
β-Asarone Alleviates High-Glucose-Induced Oxidative Damage via Inhibition of ROS Generation and Inactivation of the NF-κB/NLRP3 Inflammasome Pathway in Human Retinal Pigment Epithelial Cells
 , , , and
, , , and 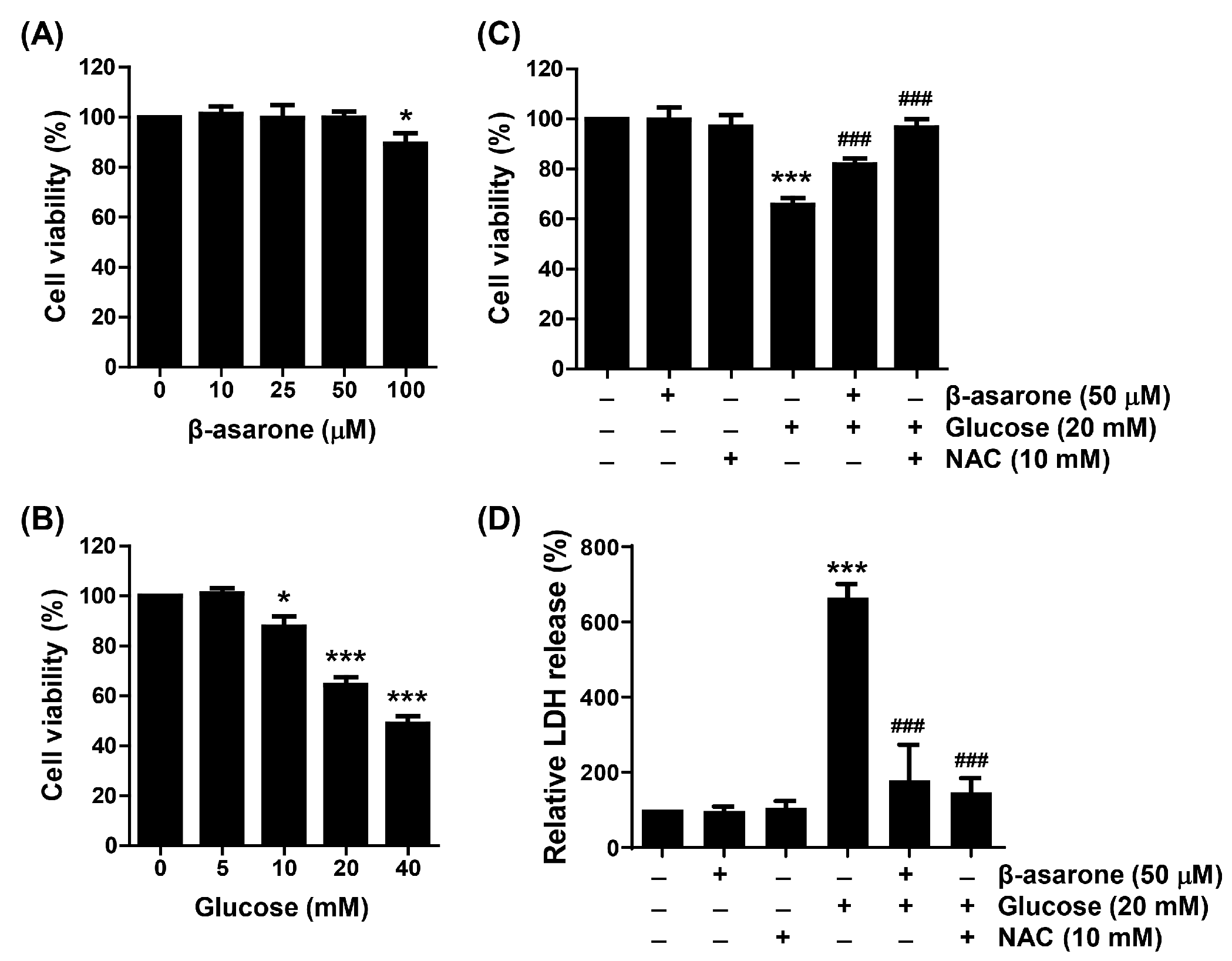{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatment
2.2. Cell Viability and Lactate Dehydrogenase (LDH) Assays
2.3. Determination of the Cell and Nuclear Morphology
2.4. Apoptosis Assay
2.5. Western Blot Analysis
2.6. Assessment of DNA Damage
2.7. ROS Assay
2.8. Evaluation of the Antioxidant Defense System
2.9. Measurement of IL-1β and IL-18 Levels
2.10. Immunofluorescence Assay for NF-κB, NLRP3, and ASC
2.11. Assay of Caspase-1 Activity
2.12. Statistical Analysis
3. Results
3.1. β-Asarone Ameliorates HG-Induced Cell Viability Reduction and Cytotoxicity in ARPE-19 Cells
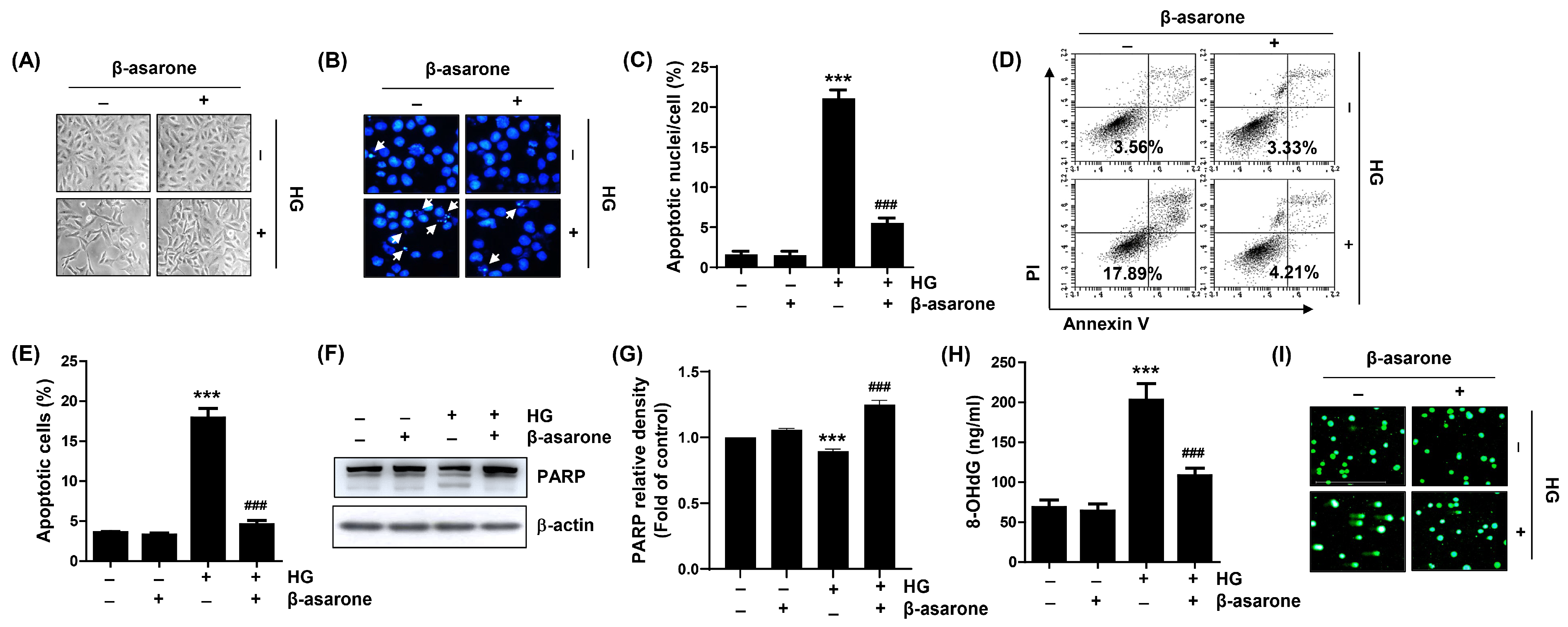
3.2. β-Asarone Reduces Apoptosis and DNA Damage in HG-Treated ARPE-19 Cells
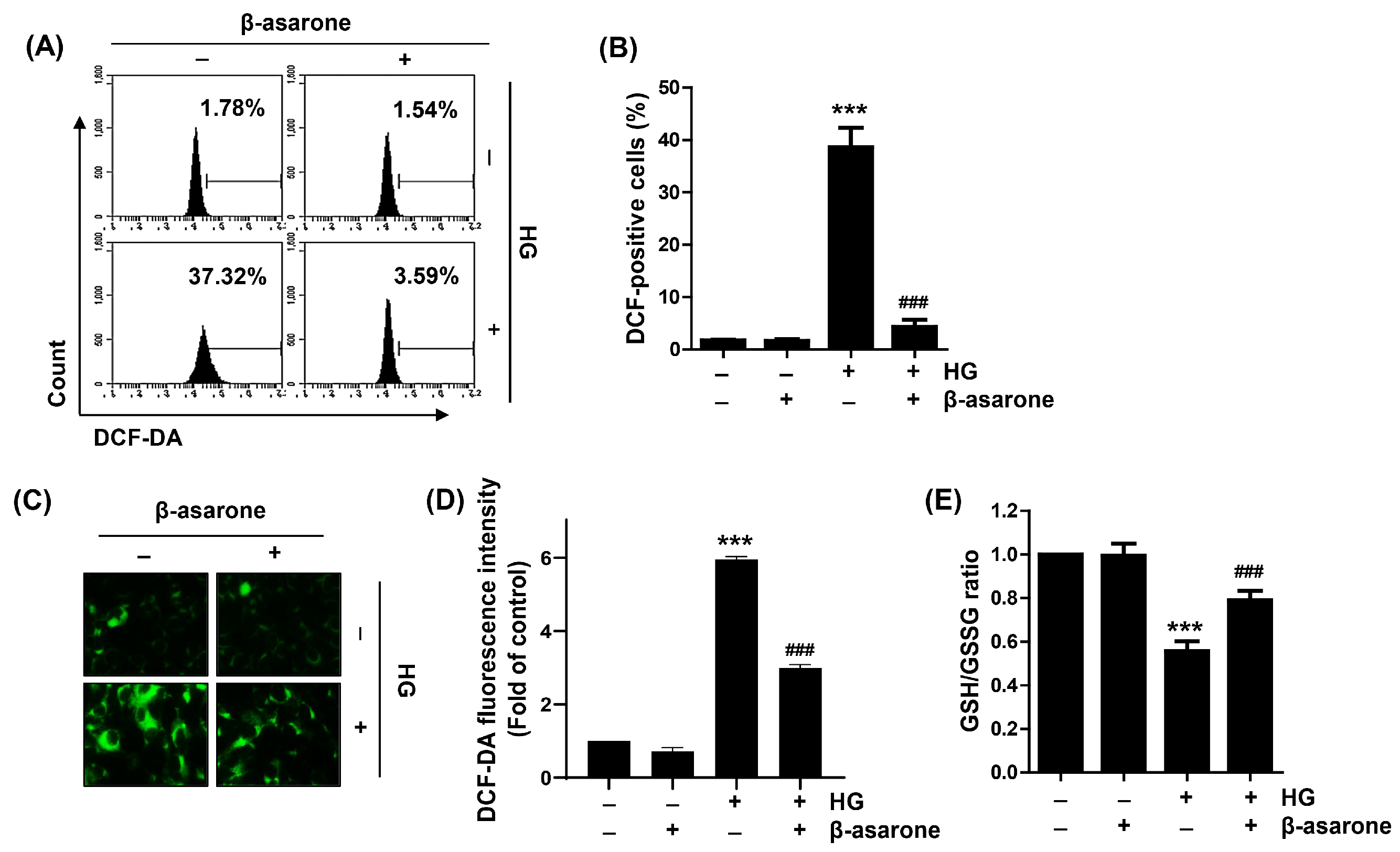
3.3. β-Asarone Decreases ROS Generation in HG-Treated ARPE-19 Cells
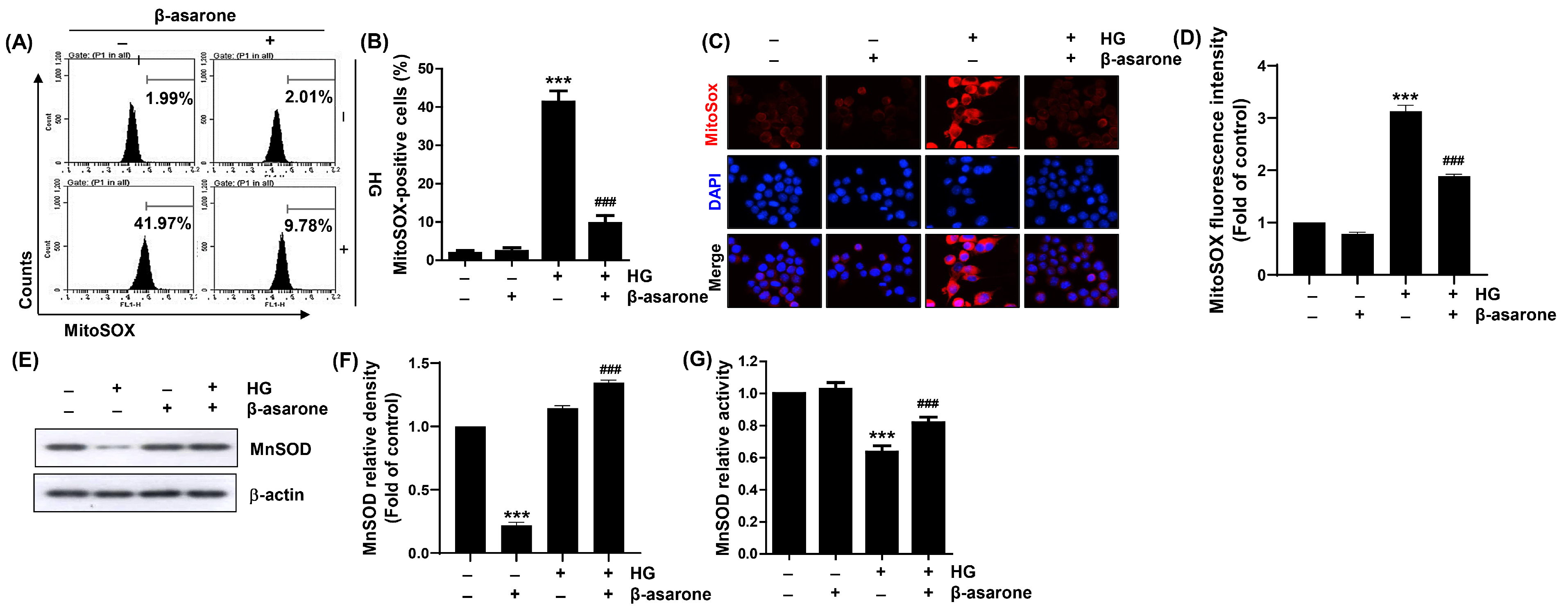
3.4. β-Asarone Attenuates HG-induced Production of Mitochondrial Superoxide Radicals and Inactivation of MnSOD in ARPE-19 Cells
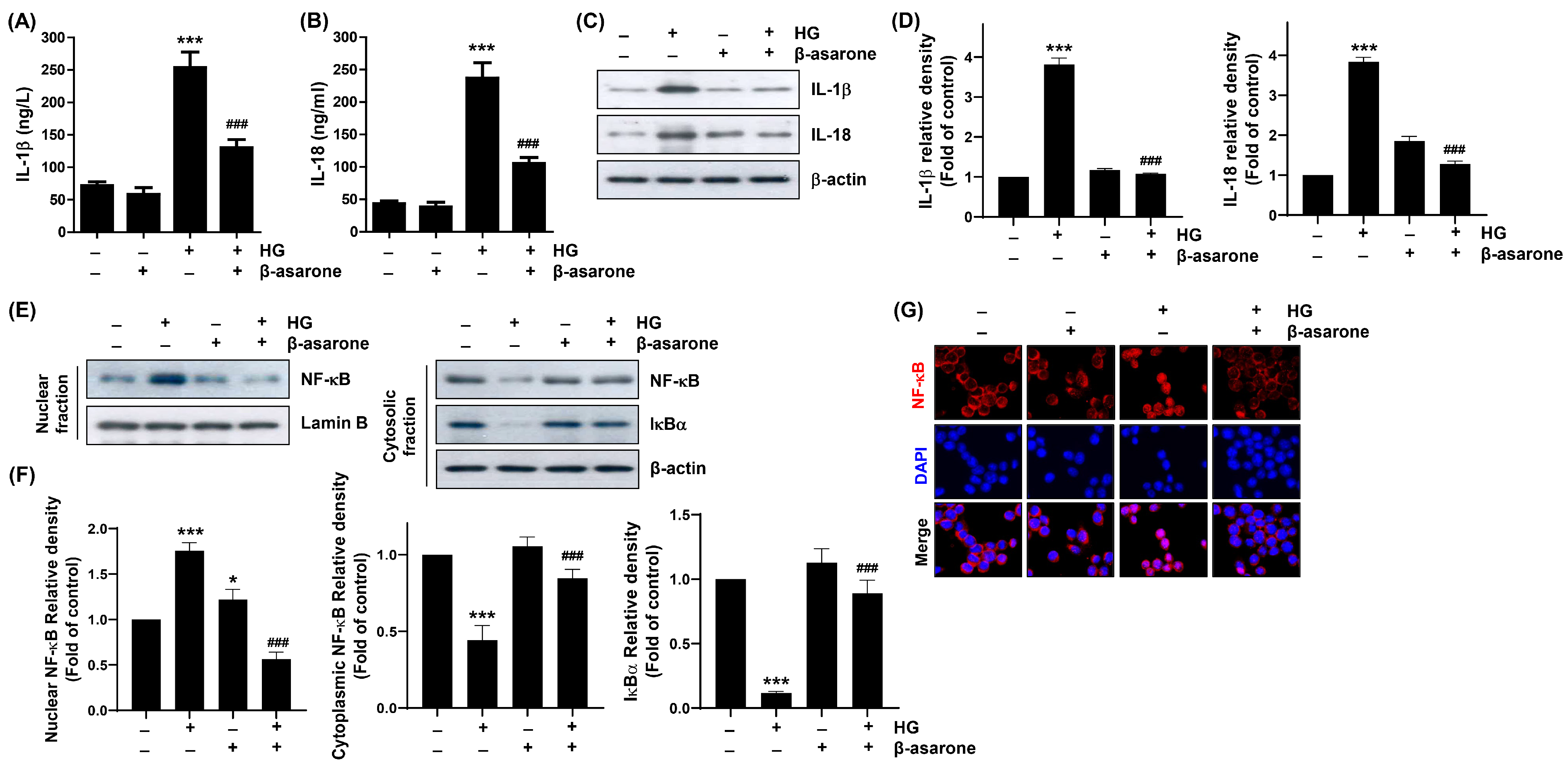
3.5. β-Asarone Alleviates HG-Induced Inflammatory Response in ARPE-19 Cells
3.6. β-Asarone Mitigates NF-κB Signaling Activation in HG-Treated ARPE-19 Cells
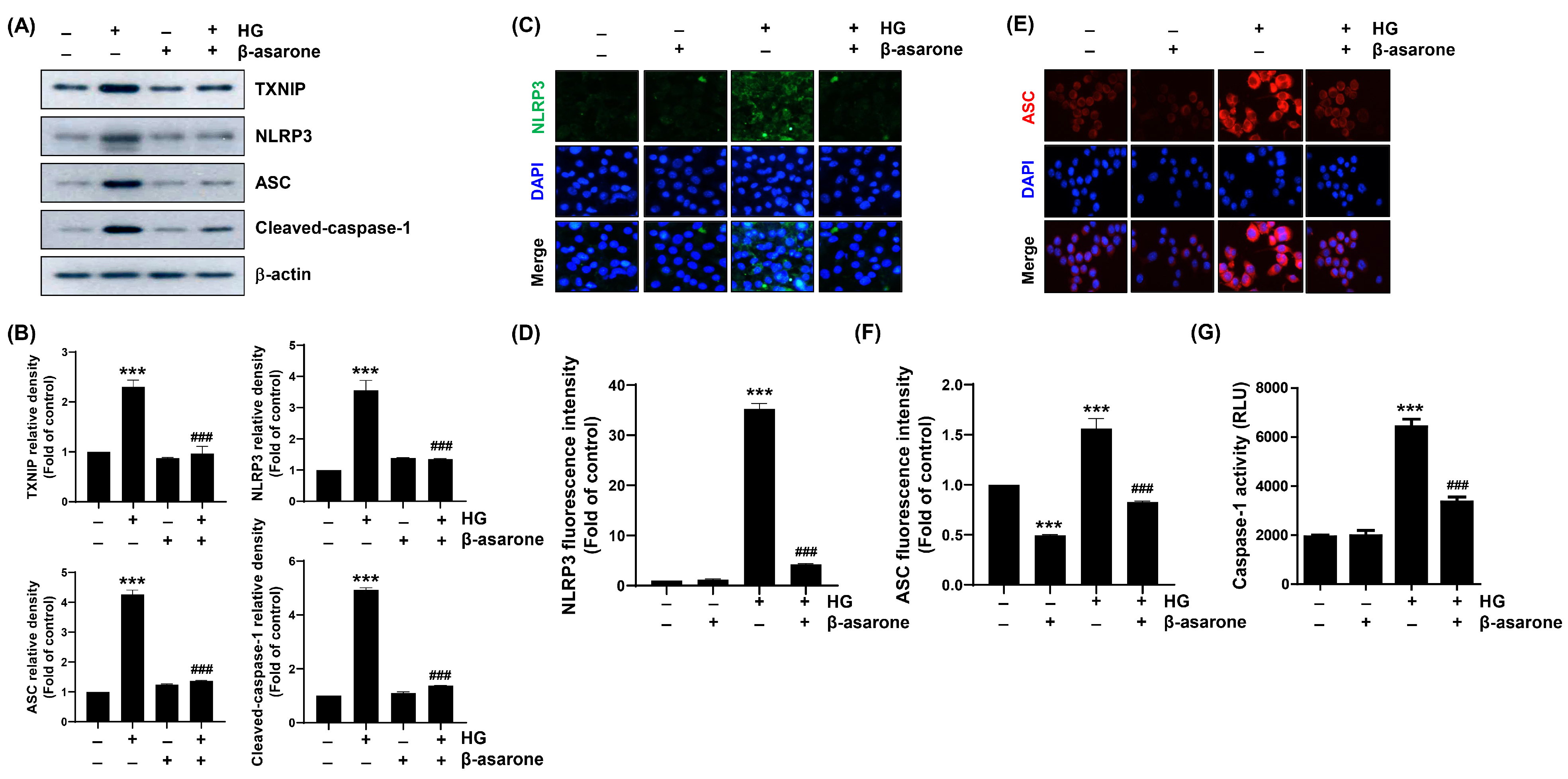
3.7. β-Asarone Attenuates HG-Induced NLRP3 Inflammasome Activation in ARPE-19 Cells
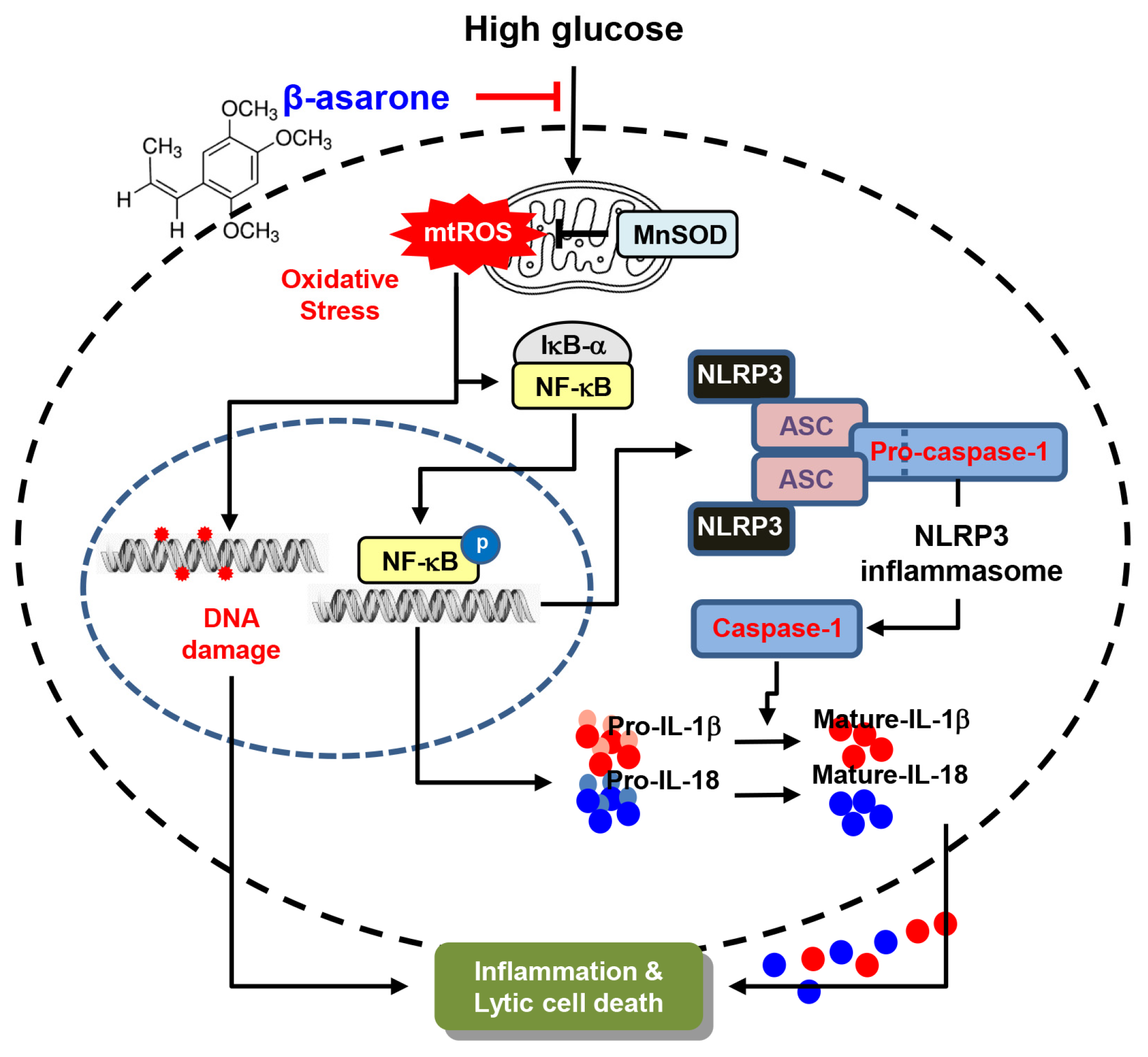
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, Y.; Ren, T.; Wang, N. Biomechanical homeostasis in ocular diseases: A mini-review. Front. Public Health 2023, 11, 1106728. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Xu, S.; Jiang, L.; Tan, Z.; Wang, J. A systematic pan-cancer analysis of CASP3 as a potential target for immunotherapy. Front. Mol. Biosci. 2022, 9, 776808. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, M.; Geng, Z.; Khattak, S.; Ji, X.; Wu, D.; Dang, Y. Role of oxidative stress in retinal disease and the early intervention strategies: A review. Oxid. Med. Cell. Longev. 2022, 2022, 7836828. [Google Scholar] [CrossRef] [PubMed]
