
Development and Recovery of Liver Injury in Piglets by Incremental Injection of LPS

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Sample Collection
2.3. Serum Biochemical Indexes
2.4. Histopathological Evaluation
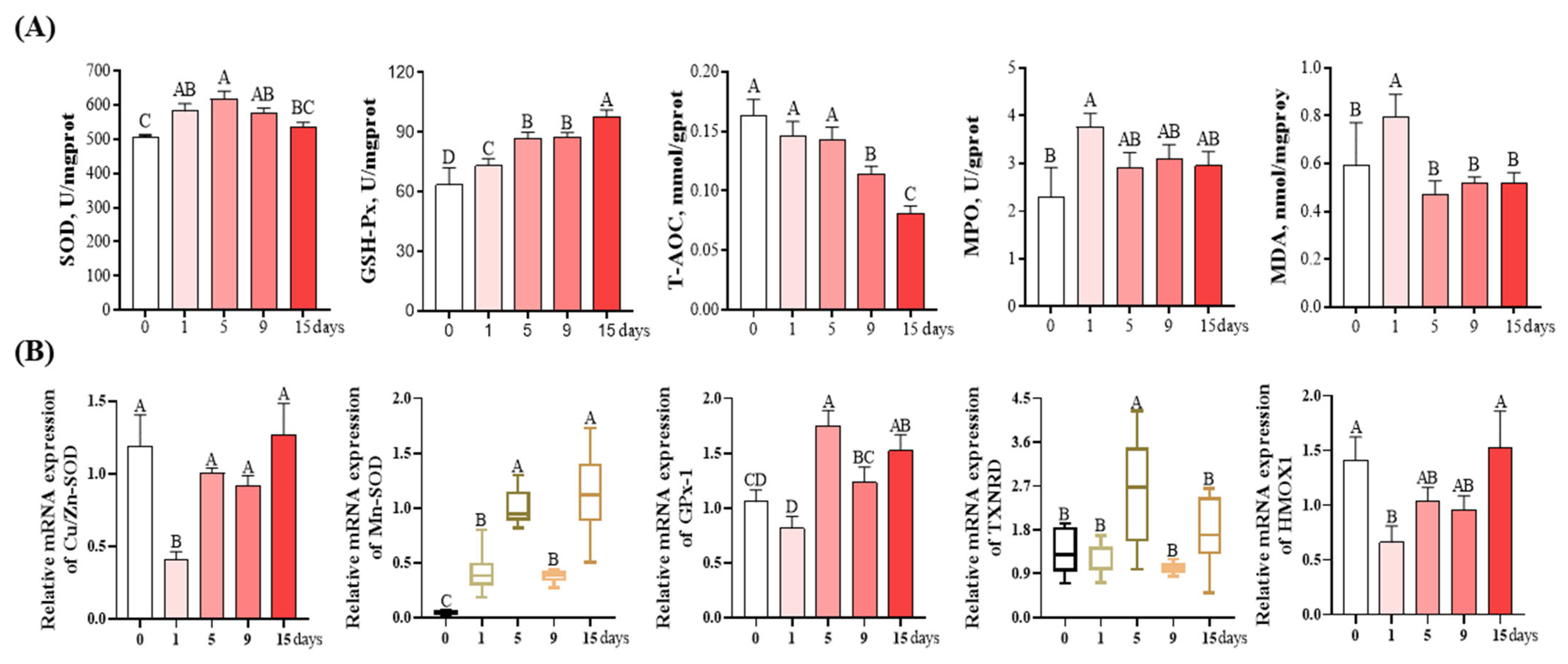
2.5. Redox Parameters, ROS, and Mitochondrial Membrane Potential Detection
2.6. Reverse Transcription and Quantitative Real-Time PCR
2.7. Immunofluorescence
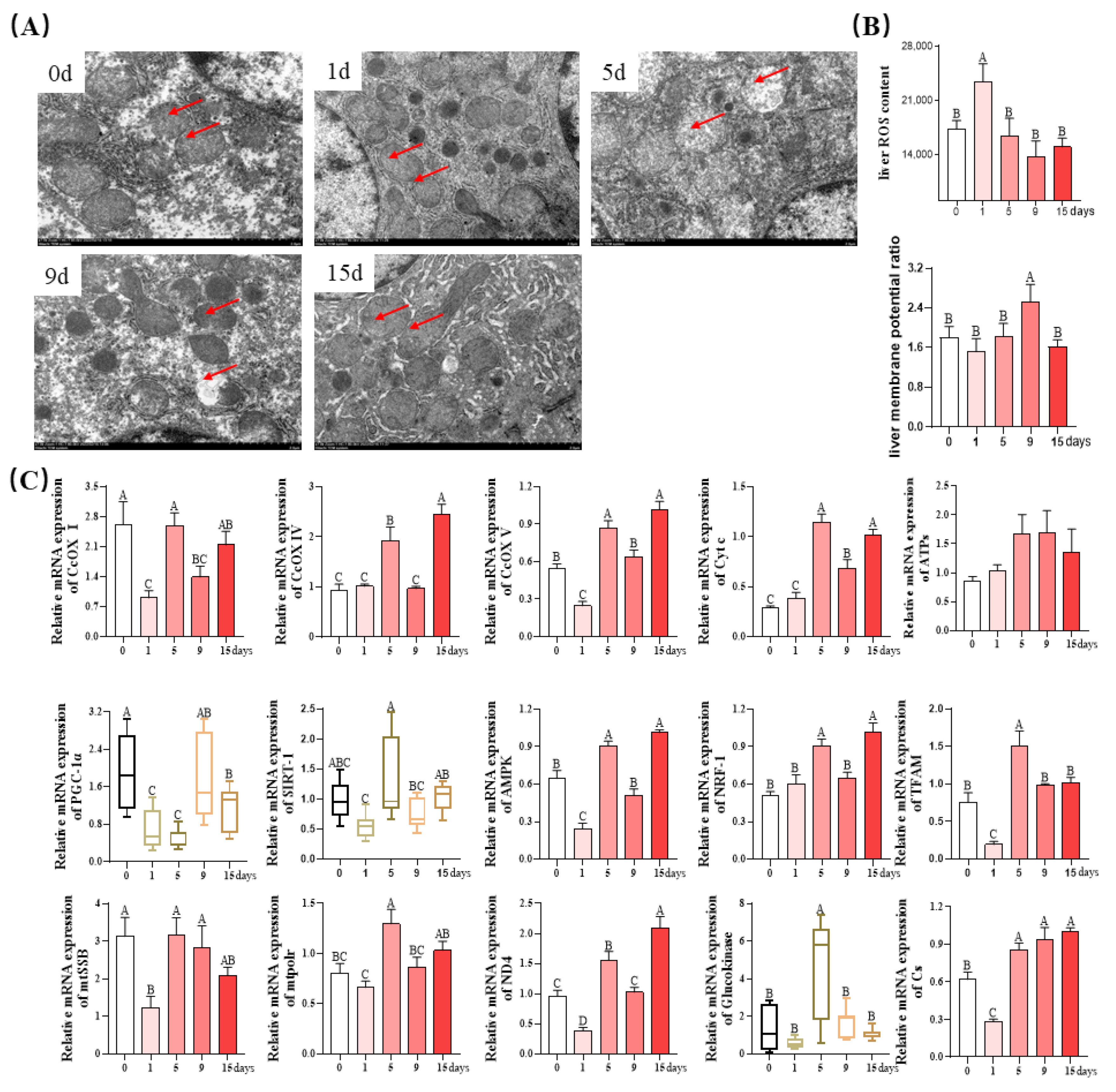
2.8. Mitochondrial Ultrastructure
2.9. Statistical Analysis
3. Results
3.1. Serum Biochemical Indexes
3.2. Histopathological Observation of Livers
3.3. Liver Inflammation
3.4. Antioxidant and Oxidative Damage-Related Parameters of livers
3.5. Mitochondrial Function of Livers
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, I.K.; Kye, Y.C.; Kim, G.; Kim, H.W.; Gu, M.J.; Umboh, J.; Maaruf, K.; Kim, S.W.; Yun, C.H. Stress, Nutrition, and Intestinal Immune Responses in Pigs—A Review. Asian-Australas. J. Anim. Sci. 2016, 29, 1075–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Chen, Y.; Li, Y.; Jia, P.; Ji, S.; Chen, Y.; Wang, T. Protective effects of pterostilbene against hepatic damage, redox imbalance, mitochondrial dysfunction, and endoplasmic reticulum stress in weanling piglets. J. Anim. Sci. 2020, 98, skaa328. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Bortoluzzi, E.M.; Goering, M.J.; Ochoa, S.J.; Holliday, A.J.; Mumm, J.M.; Nelson, C.E.; Wu, H.; Mote, B.E.; Psota, E.T.; Schmidt, T.B.; et al. Evaluation of Precision Livestock Technology and Human Scoring of Nursery Pigs in a Controlled Immune Challenge Experiment. Animals 2023, 13, 246. [Google Scholar] [CrossRef]
- Yu, J.; Zheng, C.; Zheng, J.; Duan, G.; Guo, Q.; Zhang, P.; Wan, M.; Duan, Y. Development of Intestinal Injury and Restoration of Weaned Piglets under Chronic Immune Stress. Antioxidants 2022, 11, 2215. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, S.; Zhong, J.; Bian, Y.; Fan, Y.; Chen, Q.; Liu, P.; Liu, Z. Rhein ameliorates lipopolysaccharide-induced intestinal barrier injury via modulation of Nrf2 and MAPKs. Life Sci. 2019, 216, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Y.; Shi, H.; Wang, X.; Zhu, H.; Pi, D.; Leng, W.; Li, S. Aspartate attenuates intestinal injury and inhibits TLR4 and NODs/NF-kappaB and p38 signaling in weaned pigs after LPS challenge. Eur. J. Nutr. 2017, 56, 1433–1443. [Google Scholar] [CrossRef]
- Wang, R.; Tang, R.; Li, B.; Ma, X.; Schnabl, B.; Tilg, H. Gut microbiome, liver immunology, and liver diseases. Cell. Mol. Immunol. 2021, 18, 4–17. [Google Scholar] [CrossRef]
