
Crop, Host, and Gut Microbiome Variation Influence Precision Nutrition: An Example of Blueberries
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diversity of Anthocyanin Profiles across Blueberry Genotypes
2.2. Diversity of Blueberry Polyphenolic Profiles
2.2.1. Blueberry Material
2.2.2. Quantification of Individual Blueberry Phenolics
2.2.3. Statistical Analysis
2.3. Diversity of Bioavailability of Blueberry Polyphenols—Acute PK Study
2.3.1. Analysis of Phenolic Metabolites from Plasma
2.3.2. Statistical Analysis
2.4. Dietary Blueberry Effects on the Gut Microbiome—Repeat Dose, Crossover
2.4.1. Study Design
2.4.2. Microbiota Extraction and Analysis
2.4.3. Microbiota Statistics
3. Results
3.1. Diversity of Anthocyanin Profiles across Blueberry Genotypes
3.2. Diversity of Blueberry Polyphenolic Profiles
Polyphenol Content of Blueberries Used in Bioavailability Studies
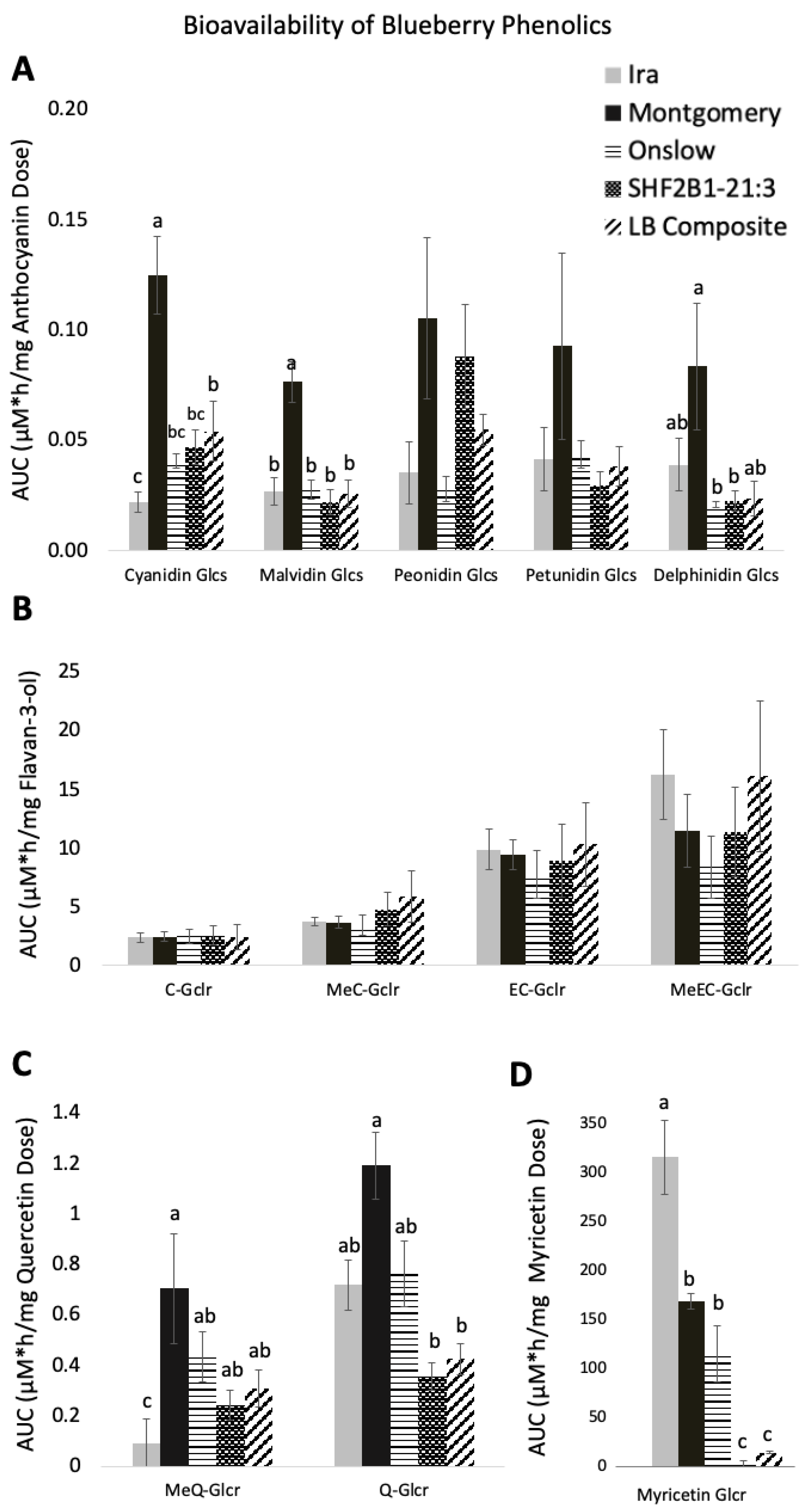
3.3. Diversity of Bioavailability of Blueberry Polyphenols
Bioavailability of Blueberry Metabolites
3.4. Dietary Blueberry Effects on the Gut Microbiome
3.4.1. Gut Microbiota
3.4.2. Taxonomic Analysis
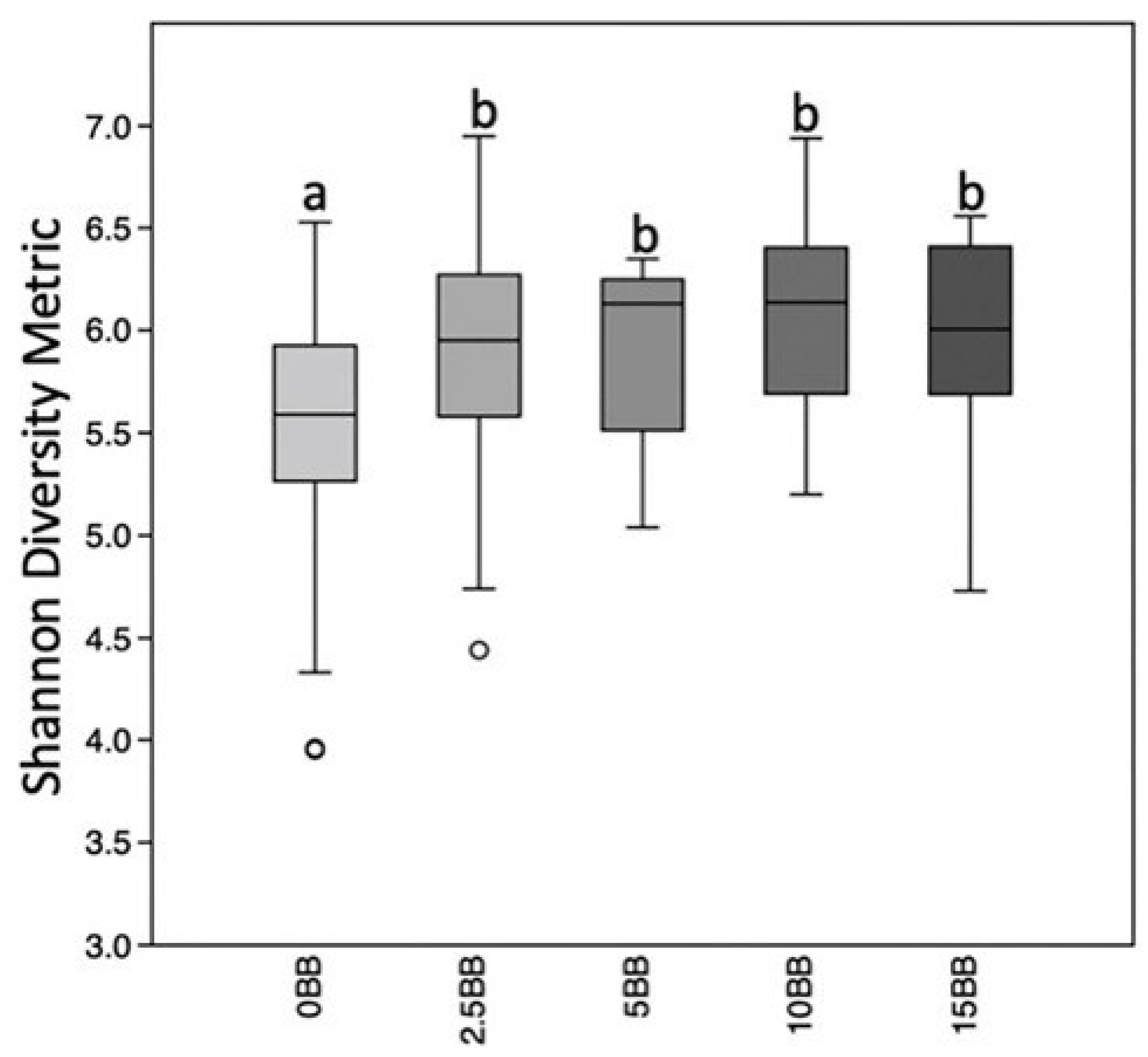
3.4.3. Alpha Diversity
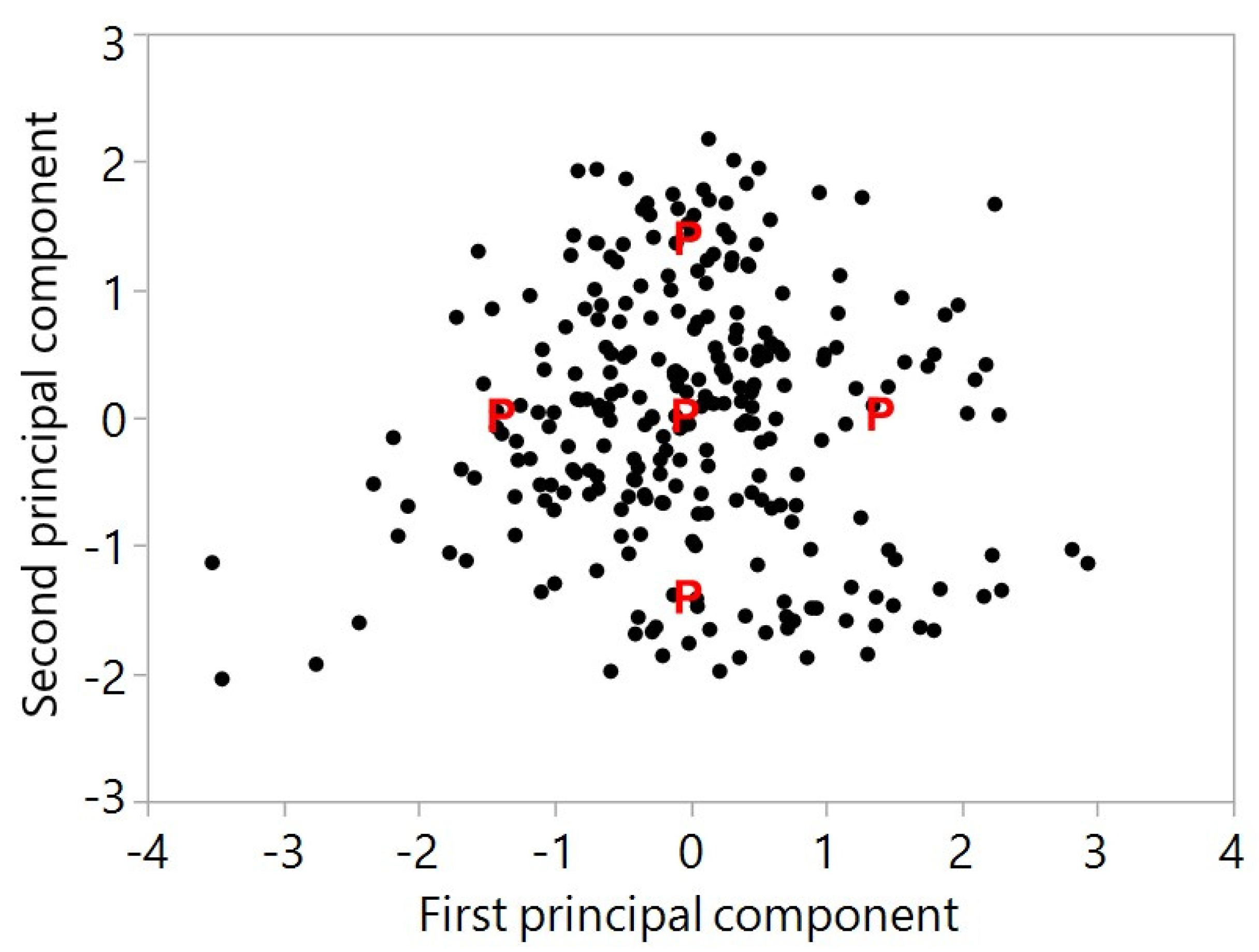
3.4.4. Beta Diversity
3.4.5. Differentially Abundant Taxa
4. Discussion
4.1. Variation in Blueberry Anthocyanin Profiles
4.2. Variation in Bioavailability of Polyphenolics
4.3. Variation in Gut Microbiome in Response to Blueberry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Esposito, K.; Kastorini, C.M.; Panagiotakos, D.B.; Giugliano, D. Prevention of Type 2 Diabetes by Dietary Patterns: A Systematic Review of Prospective Studies and Meta-Analysis. Metab. Syndr. Relat. Disor. 2010, 8, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.P.E. The impact of fruit flavonoids on memory and cognition. Br. J. Nutr. 2010, 104, S40–S47. [Google Scholar] [CrossRef] [PubMed]
- Boeing, H.; Bechthold, A.; Bub, A.; Ellinger, S.; Haller, D.; Kroke, A.; Leschik-Bonnet, E.; Muller, M.J.; Oberritter, H.; Schulze, M.; et al. Critical review: Vegetables and fruit in the prevention of chronic diseases. Eur. J. Nutr. 2012, 51, 637–663. [Google Scholar] [CrossRef] [PubMed]
