
Potential of Dry-Cured Ham Bones as a Sustainable Source to Obtain Antioxidant and DPP-IV Inhibitory Extracts



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample Preparation and Enzymatic Hydrolysis
2.3. Antioxidant Activity
2.3.1. DPPH Radical Scavenging Activity
2.3.2. Ferric-Reducing Antioxidant Power (FRAP)
2.3.3. ABTS
2.4. DPP-IV Inhibitory Activity
2.5. Characterization of the Peptide Fraction
2.5.1. Peptide Separation by Reversed-Phase Chromatography (RP-HPLC)
2.5.2. Peptide Separation by Hydrophilic Interaction Liquid Chromatography (HILIC)
2.5.3. Identification of Peptides by Mass Spectrometry in Tandem
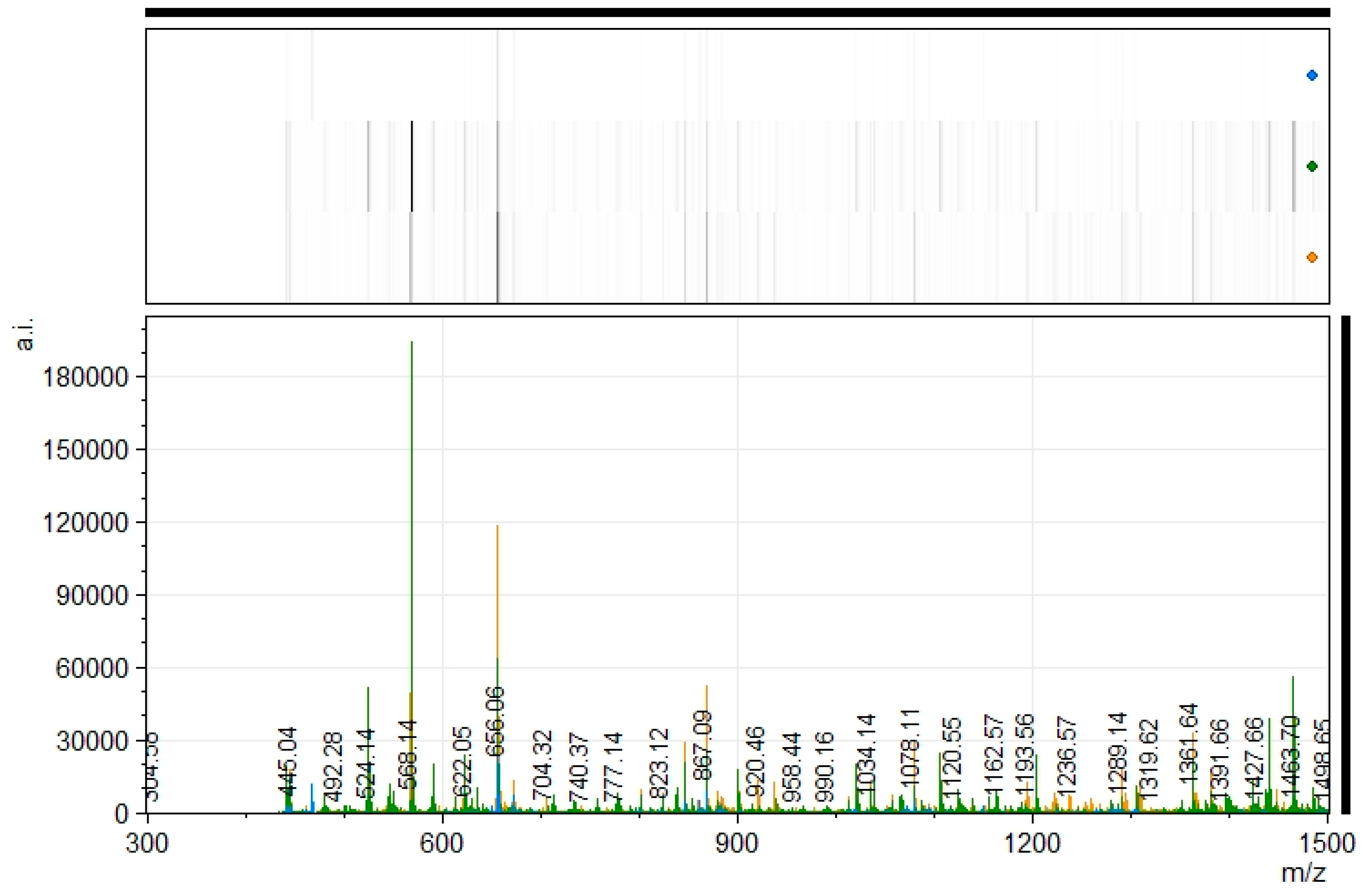
2.5.4. Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-ToF MS) Analysis
2.5.5. Free Amino Acids (FAAs)
2.6. Statistical Analysis
3. Results and Discussion
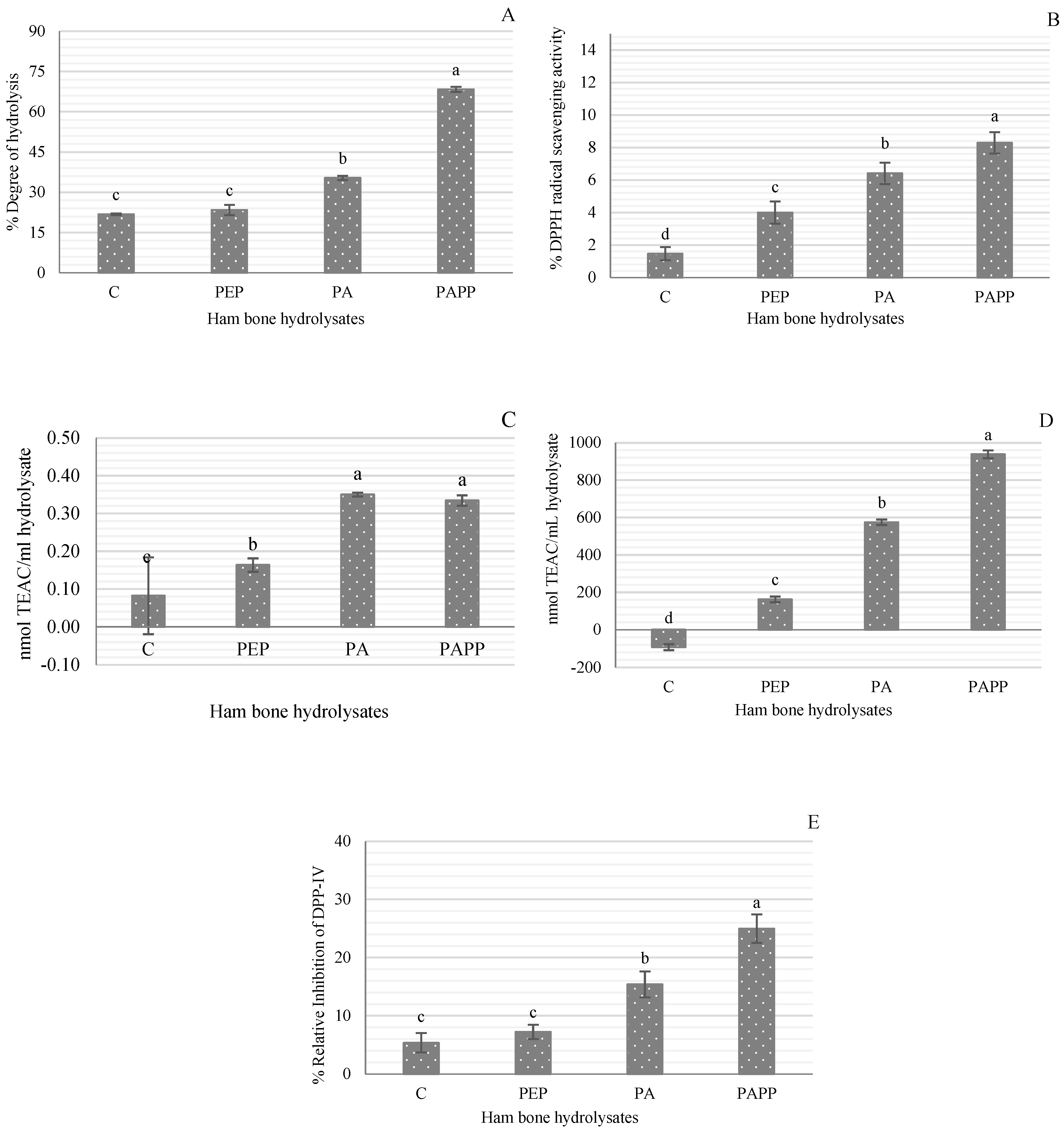
3.1. Degree of Hydrolysis (DH)
3.2. Antioxidant Activity
3.3. DPP-IV Inhibitory Activity
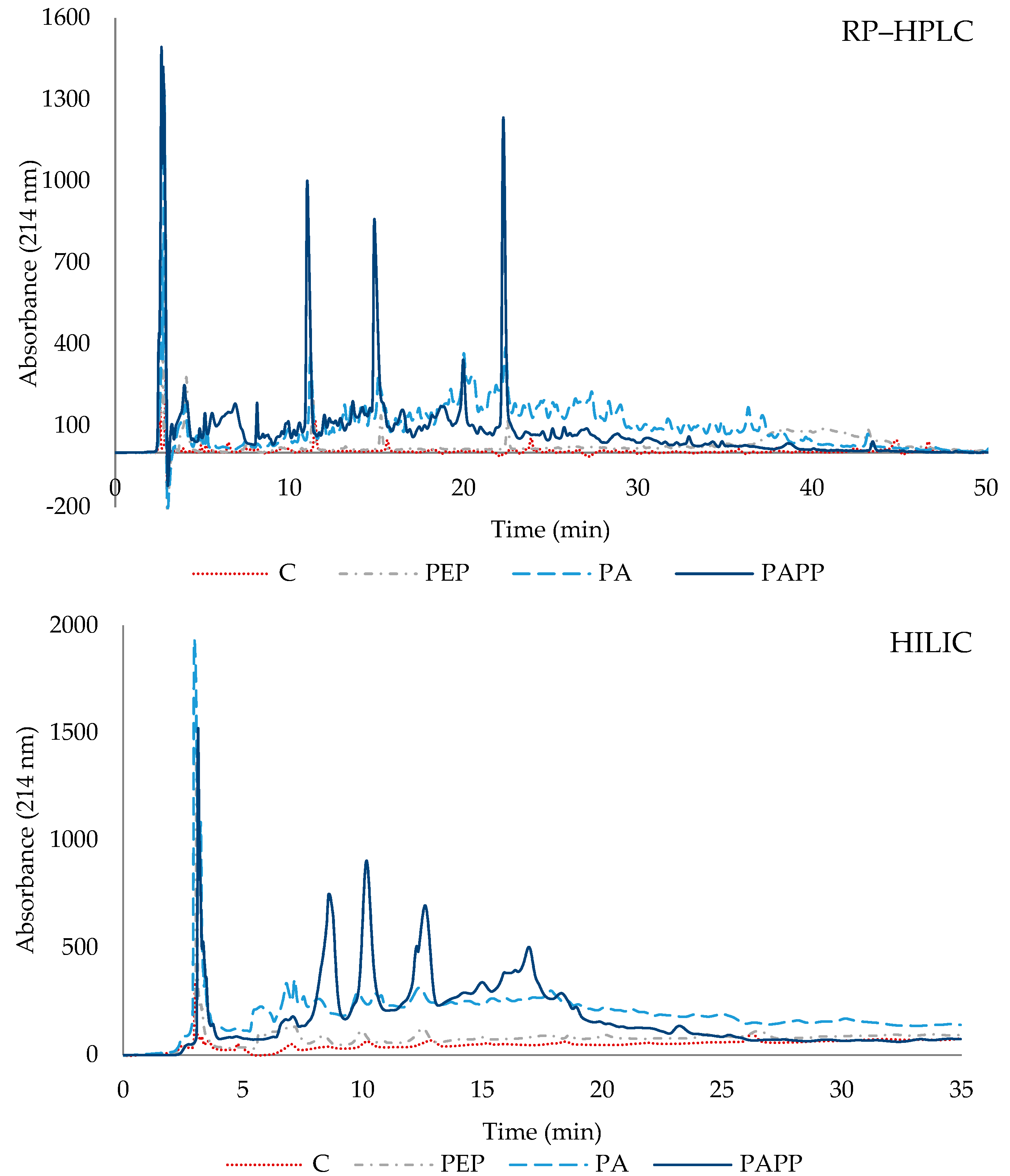
3.4. Characterization of the Generated Peptides Profile by HPLC and MALDI-MS Analysis
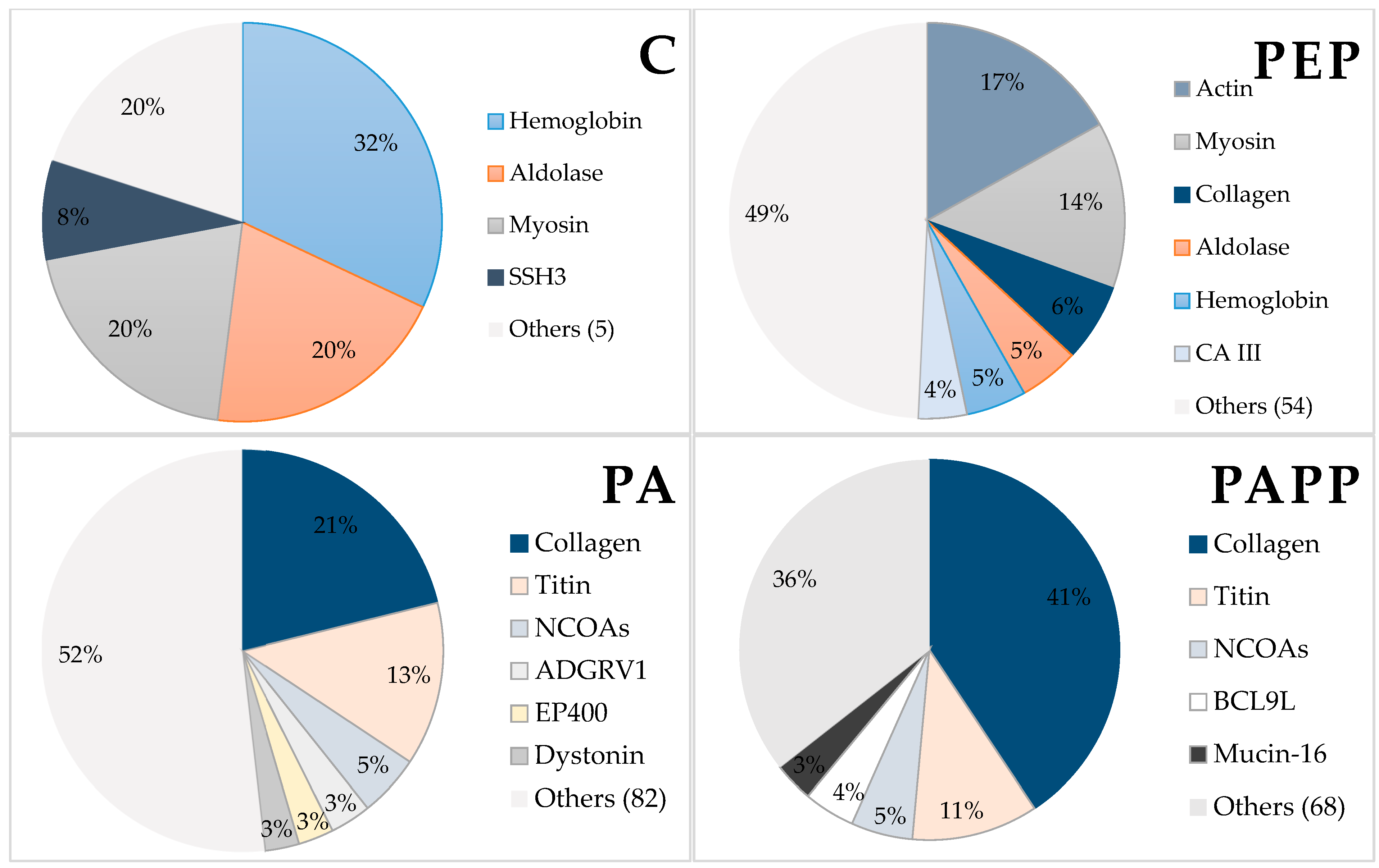
3.5. Identification of the Generated Peptides Using Mass Spectrometry in Tandem (LC-MS/MS)
3.6. Free Amino Acids
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Udenigwe, C.C.; Girgih, A.T.; Mohan, A.; Gong, M.; Malomo, S.A.; Aluko, R.E. Antihypertensive and bovine plasma oxidation-inhibitory activities of spent hen meat protein hydrolysates. J. Food Biochem. 2017, 41, e12378. [Google Scholar] [CrossRef]
