
Components of the Glutathione Cycle as Markers of Biological Age: An Approach to Clinical Application in Aging
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Objective 1: To Verify Whether Oxidative Stress in Leukocytes is Associated with Immunosenescence
2.1.2. Objective 2: To Identify the Best Blood Fraction to Measure the Glutathione Cycle Parameters
2.2. Participants and Peripheral Blood Extraction
2.3. Obtaining Different Blood Fractions: Whole Blood, Total Blood Cells, Isolated Erythrocytes, Plasma, and Leukocytes
2.4. Immune Function Tests
2.5. Calculation of ImmunolAge Using the Immunity Clock
2.6. Assessment of the Components of the Glutathione Cycle
2.7. Statistical Analysis
3. Results
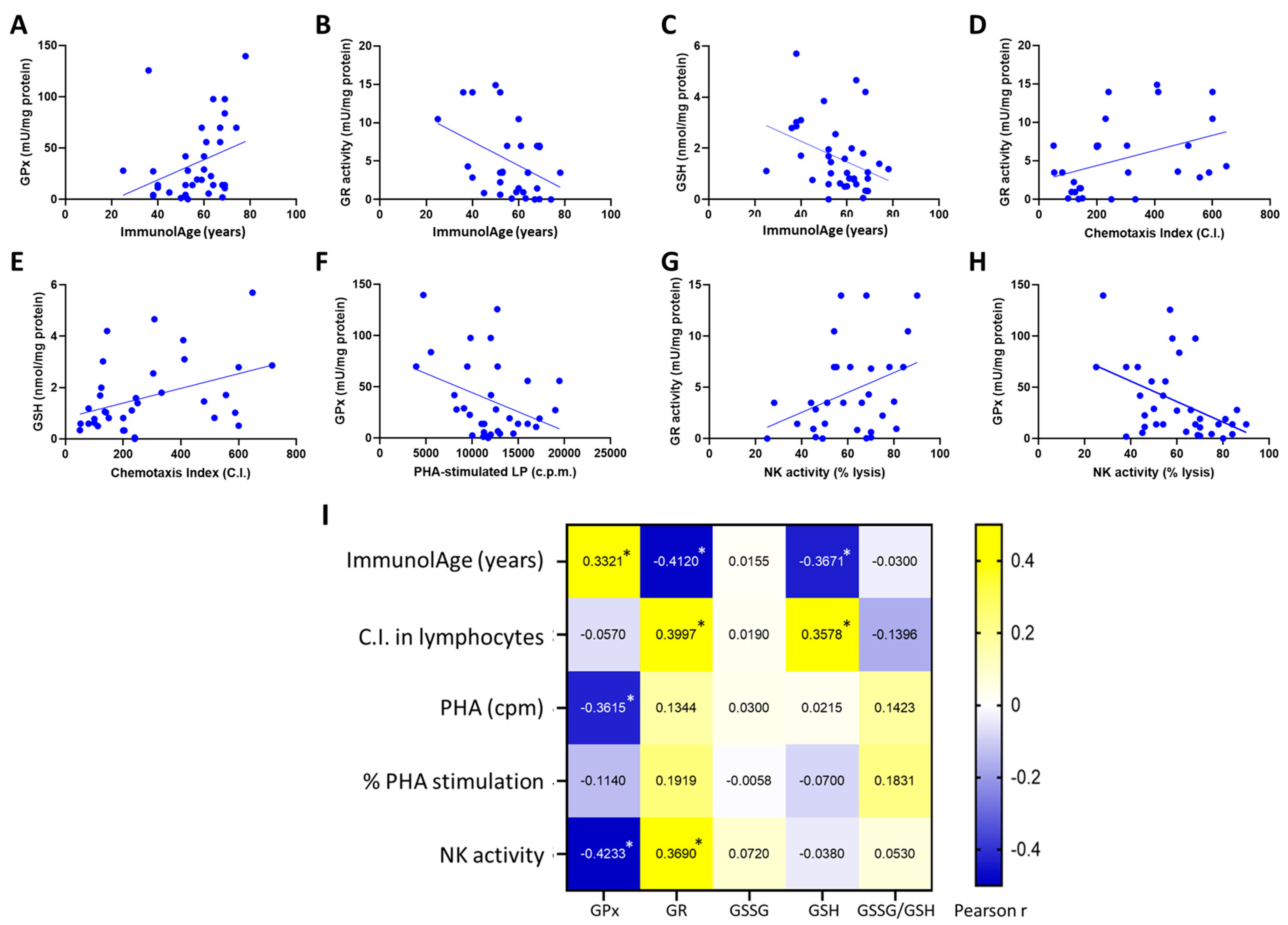
3.1. Parameters of the Glutathione Cycle Correlate with the Functionality of Peripheral Neutrophils and with ImmunolAge
3.2. Parameters of the Glutathione Cycle Correlate with the Functionality of Peripheral Lymphocytes and with ImmunolAge
3.3. Blood Cells Are the Most Suitable Blood Fraction to Assess Parameters of the Glutathione Cycle to Estimate the Rate of Aging in Humans
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De la Fuente, M.; Miquel, J. An update of the oxidation-inflammation theory of aging: The involvement of the immune system in oxi-inflamm-aging. Curr. Pharm. Des. 2009, 15, 3003–3026. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miquel, J.; Economos, A.C.; Fleming, J.; Johnson, J.E., Jr. Mitochondrial role in cell aging. Exp. Gerontol. 1980, 15, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. Aging in vertebrates and the effect of caloric restriction: A mitochondrial free radical production-DNA damage mechanism? Biol. Rev. 2004, 79, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. Free radicals and aging. Trends Neurosci. 2004, 27, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Barja, G. The mitochondrial free radical theory of aging. Prog. Mol. Biol. Transl. Sci. 2014, 127, 1–27. [Google Scholar]
- Sedlak, T.W.; Paul, B.D.; Parker, G.M.; Hester, L.D.; Snowman, A.M.; Taniguchi, Y.; Kamiya, A.; Snyder, S.H.; Sawa, A. The glutathione cycle shapes synaptic glutamate activity. Proc. Natl. Acad. Sci. USA 2019, 116, 2701–2706. [Google Scholar] [CrossRef] [Green Version]
- Labarrere, C.A.; Kassab, G.S. Glutathione: A Samsonian life-sustaining small molecule that protects against oxidative stress, ageing and damaging inflammation. Front. Nutr. 2022, 9, 1007816. [Google Scholar] [CrossRef]
- Barros, M.P.; Rodrigo, M.J.; Zacarias, L. Dietary Carotenoid Roles in Redox Homeostasis and Human Health. J. Agric. Food Chem. 2018, 66, 5733–5740. [Google Scholar] [CrossRef] [Green Version]
- Iskusnykh, I.Y.; Zakharova, A.A.; Pathak, D. Glutathione in Brain Disorders and Aging. Molecules 2022, 27, 324. [Google Scholar] [CrossRef]