- Oyebode, O.; Gordon-Dseagu, V.; Walker, A.; Mindell, J.S. Fruit and vegetable consumption and all-cause, cancer and CVD mortality: Analysis of Health Survey for England data. J. Epidemiol. Community Health 2014, 68, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Dillard, C.J.; German, J.B. Phytochemicals: Nutraceuticals and human health. J. Sci. Food Agric. 2000, 80, 1744–1756. [Google Scholar] [CrossRef]
- Son, T.G.; Camandola, S.; Mattson, M.P. Hormetic Dietary Phytochemicals. Neuromol. Med. 2008, 10, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xiao, Y.Y. Grape Phytochemicals and Associated Health Benefits. Crit. Rev. Food Sci. Nutr. 2013, 53, 1202–1225. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)phenolics in Human Health: Structures,Bioavailability, and Evidence of Protective Effects against Chronic Diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Wu, X.L.; Beecher, G.R.; Holden, J.M.; Haytowitz, D.B.; Gebhardt, S.E.; Prior, R.L. Concentrations of anthocyaninsin common foods in the United States and estimation of normal consumption. J. Agric. Food Chem. 2006, 54, 4069–4075. [Google Scholar] [CrossRef]
- Erlund, I.; Koli, R.; Alfthan, G.; Marniemi, J.; Puukka, P.; Mustonen, P.; Mattila, P.; Jula, A. Favorable effects of berry consumption on platelet function, blood pressure, and HDL cholesterol. Am. J. Clin. Nutr. 2008, 87, 323–331. [Google Scholar] [CrossRef]
- Basu, A.; Du, M.; Leyva, M.J.; Sanchez, K.; Betts, N.M.; Wu, M.Y.; Aston, C.E.; Lyons, T.J. Blueberries Decrease Cardiovascular Risk Factors in Obese Men and Women with Metabolic Syndrome. J. Nutr. 2010, 140, 1582–1587. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.A.; Shukitt-Hale, B.; Denisova, N.A.; Bielinski, D.; Martin, A.; McEwen, J.J.; Bickford, P.C. Reversals of age-related declines in neuronal signal transduction, cognitive, and motor behavioral deficits with blueberry, spinach, or strawberry dietary supplementation. J. Neurosci. 1999, 19, 8114–8121. [Google Scholar] [CrossRef] [PubMed]