- Dammak, A.; Huete-Toral, F.; Carpena-Torres, C.; Martin-Gil, A.; Pastrana, C.; Carracedo, G. From oxidative stress to inflammation in the posterior ocular diseases: Diagnosis and treatment. Pharmaceutics 2021, 13, 1376. [Google Scholar] [CrossRef] [PubMed]
- Kang, Q.; Yang, C. Oxidative stress and diabetic retinopathy: Molecular mechanisms, pathogenetic role and therapeutic implications. Redox Biol. 2020, 37, 101799. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, P.; Mai, J.; Choi, E.T.; Wang, H.; Yang, X.F. Targeting mitochondrial reactive oxygen species as novel therapy for inflammatory diseases and cancers. J. Hematol. Oncol. 2013, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Patergnani, S.; Bouhamida, E.; Leo, S.; Pinton, P.; Rimessi, A. Mitochondrial oxidative stress and “Mito-Inflammation”: Actors in the diseases. Biomedicines 2021, 9, 216. [Google Scholar] [CrossRef]
- Naik, E.; Dixit, V.M. Mitochondrial reactive oxygen species drive proinflammatory cytokine production. J. Exp. Med. 2011, 208, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.Y.; Maran, J.J.; Jamieson, E.G.; Rupenthal, I.D.; Murphy, R.; Mugisho, O.O. Characterization of NLRP3 inflammasome activation in the onset of diabetic retinopathy. Int. J. Mol. Sci. 2022, 23, 14471. [Google Scholar] [CrossRef]
- Zhao, M.; Li, S.; Matsubara, J.A. Targeting pyroptotic cell death pathways in retinal disease. Front. Med. 2022, 8, 802063. [Google Scholar] [CrossRef]
- Al Mamun, A.; Mimi, A.A.; Zaeem, M.; Wu, Y.; Monalisa, I.; Akter, A.; Munir, F.; Xiao, J. Role of pyroptosis in diabetic retinopathy and its therapeutic implications. Eur. J. Pharmacol. 2021, 904, 174166. [Google Scholar] [CrossRef]