- Stohs, S.J.; Lawson, T.; Al-Turk, W.A. Changes in glutathione and glutathione metabolizing enzymes in erythrocytes and lymphocytes of mice as a function of age. Gen. Pharmacol. 1984, 15, 267–270. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, M.Y.; Day, W.W.; Zamorano, D.M. Glutathione and lipid peroxidation in the aging rat. Comp. Biochem. Physiol. B 1987, 88, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, L.S.; Machado, P.E. Age-related changes of glutathione content, glutathione reductase and glutathione peroxidase activity of human erythrocytes. Braz. J. Med. Biol. Res. 1991, 24, 449–454. [Google Scholar] [PubMed]
- Lang, C.A.; Naryshkin, S.; Schneider, D.L.; Mills, B.J.; Lindeman, R.D. Low blood glutathione levels in healthy aging adults. J. Lab. Clin. Med. 1992, 120, 720–725. [Google Scholar] [PubMed]
- Loguercio, C.; Taranto, D.; Vitale, L.M.; Beneduce, F.; Del Vecchio Blanco, C. Effect of liver cirrhosis and age on the glutathione concentration in the plasma, erythrocytes, and gastric mucosa of man. Free Radic. Biol. Med. 1996, 20, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Rikans, L.E.; Hornbrook, K.R. Lipid peroxidation, antioxidant protection and aging. Biochim. Biophys. Acta 1997, 1362, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Samiec, P.S.; Drews-Botsch, C.; Flagg, E.W.; Kurtz, J.C.; Sternberg, P., Jr.; Reed, R.L.; Jones, D.P. Glutathione in human plasma: Decline in association with aging, age-related macular degeneration, and diabetes. Free Radic. Biol. Med. 1998, 24, 99–704. [Google Scholar] [CrossRef]
- Liu, R.; Choi, J. Age-associated decline in gamma-glutamylcysteine synthetase gene expression in rats. Free Radic. Biol. Med. 2000, 28, 566–574. [Google Scholar] [CrossRef]
- Erden-Inal, M.; Sunal, E.; Kanbak, G. Age-related changes in the glutathione redox system. Cell Biochem. Funct. 2002, 20, 61–66. [Google Scholar] [CrossRef]
- Rebrin, I.; Sohal, R.S. Pro-oxidant shift in glutathione redox state during aging. Adv. Drug. Deliv. Rev. 2008, 60, 1545–1552. [Google Scholar] [CrossRef] [Green Version]
- Martínez de Toda, I.; Vida, C.; Garrido, A.; De la Fuente, M. Redox Parameters as Markers of the Rate of Aging and Predictors of Life Span. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2020, 75, 613–620. [Google Scholar] [CrossRef]
- Martínez de Toda, I.; Maté, I.; Vida, C.; Cruces, J.; De la Fuente, M. Immune function parameters as markers of biological age and predictors of longevity. Aging 2016, 8, 3110–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez de Toda, I.; Ceprián, N.; Díaz-Del Cerro, E.; De la Fuente, M. The Role of Immune Cells in Oxi-Inflamm-Aging. Cells 2021, 10, 2974. [Google Scholar] [CrossRef]
- Martínez de Toda, I.; Vida, C.; Díaz-Del Cerro, E.; De la Fuente, M. The Immunity Clock. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 1939–1945. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef]
- Massey, V.; Williams, C.H., Jr. On the reaction mechanism of yeast glutathione reductase. J. Biol. Chem. 1965, 240, 4470–4480. [Google Scholar] [CrossRef] [PubMed]
- Hissin, P.J.; Hilf, R. A fluorometric method for determination of oxidized and reduced glutathione in tissues. Anal. Biochem. 1976, 74, 214–226. [Google Scholar] [CrossRef] [PubMed]
- United Nations Department of Economic and Social Affairs. Population Division. In World Population Ageing 2020 Highlights: Living Arrangements of Older Persons (ST/ESA/SER.A/451); United Nations Department of Economic and Social Affairs: New York, NY, USA, 2020. [Google Scholar]