- Krikorian, R.; Shidler, M.D.; Nash, T.A.; Kalt, W.; Vinqvist-Tymchuk, M.R.; Shukitt-Hale, B.; Joseph, J.A. Blueberry Supplementation Improves Memory in Older Adults. J. Agric. Food Chem. 2010, 58, 3996–4000. [Google Scholar] [CrossRef] [PubMed]
- Andres-Lacueva, C.; Shukitt-Hale, B.; Galli, R.L.; Jauregui, O.; Lamuela-Raventos, R.M.; Joseph, J.A. Anthocyanins in aged blueberry-fed rats are found centrally and may enhance memory. Nutr. Neurosci. 2005, 8, 111–120. [Google Scholar] [CrossRef]
- Zhu, Y.N.; Xia, M.; Yang, Y.; Liu, F.Q.; Li, Z.X.; Hao, Y.T.; Mi, M.T.; Jin, T.R.; Ling, W.H. Purified Anthocyanin Supplementation Improves EndothelialFunction via NO-cGMP Activation in Hypercholesterolemic Individuals. Clin. Chem. 2011, 57, 1524–1533. [Google Scholar] [CrossRef]
- Yousef, G.G.; Brown, A.F.; Funakoshi, Y.; Mbeunkui, F.; Grace, M.H.; Ballington, J.R.; Loraine, A.; Lila, M.A. Efficient quantification of the health-relevant anthocyanin and phenolic acid profiles in commercial cultivars and breeding sections of blueberries (Vaccinium spp.). J. Agric. Food Chem. 2013, 61, 4806–4815. [Google Scholar] [CrossRef]
- Kalt, W.; Ryan, D.A.; Duy, J.C.; Prior, R.L.; Ehlenfeldt, M.K.; Vander Kloet, S.P. Interspecific variabion in anthocyanins, phenolics, and antioxidant capacity among genotypes of high bush and lowbush blueberries (Vaccinium section cyanococcus spp.). J. Agric. Food Chem. 2001, 49, 4761–4767. [Google Scholar] [CrossRef]
- Lila, M.A. The nature-versus-nurture debate on bioactive phytochemicals: The genome versus terroir. J. Sci. Food Agric. 2006, 86, 2510–2515. [Google Scholar] [CrossRef]
- Rodriguez-Mateo, A.; Cifuentes-Gomez, T.; Tabatabaee, S.; Lecras, C.; Spencer, P.E. Procyanidin, anthocyanin, and chlorogenic acid contents of highbush and lowbush blueberries. J. Agric. Food Chem. 2012, 60, 5772–5778. [Google Scholar] [CrossRef]
