] [CrossRef]
- Çulhaoğlu, B.; Yapar, G.; Dirmenci, T.; Topçu, G. Bioactive constituents of Salvia chrysophylla Stapf. Nat. Prod. Res. 2013, 27, 438–447. [Google Scholar] [CrossRef]
- Mata, A.T.; Proença, C.; Ferreira, A.R.; Serralheiro, M.L.M.; Nogueira, J.M.F.; Araújo, M.E.M. Antioxidant and antiacetylcholinesterase activities of five plants used as Portuguese food spices. Food Chem. 2007, 103, 778–786. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S.; Jamous, R.M.; Zaitoun, S.Y.A.; Qasem, I.B. In-vitro screening of acetylcholinesterase inhibitory activity of extracts from Palestinian indigenous flora in relation to the treatment of Alzheimer’s disease. Funct. Foods Health Dis. 2014, 4, 381–400. [Google Scholar] [CrossRef]
- Kawamura-Konishi, Y.; Watanabe, N.; Saito, M.; Nakajima, N.; Sakaki, T.; Katayama, T.; Enomoto, T. Isolation of a new phlorotannin, a potent inhibitor of carbohydrate-hydrolyzing enzymes, from the brown alga Sargassum patens. J. Agric. Food Chem. 2012, 60, 5565–5570. [Google Scholar] [CrossRef]
- Lo Piparo, E.; Scheib, H.; Frei, N.; Williamson, G.; Grigorov, M.; Chou, C.J. Flavonoids for controlling starch digestion: Structural requirements for inhibiting human α-amylase. J. Med. Chem. 2008, 51, 3555–3561. [Google Scholar] [CrossRef]
- Buchholz, T.; Melzig, M.F. Medicinal plants traditionally used for treatment of obesity and diabetes mellitus–screening for pancreatic lipase and α-Amylase inhibition. Phytother. Res. 2016, 30, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Mansouri Torghabeh, F.; Rostamzadeh, P.; Davoudi, S.; Keivan, M.; Shokri-Asl, V. Effects of Rosmarinus officinalis on orchitis following spermatic cord torsion-detorsion in male mice with emphasis on anti-inflammatory and antioxidant properties. Andrologia 2022, 54, e14252. [Google Scholar] [CrossRef] [PubMed]
- Grigore, A.; Vulturescu, V.; Neagu, G.; Ungureanu, P.; Panteli, M.; Rasit, I. Antioxidant–Anti-Inflammatory Evaluation of a Polyherbal Formula. Pharmaceuticals 2022, 15, 114. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Anti-Inflammatory Therapeutic Mechanisms of Natural Products: Insight from Rosemary Diterpenes, Carnosic Acid and Carnosol. Biomedicines 2023, 11, 545. [Google Scholar] [CrossRef]
- Danisman, B.; Cicek, B.; Yildirim, S.; Bolat, I.; Kantar, D.; Golokhvast, K.S.; Nikitovic, D.; Tsatsakis, A.; Taghizadehghalehjoughi, A. Carnosic Acid Ameliorates Indomethacin-Induced Gastric Ulceration in Rats by Alleviating Oxidative Stress and Inflammation. Biomedicines 2023, 11, 829. [Google Scholar] [CrossRef]
- Yamamoto, T.; Xing, J.; Katayama, Y.; Tsubokawa, T.; Hirayama, T.; Maejima, S. Spinal cord responses to feline transcranial brain stimulation: Evidence for involvement of cerebellar pathways. J. Neurotrauma 1990, 7, 247–256. [Google Scholar] [CrossRef]
- Hwang, C.J.; Choi, J.Y.; Park, M.H.; Song, M.J.; Oh, K.W.; Son, D.J.; Lee, S.H.; Han, S.B.; Hong, J.T. Inhibitory effect of carnosol on phthalic anhydride-induced atopic dermatitis via inhibition of STAT3. Biomol. Ther. 2017, 25, 535. [Google Scholar] [CrossRef]
- Guimarães, N.S.; Ramos, V.S.; Prado-Souza, L.F.; Lopes, R.M.; Arini, G.S.; Feitosa, L.G.; Silva, R.R.; Nantes, I.L.; Damasceno, D.C.; Lopes, N.P.; et al. Rosemary (Rosmarinus officinalis L.) Glycolic Extract Protects Liver Mitochondria from Oxidative Damage and Prevents Acetaminophen-Induced Hepatotoxicity. Antioxidants 2023, 12, 628. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Family | tR (min) | Measured m/z | Molecular Formula | Identification | WRO | CRO |