- Martínez de Toda, I.; Vida, C.; Sanz San Miguel, L.; De la Fuente, M. When will my mouse die? Life span prediction based on immune function, redox and behavioural parameters in female mice at the adult age. Mech. Ageing Dev. 2019, 182, 111125. [Google Scholar] [CrossRef]
- Martínez de Toda, I.; Vida, C.; Sanz San Miguel, L.; De la Fuente, M. Function, Oxidative, and Inflammatory Stress Parameters in Immune Cells as Predictive Markers of Lifespan throughout Aging. Oxid. Med. Cell. Longev. 2019, 2019, 4574276. [Google Scholar] [CrossRef] [Green Version]
- Garrido, A.; Cruces, J.; Ceprián, N.; Vara, E.; De la Fuente, M. Oxidative-Inflammatory Stress in Immune Cells from Adult Mice with Premature Aging. Int. J. Mol. Sci. 2019, 20, 769. [Google Scholar] [CrossRef] [Green Version]
- Veskoukis, A.S.; Nikolaidis, M.G.; Kyparos, A.; Kouretas, D. Blood reflects tissue oxidative stress depending on biomarker and tissue studied. Free Radic. Biol. Med. 2009, 47, 1371–1374. [Google Scholar] [CrossRef] [PubMed]
- Margaritelis, N.V.; Veskoukis, A.S.; Paschalis, V.; Vrabas, I.S.; Dipla, K.; Zafeiridis, A.; Kyparos, A.; Nikolaidis, M.G. Blood reflects tissue oxidative stress: A systematic review. Biomarkers 2015, 20, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Giustarini, D.; Dalle-Donne, I.; Colombo, R.; Milzani, A.; Rossi, R. An improved HPLC measurement for GSH and GSSG in human blood. Free Radic. Biol. Med. 2003, 35, 1365–1372. [Google Scholar] [CrossRef]
- Hernanz, A.; Fernández-Vivancos, E.; Montiel, C.; Vazquez, J.J.; Arnalich, F. Changes in the intracellular homocysteine and glutathione content associated with aging. Life Sci. 2000, 67, 1317–1324. [Google Scholar] [CrossRef] [PubMed]
- Dröge, W. Oxidative aging and insulin receptor signaling. J. Gerontol. A Biol. Sci. Med. Sci. 2005, 60, 1378–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115, Correction in Genome Biol. 2015, 16, 96. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Epel, E. Stress and telomere shortening: Insights from cellular mechanisms. Ageing Res. Rev. 2022, 73, 101507. [Google Scholar] [CrossRef]
- Blackburn, E.H.; Greider, C.W.; Szostak, J.W. Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nat. Med. 2006, 12, 1133–1138. [Google Scholar] [CrossRef]
- Lucas, H.R.; Rifkind, J.M. Considering the vascular hypothesis of Alzheimer’s disease: Effect of copper associated amyloid on red blood cells. Adv. Exp. Med. Biol. 2013, 765, 131–138. [Google Scholar] [PubMed]
- Jacob, K.D.; Noren Hooten, N.; Trzeciak, A.R.; Evans, M.K. Markers of oxidant stress that are clinically relevant in aging and age-related disease. Mech. Ageing Dev. 2013, 134, 139–157. [Google Scholar] [CrossRef] [Green Version]
- Elson, C.J.; Jablonska, K.F.; Taylor, R.B. Functional half-life of virgin and primed B lymphocytes. Eur. J. Immunol. 1976, 6, 634–638. [Google Scholar] [CrossRef]
- Tak, T.; Tesselaar, K.; Pillay, J.; Borghans, J.A.; Koenderman, L. What’s your age again? Determination of human neutrophil half-lives revisited. J. Leukoc. Biol. 2013, 94, 595–601. [Google Scholar] [CrossRef]
