
Strigolactone Alleviates the Adverse Effects of Salt Stress on Seed Germination in Cucumber by Enhancing Antioxidant Capacity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Determination of Germination Rate, Radicle Long, and Lateral Root Number of Seeds
2.3. Determination of MDA, H2O2, O2−, and Proline Contents
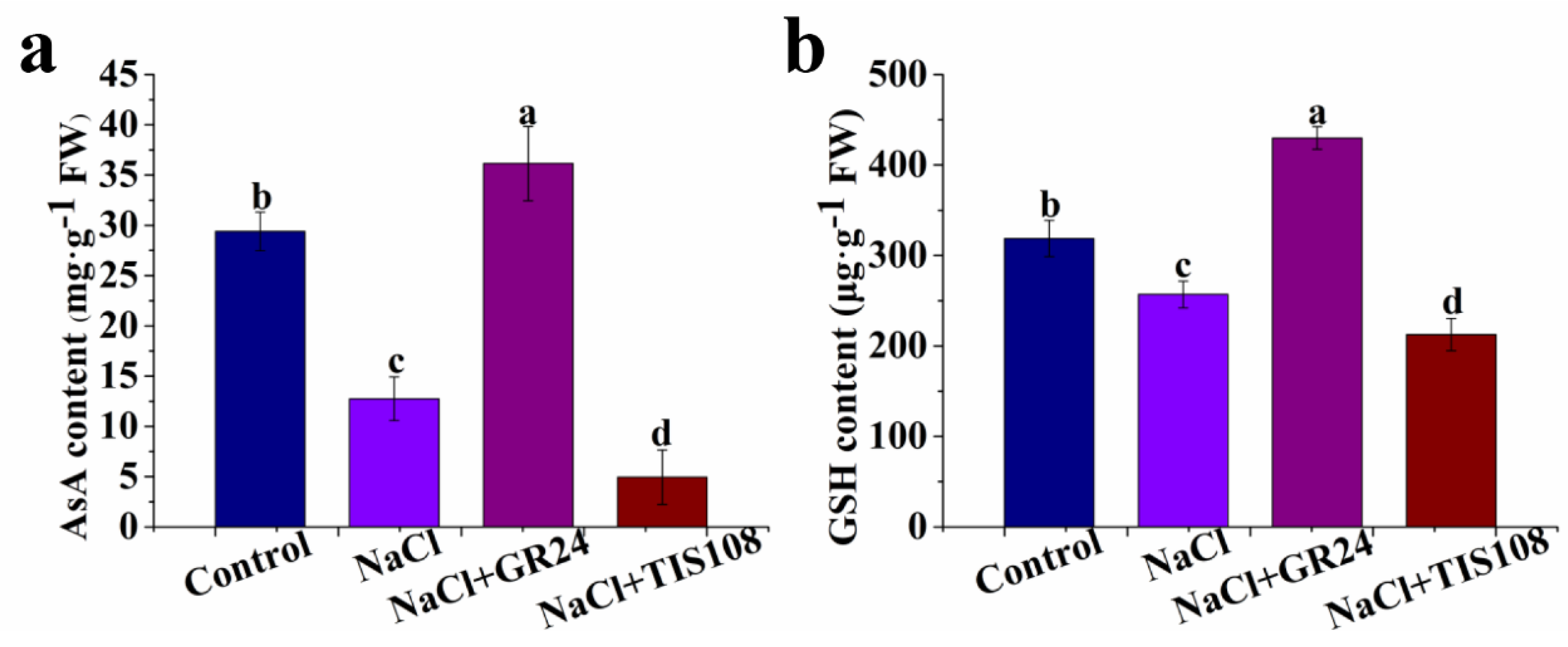