- Chen, F.; Liu, Y.; Zhu, H.; Hong, Y.; Wu, Z.; Hou, Y.; Li, Q.; Ding, B.; Yi, D.; Chen, H. Fish oil attenuates liver injury caused by LPS in weaned pigs associated with inhibition of TLR4 and nucleotide-binding oligomerization domain protein signaling pathways. Innate Immun. 2013, 19, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Kang, P.; Liu, Y.; Zhu, H.; Zhang, J.; Shi, H.; Li, S.; Pi, D.; Leng, W.; Wang, X.; Wu, H.; et al. The effect of dietary asparagine supplementation on energy metabolism in liver of weaning pigs when challenged with lipopolysaccharide. Asian-Australas. J. Anim. Sci. 2018, 31, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Leng, W.; Liu, Y.; Shi, H.; Li, S.; Zhu, H.; Pi, D.; Hou, Y.; Gong, J. Aspartate alleviates liver injury and regulates mRNA expressions of TLR4 and NOD signaling-related genes in weaned pigs after lipopolysaccharide challenge. J. Nutr. Biochem. 2014, 25, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, X.L.; Jiang, X.M.; Chen, L.; Hong, L.; Zhuo, Y.; Lin, Y.; Fang, Z.F.; Che, L.Q.; Feng, B.; et al. Effects of dietary supplementation with exogenous catalase on growth performance, oxidative stress, and hepatic apoptosis in weaned piglets challenged with lipopolysaccharide. J. Anim. Sci. 2020, 98, skaa067. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Y.; Cao, K.X.; Yang, R.G.; Xu, B.Y.; Zhang, W.P.; Batonon-Alavo, D.I.; Zhang, S.J.; Sun, L.H. Increased Ingestion of Hydroxy-Methionine by Both Sows and Piglets Improves the Ability of the Progeny to Counteract LPS-Induced Hepatic and Splenic Injury with Potential Regulation of TLR4 and NOD Signaling. Antioxidants 2022, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hou, Y.; Yi, D.; Li, Y.; Ding, B.; Zhu, H.; Liu, J.; Xiao, H.; Wu, G. Dietary supplementation with glutamate precursor alpha-ketoglutarate attenuates lipopolysaccharide-induced liver injury in young pigs. Amino Acids 2015, 47, 1309–1318. [Google Scholar] [CrossRef]
- Yi, D.; Hou, Y.; Wang, L.; Ding, B.; Yang, Z.; Li, J.; Long, M.; Liu, Y.; Wu, G. Dietary N-acetylcysteine supplementation alleviates liver injury in lipopolysaccharide-challenged piglets. Br. J. Nutr. 2014, 111, 46–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, D.; Wu, T.; Yi, D.; Wang, L.; Li, P.; Zhang, J.; Hou, Y.; Wu, G. Dietary Supplementation with Lactobacillus casei Alleviates Lipopolysaccharide-Induced Liver Injury in a Porcine Model. Int. J. Mol. Sci. 2017, 18, 2535. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.Y.; Lee, S.H.; Han, E.; Hwang, J.S.; Han, S.; Kim, M.K.; Jang, B.K. Evogliptin Directly Inhibits Inflammatory and Fibrotic Signaling in Isolated Liver Cells. Int. J. Mol. Sci. 2022, 23, 11636. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Du, S.Y.; Kong, Q.Y.; Zhang, X.J.; Jiang, S.R.; Cao, X.F.; Li, Y.H.; Li, C.F.; Chen, H.Q.; Ding, Z.N.; et al. HSPA12A attenuates lipopolysaccharide-induced liver injury through inhibiting caspase-11-mediated hepatocyte pyroptosis via PGC-1 alpha-dependent acyloxyacyl hydrolase expression. Cell Death Differ. 2020, 27, 2651–2667. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Gordillo, K.; Shah, R.; Muriel, P. Oxidative Stress and Inflammation in Hepatic Diseases: Current and Future Therapy. Oxidative Med. Cell. Longev. 2017, 2017, 3140673. [Google Scholar] [CrossRef]