- Neves, A.C.; Harnedy, P.A.; O’Keeffe, M.B.; Alashi, M.A.; Aluko, R.E.; FitzGerald, R.J. Peptide identification in a salmon gelatin hydrolysate with antihypertensive, dipeptidyl peptidase IV inhibitory and antioxidant activities. Food Res. Int. 2017, 100, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Reinoso, Z.; Cournoyer, A.; Thibodeau, J.; Said, L.B.; Fliss, I.; Bazinet, L.; Mikhaylin, S. Effect of pH on the Antimicrobial Activity and Peptide Population of Pepsin Hydrolysates Derived from Bovine and Porcine Hemoglobins. ACS Food Sci. Technol. 2021, 1, 1687–1701. [Google Scholar] [CrossRef]
- Wang, K.; Han, L.; Tan, Y.; Hong, H.; Luo, Y. Generation of novel antioxidant peptides from silver carp muscle hydrolysate: Gastrointestinal digestion stability and transepithelial absorption property. Food Chem. 2023, 403, 134136. [Google Scholar] [CrossRef]
- Gallego, M.; Mora, L.; Hayes, M.; Reig, M.; Toldrá, F. Effect of cooking and in vitro digestion on the antioxidant activity of dry-cured ham by-products. Food Res. Int. 2017, 97, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Z.; Zhang, H.; Zhang, M.; Yao, W.T.; Mao, X.Y.; Ren, F.Z. Antioxidant activity of hydrolysates and peptide fractions of porcine plasma albumin and globulin. J. Food Biochem. 2008, 32, 693–707. [Google Scholar] [CrossRef]
- He, L.; Gao, Y.; Wang, X.; Han, L.; Yu, Q.; Shi, H.; Song, R. Ultrasonication promotes extraction of antioxidant peptides from oxhide gelatin by modifying collagen molecule structure. Ultrason. Sonochem. 2021, 78, 105738. [Google Scholar] [CrossRef]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of novel DPP–IV inhibitory peptides from Atlantic salmon (Salmo salar) skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef]