- Xiaodong, L.; Xuejun, X. GSDMD-mediated pyroptosis in retinal vascular inflammatory diseases: A review. Int. Ophthalmol. 2023, 43, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.R.; Wieser, M.E.; Ganga, R.R.; Barathi, V.A.; Lakshminarayanan, R.; Mohan, R.R.; Hainsworth, D.P.; Chaurasia, S.S. NOD-like receptors in the eye: Uncovering its role in diabetic retinopathy. Int. J. Mol. Sci. 2020, 21, 899. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhang, X.; Liao, N.; Mi, L.; Peng, Y.; Liu, B.; Zhang, S.; Wen, F. Enhanced expression of NLRP3 inflammasome-related inflammation in diabetic retinopathy. Invest. Ophthalmol. Vis. Sci. 2018, 59, 978–985. [Google Scholar] [CrossRef]
- Paerewijck, O.; Lamkanfi, M. The human inflammasomes. Mol. Asp. Med. 2022, 88, 101100. [Google Scholar] [CrossRef] [PubMed]
- Duez, H.; Pourcet, B. Nuclear receptors in the control of the NLRP3 inflammasome pathway. Front. Endocrinol. 2021, 12, 630536. [Google Scholar] [CrossRef] [PubMed]
- Mathur, A.; Hayward, J.A.; Man, S.M. Molecular mechanisms of inflammasome signaling. J. Leukoc. Biol. 2018, 103, 233–257. [Google Scholar] [CrossRef] [Green Version]
- Mandell, J.T.; de Rivero Vaccari, J.P.; Sabater, A.L.; Galor, A. The inflammasome pathway: A key player in ocular surface and anterior segment diseases. Surv. Ophthalmol. 2023, 68, 280–289. [Google Scholar] [CrossRef]
- Wen, J.; Yang, Y.; Hao, J. Acori tatarinowii Rhizoma: A comprehensive review of its chemical composition, pharmacology, pharmacokinetics and toxicity. Front. Pharmacol. 2023, 14, 1090526. [Google Scholar] [CrossRef]
- Chellian, R.; Pandy, V.; Mohamed, Z. Pharmacology and toxicology of α- and β-asarone: A review of preclinical evidence. Phytomedicine 2017, 32, 41–58. [Google Scholar] [CrossRef]