- Yousef, G.G.; Lila, M.A.; Guzman, I.; Ballington, J.R.; Brown, A.F. Impact of Interspecific Introgression on Anthocyanin Profiles of Southern Highbush Blueberry. J. Am. Soc. Hortic. Sci. 2014, 139, 99–112. [Google Scholar] [CrossRef]
- Joliffe, I.T. Principal Component Analysis, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Furrer, A.; Cladis, D.P.; Kurilich, A.; Manohara, R.; Ferruzzi, M.G. Changes in phenolic content of commercial potato varieties through industrial processing and fresh preparation. Food Chem. 2017, 218, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Cladis, D.P.; Li, S.; Reddivari, L.; Cox, A.; Ferruzzi, M.G.; Weaver, C.M. A 90 day oral toxicity study of blueberry polyphenols in ovariectomized sprague-dawley rats. Food Chem. Toxicol. 2020, 139, 111254. [Google Scholar] [CrossRef] [PubMed]
- Song, B.J.; Sapper, T.N.; Burtch, C.E.; Brimmer, K.; Goldschmidt, M.; Ferruzzi, M.G. Photo- and thermodegradation of anthocyanins from grape and purple sweet potato in model beverage systems. J. Agric. Food Chem. 2013, 61, 1364–1372. [Google Scholar] [CrossRef]
- Lee, J.; Durst, R.W.; Wrolstad, R.E.; Collaborators: Eisele, T.; Giusti, M.M.; Hach, J.; Hofsommer, H.; Koswig, S.; Krueger, D.A.; Kupina, S.; et al. Determination of total monomeric anthocyanin pigment content of fruit juices, beverages, natural colorants, and wines by the pH differential method: Collaborative study. J. AOAC Int. 2005, 88, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.Y.; Kritchevsky, J.; Hargett, K.; Feller, K.; Klobusnik, R.; Song, B.J.; Cooper, B.; Jouni, Z.; Ferruzzi, M.G.; Janle, E.M. Plasma bioavailability and regional brain distribution of polyphenols from apple/grape seed and bilberry extracts in a young swine model. Mol. Nutr. Food Res. 2015, 59, 2432–2447. [Google Scholar] [CrossRef] [PubMed]
- Hodges, J.K.; Maiz, M.; Cao, S.; Lachcik, P.J.; Peacock, M.; McCabe, G.P.; McCabe, L.D.; Cladis, D.P.; Jackson, G.S.; Ferruzzi, M.G.; et al. Moderate Consumption of Freeze-dried Blueberry Powder Increased Net Bone Calcium Retention in Healthy Postmenopausal Women: A Randomized Crossover Trial. Am. J. Clin. Nutr. Under Review.