- Carrera-Alvarado, G.; Toldrá, F.; Mora, L. DPP-IV Inhibitory Peptides GPF, IGL, and GGGW Obtained from Chicken Blood Hydrolysates. Int. J. Mol. Sci. 2022, 23, 14140. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Suwal, S.; Huang, J.Y.; Liceaga, A.M. Selective separation and characterisation of dual ACE and DPP-IV inhibitory peptides from rainbow trout (Oncorhynchus mykiss) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 1062–1073. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Parthsarathy, V.; McLaughlin, C.M.; O’Keeffe, M.B.; Allsopp, P.J.; McSorley, E.M.; O’Harte, F.P.M.; FitzGerald, R.J. Atlantic salmon (Salmo salar) co-product-derived protein hydrolysates: A source of antidiabetic peptides. Food Res. Int. 2018, 106, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Power, O.; Nongonierma, A.B.; Jakeman, P.; Fitzgerald, R.J. Food protein hydrolysates as a source of dipeptidyl peptidase IV inhibitory peptides for the management of type 2 diabetes. Proc. Nutr. Soc. 2014, 73, 34–46. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Features of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides from dietary proteins. J. Food Biochem. 2019, 43, e12451. [Google Scholar] [CrossRef] [PubMed]
- Heres, A.; Saldaña, C.; Toldrá, F.; Mora, L. Identification of dipeptides by MALDI-ToF mass spectrometry in long-processing Spanish dry-cured ham. Food Chem. Mol. Sci. 2021, 3, 100048. [Google Scholar] [CrossRef]
- Pagán, J.; Benítez, R.; Ibarz, A. Effect of enzymatic hydrolyzed protein from pig bones on some biological and functional properties. J. Food Sci. Technol. 2021, 58, 4626–4635. [Google Scholar] [CrossRef] [PubMed]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Bersuder, P.; Hole, M.; Smith, G. Antioxidants from a heated histidine-glucose model system. I: Investigation of the antioxidant role of histidine and isolation of antioxidants by high-performance liquid chromatography. J. Am. Oil Chem. Soc. 1998, 75, 181–187. [Google Scholar] [CrossRef]
