
Discrimination of Green Coffee (Coffea arabica and Coffea canephora) of Different Geographical Origin Based on Antioxidant Activity, High-Throughput Metabolomics, and DNA RFLP Fingerprinting



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Extraction
2.2. Total Polyphenol Content (TPC)
2.3. Total Flavonoid Content
2.4. Total Flavonol Content
2.5. Total Flavan-3-ol Content
2.6. Radical-Scavenging Activity
2.7. Reducing Activity
2.8. Identification and Quantification of Plant Bioactive Compounds
2.9. Identification and Quantification of Fatty Acids
2.10. DNA Extraction, PCR Amplification, Subcloning, and Sequencing
2.11. PCR-RFLP Analysis
2.12. Statistical Analysis
3. Results and Discussion
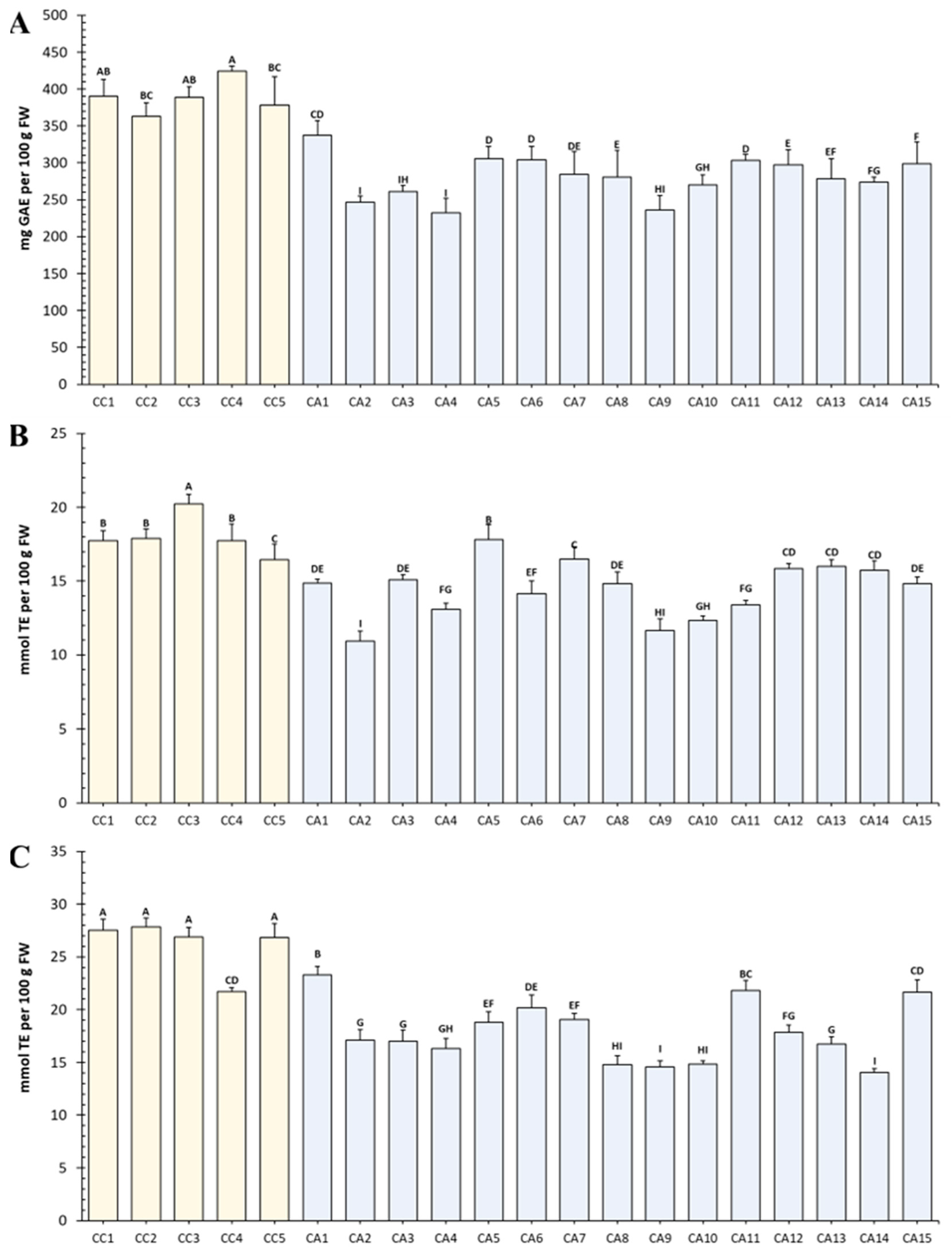
3.1. The Total Polyphenol Content Correlates with the Antioxidant Power of Green Coffee Extracts
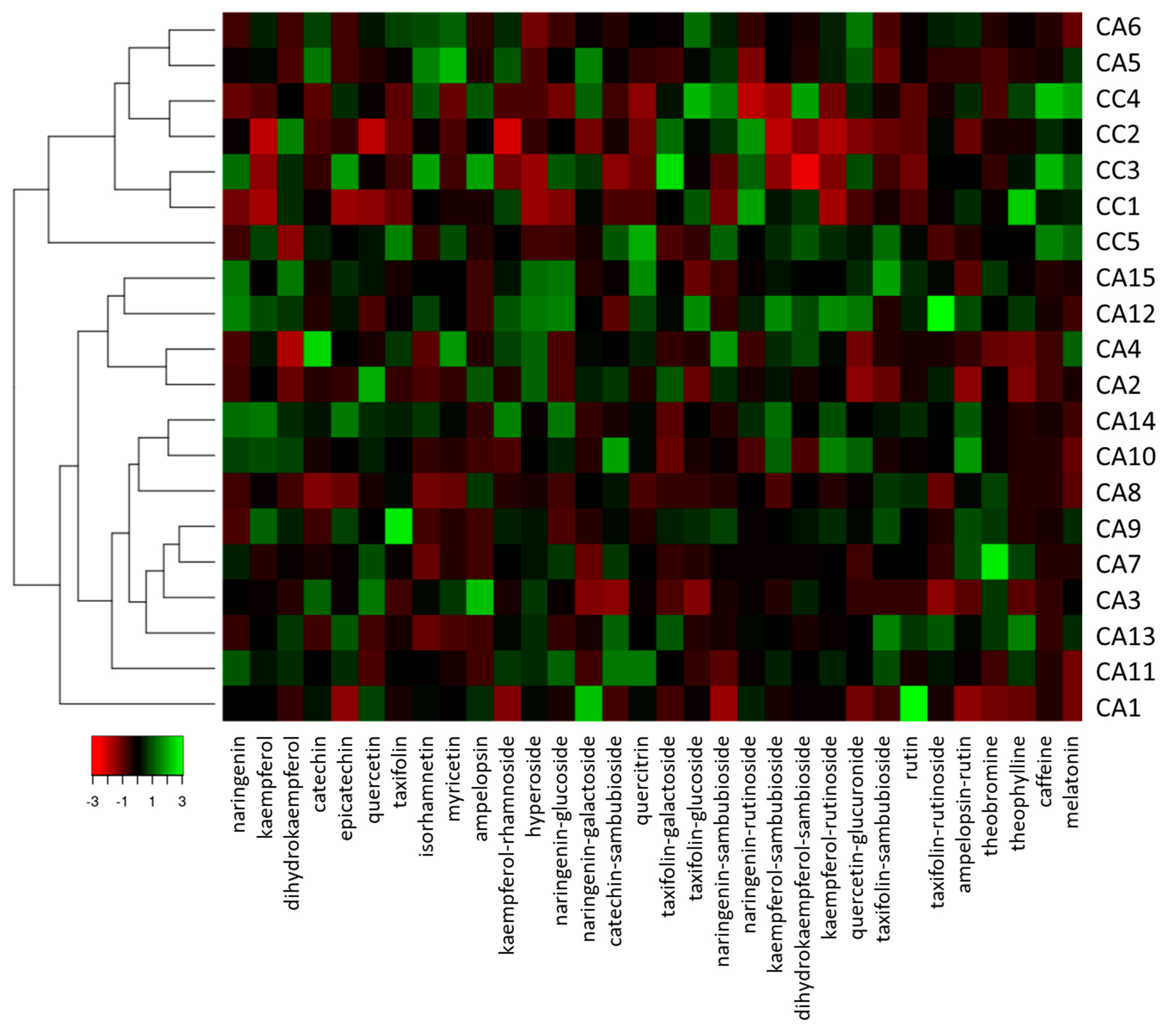
3.2. Phenolic Compounds, Xanthine Derivatives, and Melatonin Contents Discriminate C. arabica from C. canephora Accessions
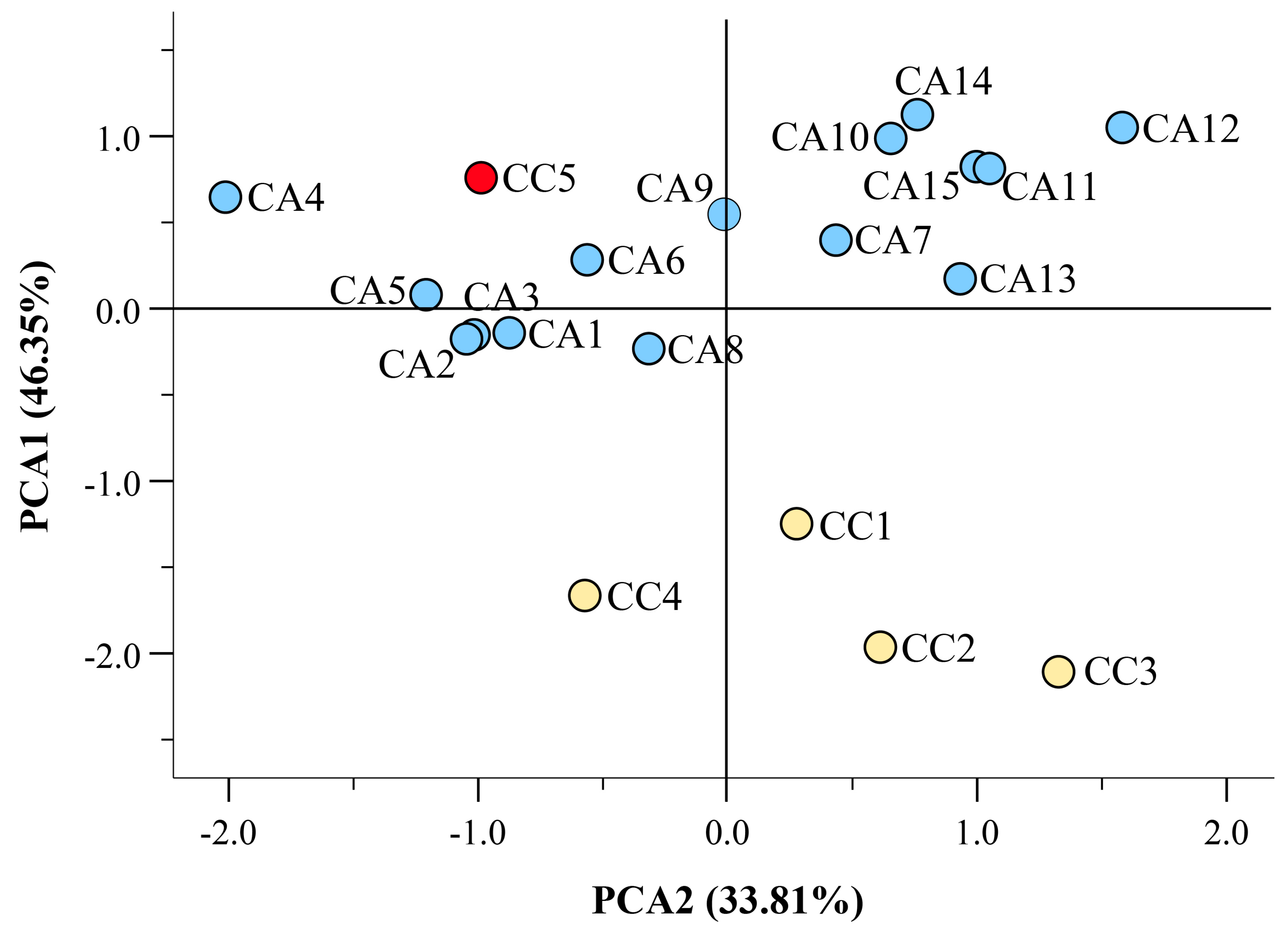
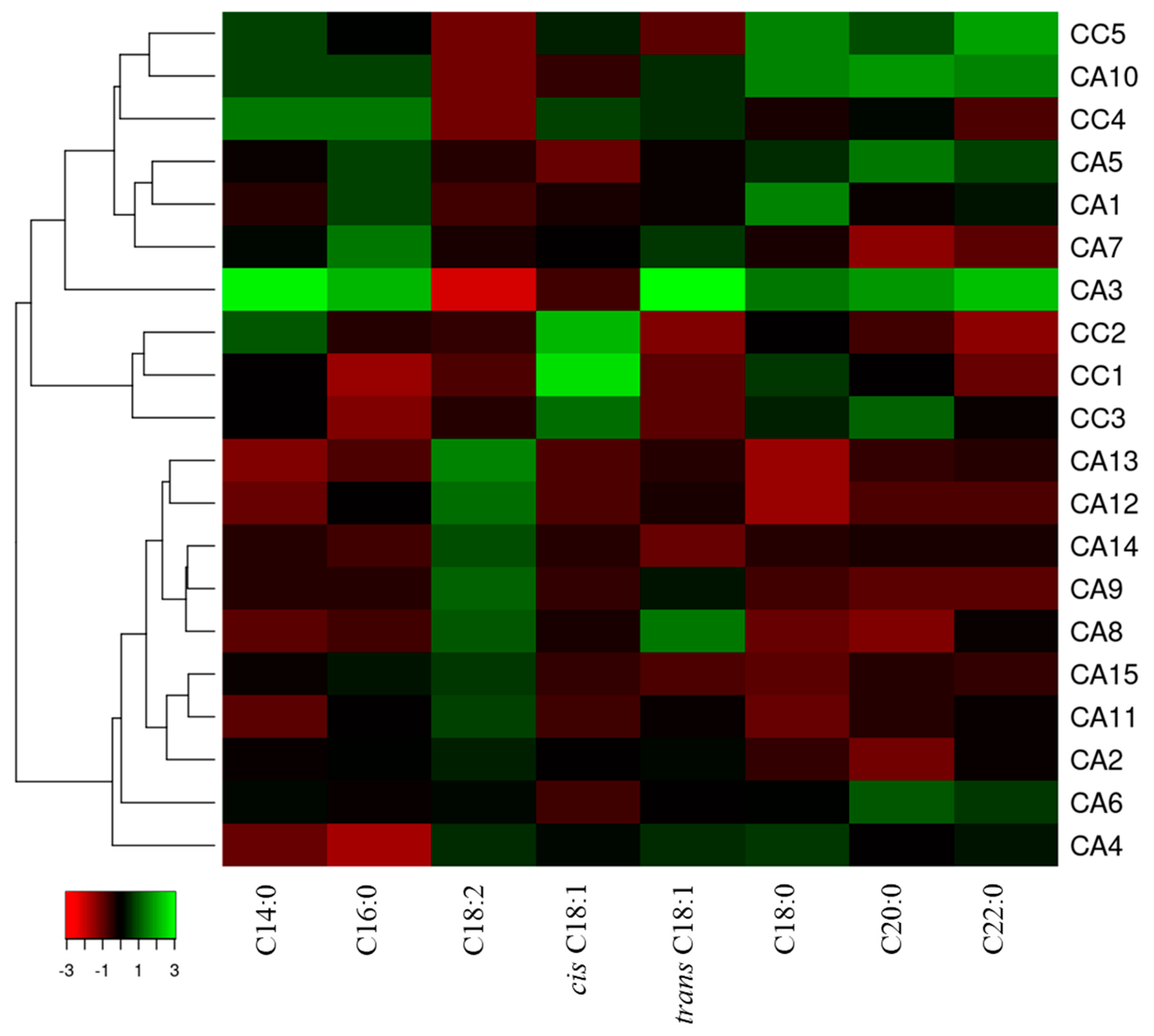
3.3. The Fatty Acid Composition and Content Discriminate C. arabica and C. canephora Accessions of Different Geographical Origin
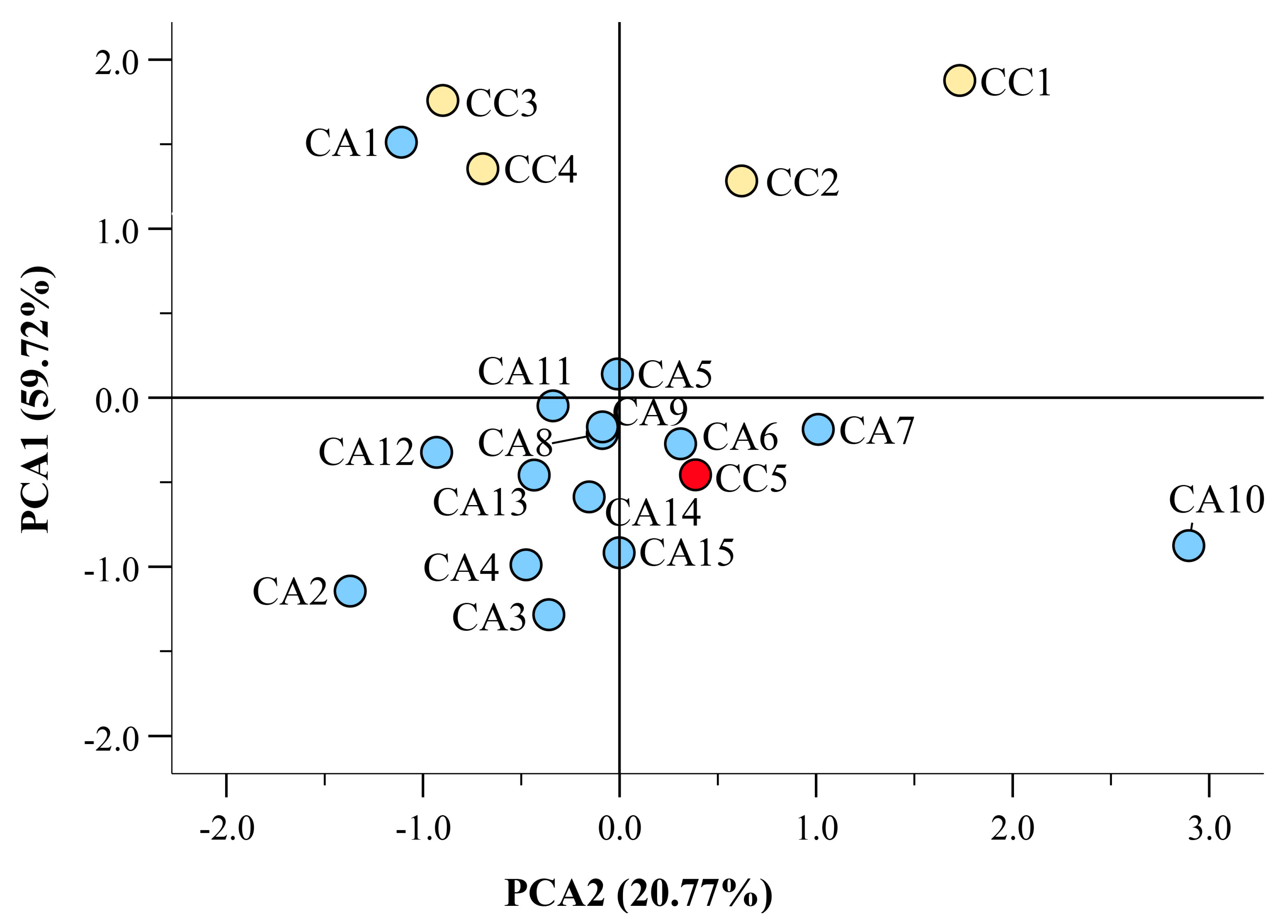
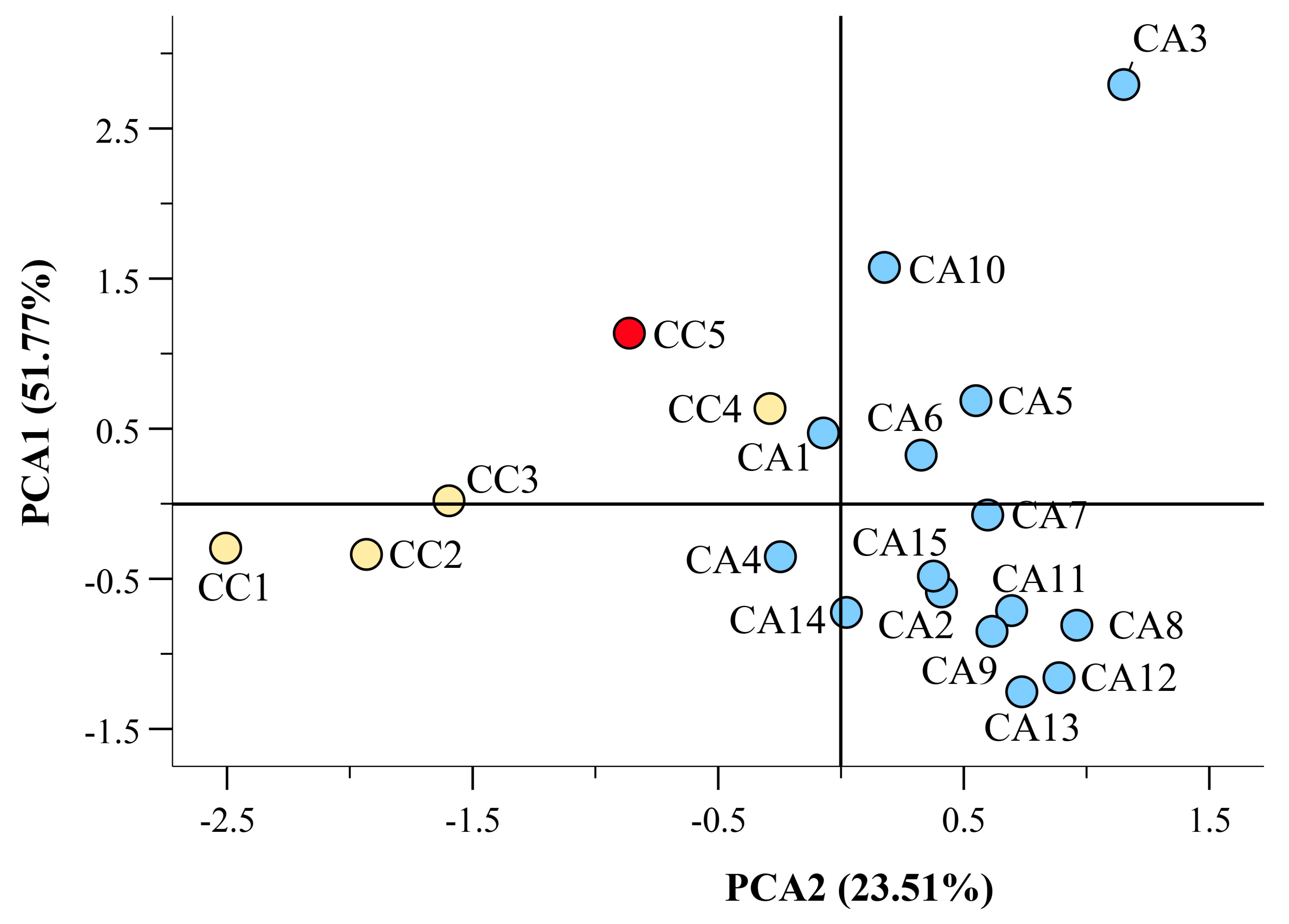
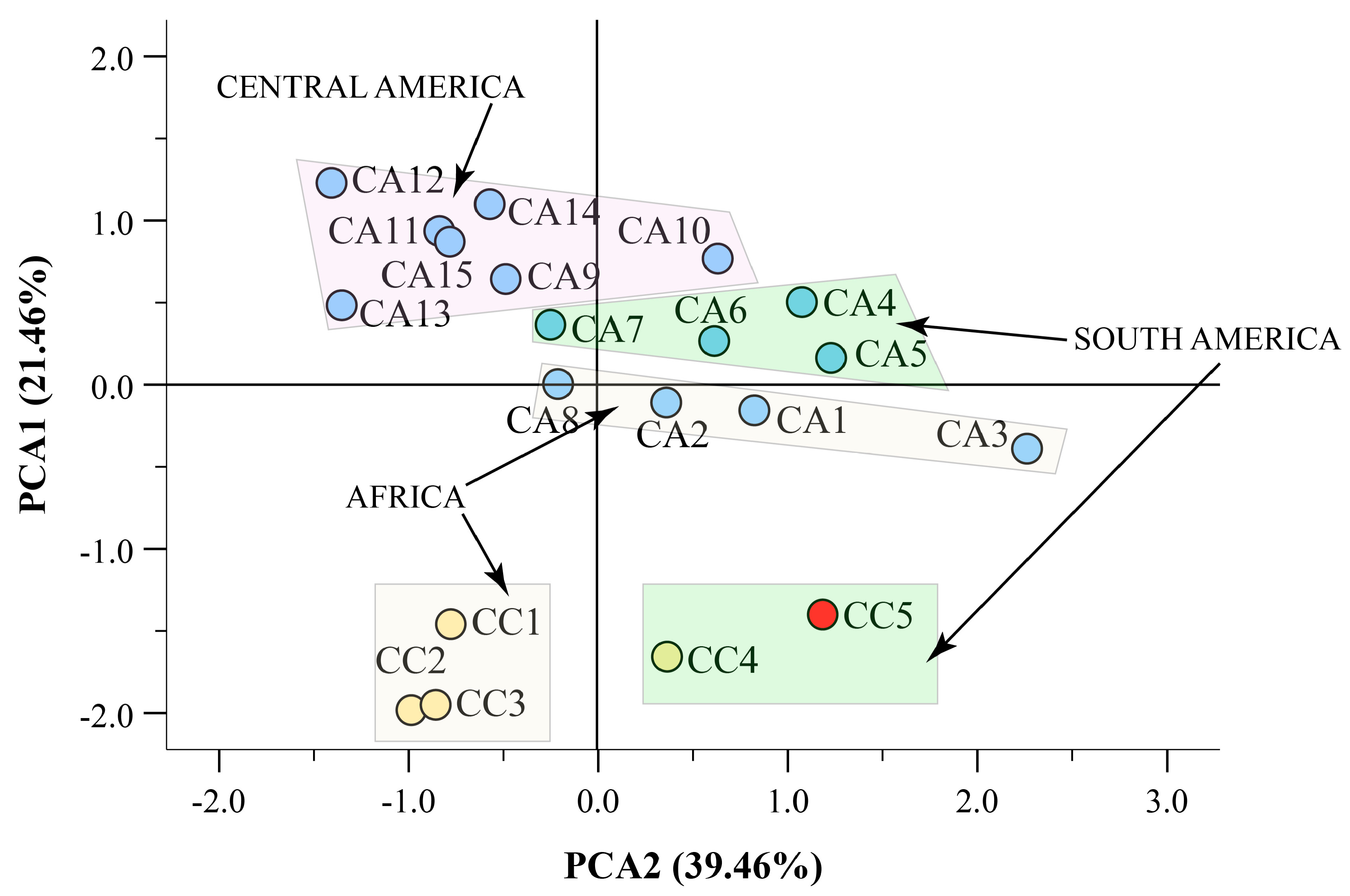
3.4. Hight Throughput Analysis Allows the Metabolic and Geographical Discrimination of C. arabica and C. canephora
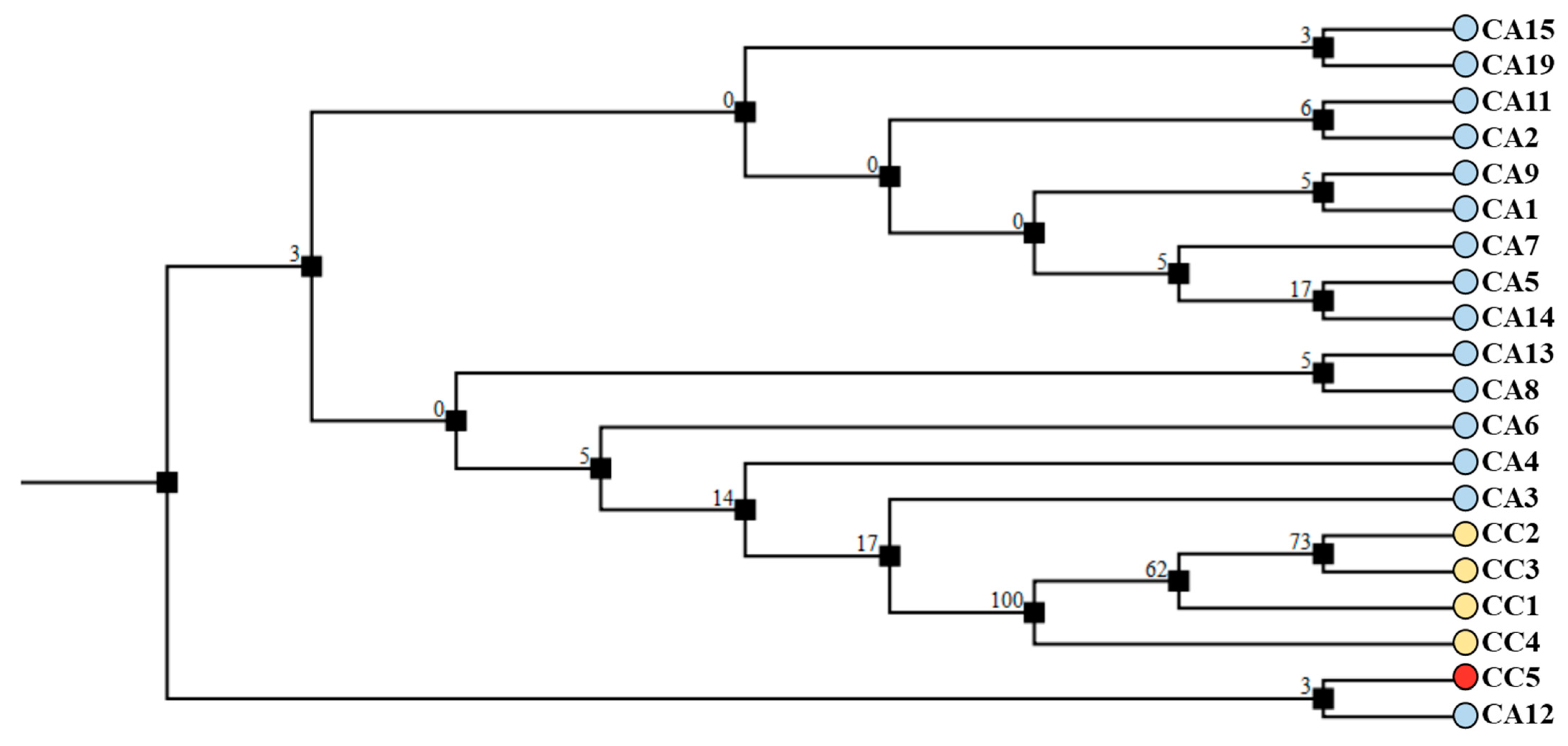
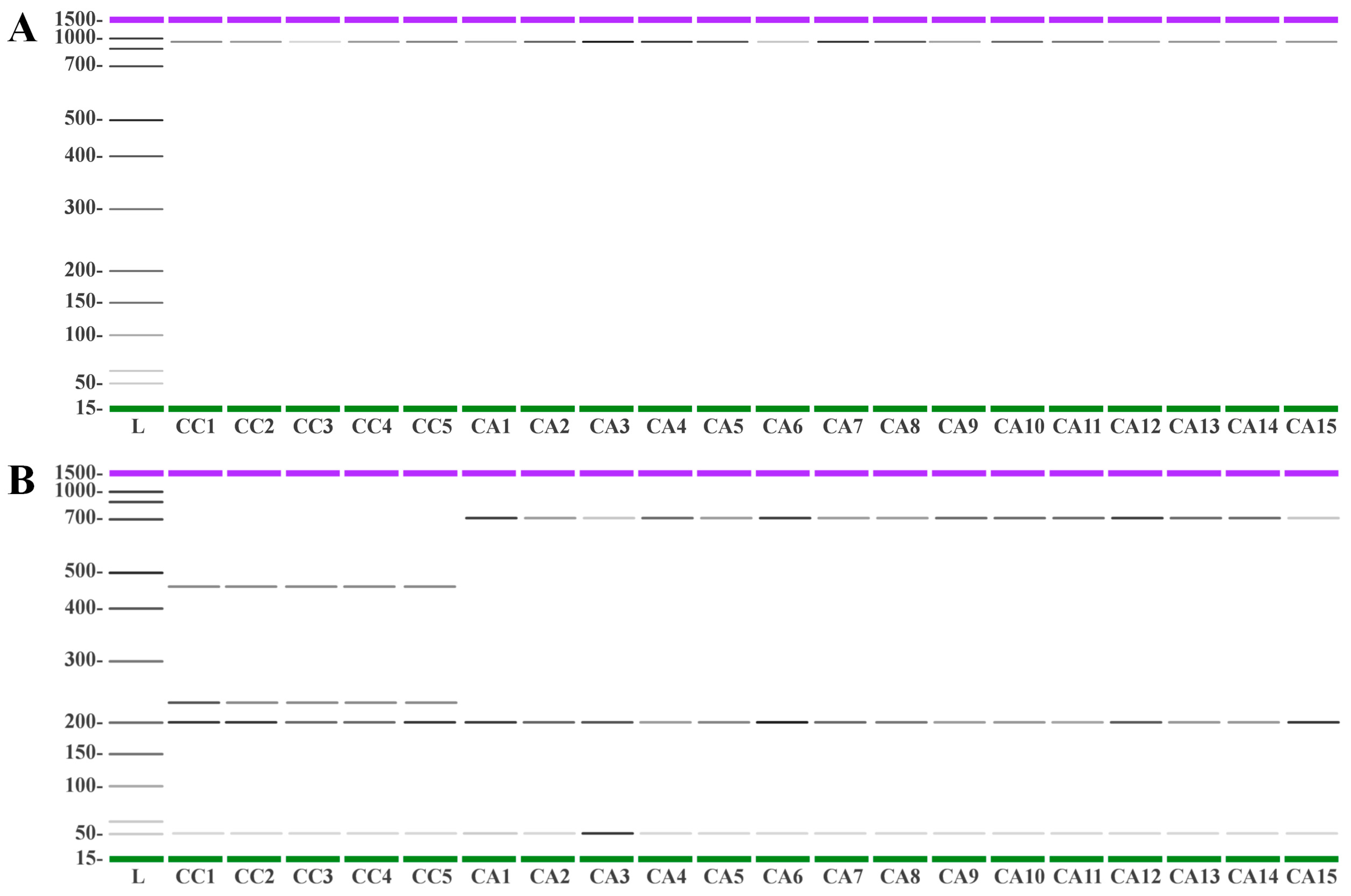
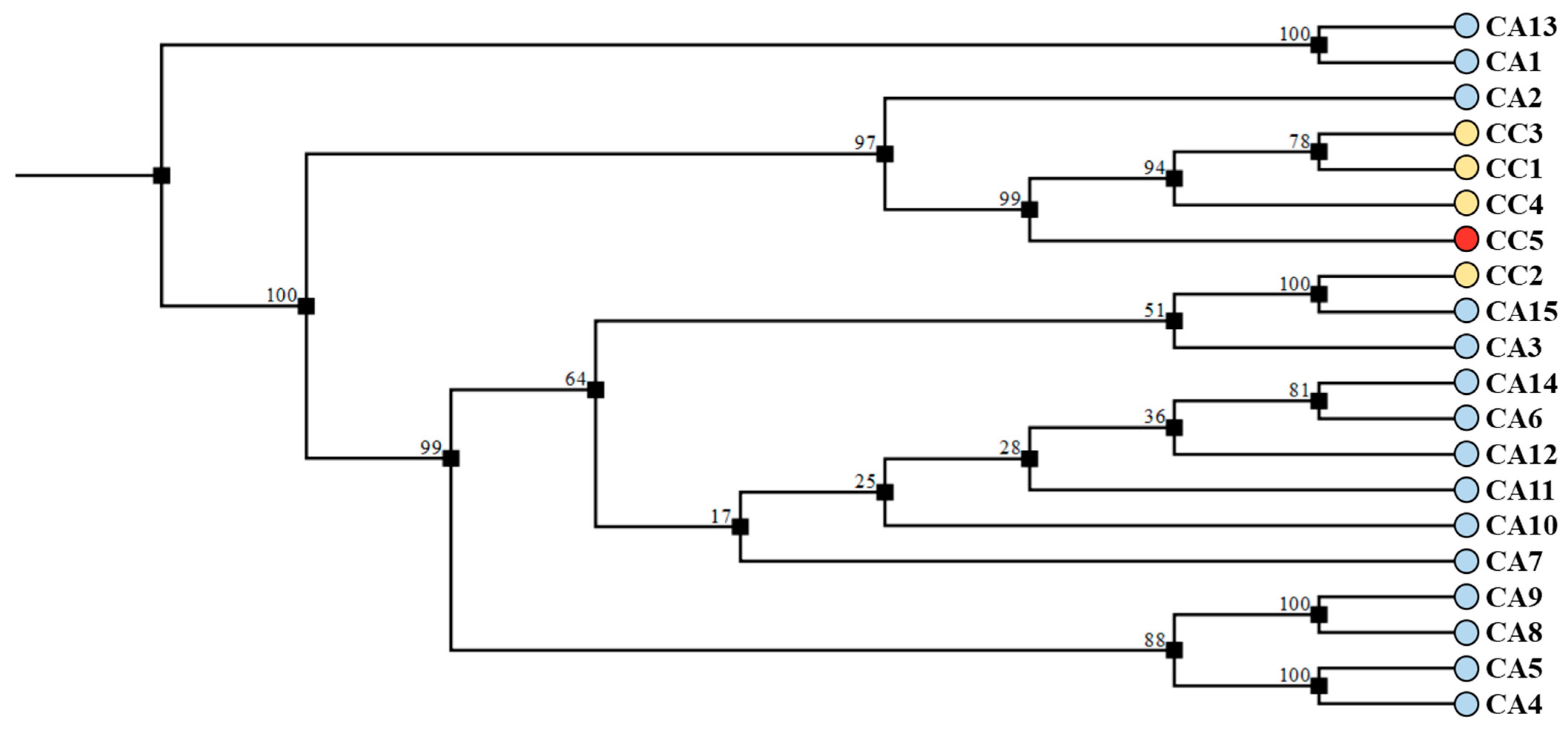
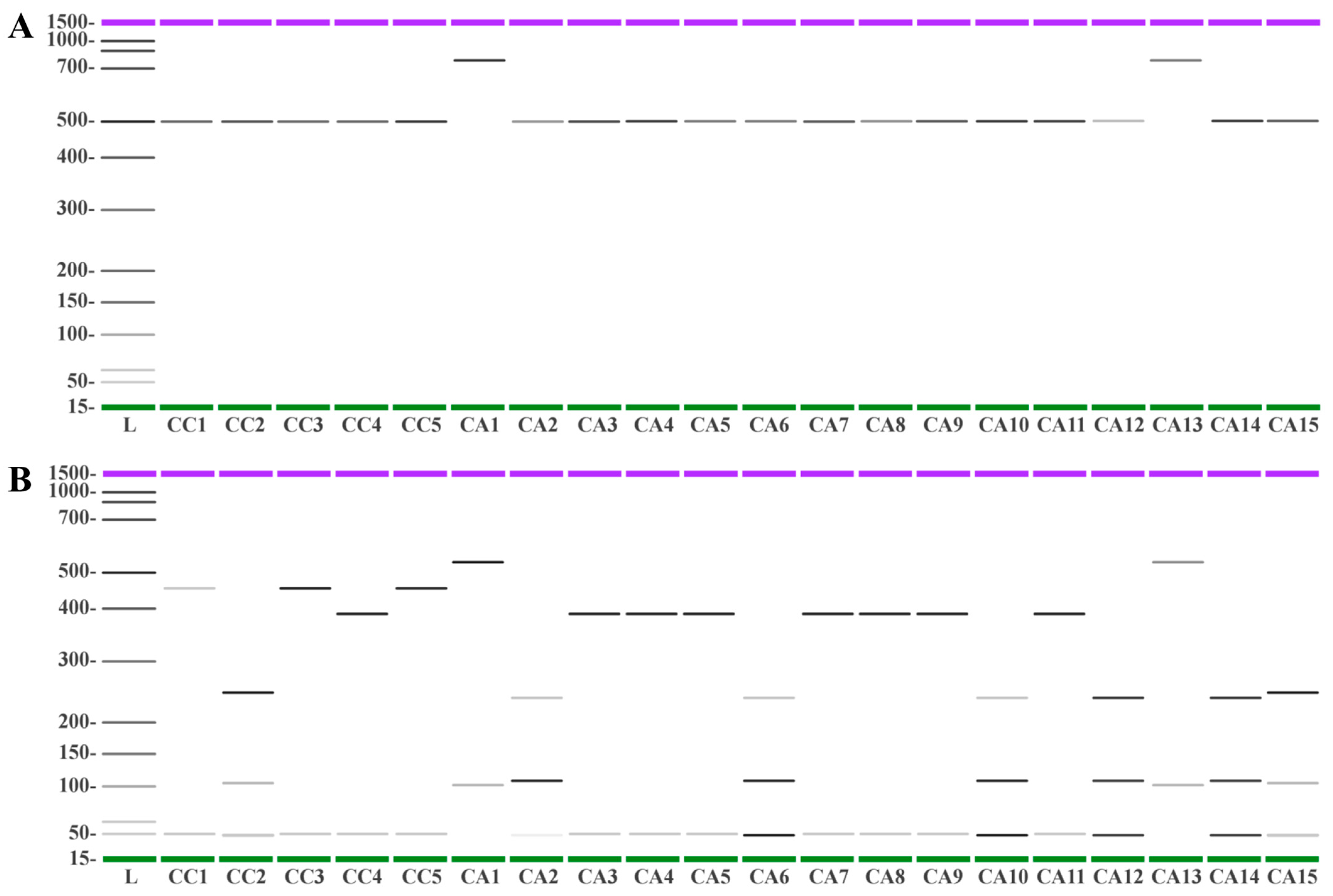
3.5. DNA Fingerprinting Using trnL-trnF and 5S-rRNA-NTS Regions Allows the Molecular Discrimination of Species and Accessions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bicho, N.C.; Lidon, F.C.; Ramalho, J.C.; Leitao, A.E. Quality assessment of Arabica and Robusta green and roasted coffees—A review. Emir. J. Food Agric. 2013, 25, 945–950. [Google Scholar] [CrossRef]
- Anderson, K.A.; Smith, B.W. Chemical profiling to differentiate geographic growing origins of coffee. J. Agric. Food Chem. 2002, 50, 2068–2075. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Govaerts, R.; Bridson, D.M.; Stoffelen, P. An annotated taxonomic conspectus of the genus Coffea (Rubiaceae). Bot. J. Linn. Soc. 2006, 152, 465–512. [Google Scholar] [CrossRef]
- Vega, F.E.; Rosenquist, E.; Collins, W. Global project needed to tackle coffee crisis. Nature 2003, 425, 343. [Google Scholar] [CrossRef]
- Robinson, J.L.; Hunter, J.M.; Reyes-Izquierdo, T.; Argumedo, R.; Brizuela-Bastien, J.; Keller, R.; Pietrzkowski, Z.J. Cognitive short- and long-term effects of coffee cherry extract in older adults with mild cognitive decline. Neuropsychol. Dev. Cognition. Sect. B Aging Neuropsychol. Cogn. 2020, 27, 918–934. [Google Scholar] [CrossRef]
- Samoggia, A.; Del Prete, M.; Argenti, C. Functional Needs, Emotions, and Perceptions of Coffee Consumers and Non-Consumers. Sustainability 2020, 12, 5694. [Google Scholar] [CrossRef]
- Achón, M.; Úbeda, N.; García-González, Á.; Partearroyo, T.; Varela-Moreiras, G. Effects of Milk and Dairy Product Consumption on Pregnancy and Lactation Outcomes: A Systematic Review. Adv. Nutr. 2019, 10, S74–S87. [Google Scholar] [CrossRef]