- Cladis, D.P.; Simpson, A.M.R.; Cooper, K.J.; Nakatsu, C.H.; Ferruzzi, M.G.; Weaver, C.M. Blueberry polyphenols alter gut microbiota and phenolic metabolism in rats. Food Funct. 2021, 12, 2442–2456. [Google Scholar] [CrossRef]
- Bolyen, E.; Rodeout, J.R.; Dillion, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glöckner, F.O. The SILVA and “All-species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Env. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Ellingsen, K.E.; Mcardle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Van Treuren, W.; White, R.; Eggesbø, M.; Knight, R.; Peddada, S. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef]
- Zhong, S.; Sandhu, A.; Edirisinghe, I.; Burton-Freeman, B. Characterization of Wild Blueberry Polyphenols Bioavailability and Kinetic Profile in Plasma over 24-h Period in Human Subjects. Mol. Nutr. Food Res. 2017, 61, 1700405. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.Y.; Ferruzzi, M.G.; Wu, Q.-L.; Simon, J.E.; Talcott, S.T.; Wang, J.; Ho, L.; Todd, G.; Cooper, B.; Pasinetti, G.M.; et al. Influence of diabetes on plasma pharmacokinetics and brain bioavailability of grape polyphenols and their Phase II metabolites in the Zucker Diabetic Fatty Rat. Mol. Nutr. Food Res. 2017, 61, 1700111. [Google Scholar] [CrossRef]
- Nohara, C.; Yokoyama, D.; Tanaka, W.; Sogon, T.; Sakono, M.; Sakakibara, H. Daily consumption of bilberry (Vaccinium myrtillus L.) extracts increases the absorption rate of anthocyanins in rats. J. Agric. Food Chem. 2018, 66, 7958–7964. [Google Scholar] [CrossRef]
- Kurilich, A.C.; Clevidence, B.A.; Britz, S.J.; Simon, P.W.; Novotny, J.A. Plasma and urine responses are lower for acylated vs nonacylated anthocyanins from raw and cooked purple carrots. J. Agric. Food Chem. 2005, 53, 6537–6542. [Google Scholar] [CrossRef]
- Charron, C.S.; Clevidence, B.A.; Britz, S.J.; Novotny, J.A. Effect of dose size on bioavailability of acylated and nonacylated anthocyanins from red cabbage (Brassica oleracea L. Var. capitata). J. Agric. Food Chem. 2007, 55, 5354–5362. [Google Scholar] [CrossRef]
- Charron, C.S.; Kurilich, A.C.; Clevidence, B.A.; Simon, P.W.; Harrison, D.J.; Britz, S.J.; Baer, D.J.; Novotny, J.A. Bioavailability of anthocyanins from purple carrot juice: Effects of acylation and plant matrix. J. Agric. Food Chem. 2009, 57, 1226–1230. [Google Scholar] [CrossRef] [PubMed]
- Terao, J. Factors modulating bioavailability of quercetin-related flavonoids and the consequences of their vascular function. Biochem. Pharmcol. 2017, 139, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Erlund, I.; Kosonen, T.; Alfthan, G.; Mäenpää, J.; Perttunen, K.; Kenraali, J.; Parantainen, J.; Aro, A. Pharmacokinetics of quercetin from quercetin aglycone and rutin in healthy volunteers. Eur. J. Clin. Pharm. 2000, 56, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.L.; Xiao, N.; Li, X.W.; Fan, Y.; Alolga, R.N.; Sun, X.Y.; Wang, S.-L.; Ping, L.; Qi, L.W. Pharmacokinetic comparison between quercetin and quercetin 3-O-β-glucuronide in rats by UHPLC-MS/MS. Sci. Rep. 2016, 6, 35460. [Google Scholar] [CrossRef]
- Canipe III, L.G.; Sioda, M.; Cheatham, C.L. Divesity of the gut-microbiome related to cognitive behavioral outcomes in healthy older adults. Arch. Gerontol. Geriatr. 2021, 96, 104464. [Google Scholar] [CrossRef]
- Sato, A.Y.; Pellegrini, G.G.; Cregor, M.; McAndrews, K.; Choi, R.B.; Maiz, M.; Johnson, O.; McCabe, L.D.; McCabe, G.P.; Ferruzzi, M.G.; et al. Skeletal protection and promotion of microbiome diversity by dietary boosting of the endogenous antioxidant response. J. Bone Min. Res. 2020, 36, 768–778. [Google Scholar] [CrossRef]