- Huang, S.-J.; Tsai, S.-Y.; Mau, J.-L. Antioxidant properties of methanolic extracts from Agrocybe cylindracea. LWT Food Sci. Technol. 2006, 39, 379–387. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Aristoy, M.C.; Toldrá, F. Deproteinization techniques for HPLC amino acid analysis in fresh pork muscle and dry-cured ham. J. Agric. Food Chem. 1991, 39, 1792–1795. [Google Scholar] [CrossRef]
- Flores, M.; Aristoy, M.C.; Spanier, A.M.; Toldrá, F. Non-Volatile Components Effects on Quality of ‘Serrano’ Dry-cured Ham as Related to Processing Time. J. Food Sci. 1997, 62, 1235–1239. [Google Scholar] [CrossRef]
- Pagán, J.; Ibarz, A.; Falguera, V.; Benítez, R. Enzymatic hydrolysis kinetics and nitrogen recovery in the protein hydrolysate production from pig bones. J. Food Eng. 2013, 119, 655–659. [Google Scholar] [CrossRef]
- Yao, Y.; Wang, M.; Liu, Y.; Han, L.; Liu, X. Insights into the improvement of the enzymatic hydrolysis of bovine bone protein using lipase pretreatment. Food Chem. 2020, 302, 125199. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Gallego, M.; Reig, M.; Aristoy, M.C.; Mora, L. Recent Progress in Enzymatic Release of Peptides in Foods of Animal Origin and Assessment of Bioactivity. J. Agric. Food Chem. 2020, 68, 12842–12855. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, I.D.; Aluko, R.E. Structural and functional properties of food protein-derived antioxidant peptides. J. Food Biochem. 2019, 43, e12761. [Google Scholar] [CrossRef]