- Acidri, R.; Sawai, Y.; Sugimoto, Y.; Handa, T.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Phytochemical Profile and Antioxidant Capacity of Coffee Plant Organs Compared to Green and Roasted Coffee Beans. Antioxidants 2020, 9, 93. [Google Scholar] [CrossRef]
- Clifford, M.N. Chlorogenic acids and other cinnamates—nature, occurrence and dietary burden. J. Sci. Food Agric. 1999, 79, 362–372. [Google Scholar] [CrossRef]
- Del Rio, D.; Stalmach, A.; Calani, L.; Crozier, A. Bioavailability of Coffee Chlorogenic Acids and Green Tea Flavan-3-ols. Nutrients 2010, 2, 820–833. [Google Scholar] [CrossRef]
- Clifford, M.N.; Knight, S. The cinnamoyl-amino acid conjugates of green robusta coffee beans. Food Chem. 2004, 87, 457–463. [Google Scholar] [CrossRef]
- Ky, C.L.; Barre, P.; Noirot, M. Genetic investigations on the caffeine and chlorogenic acid relationship in an interspecific cross between Coffea liberica dewevrei and C. pseudozanguebariae. Tree Genet. Genomes 2013, 9, 1043–1049. [Google Scholar] [CrossRef]
- Baeza, G.; Amigo-Benavent, M.; Sarria, B.; Goya, L.; Mateos, R.; Bravo, L. Green coffee hydroxycinnamic acids but not caffeine protect human HepG2 cells against oxidative stress. Food Res. Int. 2014, 62, 1038–1046. [Google Scholar] [CrossRef]
- Febrianto, N.A.; Zhu, F. Coffee bean processing: Emerging methods and their effects on chemical, biological and sensory properties. Food Chem. 2023, 412, 20. [Google Scholar] [CrossRef] [PubMed]
- Pimpley, V.A.; Maity, S.; Murthy, P.S. Green coffee polyphenols in formulations of functional yoghurt and their quality attributes. Int. J. Dairy Technol. 2022, 75, 159–170. [Google Scholar] [CrossRef]
- Klikarova, J.; Ceslova, L. Targeted and Non-Targeted HPLC Analysis of Coffee-Based Products as Effective Tools for Evaluating the Coffee Authenticity. Molecules 2022, 27, 24. [Google Scholar] [CrossRef] [PubMed]
- Macedo, L.L.; Araujo, C.D.; Vimercati, W.C.; Hein, P.R.G.; Pimenta, C.J.; Saraiva, S.H. Evaluation of chemical properties of intact green coffee beans using near-infrared spectroscopy. J. Sci. Food Agric. 2021, 101, 3500–3507. [Google Scholar] [CrossRef]
- Belchior, V.; Botelho, B.G.; Franca, A.S. Comparison of Spectroscopy-Based Methods and Chemometrics to Confirm Classification of Specialty Coffees. Foods 2022, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.J.; Jeong, H.J.; Moon, H.; Kim, H.N.; Cho, J.H.; Lee, J.E.; Hong, K.S.; Hong, Y.S. Assessment of green coffee bean metabolites dependent on coffee quality using a H-1 NMR-based metabolomics approach. Food Res. Int. 2015, 67, 175–182. [Google Scholar] [CrossRef]
- Farghal, H.H.; Mansour, S.T.; Khattab, S.; Zhao, C.; Farag, M.A. A comprehensive insight on modern green analyses for quality control determination and processing monitoring in coffee and cocoa seeds. Food Chem. 2022, 394, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Lim, L.T.; Fu, Y.C. Review of Analytical Methods to Detect Adulteration in Coffee. J. AOAC Int. 2020, 103, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Bressanello, D.; Liberto, E.; Cordero, C.; Sgorbini, B.; Rubiolo, P.; Pellegrino, G.; Ruosi, M.R.; Bicchi, C. Chemometric Modeling of Coffee Sensory Notes through Their Chemical Signatures: Potential and Limits in Defining an Analytical Tool for Quality Control. J. Agric. Food Chem. 2018, 66, 7096–7109. [Google Scholar] [CrossRef] [PubMed]