|---|---|---|---|---|---|---|---|
| 1 | flavonoid glycoside | 8.45 | 479.1167 | C22H22O12 | Isorhamnetin-3-O-glucoside | X | X |
| 2 | flavonoid glycoside | 8.78 | 463.1220 | C22H22O11 | Homoplantaginin (Hispidulin-7- Glucoside) | X | X |
| 3 | polyphenol | 8.94 | 361.0909 | C18H16O8 | Rosmarinic acid | X | X |
| 4 | flavonoid glycoside | 9.43 | 505.0963 | C23H21O13 | Luteolin-3′-O-(3″-O-acetyl)-β-D-glucuronide | X | X |
| 5 | flavonoid glycoside | 9.55 | 505.0957 | C23H21O13 | Luteolin-3′-O-(2″-O-acetyl)-β-D-glucuronide | X | X |
| 6 | flavonoid | 9.83 | 317.0644 | C16H12O7 | Isorhamnetin | X | |
| 7 | flavonoid | 10.53 | 301.0699 | C16H12O6 | Diosmetin | X | |
| 8 | flavonoid | 11.30 | 315.0853 | C17H14O6 | Pectolinarigenin | X | X |
| 9 | flavonoid | 11.93 | 285.0754 | C16H12O5 | Genkwanin | X | X |
| 10 | terpenoid | 12.97 | 329.1008 | C18H16O6 | Salvigenin | X | X |
| 11 | terpenoid | 13.24 | 331.1898 | C20H26O4 | Carnosol | X | X |
| 12 | terpenoid | 13.75 | 317.2103 | C20H28O3 | Rosmaridiphenol | X | X |
| 13 | terpenoid | 14.08 | 333.2052 | C20H28O4 | Carnosic acid | X | X |
| 14 | terpenoid | 14.30 | 345.2049 | C21H28O4 | Rosmadial | X |
| Compound | Amount mg/g | |
|---|---|---|
| WRO | CRO | |
| 1 | 9.03 | 72.93 |
| 2 | 30.21 | 313.12 |
| 3 | 6840.56 | 6494.65 |
| 4 | 1.33 | 265.35 |
| 5 | 0.15 | 22.49 |
| 6 | 0.00 | 1373.01 |
| 7 | 0.00 | 169.21 |
| 8 | 4.06 | 258.07 |
| 10 | 203.74 | 1128.92 |
| 11 | 60.52 | 38.48 |
| 13 | 1.77 | 46.39 |
| TPC mg GAE 1/g Extract (Mean ± SD)4 | TFC mg QE 2/g Extract (Mean ± SD)4 | DPPH IC50 3(µg/mL) (Mean ± SD)4 | FRAP mmol Fe2+ Equivalents/g Extract (Mean ± SD) 4 | |
|---|---|---|---|---|
| WRO | 129.36 ± 9.23 a | 61.67 ± 2.33 a | 17.58 ± 0.98 a | 2.25 ± 0.55 c |
| CRO | 175.64 ± 42.61 a | 49.47 ± 6.35 b | 14.93 ± 0.92 b | 3.66 ± 0.72 b |
| Trolox | / | / | 3.65 ± 0.08 c | 5.48 ± 0.34 a |
| AChE IC50 1 (µg/mL) (Mean ± SD) 2 | BChE IC501 (µg/mL) (Mean ± SD) 2 | α-Amylase IC501 (µg/mL) (Mean ± SD) 2 | |
|---|---|---|---|
| WRO | 96.39 ± 3.22 b | 243.35 ± 11.25 a | 40.52 ± 3.57 b |
| CRO | 216.50 ± 19.90 a | 257.20 ± 5.50 a | 52.68 ± 4.38 a |
| Galantamine | 0.79 ± 0.01 c | 1.97 ± 0.14 b | / |
| Acarbose | / | / | 0.95 ± 0.31 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francolino, R.; Martino, M.; Caputo, L.; Amato, G.; Chianese, G.; Gargiulo, E.; Formisano, C.; Romano, B.; Ercolano, G.; Ianaro, A.; et al. Phytochemical Constituents and Biological Activity of Wild and Cultivated Rosmarinus officinalis Hydroalcoholic Extracts. Antioxidants 2023, 12, 1633. https://doi.org/10.3390/antiox12081633
Francolino R, Martino M, Caputo L, Amato G, Chianese G, Gargiulo E, Formisano C, Romano B, Ercolano G, Ianaro A, et al. Phytochemical Constituents and Biological Activity of Wild and Cultivated Rosmarinus officinalis Hydroalcoholic Extracts. Antioxidants. 2023; 12(8):1633. https://doi.org/10.3390/antiox12081633
Chicago/Turabian StyleFrancolino, Rosaria, Mara Martino, Lucia Caputo, Giuseppe Amato, Giuseppina Chianese, Ernesto Gargiulo, Carmen Formisano, Benedetta Romano, Giuseppe Ercolano, Angela Ianaro, and et al. 2023. "Phytochemical Constituents and Biological Activity of Wild and Cultivated Rosmarinus officinalis Hydroalcoholic Extracts" Antioxidants 12, no. 8: 1633. https://doi.org/10.3390/antiox12081633