- Dare, A.J.; Phillips, A.R.; Hickey, A.J.; Mittal, A.; Loveday, B.; Thompson, N.; Windsor, J.A. A systematic review of experimental treatments for mitochondrial dysfunction in sepsis and multiple organ dysfunction syndrome. Free Radic. Biol. Med. 2009, 47, 1517–1525. [Google Scholar] [CrossRef]
- David, J.A.; Rifkin, W.J.; Rabbani, P.S.; Ceradini, D.J. The Nrf2/Keap1/ARE Pathway and Oxidative Stress as a Therapeutic Target in Type II Diabetes Mellitus. J. Diabetes Res. 2017, 2017, 4826724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doise, J.M.; Aho, L.S.; Quenot, J.P.; Guilland, J.C.; Zeller, M.; Vergely, C.; Aube, H.; Blettery, B.; Rochette, L. Plasma antioxidant status in septic critically ill patients: A decrease over time. Fundam. Clin. Pharmacol. 2008, 22, 203–209. [Google Scholar] [CrossRef]
- Huang, F.F.; Yang, Y.; Wang, L.M.; Wang, H.; Li, P.; Xiao, K.; Xu, X.; Liu, J.S.; Liu, Y.L.; Zhu, H.L. Holly polyphenols attenuate liver injury, suppression inflammation and oxidative stress in lipopolysaccharide-challenged weaned pigs. Food Agric. Immunol. 2022, 33, 35–46. [Google Scholar] [CrossRef]
- Svistunenko, D.A.; Davies, N.; Brealey, D.; Singer, M.; Cooper, C.E. Mitochondrial dysfunction in patients with severe sepsis: An EPR interrogation of individual respiratory chain components. Biochim. Biophys. Acta 2006, 1757, 262–272. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Rui, D.; Yan, Y.; Xu, S.; Niu, Q.; Feng, G.; Wang, Y.; Li, S.; Jing, M. Oxidative Damage Induced by Arsenic in Mice or Rats: A Systematic Review and Meta-Analysis. Biol. Trace Elem. Res. 2017, 176, 154–175. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Y.L.; Che, Z.Q.; Zhu, H.L.; Meng, G.Q.; Hou, Y.Q.; Ding, B.Y.; Yin, Y.L.; Chen, F. Dietary L-arginine supplementation alleviates liver injury caused by Escherichia coli LPS in weaned pigs. Innate Immun. 2012, 18, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zheng, C.; Zhong, Y.; Song, B.; Yan, Z.; Kong, X.; Deng, J.; Li, F.; Yin, Y. Beta-hydroxy beta-methyl butyrate decreases muscle protein degradation via increased Akt/FoxO3a signaling and mitochondrial biogenesis in weanling piglets after lipopolysaccharide challenge. Food Funct. 2019, 10, 5152–5165. [Google Scholar] [CrossRef]
- Escribano, D.; Campos, P.H.R.F.; Gutierrez, A.M.; Le Floc’h, N.; Ceron, J.J.; Merlot, E. Effect of repeated administration of lipopolysaccharide on inflammatory and stress markers in saliva of growing pigs. Vet. J. 2014, 200, 393–397. [Google Scholar] [CrossRef]