- Yeager, S.E.; Batali, M.E.; Guinard, J.X.; Ristenpart, W.D. Acids in coffee: A review of sensory measurements and meta-analysis of chemical composition. Crit. Rev. Food Sci. Nutr. 2023, 63, 1010–1036. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, N.; Whitworth, M.B.; Grebby, S.; Fisk, I.D. Rapid prediction of single green coffee bean moisture and lipid content by hyperspectral imaging. J. Food Eng. 2018, 227, 18–29. [Google Scholar] [CrossRef]
- Pazmino-Arteaga, J.; Gallardo, C.; Gonzalez-Rodriguez, T.; Winkler, R. Loss of Sensory Cup Quality: Physiological and Chemical Changes during Green Coffee Storage. Plant. Food Hum. Nutr. 2022, 77, 1–11. [Google Scholar] [CrossRef]
- Cotter, A.R.; Hopfer, H. The Effects of Storage Temperature on the Aroma of Whole Bean Arabica Coffee Evaluated by Coffee Consumers and HS-SPME-GC-MS. Beverages 2018, 4, 10. [Google Scholar] [CrossRef]
- Barbosa, M.D.G.; Scholz, M.B.D.; Kitzberger, C.S.G.; Benassi, M.D. Correlation between the composition of green Arabica coffee beans and the sensory quality of coffee brews. Food Chem. 2019, 292, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Babova, O.; Occhipinti, A.; Maffei, M.E. Chemical partitioning and antioxidant capacity of green coffee (Coffea arabica and Coffea canephora) of different geographical origin. Phytochemistry 2016, 123, 33–39. [Google Scholar] [CrossRef]
- Malta, M.R.; Fassio, L.d.O.; Liska, G.R.; Carvalho, G.R.; Pereira, A.A.; Botelho, C.E.; Ferraz, V.P.; Silva, A.D.; Pedrosa, A.W.; Alvaro, L.N.; et al. Discrimination of genotypes coffee by chemical composition of the beans: Potential markers in natural coffees. Food Res. Int. 2020, 134, 109219. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.T.; Long, Y.; Ma, Y.J.; Chen, Y.; Yu, Q.; Xie, J.H.; Li, B.; Tian, J.L. Comparison of chemical and fatty acid composition of green coffee bean (Coffea arabica L.) from different geographical origins. LWT 2021, 140, 9. [Google Scholar] [CrossRef]
- Cai, Z.H.; Li, P.; Dong, T.T.X.; Tsim, K.W.K. Molecular diversity of 5S-rRNA spacer domain in Fritillaria species revealed by PCR analysis. Planta Med. 1999, 65, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Rubiolo, P.; Matteodo, M.; Bicchi, C.; Appendino, G.; Gnavi, G.; Bertea, C.M.; Maffei, M.E. Chemical and Biomolecular Characterization of Artemisia umbelliformis Lam., an Important Ingredient of the Alpine Liqueur “Genepi”. J. Agric. Food Chem. 2009, 57, 3436–3443. [Google Scholar] [CrossRef] [PubMed]
- Gnavi, G.; Bertea, C.M.; Maffei, M.E. PCR, sequencing and PCR-RFLP of the 5S-rRNA-NTS region as a tool for the DNA fingerprinting of medicinal and aromatic plants. Flavour. Frag. J. 2010, 25, 132–137. [Google Scholar] [CrossRef]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.H.; Zhang, Y.; Zhou, S.L. Barcoding the kingdom Plantae: New PCR primers for ITS regions of plants with improved universality and specificity. Mol. Ecol. Resour. 2016, 16, 138–149. [Google Scholar] [CrossRef]
- Mannino, G.; Gentile, C.; Maffei, M.E. Chemical partitioning and DNA fingerprinting of some pistachio (Pistacia vera L.) varieties of different geographical origin. Phytochemistry 2019, 160, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Petry, R.D.; Ortega, G.G.; Silva, W.B. Flavonoid content assay: Influence of the reagent concentration and reaction time on the spectrophotometric behavior of the aluminium chloride--flavonoid complex. Die Pharm. 2001, 56, 465–470. [Google Scholar]