- Wang, K.; Han, L.; Hong, H.; Pan, J.; Liu, H.; Luo, Y. Purification and identification of novel antioxidant peptides from silver carp muscle hydrolysate after simulated gastrointestinal digestion and transepithelial transport. Food Chem. 2021, 342, 128275. [Google Scholar] [CrossRef]
- Cai, W.-W.; Hu, X.-M.; Wang, Y.-M.; Chi, C.-F.; Wang, B. Bioactive Peptides from Skipjack Tuna Cardiac Arterial Bulbs: Preparation, Identification, Antioxidant Activity, and Stability against Thermal, pH, and Simulated Gastrointestinal Digestion Treatments. Mar. Drugs 2022, 20, 626. [Google Scholar] [CrossRef]
- Cotabarren, J.; Rosso, A.M.; Tellechea, M.; García-Pardo, J.; Lorenzo, J.; Obregón, W.D.; Parisi, M.G. Adding value to the chia (Salvia hispanica L.) expeller: Production of bioactive peptides with antioxidant properties by enzymatic hydrolysis with Papain. Food Chem. 2019, 274, 848–856. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. Production of bioactive peptides using enzymatic hydrolysis and identification antioxidative peptides from patin (Pangasius sutchi) sarcoplasmic protein hydolysate. J. Funct. Foods 2014, 9, 280–289. [Google Scholar] [CrossRef]
- Gallego, M.; Mauri, L.; Aristoy, M.C.; Toldrá, F.; Mora, L. Antioxidant peptides profile in dry-cured ham as affected by gastrointestinal digestion. J. Funct. Foods 2020, 69, 103956. [Google Scholar] [CrossRef]
- Li, C.; Mora, L.; Toldrá, F. Characterization of antioxidant efficacy of peptide extracts as affected by peptide interactions during the ripening of Spanish dry-cured ham. Food Res. Int. 2021, 147, 110525. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; Escudero, E.; Fraser, P.D.; Aristoy, M.C.; Toldrá, F. Proteomic identification of antioxidant peptides from 400 to 2500 Da generated in Spanish dry-cured ham contained in a size-exclusion chromatography fraction. Food Res. Int. 2014, 56, 68–76. [Google Scholar] [CrossRef]