- Rakhshandeh, A.; de Lange, C.F.M. Evaluation of chronic immune system stimulation models in growing pigs. Animal 2012, 6, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Hamza, A.A.; Mohamed, M.G.; Lashin, F.M.; Amin, A. Dandelion prevents liver fibrosis, inflammatory response, and oxidative stress in rats. J. Basic Appl. Zool. 2020, 81, 43. [Google Scholar] [CrossRef]
- Zeng, M.; Zhang, B.; Li, B.; Kan, Y.; Wang, S.; Feng, W.; Zheng, X. Adenosine Attenuates LPS-Induced Cardiac Dysfunction by Inhibition of Mitochondrial Function via the ER Pathway. Evid. Based Complement. Altern. Med. 2019, 2019, 1832025. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Deng, Z.Z.; Geng, L.H.; Wang, J.; Zhang, Q.B. In vitro evaluation of the neuroprotective effect of oligo-porphyran from Porphyra yezoensis in PC12 cells. J. Appl. Phycol. 2019, 31, 2559–2571. [Google Scholar] [CrossRef]
- Duan, Y.H.; Li, F.N.; Li, L.L.; Fan, J.X.; Sun, X.M.; Yin, Y.L. n-6:n-3 PUFA ratio is involved in regulating lipid metabolism and inflammation in pigs. Br. J. Nutr. 2014, 111, 445–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra-Frutos, I. Preliminary tests when comparing means. Comput. Stat. 2016, 31, 1607–1631. [Google Scholar] [CrossRef]
- Hamesch, K.; Borkham-Kamphorst, E.; Strnad, P.; Weiskirchen, R. Lipopolysaccharide-induced inflammatory liver injury in mice. Lab. Anim. 2015, 49, 37–46. [Google Scholar] [CrossRef]
- Kang, P.; Liu, Y.; Zhu, H.; Li, S.; Shi, H.; Chen, F.; Leng, W.; Pi, D.; Hou, Y.; Yi, D. The effect of aspartate on the energy metabolism in the liver of weanling pigs challenged with lipopolysaccharide. Eur. J. Nutr. 2015, 54, 581–588. [Google Scholar] [CrossRef]
- Kwo, P.Y.; Cohen, S.M.; Lim, J.K. ACG Clinical Guideline: Evaluation of Abnormal Liver Chemistries. Am. J. Gastroenterol. 2017, 112, 18–35. [Google Scholar] [CrossRef]
- Wan, J.Y.; Gong, X.; Zhang, L.; Li, H.Z.; Zhou, Y.F.; Zhou, Q.X. Protective effect of baicalin against Lipopolysaccharide/D-galactosamine-induced liver injury in mice by up-regulation of Heme oxygenase-1. Eur. J. Pharmacol. 2008, 587, 302–308. [Google Scholar] [CrossRef]
- Tan, L.; Meng, Y.; Zeng, T.; Wang, Q.; Long, T.; Wu, S.; Guan, X.; Fu, H.; Zheng, W.; Tian, Y.; et al. Clinical diagnostic significance of prealbumin, cholinesterase and retinol binding protein in liver cirrhosis combined with encephalopathy. Br. J. Biomed. Sci. 2019, 76, 24–28. [Google Scholar] [CrossRef]