- Lim, H.W.; Kumar, H.; Kim, B.W.; More, S.V.; Kim, I.W.; Park, J.I.; Park, S.Y.; Kim, S.K.; Choi, D.K. β-asarone (cis-2,4,5-trimethoxy-1-allyl phenyl), attenuates pro-inflammatory mediators by inhibiting NF-κB signaling and the JNK pathway in LPS activated BV-2 microglia cells. Food Chem. Toxicol. 2014, 72, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Hermes, L.; Haupenthal, S.; Uebel, T.; Esselen, M. DNA double strand break repair as cellular response to genotoxic asarone isomers considering phase I metabolism. Food Chem. Toxicol. 2020, 142, 111484. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Cai, M.; Xia, W.; Jiang, L.; Song, H.; Chen, X. Explore the mechanism of β-asarone on improving cognitive dysfunction in rats with diabetic encephalopathy. J. Alzheimers Dis. Rep. 2022, 6, 195–206. [Google Scholar] [CrossRef]
- Xiao, B.; Huang, X.; Wang, Q.; Wu, Y. β-asarone alleviates myocardial ischemia-reperfusion injury by inhibiting inflammatory response and NLRP3 inflammasome mediated pyroptosis. Biol. Pharm. Bull. 2020, 43, 1046–1051. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.H.; Park, C.; Hwangbo, B.; Bang, E.J.; Kim, S.O.; Shim, J.H.; Park, S.H.; Lee, H.; Leem, S.H.; Kim, G.Y.; et al. Activation of heme oxygenase-1 is involved in the preventive effect of honokiol against oxidative damage in human retinal pigment epithelial cells. Biotechnol. Bioprocess Eng. 2022, 27, 975–986. [Google Scholar] [CrossRef]
- Kim, C.; Ryu, S.H.; Choi, H.; Park, D.H.; Bae, J.S. The inhibitory functions of sparstolonin B against ambient fine particulate matter induced lung injury. Biotechnol. Bioprocess Eng. 2022, 27, 949–960. [Google Scholar] [CrossRef]
- Choi, Y.H. Tacrolimus induces apoptosis in leukemia Jurkat cells through inactivation of the reactive oxygen species-dependent phosphoinositide-3-kinase/Akt signaling pathway. Biotechnol. Bioprocess Eng. 2022, 27, 183–192. [Google Scholar] [CrossRef]
- di Vito, R.; Levorato, S.; Fatigoni, C.; Acito, M.; Sancineto, L.; Traina, G.; Villarini, M.; Santi, C.; Moretti, M. In vitro toxicological assessment of PhSeZnCl in human liver cells. Toxicol. Res. 2023, 39, 105–114. [Google Scholar] [CrossRef]