- Nayar, S.; Dasgupta, P.; Galustian, C. Extending the lifespan and efficacies of immune cells used in adoptive transfer for cancer immunotherapies—A review. Oncoimmunology 2015, 19, e1002720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeandel, C.; Nicolas, M.B.; Dubois, F.; Nabet-Belleville, F.; Penin, F.; Cuny, G. Lipid peroxidation and free radical scavengers in Alzheimer’s disease. Gerontology 1989, 35, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Bourdel-Marchasson, I.; Delmas-Beauvieux, M.C.; Peuchant, E.; Richard-Harston, S.; Decamps, A.; Reignier, B.; Emeriau, J.P.; Rainfray, M. Antioxidant defences and oxidative stress markers in erythrocytes and plasma from normally nourished elderly Alzheimer patients. Age Ageing 2001, 30, 235–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casado, A.; Castellanos, A.; López-Fernández, M.E.; Ruíz, R.; Aroca, C.G.; Noriega, F. Relationship between oxidative and occupational stress and aging in nurses of an intensive care unit. Age 2008, 30, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junqueira, V.B.; Barros, S.B.; Chan, S.S.; Rodrigues, L.; Giavarotti, L.; Abud, R.L.; Deucher, G.P. Aging and oxidative stress. Mol. Aspects Med. 2004, 25, 5–16. [Google Scholar] [CrossRef]
- Ismail, T.; Kim, Y.; Lee, H.; Lee, D.S.; Lee, H.S. Interplay Between Mitochondrial Peroxiredoxins and ROS in Cancer Development and Progression. Int. J. Mol. Sci. 2019, 20, 4407. [Google Scholar] [CrossRef] [Green Version]
- Detienne, G.; De Haes, W.; Mergan, L.; Edwards, S.L.; Temmerman, L.; Van Bael, S. Beyond ROS clearance: Peroxiredoxins in stress signaling and aging. Ageing Res. Rev. 2018, 44, 33–48. [Google Scholar] [CrossRef]
- Oláhová, M.; Taylor, S.R.; Khazaipoul, S.; Wang, J.; Morgan, B.A.; Matsumoto, K.; Blackwell, T.K.; Veal, E.A. A redox-sensitive peroxiredoxin that is important for longevity has tissue- and stress-specific roles in stress resistance. Proc. Natl. Acad. Sci. USA 2008, 105, 19839–19844. [Google Scholar] [CrossRef]
- Xiao, Z.; La Fontaine, S.; Bush, A.I.; Wedd, A.G. Molecular Mechanisms of Glutaredoxin Enzymes: Versatile Hubs for Thiol-Disulfide Exchange between Protein Thiols and Glutathione. J. Mol. Biol. 2019, 431, 158–177. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaz-Del Cerro, E.; Martinez de Toda, I.; Félix, J.; Baca, A.; De la Fuente, M. Components of the Glutathione Cycle as Markers of Biological Age: An Approach to Clinical Application in Aging. Antioxidants 2023, 12, 1529. https://doi.org/10.3390/antiox12081529
Diaz-Del Cerro E, Martinez de Toda I, Félix J, Baca A, De la Fuente M. Components of the Glutathione Cycle as Markers of Biological Age: An Approach to Clinical Application in Aging. Antioxidants. 2023; 12(8):1529. https://doi.org/10.3390/antiox12081529
Chicago/Turabian StyleDiaz-Del Cerro, Estefania, Irene Martinez de Toda, Judith Félix, Adriana Baca, and Monica De la Fuente. 2023. "Components of the Glutathione Cycle as Markers of Biological Age: An Approach to Clinical Application in Aging" Antioxidants 12, no. 8: 1529. https://doi.org/10.3390/antiox12081529