- Kwan, S.Y.; Jiao, J.J.; Qi, J.; Wang, Y.; Wei, P.; McCormick, J.B.; Fisher-Hoch, S.P.; Beretta, L. Bile Acid Changes Associated with Liver Fibrosis and Steatosis in the Mexican-American Population of South Texas. Hepatol. Commun. 2020, 4, 555–568. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Guo, J.; Li, X.; Wang, Y.; Wang, D.; Xiao, K.; Zhu, H.; Wang, X.; Hu, C.A.; Zhang, G.; et al. Necroptosis Underlies Hepatic Damage in a Piglet Model of Lipopolysaccharide-Induced Sepsis. Front. Immunol. 2021, 12, 633830. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F.; Luedde, T.; Trautwein, C. Inflammatory pathways in liver homeostasis and liver injury. Clin. Rev. Allergy Immunol. 2009, 36, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yang, L.; Dong, C.; Li, L. The class D scavenger receptor CD68 contributes to mouse chronic liver injury. Immunol. Res. 2018, 66, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Zen, Y.; Yeh, M.M. Hepatotoxicity of immune checkpoint inhibitors: A histology study of seven cases in comparison with autoimmune hepatitis and idiosyncratic drug-induced liver injury. Mod. Pathol. 2018, 31, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Knolle, P.A. Involvement of the liver in the induction of CD8 T cell tolerance towards oral antigen. Z. Gastroenterol. 2006, 44, 51–56. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion-from mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef] [Green Version]
- Klune, J.R.; Tsung, A. Molecular Biology of Liver Ischemia/Reperfusion Injury: Established Mechanisms and Recent Advancements. Surg. Clin. N. Am. 2010, 90, 665–677. [Google Scholar] [CrossRef]
- Recknagel, P.; Gonnert, F.A.; Westermann, M.; Lambeck, S.; Lupp, A.; Rudiger, A.; Dyson, A.; Carre, J.E.; Kortgen, A.; Krafft, C.; et al. Liver Dysfunction and Phosphatidylinositol-3-Kinase Signalling in Early Sepsis: Experimental Studies in Rodent Models of Peritonitis. PLoS Med. 2012, 9, e1001338. [Google Scholar] [CrossRef]
- Zhang, F.; Qiu, X.M.; Liu, Y.; Wang, J.; Li, X.P.; Wang, X.L. Expression analysis of three immune genes Interferongamma, Mx and Interferon regulatory factor-1 of Japanese flounder (Paralichthys olivaceus). Braz. Arch. Biol. Technol. 2017, 60, e17160243. [Google Scholar] [CrossRef] [Green Version]
- Maki, T. Pathogenesis of calcium bilirubinate gallstone: Role of E. coli, beta-glucuronidase and coagulation by inorganic ions, polyelectrolytes and agitation. Ann. Surg. 1966, 164, 90–100. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. TLR signaling. Cell Death Differ. 2006, 13, 816–825. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; He, Q.; Li, S.; Liu, T.; Zhang, J. Hydrogen-Rich Water Mitigates LPS-Induced Chronic Intestinal Inflammatory Response in Rats via Nrf-2 and NF-kappaB Signaling Pathways. Vet. Sci. 2022, 9, 621. [Google Scholar] [CrossRef]
- Bailey, S.M.; Cunningham, C.C. Contribution of mitochondria to oxidative stress associated with alcoholic liver disease. Free Radic. Biol. Med. 2002, 32, 11–16. [Google Scholar] [CrossRef]