- Park, C.; Lee, H.; Kim, S.O.; Lee, E.W.; Lee, H.T.; Kwon, H.J.; Kim, B.W.; Kim, G.Y.; Kim, M.R.; Choi, Y.H. The preventive effect of Mori Ramulus on oxidative stress-induced cellular damage in skeletal L6 myoblasts through Nrf2-mediated activation of HO-1. Toxicol. Res. 2022, 39, 25–36. [Google Scholar] [CrossRef]
- Kim, M.Y.; Bang, E.; Hwangbo, H.; Ji, S.Y.; Kim, D.H.; Lee, H.; Park, C.; Hong, S.H.; Kim, G.Y.; Choi, Y.H. Diallyl trisulfide inhibits monosodium urate-induced NLRP3 inflammasome activation via NOX3/4-dependent mitochondrial oxidative stress in RAW 264.7 and bone marrow-derived macrophages. Phytomedicine 2023, 112, 154705. [Google Scholar] [CrossRef]
- Hwangbo, H.; Kim, M.Y.; Ji, S.Y.; Kim, S.Y.; Lee, H.; Kim, G.Y.; Park, C.; Keum, Y.S.; Hong, S.H.; Cheong, J.; et al. Auranofin attenuates non-alcoholic fatty liver disease by suppressing lipid accumulation and NLRP3 inflammasome-mediated hepatic inflammation in vivo and in vitro. Antioxidants 2020, 9, 1040. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Choi, M.; Yun, J.W. Trans-anethole induces thermogenesis via activating SERCA/SLN axis in C2C12 muscle cells. Biotechnol. Bioprocess Eng. 2022, 27, 938–948. [Google Scholar] [CrossRef]
- AbuArrah, M.; Yuli Setianto, B.; Faisal, A.; Hamim Sadewa, A. 8-Hydroxy-2-deoxyguanosine as oxidative DNA damage biomarker of medical ionizing radiation: A scoping review. J. Biomed. Phys. Eng. 2021, 11, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Møller, P.; Stopper, H.; Collins, A.R. Measurement of DNA damage with the comet assay in high-prevalence diseases: Current status and future directions. Mutagenesis 2020, 35, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, X.; Chen, B.; Dai, R.; Xi, Z.; Xu, H. Insights into manganese superoxide dismutase and human diseases. Int. J. Mol. Sci. 2022, 23, 15893. [Google Scholar] [CrossRef]