- Miliauskas, G.; Venskutonis, P.R.; van Beek, T.A. Screening of radical scavenging activity of some medicinal and aromatic plant extracts. Food Chem. 2004, 85, 231–237. [Google Scholar] [CrossRef]
- Prior, R.L.; Fan, E.; Ji, H.; Howell, A.; Nio, C.; Payne, M.J.; Reed, J. Multi-laboratory validation of a standard method for quantifying proanthocyanidins in cranberry powders. J. Sci. Food Agric. 2010, 90, 1473–1478. [Google Scholar] [CrossRef]
- George, J.; Edwards, D.; Pun, S.; Williams, D. Evaluation of Antioxidant Capacity (ABTS and CUPRAC) and Total Phenolic Content (Folin-Ciocalteu) Assays of Selected Fruit, Vegetables, and Spices. Int. J. Food Sci. 2022, 2022, 2581470. [Google Scholar] [CrossRef]
- Svečnjak, L.; Marijanović, Z.; Okińczyc, P.; Marek Kuś, P.; Jerković, I. Mediterranean Propolis from the Adriatic Sea Islands as a Source of Natural Antioxidants: Comprehensive Chemical Biodiversity Determined by GC-MS, FTIR-ATR, UHPLC-DAD-QqTOF-MS, DPPH and FRAP Assay. Antioxidants 2020, 9, 337. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Maffei, M.E. Metabolomics-Based Profiling, Antioxidant Power, and Uropathogenic Bacterial Anti-Adhesion Activity of SP4(TM), a Formulation with a High Content of Type-A Proanthocyanidins. Antioxidants 2022, 11, 1234. [Google Scholar] [CrossRef] [PubMed]
- Mannino, G.; Casacci, L.P.; Bianco Dolino, G.; Badolato, G.; Maffei, M.E.; Barbero, F. The Geomagnetic Field (GMF) Is Necessary for Black Garden Ant (Lasius niger L.) Foraging and Modulates Orientation Potentially through Aminergic Regulation and MagR Expression. Int. J. Mol. Sci. 2023, 24, 4387. [Google Scholar] [CrossRef] [PubMed]
- Vigliante, I.; Mannino, G.; Maffei, M.E. Chemical Characterization and DNA Fingerprinting of Griffonia simplicifolia Baill. Molecules 2019, 24, 9. [Google Scholar] [CrossRef]
- Capuzzo, A.; Maffei, M.E. Molecular fingerprinting of peppermint (Mentha piperita) and some Mentha hybrids by sequencing and RFLP analysis of the 5S rRNA Non-Transcribed Spacer (NTS) region. Plant. Biosyst. 2016, 150, 236–243. [Google Scholar] [CrossRef]
- Mehari, B.; Chandravanshi, B.S.; Redi-Abshiro, M.; Combrinck, S.; McCrindle, R.; Atlabachew, M. Polyphenol contents of green coffee beans from different regions of Ethiopia. Int. J. Food Prop. 2021, 24, 17–27. [Google Scholar] [CrossRef]
- Bobková, A.; Hudáček, M.; Jakabová, S.; Belej, Ľ.; Capcarová, M.; Čurlej, J.; Bobko, M.; Árvay, J.; Jakab, I.; Čapla, J.; et al. The effect of roasting on the total polyphenols and antioxidant activity of coffee. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2020, 55, 495–500. [Google Scholar] [CrossRef]
- Sun, M.F.; Jiang, C.L.; Kong, Y.S.; Luo, J.L.; Yin, P.; Guo, G.Y. Recent Advances in Analytical Methods for Determination of Polyphenols in Tea: A Comprehensive Review. Foods 2022, 11, 1425. [Google Scholar] [CrossRef]
- González-de-Peredo, A.V.; Vázquez-Espinosa, M.; Espada-Bellido, E.; Carrera, C.; Ferreiro-González, M.; Barbero, G.F.; Palma, M. Flavonol Composition and Antioxidant Activity of Onions (Allium cepa L.) Based on the Development of New Analytical Ultrasound-Assisted Extraction Methods. Antioxidants 2021, 10, 273. [Google Scholar] [CrossRef]
- Birmingham, A.D.; Esquivel-Alvarado, D.; Maranan, M.; Krueger, C.G.; Reed, J.D. Inter-Laboratory Validation of 4-(Dimethylamino) Cinnamaldehyde (DMAC) Assay Using Cranberry Proanthocyanidin Standard for Quantification of Soluble Proanthocyanidins in Cranberry Foods and Dietary Supplements, First Action Official MethodSM: 2019.06. J. AOAC Int. 2021, 104, 216–222. [Google Scholar] [CrossRef]