- Cecerska-Heryc, E.; Surowska, O.; Heryc, R.; Serwin, N.; Napiontek-Balinska, S.; Dolegowska, B. Are antioxidant enzymes essential markers in the diagnosis and monitoring of cancer patients—A review. Clin. Biochem. 2021, 93, 1–8. [Google Scholar] [CrossRef]
- Saita, E.; Kondo, K.; Momiyama, Y. Anti-Inflammatory Diet for Atherosclerosis and Coronary Artery Disease: Antioxidant Foods. Clin. Med. Insights Cardiol. 2015, 8, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Wu, M.M.; Xiao, H.; Ren, W.K.; Duan, J.L.; Yang, G.; Li, T.J.; Yin, Y.L. Development of an antioxidant system after early weaning in piglets. J. Anim. Sci. 2014, 92, 612–619. [Google Scholar] [CrossRef]
- Davies, M.J.; Hawkins, C.L. The Role of Myeloperoxidase in Biomolecule Modification, Chronic Inflammation, and Disease. Antioxid. Redox Signal. 2020, 32, 957–981. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Buettner, G.R. Superoxide Dismutase in Redox Biology: The Roles of Superoxide and Hydrogen Peroxide. Anti-Cancer Agents Med. Chem. 2011, 11, 341–346. [Google Scholar] [CrossRef]
- Tu, H.K.; Pan, K.F.; Zhang, Y.; Li, W.Q.; Zhang, L.; Ma, J.L.; Li, J.Y.; You, W.C. Manganese Superoxide Dismutase Polymorphism and Risk of Gastric Lesions, and Its Effects on Chemoprevention in a Chinese Population. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1089–1097. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Kou, Y.; Gao, Y.; Yang, P.; Liu, S.; Zhang, F.; Li, M. Eldecalcitol inhibits the progression of oral cancer by suppressing the expression of GPx-1. Oral. Dis. 2023, 29, 615–627. [Google Scholar] [CrossRef]
- Jaeschke, H.; McGill, M.R.; Ramachandran, A. Oxidant stress, mitochondria, and cell death mechanisms in drug-induced liver injury: Lessons learned from acetaminophen hepatotoxicity. Drug Metab. Rev. 2012, 44, 88–106. [Google Scholar] [CrossRef] [Green Version]
- Niknahad, H.; Heidari, R.; Alzuhairi, A.M.; Najibi, A. Mitochondrial dysfunction as a mechanism for pioglitazone-induced injury toward HepG2 cell line. Pharm. Sci. 2014, 20, 169–174. [Google Scholar]
- Ahmadian, E.; Babaei, H.; Mohajjel Nayebi, A.; Eftekhari, A.; Eghbal, M.A. Mechanistic Approach for Toxic Effects of Bupropion in Primary Rat Hepatocytes. Drug. Res. 2017, 67, 217–222. [Google Scholar] [CrossRef]
- Labbe, G.; Pessayre, D.; Fromenty, B. Drug-induced liver injury through mitochondrial dysfunction: Mechanisms and detection during preclinical safety studies. Fundam. Clin. Pharmacol. 2008, 22, 335–353. [Google Scholar] [CrossRef]
- Brookes, P.S.; Yoon, Y.; Robotham, J.L.; Anders, M.W.; Sheu, S.S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am. J. Physiol. Cell Physiol. 2004, 287, C817–C833. [Google Scholar] [CrossRef]