- Rodriquez-Daza, M.-C.; Roquim, M.; Dudonne, S.; Pilon, G.; Levy, E.; Marette, A.; Roy, D.; Desardins, Y. Berry polyphenols and fibers modulate distinct microbial metabolic functions and gut microbiota enterotype-like clustering in obese mice. Front. Microbiol. 2020, 11, 2032. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | SIR for Quantitation (m/z) | MRM Transition for Quantitation (m/z) | Standard Used for Quantification |
|---|---|---|---|
| Anthocyanins | |||
| Cyanidin * | 287 | 463 ⟶ 287 | Cyanidin-3-O-glucoside |
| Peonidin * | 301 | 463 ⟶ 301 | |
| Delphinidin * | 303 | 465 ⟶ 303 | |
| Petunidin * | 317 | 479 ⟶ 317 | |
| Malvidin * | 331 | 493 ⟶ 331 | |
| Phenolic Acids ** | |||
| Gallic acid | 169 | Gallic acid | |
| Chlorogenic acid | 353 | Chlorogenic acid | |
| Flavan-3-ols | |||
| Catechin/Epicatechin | 289 | Epicatechin | |
| Catechin/Epicatechin-O-glucuronide | 465 ⟶ 289 | ||
| Me-Catechin/Epicatechin-O-glucuronide | 479⟶ 303 | ||
| Procyanidins B1 and B2 | 577 | ||
| Flavonols | |||
| Quercetin-3-O-arabinoside | 433 | Quercetin-3-O-glucoside | |
| Quercetin-3-O-galactoside/glucoside | 463 | ||
| Quercetin-3-O-glucuronide | 477 | 477 ⟶ 301 | |
| Quercetin-3-O-rhamnoside | 447 | ||
| Me-Quercetin-3-O-glucuronide | 491 ⟶ 315 | ||
| Quercetin-3-O-rutinoside | 609 | Quercetin-3-O-rutinoside | |
| Kaempferol-3-O-galactoside/glucoside | 447 | Kaempferol-3-O-rutinoside | |
| Kaempferol-3-O-rutinoside | 593 | ||
| Myricetin-3-O-galactoside/glucoside | 479 | Quercetin-3-O-glucoside | |
| Myricetin-3-O-glucuronide | 493 ⟶ 317 |
| Delphinidin | Malvidin | Cyanidin | Petunidin | Peonidin | Total | ||
|---|---|---|---|---|---|---|---|
| Ira | 3-O-gal | 150 ± 3.78 d,e | 237 ± 13.2 a | 230 ± 6.0 b | 142 ± 5.5 b | 159 ± 9.3 b | 916 ± 38 A |
| 3-O-glu | 3.1 ± 0.2 f | 14.5 ± 0.5 e | 8.5 ± 0.1 f | 3.7 ± 0.5 e | 15.0 ± 0.6 g | 44.8 ± 2.0 E | |
| 3-O-ara | 78.5 ± 4.3 d,e | 190 ± 15.4 a | 134 ± 5.9 b | 79.7 ± 5.5 c,d | 99.7 ± 3.9 b | 582 ± 35 B | |
| Other forms | 42.5 ± 5.9 e | 42.4 ± 10.1 d,e | 40.0 ± 8.6 b,c,d | 16.7 ± 4.2 e,f | 30.4 ± 6.4 c,d | 172 ± 35 E | |
| Total | 274 ± 14 ζ | 483 ± 39 γ,δ | 412 ± 21 β | 242 ± 16 ε | 304 ± 20 β | 1715 | |
| Legacy | 3-O-gal | 184 ± 15.9 b,c | 195 ± 12.8 b | 41.5 ± 4.0 i | 112 ± 9.9 c | 17.7 ± 1.4 f | 551 ± 44 C |
| 3-O-glu | 3.5 ± 0.3 f | 7.3 ± 1.4 e | 0.2 ± 0.2 g | 4.5 ± 1.3 e | 0.2 ± 0.2 h | 15.6 ± 3.3 E | |
| 3-O-ara | 101 ± 7.7 c | 156 ± 7.9 b | 28.6 ± 2.6 f | 62.0 ± 3.9 e | 14.0 ± 1.2 f | 362 ± 23 E,F | |
| Other forms | 62.4 ± 3.9 d | 13.0 ± 5.1 f | 2.5 ± 0.9 f | 5.3 ± 1.6 g | 0.5 ± 0.5 e | 83.8 ± 12 F | |
| Total | 351 ± 26 δ,ε | 372 ± 17 ε | 72.8 ± 5.9 η | 184 ± 14 ζ | 32.4 ± 2.6 ζ | 1013 | |
| Montgomery | 3-O-gal | 58.8 ± 12.8 g | 94.9 ± 15.9 d,e | 62.9 ± 14.9 g,h | 40.5 ± 8.3 f | 33.6 ± 5.4 e,f | 291 ± 57 D |
| 3-O-glu | 24.3 ± 5.7 e | 74.1 ± 11.3 d | 20.1 ± 4.2 e | 28.0 ± 5.6 d | 33.9 ± 5.6 f | 180 ± 32 D | |
| 3-O-ara | 28.5 ± 6.0 g | 73.1 ± 8.3 e | 26.0 ± 4.7 f | 22.3 ± 3.9 g | 21.5 ± 2.4 e,f | 171 ± 25 G | |
| Other forms | 16.2 ± 1.3 g | 11.1 ± 2.2 f | 0.3 ± 0.6 f | 4.3 ± 0.5 g | 5.1 ± 1.2 e | 37.1 ± 5.8 F | |
| Total | 128 ± 26 η | 253 ± 34 ζ | 109 ± 23 ζ,η | 95 ± 18 η | 94 ± 13 ε | 679 | |
| Onslow | 3-O-gal | 139 ± 2.6 e | 142 ± 1.3 c | 191 ± 2.7 c | 97.4 ± 1.7 c,d,e | 98.3 ± 3.0 c | 668 ± 11 B |
| 3-O-glu | 69.7 ± 1.0 d | 156 ± 1.8 c | 85.2 ± 1.1 b | 78.5 ± 0.3 c | 97.4 ± 0.4 b | 486 ± 4.6 C | |
| 3-O-ara | 64.1 ± 0.9 e,f | 107 ± 0.9 d | 100 ± 1.3 c | 50.7 ± 0.3 f | 60.7 ± 1.6 c | 382 ± 5.1 E | |