- Lopes, C.R.; Shepherd, G.P. phylogenetic Studies of Some Species of the Genus Coffea. 1. Numerical-Analysis of Flavonoid Compounds. Rev. Bras. Genética 1991, 14, 425–435. [Google Scholar]
- Haile, M.; Kang, W.H. Antioxidant Activity, Total Polyphenol, Flavonoid and Tannin Contents of Fermented Green Coffee Beans with Selected Yeasts. Fermentation 2019, 5, 29. [Google Scholar] [CrossRef]
- Giomo, G.S.; Nakagawa, J.; Gallo, P.B. Beneficiamento de sementes de café e efeitos na qualidade física. Bragantia 2008, 67, 997–1010. [Google Scholar] [CrossRef]
- Cilas, C.; Bouharmont, P. Genetic studies on several bean traits of Coffea canephora coffee in Cameroon. J. Sci. Food Agric. 2005, 85, 2369–2374. [Google Scholar] [CrossRef]
- Novaes, P.; Torres, P.B.; Cornu, T.A.; Lopes, J.d.C.; Ferreira, M.J.P.; Santos, D.Y.A.C.d. Comparing antioxidant activities of flavonols from Annona coriacea by four approaches. S. Afr. J. Bot. 2019, 123, 253–258. [Google Scholar] [CrossRef]
- De Colmenares, N.G.; Ramírez-Martínez, J.R.; Aldana, J.O.; Clifford, M.N. Analysis of proanthocyanidins in coffee pulp. J. Sci. Food Agric. 1994, 65, 157–162. [Google Scholar] [CrossRef]
- de Colmenares, N.G.; Ramirez-Martinez, J.R.; Aldana, J.O.; Ramos-Nino, M.E.; Clifford, M.N.; Pekerar, S.; Mendez, B. Isolation, characterisation and determination of biological activity of coffee proanthocyanidins. J. Sci. Food Agric. 1998, 77, 368–372. [Google Scholar] [CrossRef]
- Iqbal, Y.; Ponnampalam, E.N.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. Extraction and characterization of polyphenols from non-conventional edible plants and their antioxidant activities. Food Res. Int. 2022, 157, 111205. [Google Scholar] [CrossRef]
- Awwad, S.; Issa, R.; Alnsour, L.; Albals, D.; Al-Momani, I. Quantification of Caffeine and Chlorogenic Acid in Green and Roasted Coffee Samples Using HPLC-DAD and Evaluation of the Effect of Degree of Roasting on Their Levels. Molecules 2021, 26, 7502. [Google Scholar] [CrossRef] [PubMed]
- Kulapichitr, F.; Borompichaichartkul, C.; Fang, M.; Suppavorasatit, I.; Cadwallader, K.R. Effect of post-harvest drying process on chlorogenic acids, antioxidant activities and CIE-Lab color of Thai Arabica green coffee beans. Food Chem. 2022, 366, 130504. [Google Scholar] [CrossRef]
- Rojas-González, A.; Figueroa-Hernández, C.Y.; González-Rios, O.; Suárez-Quiroz, M.L.; González-Amaro, R.M.; Hernández-Estrada, Z.J.; Rayas-Duarte, P. Coffee Chlorogenic Acids Incorporation for Bioactivity Enhancement of Foods: A Review. Molecules 2022, 27, 3400. [Google Scholar] [CrossRef] [PubMed]
- Iwashina, T. The structure and distribution of the flavonoids in plants. J. Plant. Res. 2000, 113, 287–299. [Google Scholar] [CrossRef]
- Mustafa, A.M.; Abouelenein, D.; Angeloni, S.; Maggi, F.; Navarini, L.; Sagratini, G.; Santanatoglia, A.; Torregiani, E.; Vittori, S.; Caprioli, G. A New HPLC-MS/MS Method for the Simultaneous Determination of Quercetin and Its Derivatives in Green Coffee Beans. Foods 2022, 11, 3033. [Google Scholar] [CrossRef]