- Mantzarlis, K.; Tsolaki, V.; Zakynthinos, E. Role of Oxidative Stress and Mitochondrial Dysfunction in Sepsis and Potential Therapies. Oxidative Med. Cell. Longev. 2017, 2017, 5985209. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, oxidative stress and cell death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef] [Green Version]
- Foster, K.A.; Galeffi, F.; Gerich, F.J.; Turner, D.A.; Muller, M. Optical and pharmacological tools to investigate the role of mitochondria during oxidative stress and neurodegeneration. Prog. Neurobiol. 2006, 79, 136–171. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.; Kang, Y.; Ji, X.; Li, S.; Li, Y.; Zhang, G.; Cui, H.; Shi, G. Testosterone Upregulates the Expression of Mitochondrial ND1 and ND4 and Alleviates the Oxidative Damage to the Nigrostriatal Dopaminergic System in Orchiectomized Rats. Oxid. Med. Cell. Longev. 2017, 2017, 1202459. [Google Scholar] [CrossRef] [Green Version]
- Halling, J.F.; Pilegaard, H. PGC-1 alpha-mediated regulation of mitochondrial function and physiological implications. Appl. Physiol. Nutr. Metab. 2020, 45, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Suliman, H.B.; Carraway, M.S.; Welty-Wolf, K.E.; Whorton, A.R.; Piantadosi, C.A. Lipopolysaccharide stimulates mitochondrial biogenesis via activation of nuclear respiratory factor-1. J. Biol. Chem. 2003, 278, 41510–41518. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequence (5′ to 3′) | Size (bp) | Gene Bank No. |
|---|---|---|---|
| Interleukin-6, IL-6 | F: AGCCCACCAGGAACGAAAGA | 119 | NM_214399.1 |
| R: AGCCATCACCAGAAGCAGCC | |||
| Tumor necrosis factor-α, TNF-α | F: ATGGATGGGTGGATGAGAAA | 129 | NM_214022.1 |
| R: TGGAAACTGTTGGGGAGAAG | |||
| Interleukin-12, IL-12 | F: TTTCAGACCCGACGAACTCT | 160 | NM_214013 |
| R: CATTGGGGTACCAGTCCAAC | |||
| Interleukin-1β, IL-1β | F: CAAAGGCCGCCAAGATATAA | 147 | NM_214055 |
| R: GAAATTCAGGCAGCAACAT | |||
| Nuclear factor kappa-B, NF-κB | F: GACAACATCTCCTTGGCGGG | 146 | NM_001048232 |
| R: TCTGCTCCTGCTGCTTTGAGG | |||
| Toll-like receptor 4, TLR4 | F: TCAGTTCTCACCTTCCTCCTG | 166 | NM_001293316.1 |
| R: GTTCATTCCTCACCCAGTCTTC | |||
| Myeloid differentiation factor 88, MyD88 | F: GATGGTAGCGGTTGTCTCTGAT | 148 | NM_001099923.1 |
| R: GATGCTGGGGAACTCTTTCTTC | |||
| TNF receptor associated factor 6, TRAF6 | F: CAAGAGAATACCCAGTCGCACA | 122 | XM_013990069.2 |
| R: ATCCGAGACAAAGGGGAAGAA | |||
| Copper and zinc superoxide dismutase, Cu/Zn-SOD | F: CAGGTCCTCACTTCAATCC | 255 | NM_001190422 |
| R: CCAAACGACTTCCASCAT | |||
| Manganese-containing superoxide dismutase, Mn-SOD | F: GGACAAATCTGAGCCCTAACG | 159 | NM_214127 |
| R: CCTTGTTGAAACCGAGCC | |||
| Glutathione peroxidase 1, GPx-1 | F: TGGGGAGATCCTGAATTG | 183 | NM_214201 |
| R: GATAAACTTGGGGTCGGT | |||
| Thioredoxin reductase 1, TXNRD | F: GTGCTGAGGAGCTTCCCGAGATGT | 118 | NM_214154.3 |
| R: TCCAGGACCATGACCCGCTTGTTAA | |||
| Heme oxygenase 1, HMOX1 | F: CGCTCCCGAATGAACACTCT | 148 | NM_001004027.1 |