- Cheung, I.W.Y.; Li-Chan, E.C.Y. Enzymatic production of protein hydrolysates from steelhead (Oncorhynchus mykiss) skin gelatin as inhibitors of dipeptidyl-peptidase IV and angiotensin-I converting enzyme. J. Funct. Foods 2017, 28, 254–264. [Google Scholar] [CrossRef]
- Wang, R.; Edrington, T.C.; Storrs, S.B.; Crowley, K.S.; Ward, J.M.; Lee, T.C.; Liu, Z.L.; Li, B.; Glenn, K.C. Analyzing pepsin degradation assay conditions used for allergenicity assessments to ensure that pepsin susceptible and pepsin resistant dietary proteins are distinguishable. PLoS ONE 2017, 12, e0171926. [Google Scholar] [CrossRef] [PubMed]
- Tarnutzer, K.; Sankar, D.S.; Dengjel, J.; Ewald, C.Y. Collagen constitutes about 12% in females and 17% in males of the total protein in mice. Sci. Rep. 2023, 13–4490. [Google Scholar] [CrossRef]
- Toldrá, F.; Reig, M.; Aristoy, M.C.; Mora, L. Generation of bioactive peptides during food processing. Food Chem. 2018, 267, 395–404. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Num. of Unique Peptides |
|---|---|
| C | 25 |
| PEP | 550 |
| PA | 1087 |
| PAPP | 1124 |
| Free Amino Acids | Sample | |||
|---|---|---|---|---|
| C | PEP | PA | PAPP | |
| Aspartic acid (Asp) | 5.74 ± 0.09 c | 3.73 ± 0.00 d | 9.96 ± 0.22 b | 38.14 ± 0.18 a |
| Glutamic acid (Glu) | 3.49 ± 0.11 d | 4.74 ± 0.06 c | 7.57 ± 0.34 b | 54.67 ± 0.42 a |
| Serine (Ser) | 2.95 ± 0.07 d | 5.3 ± 0.03 c | 6.7 ± 0.09 b | 34.14 ± 0.28 a |
| Asparagine (Asn) | 23.37 ± 0.59 a | |||
| Glycine (Gly) | 2.31 ± 0.11 c | 3.86 ± 0.17 b c | 4.91 ± 0.19 b | 47.76 ± 1.66 a |
| Glutamine (Gln) | 26.73 ± 0.81 a | |||
| Histidine (His) | 1.82 ± 0.16 b | 9.43 ± 0.33 a | ||