- Liu, G.F.; Hu, Q.; Zhang, X.; Jiang, J.F.; Zhang, Y.; Zhang, Z.X. Melatonin biosynthesis and signal transduction in plants in response to environmental conditions. J. Exp. Bot. 2022, 73, 5818–5827. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Salces, R.M.; Serra, F.; Reniero, F.; Heberger, K. Botanical and Geographical Characterization of Green Coffee (Coffea arabica and Coffea canephora): Chemometric Evaluation of Phenolic and Methylxanthine Contents. J. Agric. Food Chem. 2009, 57, 4224–4235. [Google Scholar] [CrossRef]
- Correia, A.; Leitao, M.C.A.; Clifford, M.N. Caffeoyl-tyrosine and Angola-II as characteristic markers for angolan robusta coffee. Food Chem. 1995, 53, 309–313. [Google Scholar] [CrossRef]
- Zhao, E.H.; Ergul, B.; Zhao, W. Caffeine’s Antioxidant Potency Optically Sensed with Double-Stranded DNA-Encased Single-Walled Carbon Nanotubes. J. Phys. Chem. B 2015, 119, 4068–4075. [Google Scholar] [CrossRef]
- Tsoi, B.; Yi, R.N.; Cao, L.F.; Li, S.B.; Tan, R.R.; Chen, M.; Li, X.X.; Wang, C.; Li, Y.F.; Kurihara, H.; et al. Comparing antioxidant capacity of purine alkaloids: A new, efficient trio for screening and discovering potential antioxidants in vitro and in vivo. Food Chem. 2015, 176, 411–419. [Google Scholar] [CrossRef]
- Ramakrishna, A.; Giridhar, P.; Sankar, K.U.; Ravishankar, G.A. Melatonin and serotonin profiles in beans of Coffea species. J. Pineal Res. 2012, 52, 470–476. [Google Scholar] [CrossRef]
- Jeszka-Skowron, M.; Frankowski, R.; Zgoła-Grześkowiak, A. Comparison of methylxantines, trigonelline, nicotinic acid and nicotinamide contents in brews of green and processed Arabica and Robusta coffee beans—Influence of steaming, decaffeination and roasting processes on coffee beans. LWT 2020, 125, 109344. [Google Scholar] [CrossRef]
- Farah, A.; de Paula Lima, J. Consumption of Chlorogenic Acids through Coffee and Health Implications. Beverages 2019, 5, 11. [Google Scholar] [CrossRef]
- Tajik, N.; Tajik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017, 56, 2215–2244. [Google Scholar] [CrossRef] [PubMed]
- Mohammad Salamatullah, A.; Hayat, K.; Mabood Husain, F.; Asif Ahmed, M.; Arzoo, S.; Mohammed Alghunaymi, A.; Alzahrani, A.; Alyahya, H.K.; Al-Badr, N.; Bourhia, M. Effect of Microwave Roasting and Extraction Solvents on the Bioactive Properties of Coffee Beans. Evid.-Based Complement. Altern. Med. eCAM 2021, 2021, 4908033. [Google Scholar] [CrossRef]
- Król, K.; Gantner, M.; Tatarak, A.; Hallmann, E. The content of polyphenols in coffee beans as roasting, origin and storage effect. Eur. Food Res. Technol. 2020, 246, 33–39. [Google Scholar] [CrossRef]
- Elgadi, S.; Ouhammou, A.; Taous, F.; Zine, H.; Papazoglou, E.G.; Elghali, T.; Amenzou, N.; El Allali, H.; Aitlhaj, A.; El Antari, A. Combination of Stable Isotopes and Fatty Acid Composition for Geographical Origin Discrimination of One Argan Oil Vintage. Foods 2021, 10, 1274. [Google Scholar] [CrossRef]
- Maffei, M.; Scannerini, S. Fatty acid variability from nonpolar lipids in some Lamiaceae. Biochem. Syst. Ecol. 1993, 21, 475–486. [Google Scholar] [CrossRef]
- Zhang, X.-J.; Qiu, J.-F.; Guo, L.-P.; Wang, Y.; Li, P.; Yang, F.-Q.; Su, H.; Wan, J.-B. Discrimination of Multi-Origin Chinese Herbal Medicines Using Gas Chromatography-Mass Spectrometry-Based Fatty Acid Profiling. Molecules 2013, 18, 15329–15343. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, D.; Laffargue, A.; Posada, H.; Bertrand, B.; Lashermes, P.; Dussert, S. Genotypic and environmental effects on coffee (Coffea arabica L.) bean fatty acid profile: Impact on variety and origin chemometric determination. J. Agric. Food Chem. 2009, 57, 11321–11327. [Google Scholar] [CrossRef]