| Other forms | 30.8 ± 2.2 f | 37.3 ± 5.4 e | 23.9 ± 5.3 e | 21.1 ± 2.9 e | 33.9 ± 5.2 c | 147 ± 21 E | |
| Total | 304 ± 3.2 ε,ζ | 442 ± 3.4 δ | 400 ± 1.8 β | 248 ± 2.6 ε | 290 ± 5.6 β | 1684 | |
| Sampson | 3-O-gal | 203 ± 6.9 b | 245 ± 2.9 a | 67.1 ± 6.5 f,g | 165 ± 3.1 a | 57.0 ± 19.7 d,e | 737 ± 39 B |
| 3-O-glu | 5.0 ± 0.4 f | 15.3 ± 1.2 e | 1.22 ± 0.2 f,g | 5.3 ± 0.3 e | 2.1 ± 1.2 h | 28.9 ± 3.4 E | |
| 3-O-ara | 129 ± 4.2 b | 209 ± 1.7 a | 53.3 ± 5.7 e | 104 ± 0.5 a | 36.9 ± 14.5 d | 532 ± 27 B,C | |
| Other forms | 62.1 ± 3.0 d | 61.4 ± 2.0 d | 2.0 ± 0.6 f | 16.3 ± 0.8 e,f | 4.4 ± 0.6 e | 146 ± 7.0 E | |
| Total | 399 ± 14 δ | 531 ± 3.7 γ | 124 ± 13 ε,ζ | 291 ± 2.8 δ | 100 ± 36 ε | 1444 | |
| SHF2 B1-21:3 | 3-O-gal | 166 ± 9.1 c,d | 108 ± 3.8 d | 93.0 ± 3.9 e | 102 ± 6.1 c,d | 39.0 ± 1.5 d,e,f | 508 ± 24 C |
| 3-O-glu | 159 ± 11.0 b | 173 ± 10.4 b | 65.8 ± 2.7 c | 143 ± 8.2 a | 48.5 ± 2.3 d,e | 589 ± 35 B | |
| 3-O-ara | 134 ± 6.4 b | 136 ± 4.3 c | 72.5 ± 2.2 d | 86.7 ± 3.0 b,c | 30.6 ± 1.2 d,e | 461 ± 17 D | |
| Other forms | 130 ± 5.7 b | 223 ± 4.3 b | 43.5 ± 3.1 b | 111 ± 2.2 a | 43.3 ± 2.2 b,c | 551 ± 17 B | |
| Total | 589 ± 26 β | 641 ± 22 β | 275 ± 8.2 δ | 442 ± 18 α | 162 ± 6.0 δ | 2108 | |
| Cranberry | 3-O-gal | nd | 9.0 ± 0.4 f | 157 ± 6.9 d | trace | 342 ± 13.3 a | 507 ± 21 C |
| 3-O-glu | nd | trace | 4.6 ± 0.2 f,g | nd | 41.7 ± 3.5 e | 46.3 ± 3.7 E | |
| 3-O-ara | nd | 11.0 ± 0.3 f | 139 ± 1.2 b | trace | 247 ± 4.7 a | 396 ± 6.3 E | |
| Other forms | 90.4 ± 2.8 c | 0.5 ± 0.5 f | 41.7 ± 8.7 b,c | 32.6 ± 0.2 d | 94.8 ± 21.1 a | 260 ± 33 D | |
| Total | 90.4 ± 2.8 η | 20.5 ± 0.4 η | 342 ± 2.6 γ | 32.6 ± 0.2 θ | 725 ± 0.2 α | 1210 | |
| Wild Blueberry | 3-O-gal | 58.8 ± 3.5 g | 113 ± 5.4 d | 42.7 ± 1.6 h,i | 52.4 ± 2.8 f | 40.7 ± 1.1 d,e,f | 307 ± 14 D |
| 3-O-glu | 78.7 ± 3.7 d | 170 ± 6.5 b,c | 50.4 ± 3.2 d | 86.6 ± 4.8 c | 55.9 ± 2.4 d | 442 ± 21 C | |
| 3-O-ara | 46.5 ± 2.5 f,g | 132 ± 6.4 c | 41.3 ± 2.5 e | 46.1 ± 2.1 f | 39.7 ± 1.5 d | 305 ± 15 F | |
| Other forms | 93.2 ± 5.3 c | 187 ± 14.0 c | 27.2 ± 2.1 d,e | 47.6 ± 3.8 c | 58.6 ± 4.3 b | 414 ± 29 C | |
| Total | 277 ± 14 ζ | 602 ± 31 β | 162 ± 9.2 ε | 233 ± 13 ε | 195 ± 8.8 γ,δ | 1468 | |
| Bilberry | 3-O-gal | 258 ± 9.4 a | 79.0 ± 3.0 e | 269 ± 11.5 a | 92.1 ± 4.3 d,e | 45.1 ± 1.2 d,e | 744 ± 29 B |
| 3-O-glu | 186 ± 9.1 a | 162 ± 3.0 b,c | 182 ± 6.3 a | 131 ± 6.1 a,b | 114 ± 2.9 a | 775 ± 27 A | |
| 3-O-ara | 277 ± 13.4 a | 98.9 ± 1.5 d | 253 ± 8.4 a | 93.3 ± 3.6 b | 43.1 ± 1.0 d | 765 ± 28 A | |
| Other forms | 20.1 ± 1.1 f,g | 13.9 ± 3.2 f | 28.8 ± 5.9 c,d,e | 13.7 ± 2.6 f | 9.0 ± 2.1 d,e | 85.6 ± 15 F | |
| Total | 741 ± 33 α | 354 ± 7.8 ε | 733 ± 28 α | 330 ± 15 γ | 211 ± 5.5 γ | 2369 | |
| Lowbush composite | 3-O-gal | 107 ± 1.0 f | 163 ± 1.7 c | 85.4 ± 1.2 e,f | 84.1 ± 0.6 e | 62.9 ± 0.8 d | 502 ± 5.2 C |
| 3-O-glu | 125 ± 1.2 c | 228 ± 2.4 a | 85.2 ± 0.9 b | 126 ± 1.0 b | 77.7 ± 0.6 c | 641 ± 6.1 B | |
| 3-O-ara | 86.6 ± 2.1 c,d | 194 ± 4.6 a | 79.0 ± 1.0 d | 76.5 ± 2.9 d | 60.7 ± 0.8 c | 497 ± 11 C,D | |
| Other forms | 168 ± 5.0 a | 334 ± 7.3 a | 65.4 ± 2.1 a | 96.5 ± 1.9 b | 96.0 ± 3.1 a | 760 ± 19 A | |
| Total | 487 ± 6.3 γ | 918 ± 9.2 α | 315 ± 3.9 γ,δ | 383 ± 3.5 β | 297 ± 3.7 β | 2400 |
| Phenolic Acids | Flavan-3-ols | |||||
|---|---|---|---|---|---|---|
| Gallic Acid | Chlorogenic Acid | Catechin | Epicatechin | Procyanidin B1 | Procyanidin B2 | |
| Ira | 0.70 ± 0.09 b,c,d | 755 ± 46.7 a | 15.9 ± 1.28 a | 1.07 ± 0.25 d,e | 8.27 ± 0.96 a | 0.75 ± 0.31 e |