2.4. Determination of AsA and GSH Contents
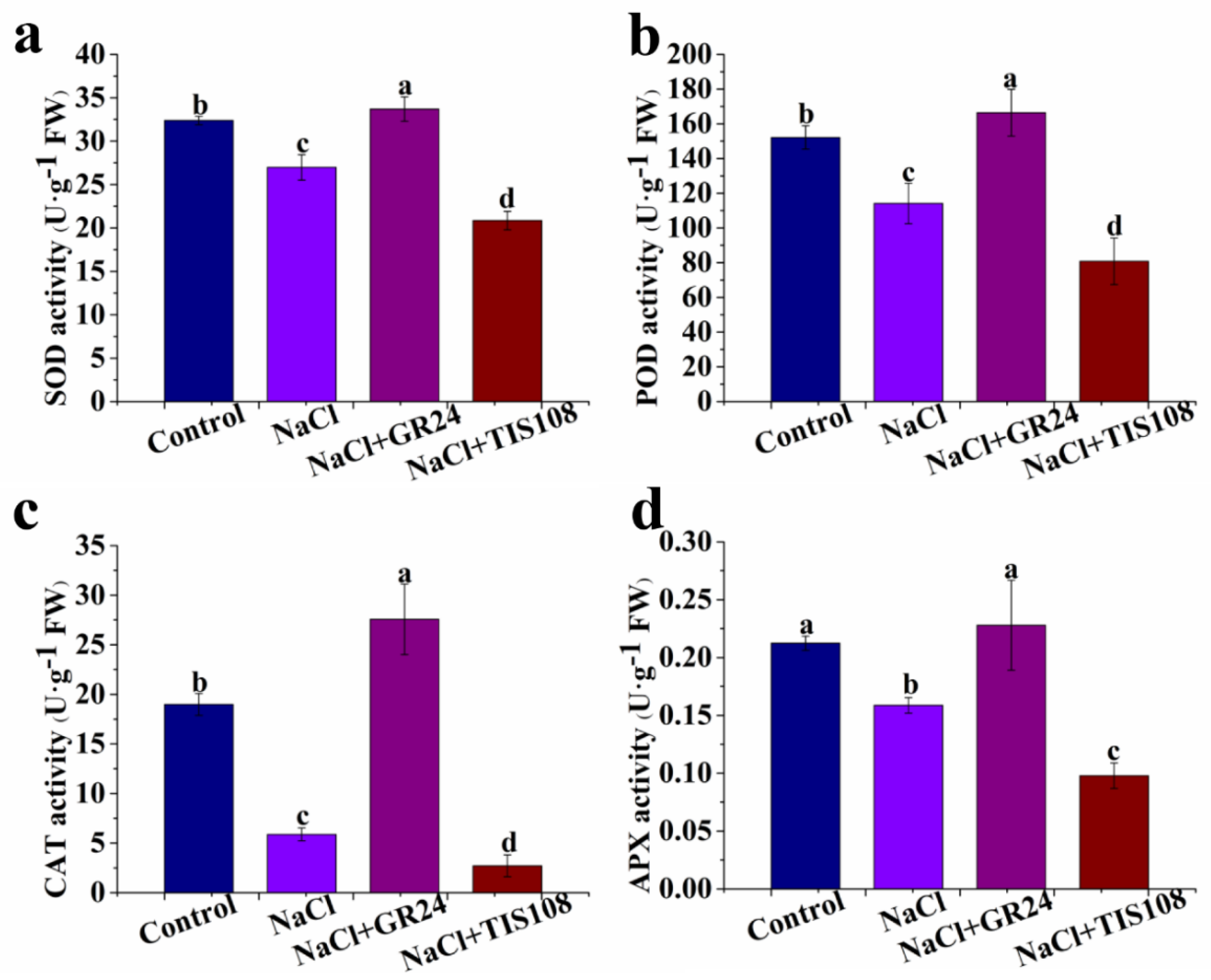
2.5. Determination of SOD, POD, CAT, and APX Activities
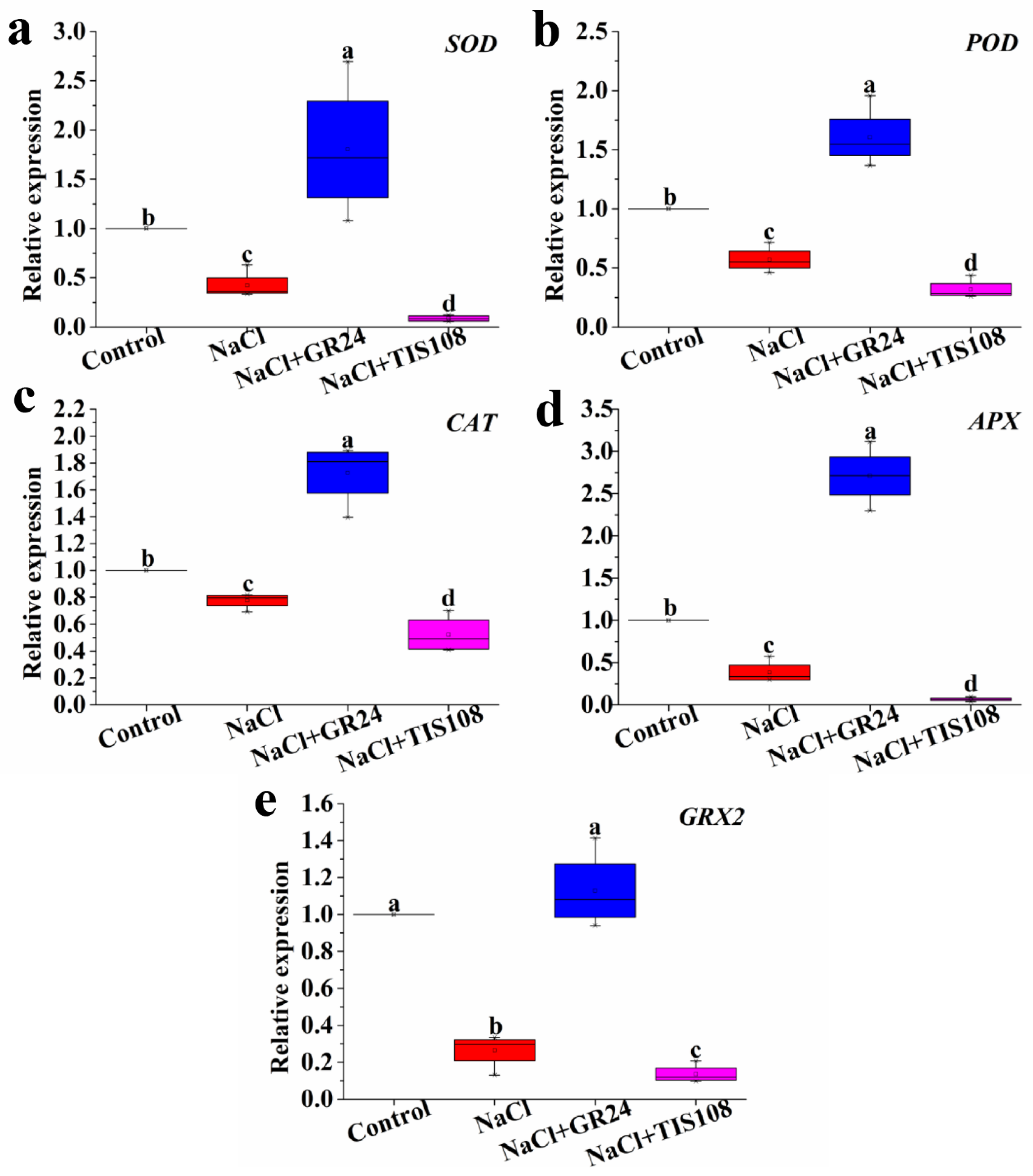