- Indo, H.P.; Hawkins, C.L.; Nakanishi, I.; Matsumoto, K.I.; Matsui, H.; Suenaga, S.; Davies, M.J.; St Clair, D.K.; Ozawa, T.; Majima, H.J. Role of mitochondrial reactive oxygen species in the activation of cellular signals, molecules, and function. Handb. Exp. Pharmacol. 2017, 240, 439–456. [Google Scholar]
- Mussbacher, M.; Derler, M.; Basílio, J.; Schmid, J.A. NF-κB in monocytes and macrophages—An inflammatory master regulator in multitalented immune cells. Front. Immunol. 2023, 14, 1134661. [Google Scholar] [CrossRef]
- Capece, D.; Verzella, D.; Flati, I.; Arboretto, P.; Cornice, J.; Franzoso, G. NF-κB: Blending metabolism, immunity, and inflammation. Trends Immunol. 2022, 43, 757–775. [Google Scholar] [CrossRef]
- Kaur, G.; Harris, N.R. Endothelial glycocalyx in retina, hyperglycemia, and diabetic retinopathy. Am. J. Physiol. Cell. Physiol. 2023, 324, C1061–C1077. [Google Scholar] [CrossRef]
- Rodríguez, M.L.; Millán, I.; Ortega, Á.L. Cellular targets in diabetic retinopathy therapy. World J. Diabetes 2021, 12, 1442–1462. [Google Scholar] [CrossRef]
- Jin, Y.; Arroo, R. The protective effects of flavonoids and carotenoids against diabetic complications-A review of in vivo evidence. Front. Nutr. 2023, 10, 1020950. [Google Scholar] [CrossRef] [PubMed]
- Khazeei Tabari, M.A.; Mirjalili, R.; Khoshhal, H.; Shokouh, E.; Khandan, M.; Hasheminasabgorji, E.; Hafezi-Moghadam, A.; Bagheri, A. Nature against diabetic retinopathy: A review on antiangiogenic, antioxidant, and anti-Inflammatory phytochemicals. Evid. Based Complement. Alternat. Med. 2022, 2022, 4708527. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.L.; Bruno, D.F.; Ambrósio, A.F.; Santos, P.F. Benefits of flavonoids in diabetic retinopathy. Nutrients 2020, 12, 3169. [Google Scholar] [CrossRef]
- Du, X.Y.; Cao, Y.S.; Yang, J.; Guo, L.C.; Zhang, T.; Yuan, Q.; Chen, X.; Hu, L.M. Preclinical evidence and possible mechanisms of β-asarone for rats and mice with Alzheimer’s disease: A systematic review and meta-analysis. Front. Pharmacol. 2022, 13, 956746. [Google Scholar] [CrossRef] [PubMed]