- Bertrand, B.; Villarreal, D.; Laffargue, A.; Posada, H.; Lashermes, P.; Dussert, S. Comparison of the effectiveness of fatty acids, chlorogenic acids, and elements for the chemometric discrimination of coffee (Coffea arabica L.) varieties and growing origins. J. Agric. Food Chem. 2008, 56, 2273–2280. [Google Scholar] [CrossRef]
- Mendes, G.d.A.; de Oliveira, M.A.L.; Rodarte, M.P.; de Carvalho dos Anjos, V.; Bell, M.J.V. Origin geographical classification of green coffee beans (Coffea arabica L.) produced in different regions of the Minas Gerais state by FT-MIR and chemometric. Curr. Res. Food Sci. 2022, 5, 298–305. [Google Scholar] [CrossRef]
- Bessada, S.M.F.; Alves, R.C.; Costa, A.S.G.; Nunes, M.A.; Oliveira, M. Coffea canephora silverskin from different geographical origins: A comparative study. Sci. Total Environ. 2018, 645, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Hao, d.C.; Huang, B.L.; Chen, S.L.; Mu, J. Evolution of the chloroplast trnL-trnF region in the gymnosperm lineages Taxaceae and Cephalotaxaceae. Biochem. Genet. 2009, 47, 351–369. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Gielly, L.; Miquel, C.; Valentini, A.; Vermat, T.; Corthier, G.; Brochmann, C.; Willerslev, E. Power and limitations of the chloroplast trnL (UAA) intron for plant DNA barcoding. Nucleic Acids Res. 2007, 35, e14. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.R.; Carlson, J.E. Molecular cytogenetics of the genes encoding 18s-5.8s-26s rRNA and 5s rRNA in two species of spruce (Picea). Theor. Appl. Genet. 1997, 95, 1–9. [Google Scholar] [CrossRef]
- Gnavi, G.; Bertea, C.M.; Usai, M.; Maffei, M.E. Comparative characterization of Santolina insularis chemotypes by essential oil composition, 5S-rRNA-NTS sequencing and EcoRV RFLP-PCR. Phytochemistry 2010, 71, 930–936. [Google Scholar] [CrossRef]
- Capuzzo, A.; Maffei, M.E. Molecular fingerprinting of some Mentha species by sequencing and RFLP analysis of the 5S-rRNA non-transcribed spacer region. Plant. Biosyst. 2014, 148, 683–690. [Google Scholar] [CrossRef]
- Bolsheva, N.L.; Melnikova, N.V.; Kirov, I.V.; Speranskaya, A.S.; Krinitsina, A.A.; Dmitriev, A.A.; Belenikin, M.S.; Krasnov, G.S.; Lakunina, V.A.; Snezhkina, A.V.; et al. Evolution of blue-flowered species of genus Linum based on high-throughput sequencing of ribosomal RNA genes. BMC Evol. Biol. 2017, 17, 14. [Google Scholar] [CrossRef]
- Simon, L.; Rabanal, F.A.; Dubos, T.; Oliver, C.; Lauber, D.; Poulet, A.; Vogt, A.; Mandlbauer, A.; Le Goff, S.; Sommer, A.; et al. Genetic and epigenetic variation in 5S ribosomal RNA genes reveals genome dynamics in Arabidopsis thaliana. Nucleic Acids Res. 2018, 46, 3019–3033. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Geographical Origin | Species | Cultivar | Commercial Name | Accession No | |
|---|---|---|---|---|---|---|
| CC1 | Uganda | Africa | C. canephora | Jolly Quartz | Robusta | 28337/1 |
| CC2 | Uganda | Africa | C. canephora | Jolly Quartz | Robusta | 28336/1 |
| CC3 | Vietnam | Southeast Asia | C. canephora | Vietnam clean | Robusta | FV2583 |
| CC4 | Vietnam | Southeast Asia | C. canephora | Vietnam unwashed | Robusta | FV2583 |
| CC5 | Brazil | South America | C. canephora | Moka Fine C | Caracol | FV1147 |
| CA1 | Kenya | Africa | C. arabica | Arabica low grade | Arabica | 6841A |