| Threonine (Thr) | 1.68 ± 0.14 d | 3.58 ± 0.09 c | 5.63 ± 0.27 b | 30.11 ± 0.81 a |
| Alanine (Ala) | 1.64 ± 0.11 d | 3.61 ± 0.30 c | 5.67 ± 0.56 b | 54.06 ± 0.7 a |
| Arginine (Arg) | 1.67 ± 0.05 d | 5.3 ± 0.36 c | 7.76 ± 0.77 b | 57.21 ± 0.78 a |
| Proline (Pro) | 19.56 ± 0.65 a | |||
| Tyrosine (Tyr) | 1.88 ± 0.15 c | 3.68 ± 0.09 b | 4.62 ± 0.08 b | 24.95 ± 1.22 a |
| Valine (Val) | 2.34 ± 0.17 b | 4.14 ± 0.13 b | 37.54 ± 1.86 a | |
| Methionine (Met) | 2.84 ± 0.48 b | 13.31 ± 0.52 a | ||
| Isoleucine (Ile) | 1.45 ± 0.10 c | 2.57 ± 0.12 b | 23.53 ± 0.54 a | |
| Leucine (Leu) | 1.83 ± 0.16 d | 4.66 ± 0.05 c | 7.29 ± 0.21 b | 50.44 ± 0.55 a |
| Phenylalanine (Phe) | 2.58 ± 0.06 c | 4.11 ± 0.49 b | 24.51 ± 0.28 a | |
| Tryptophan (Trp) | 6.97 ± 0.1 a | |||
| Lysine (Lys) | 3.74 ± 0.07 d | 9.79 ± 0.07 c | 12.53 ± 0.88 b | 92.06 ± 0.64 a |
| Total (mg/100 mL hydrolysate) | 26.37 ± 0.45 d | 54.62 ± 0.07 c | 88.12 ± 1.36 b | 668.46 ± 5.64 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrera-Alvarado, G.; Toldrá, F.; Mora, L. Potential of Dry-Cured Ham Bones as a Sustainable Source to Obtain Antioxidant and DPP-IV Inhibitory Extracts. Antioxidants 2023, 12, 1151. https://doi.org/10.3390/antiox12061151
Carrera-Alvarado G, Toldrá F, Mora L. Potential of Dry-Cured Ham Bones as a Sustainable Source to Obtain Antioxidant and DPP-IV Inhibitory Extracts. Antioxidants. 2023; 12(6):1151. https://doi.org/10.3390/antiox12061151
Chicago/Turabian StyleCarrera-Alvarado, Gisela, Fidel Toldrá, and Leticia Mora. 2023. "Potential of Dry-Cured Ham Bones as a Sustainable Source to Obtain Antioxidant and DPP-IV Inhibitory Extracts" Antioxidants 12, no. 6: 1151. https://doi.org/10.3390/antiox12061151