- Meng, M.; Zhang, L.; Ai, D.; Wu, H.; Peng, W. β-asarone ameliorates β-amyloid-induced neurotoxicity in PC12 cells by activating P13K/Akt/Nrf2 signaling pathway. Front. Pharmacol. 2021, 12, 659955. [Google Scholar] [CrossRef]
- Hei, X.; Xie, M.; Xu, J.; Li, J.; Liu, T. β-asarone exerts antioxidative effects on H2O2-stimulated PC12 cells by activating Nrf2/HO-1 pathway. Neurochem. Res. 2020, 45, 1953–1961. [Google Scholar] [CrossRef]
- Chang, W.; Teng, J. b-asarone prevents Aβ25-35-induced inflammatory responses and autophagy in SH-SY5Y cells: Down expression Beclin-1, LC3B and up expression Bcl-2. Int. J. Clin. Exp. Med. 2015, 8, 20658–20663. [Google Scholar]
- Daldal, H.; Nazıroğlu, M. Carvacrol protects the ARPE19 retinal pigment epithelial cells against high glucose-induced oxidative stress, apoptosis, and inflammation by suppressing the TRPM2 channel signaling pathways. Graefes. Arch. Clin. Exp. Ophthalmol. 2022, 260, 2567–2583. [Google Scholar] [CrossRef]
- Li, M.; Tian, M.; Jiang, X.; Liu, Y.; Wang, Y.; Li, Y. Inhibition of galectin-3 ameliorates high-glucose-induced oxidative stress and inflammation in ARPE-19 cells. Cutan. Ocul. Toxicol. 2022, 41, 179–186. [Google Scholar] [CrossRef]
- Chiang, Y.F.; Chen, H.Y.; Chang, Y.J.; Shih, Y.H.; Shieh, T.M.; Wang, K.L.; Hsia, S.M. Protective effects of fucoxanthin on high glucose- and 4-hydroxynonenal (4-HNE)-induced injury in human retinal pigment epithelial cells. Antioxidants 2020, 9, 1176. [Google Scholar] [CrossRef]
- Tokarz, P.; Kaarniranta, K.; Blasiak, J. Inhibition of DNA methyltransferase or histone deacetylase protects retinal pigment epithelial cells from DNA damage induced by oxidative stress by the stimulation of antioxidant enzymes. Eur. J. Pharmacol. 2016, 776, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiao, Y.; Li, X.; Gao, S.; Zhou, N.; Duan, J.; Zhang, M. Pyroptosis: A new insight into eye disease therapy. Front. Pharmacol. 2021, 12, 797110. [Google Scholar] [CrossRef] [PubMed]