| Legacy | 0.52 ± 0.04 c,d | 520 ± 58.5 b | 8.11 ± 0.67 c | 0.71 ± 0.27 e | 4.02 ± 0.23 c | 2.51 ± 0.44 d,e |
| Montgomery | 0.47 ± 0.03 c,d | 493 ± 48.9 b | 12.8 ± 0.29 b | 1.63 ± 0.07 c,d,e | 6.56 ± 0.49 b | 2.03 ± 0.23 e |
| Onslow | 0.57 ± 0.19 c,d | 496 ± 30.5 b | 15.2 ± 0.57 a | 3.24 ± 0.09 c | 7.17 ± 0.29 a,b | 4.23 ± 0.24 c,d |
| Sampson | 0.92 ± 0.09 b,c | 269 ± 9.03 c | 3.90 ± 0.16 d | 0.41 ± 0.28 e | 0.91 ± 0.19 d | 0.89 ± 0.44 e |
| SHF2B1-21:3 | 0.66 ± 0.04 b,c,d | 492 ± 26.4 b | 6.97 ± 0.10 c | 1.89 ± 0.88 c,d,e | 3.23 ± 0.26 c | 2.67 ± 0.62 d,e |
| Cranberry | 0.38 ± 0.08 d | 20.3 ± 0.64 d | 1.93 ± 0.07 e | 19.3 ± 0.82 a | trace | 13.8 ± 1.49 b |
| Wild BB | 0.52 ± 0.02 c,d | 566 ± 31.6 b | 7.64 ± 0.78 c | 3.04 ± 0.32 c,d | 6.21 ± 0.31 b | 4.62 ± 0.48 c,d |
| Bilberry | 2.04 ± 0.37 a | 104 ± 3.58 c,d | 0.06 ± 0.06 f | 10.9 ± 1.75 b | trace | 19.3 ± 1.33 a |
| LB composite | 1.11 ± 0.32 b | 740 ± 25.1 a | 8.02 ± 0.43 c | 1.67 ± 0.64 c,d,e | 7.02 ± 0.53 a,b | 5.44 ± 0.62 c |
| Quer 3-O-arb | Quer 3-O-rham | Quer 3-O-glcs | Quer 3-O-gcnd | Quer 3-O-rut | Total Quer | Kaemp 3-O-glcs | Kaemp 3-O-rut | Myr 3-O-glcs | |
|---|---|---|---|---|---|---|---|---|---|
| Ira | 0.34 ± 0.14 f | 58.1 ± 9.21 b | 26.1 ± 3.18 d | 8.92 ± 0.34 c,d | 14.3 ± 1.30 c,d,e | 108 ± 11.7 e | 0.09 ± 0.08 d | trace | 2.20 ± 0.12 e |
| Legacy | 28.1 ± 7.30 d | 24.7 ± 6.50 c | 249 ± 29.1 a | 1.14 ± 0.48 d | 9.18 ± 2.16 d,e,f | 313 ± 43.7 b,c | 3.35 ± 1.03 a | 0.57 ± 0.31 b,c | 19.0 ± 1.40 c,d |
| Montgomery | 17.3 ± 1.40 d,e,f | 27.7 ± 1.35 c | 23.9 ± 2.83 d | 6.04 ± 0.59 d | 17.6 ± 1.15 c,d | 92.5 ± 5.48 e | 0.59 ± 0.06 c,d | 0.71 ± 0.11 b,c | 4.86 ± 0.10 d,e |
| Onslow | 26.3 ± 4.12 d | 47.8 ± 4.76 b | 33.3 ± 4.14 d | 15.5 ± 0.44 c | 7.68 ± 1.85 e,f | 131 ± 5.70 e | 0.73 ± 0.12 c,d | 0.34 ± 0.19 c | 2.35 ± 0.54 e |
| Sampson | 92.7 ± 8.20 b | 113 ± 6.15 a | 124 ± 13.0 c | trace | 31.3 ± 3.35 a | 360 ± 22.4 a,b | 1.10 ± 0.12 c,d | 1.77 ± 0.36 a | 26.1 ± 4.18 c |
| SHF2B1-21:3 | 20.6 ± 2.35 d,e | 13.9 ± 2.14 c,d | 156 ± 16.6 c | trace | 4.81 ± 0.74 f | 194 ± 19.1 d | 0.64 ± 0.07 c,d | trace | 52.1 ± 15.7 a,b |
| Cranberry | 136 ± 11.0 a | 21.1 ± 3.05 c | 206 ± 15.8 b | trace | trace | 363 ± 22.6 a,b | 0.72 ± 0.26 c,d | nd | 58.7 ± 9.74 a |
| Wild BB | 26.9 ± 2.12 d | 20.7 ± 1.94 c | 161 ± 21.9 c | 40.3 ± 6.84 b | 20.9 ± 3.06 b,c | 269 ± 25.5 c | 1.59 ± 0.30 b,c | 1.32 ± 0.22 a,b | 22.4 ± 3.80 c,d |
| Bilberry | 7.37 ± 1.26 e,f | 1.09 ± 0.21 d | 47.5 ± 6.38 d | 38.5 ± 1.40 b | trace | 94.5 ± 8.75 e | trace | trace | 35.3 ± 8.01 b,c |
| LB composite | 64.1 ± 12.4 c | 54.6 ± 7.43 b | 210 ± 9.01 a,b | 60.5 ± 4.19 a | 29.0 ± 7.50 a,b | 418 ± 20.6 a | 2.28 ± 0.30 a,b | 2.00 ± 0.51 a | 27.8 ± 1.52 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weaver, C.M.; Ferruzzi, M.G.; Maiz, M.; Cladis, D.P.; Nakatsu, C.H.; McCabe, G.P.; Lila, M.A. Crop, Host, and Gut Microbiome Variation Influence Precision Nutrition: An Example of Blueberries. Antioxidants 2023, 12, 1136. https://doi.org/10.3390/antiox12051136
Weaver CM, Ferruzzi MG, Maiz M, Cladis DP, Nakatsu CH, McCabe GP, Lila MA. Crop, Host, and Gut Microbiome Variation Influence Precision Nutrition: An Example of Blueberries. Antioxidants. 2023; 12(5):1136. https://doi.org/10.3390/antiox12051136
Chicago/Turabian StyleWeaver, Connie M., Mario G. Ferruzzi, Maria Maiz, Dennis P. Cladis, Cindy H. Nakatsu, George P. McCabe, and Mary Ann Lila. 2023. "Crop, Host, and Gut Microbiome Variation Influence Precision Nutrition: An Example of Blueberries" Antioxidants 12, no. 5: 1136. https://doi.org/10.3390/antiox12051136