| CA2 | Kenya | Africa | C. arabica | Arabica AK3 | Arabica | AK-3 |
| CA3 | Kenya | Africa | C. arabica | Arabica AK2 | Arabica | AK-2 |
| CA4 | Brazil | South America | C. arabica | Arabica Natural Terraforte | Arabica | 401|CA0839 |
| CA5 | Peru | South America | C. arabica | Tinamous | Arabica | 6890 |
| CA6 | Peru | South America | C. arabica | Jacamar | Arabica | 6874 |
| CA7 | Colombia | South America | C. arabica | Pacorini Silocaf | Arabica | 240229/1 |
| CA8 | Ethiopia | Africa | C. arabica | Sidamo Grade 2 | Arabica | 1168 |
| CA9 | Mexico | Central America | C. arabica | PW ep | Arabica | 1190 |
| CA10 | Guatemala | Central America | C. arabica | HB ep | Arabica | 1191 |
| CA11 | Honduras | Central America | C. arabica | HG ep “Margay” | Arabica | 1195 |
| CA12 | Honduras | Central America | C. arabica | HG ep “Margay” | Arabica | 1195 |
| CA13 | Honduras | Central America | C. arabica | Catuahi, Caturra, Icatu | Arabica | 801 |
| CA14 | Honduras | Central America | C. arabica | HG ep “Margay” | Arabica | 1145 |
| CA15 | Honduras | Central America | C. arabica | HG ep “Margay” | Arabica | 1195 |
| Code | TFdC (mg RE·100 g−1) | TFlC (mg QE·100 g−1) | TPAC (mg PACE·100 g−1) |
|---|---|---|---|
| CC1 | 125.59 ± 10.09 a | 68.65 ± 4.25 bc | 2.72 ± 0.22 a |
| CC2 | 105.23 ± 12.99 b | 68.66 ± 9.38 bc | 2.33 ± 0.33 ab |
| CC3 | 121.87 ± 8.54 ab | 70.69 ± 4.71 ab | 2.06 ± 0.12 cd |
| CC4 | 83.26 ± 8.85 c | 81.12 ± 5.57 a | 2.05 ± 0.16 cd |
| CC5 | 74.92 ± 7.45 hi | 42.58 ± 4.82 i | 1.91 ± 0.36 cd |
| CA1 | 136.11 ± 18.43 a | 75.46 ± 5.19 ab | 1.92 ± 0.28 cd |
| CA2 | 33.58 ± 3.77 l | 49.13 ± 5.84 fg | 1.61 ± 0.15 e |
| CA3 | 38.83 ± 6.89 l | 47.46 ± 4.81 fg | 1.73 ± 0.35 de |
| CA4 | 42.87 ± 3.59 il | 52.95 ± 1.44 ef | 1.86 ± 0.29 cd |
| CA5 | 46.79 ± 4.82 hi | 64.79 ± 1.34 cd | 2.07 ± 0.28 cd |
| CA6 | 39.89 ± 5.55 i | 56.29 ± 7.14 de | 2.15 ± 0.31 cd |
| CA7 | 35.65 ± 1.87 i | 62.57 ± 8.29 cd | 2.33 ± 0.37 ab |
| CA8 | 59.81 ± 1.02 ef | 53.52 ± 9.94 ef | 2.05 ± 0.21 cd |
| CA9 | 31.23 ± 1.05 i | 64.76 ± 6.25 cd | 2.21 ± 0.29 ab |
| CA10 | 57.94 ± 5.74 gh | 44.23 ± 3.63 hi | 2.64 ± 0.15 a |
| CA11 | 78.01 ± 8.71 de | 60.57 ± 4.45 cd | 1.86 ± 0.28 cd |
| CA12 | 39.01 ± 4.59 cd | 57.61 ± 3.72 de | 1.75 ± 0.22 de |
| CA13 | 62.27 ± 6.47 l | 52.29 ± 5.34 ef | 1.91 ± 0.32 cd |
| CA14 | 73.84 ± 8.22 cd | 46.42 ± 7.48 gh | 1.91 ± 0.23 cd |
| CA15 | 79.79 ± 6.01 cd | 44.59 ± 7.11 hi | 1.73 ± 0.18 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mannino, G.; Kunz, R.; Maffei, M.E. Discrimination of Green Coffee (Coffea arabica and Coffea canephora) of Different Geographical Origin Based on Antioxidant Activity, High-Throughput Metabolomics, and DNA RFLP Fingerprinting. Antioxidants 2023, 12, 1135. https://doi.org/10.3390/antiox12051135
Mannino G, Kunz R, Maffei ME. Discrimination of Green Coffee (Coffea arabica and Coffea canephora) of Different Geographical Origin Based on Antioxidant Activity, High-Throughput Metabolomics, and DNA RFLP Fingerprinting. Antioxidants. 2023; 12(5):1135. https://doi.org/10.3390/antiox12051135
Chicago/Turabian StyleMannino, Giuseppe, Ronja Kunz, and Massimo E. Maffei. 2023. "Discrimination of Green Coffee (Coffea arabica and Coffea canephora) of Different Geographical Origin Based on Antioxidant Activity, High-Throughput Metabolomics, and DNA RFLP Fingerprinting" Antioxidants 12, no. 5: 1135. https://doi.org/10.3390/antiox12051135