- Singh, L.P.; Devi, T.S.; Yumnamcha, T. The role of Txnip in mitophagy dysregulation and inflammasome activation in diabetic retinopathy: A new perspective. JOJ Ophthalmol. 2017, 4, 555643. [Google Scholar] [CrossRef] [PubMed]
- Yumnamcha, T.; Devi, T.S.; Singh, L.P. Auranofin mediates mitochondrial dysregulation and inflammatory cell death in human retinal pigment epithelial cells: Implications of retinal neurodegenerative diseases. Front. Neurosci. 2019, 13, 1065. [Google Scholar] [CrossRef]
- Gan, J.; Huang, M.; Lan, G.; Liu, L.; Xu, F. High glucose induces the loss of retinal pericytes partly via NLRP3-caspase-1-GSDMD-mediated pyroptosis. Biomed. Res. Int. 2020, 2020, 4510628. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.; Cha, H.-J.; Hwangbo, H.; Bang, E.; Hong, S.H.; Song, K.S.; Noh, J.S.; Kim, D.-H.; Kim, G.-Y.; Choi, Y.H. β-Asarone Alleviates High-Glucose-Induced Oxidative Damage via Inhibition of ROS Generation and Inactivation of the NF-κB/NLRP3 Inflammasome Pathway in Human Retinal Pigment Epithelial Cells. Antioxidants 2023, 12, 1410. https://doi.org/10.3390/antiox12071410
Park C, Cha H-J, Hwangbo H, Bang E, Hong SH, Song KS, Noh JS, Kim D-H, Kim G-Y, Choi YH. β-Asarone Alleviates High-Glucose-Induced Oxidative Damage via Inhibition of ROS Generation and Inactivation of the NF-κB/NLRP3 Inflammasome Pathway in Human Retinal Pigment Epithelial Cells. Antioxidants. 2023; 12(7):1410. https://doi.org/10.3390/antiox12071410
Chicago/Turabian StylePark, Cheol, Hee-Jae Cha, Hyun Hwangbo, EunJin Bang, Su Hyun Hong, Kyoung Seob Song, Jeong Sook Noh, Do-Hyung Kim, Gi-Young Kim, and Yung Hyun Choi. 2023. "β-Asarone Alleviates High-Glucose-Induced Oxidative Damage via Inhibition of ROS Generation and Inactivation of the NF-κB/NLRP3 Inflammasome Pathway in Human Retinal Pigment Epithelial Cells" Antioxidants 12, no. 7: 1410. https://doi.org/10.3390/antiox12071410