| R: GCGAGGGTCTCTGGTCCTTA | |||
| Peroxisome proliferator-activated receptor g coactivator-1α, PGC-1α | F: CCCGAAACAGTAGCAGAGACAAG | 111 | NM 213963 |
| R: CTGGGGTCAGAGGAAGAGATAAAG | |||
| Nuclear respiratory factor-1, NRF-1 | F: GCCAGTGAGATGAAGAGAAACG | 166 | AK237171.1 |
| R; CTACAGCAGGGACCAAAGTTCAC | |||
| Transcription factor a mitochondrial, TFAM | F: GGTCCATCACAGGTAAAGCTGAA | 167 | AY923074.1 |
| R: ATAAGATCGTTTCGCCCAACTTC | |||
| Silent information regulator transcript 1, Sirt-1 | F: TGACTGTGAAGCTGTACGAGGAG | 143 | EU030283.2 |
| R: TGGCTCTATGAAACTGCTCTGG | |||
| Mitochondrial single-strand DNA-binding protein, mtSSB | F: CTTTGAGGTAGTGCTGTGTCG | 143 | AK352341.1 |
| R: CTCACCCCTGACGATGAAGAC | |||
| Mitochondrial RNA polymerase, mtpolr | F: CTTTGAGGTTTTCCAGCAGCAG | 119 | XM 001927064.1 |
| R: GCTCCCAGTTTTGGTTGACAG | |||
| NADH dehydrogenase subunit 4, ND4 | F: TTATTGGTGCCGGAGGTACTG | 112 | NM 001097468 |
| R: CCCAGTTTATTCCAGGGTTCTG | |||
| Glucokinase | F: CTTTTCCCTCCCACACTGCTAT | 119 | AK233298.1 |
| R: GACTCCTCTTCCTGAGACCCTCT | |||
| Citrate synthase, Cs | F: CCTTTCAGACCCCTACTTGTCCT | 127 | M21197.1 |
| R: CACATCTTTGCCGACTTCCTTC | |||
| AMP-activated protein kinase, AMPK | F: ACCAGGACCCTTTGGCAGTT | 100 | NM_001167633.1 |
| R: GAATCAGGTGGGCTTGTTGC | |||
| Cytochrome c oxidase I, CcOX Ⅰ | F: ATTATCCTGACGCATACACAGCA | 127 | AJ950517.1 |
| R: GCAGATACTTCTCGTTTTGATGC | |||
| Cytochrome c oxidase Ⅳ, CcOX Ⅳ | F: CCAAGTGGGACTACGACAAGAAC | 131 | AK233334.1 |
| R: CCTGCTCGTTTATTAGCACTGG | |||
| Cytochrome c oxidase Ⅳ, CcOX V | F: ATCTGGAGGTGGTGTTCCTACTG | 160 | AY786556.1 |
| R: GTTGGTGATGGAGGGGACTAAA | |||
| Cytochrome c, Cyt c | F: TAGAAAAGGGAGGCAAACACAAG | 154 | NM 001129970.1 |
| R: GGATTCTCCAGGTACTCCATCAG | |||
| ATP synthase, ATPs | F: TGTCCTCCTCCCTATCACACATT | 116 | AK230503 |
| R: TAGTGGTTATGACGTTGGCTTGA | |||
| β-actin | F: TCTTTTCCAGCCTTCCTTCTTG | 100 | NM_007393 |
| R: GAGGTCTTTACGGATGTCAACG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, G.; Huang, P.; Zheng, C.; Zheng, J.; Yu, J.; Zhang, P.; Wan, M.; Li, F.; Guo, Q.; Yin, Y.; et al. Development and Recovery of Liver Injury in Piglets by Incremental Injection of LPS. Antioxidants 2023, 12, 1143. https://doi.org/10.3390/antiox12061143
Duan G, Huang P, Zheng C, Zheng J, Yu J, Zhang P, Wan M, Li F, Guo Q, Yin Y, et al. Development and Recovery of Liver Injury in Piglets by Incremental Injection of LPS. Antioxidants. 2023; 12(6):1143. https://doi.org/10.3390/antiox12061143
Chicago/Turabian StyleDuan, Geyan, Pan Huang, Changbing Zheng, Jie Zheng, Jiayi Yu, Peiwen Zhang, Mengliao Wan, Fengna Li, Qiuping Guo, Yulong Yin, and et al. 2023. "Development and Recovery of Liver Injury in Piglets by Incremental Injection of LPS" Antioxidants 12, no. 6: 1143. https://doi.org/10.3390/antiox12061143