2.6. Total RNA Extraction and Gene Expression Analysis
2.7. Statistical Analysis
3. Results
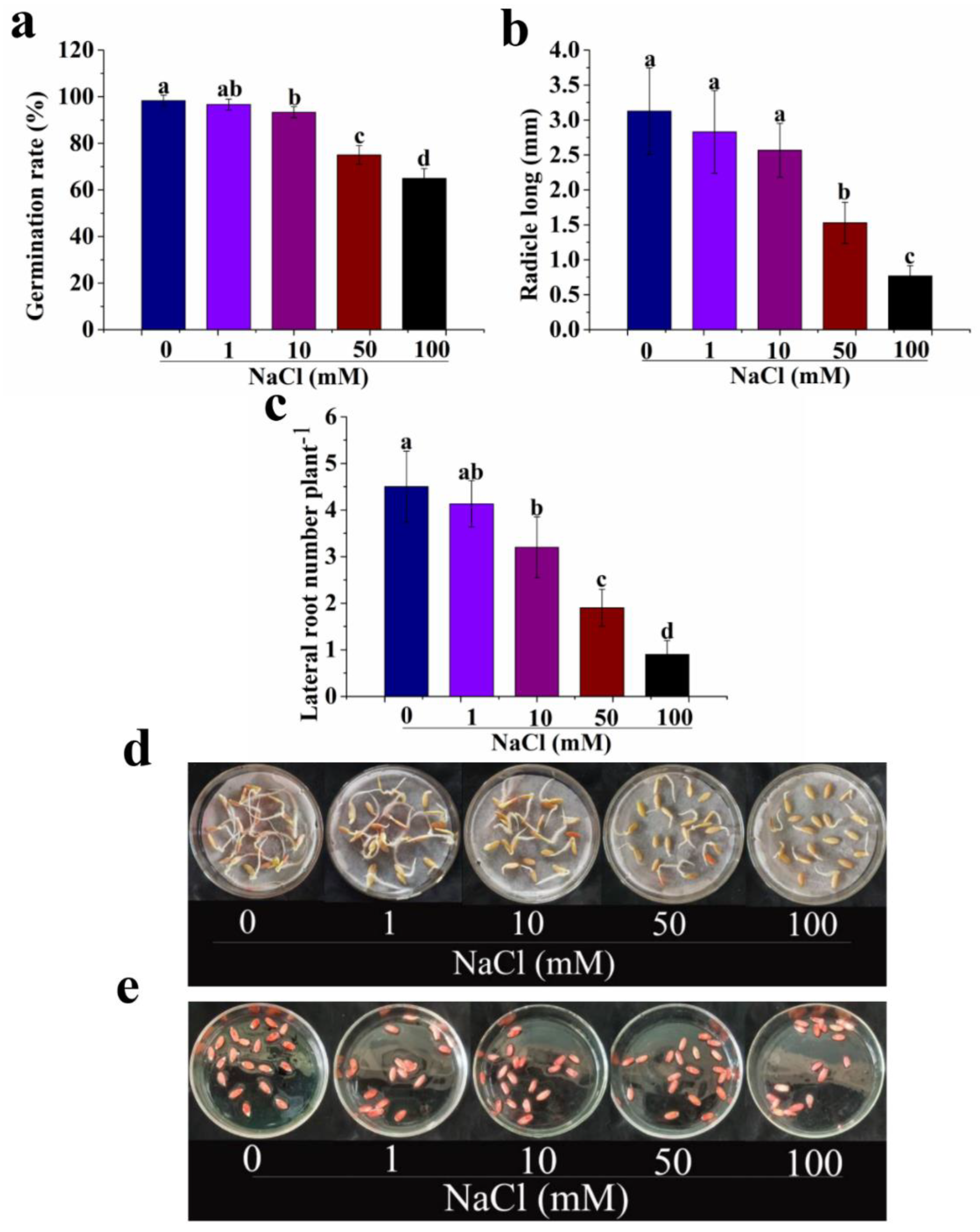
3.1. Salt Stress Inhibits Seed Germination in Cucumber
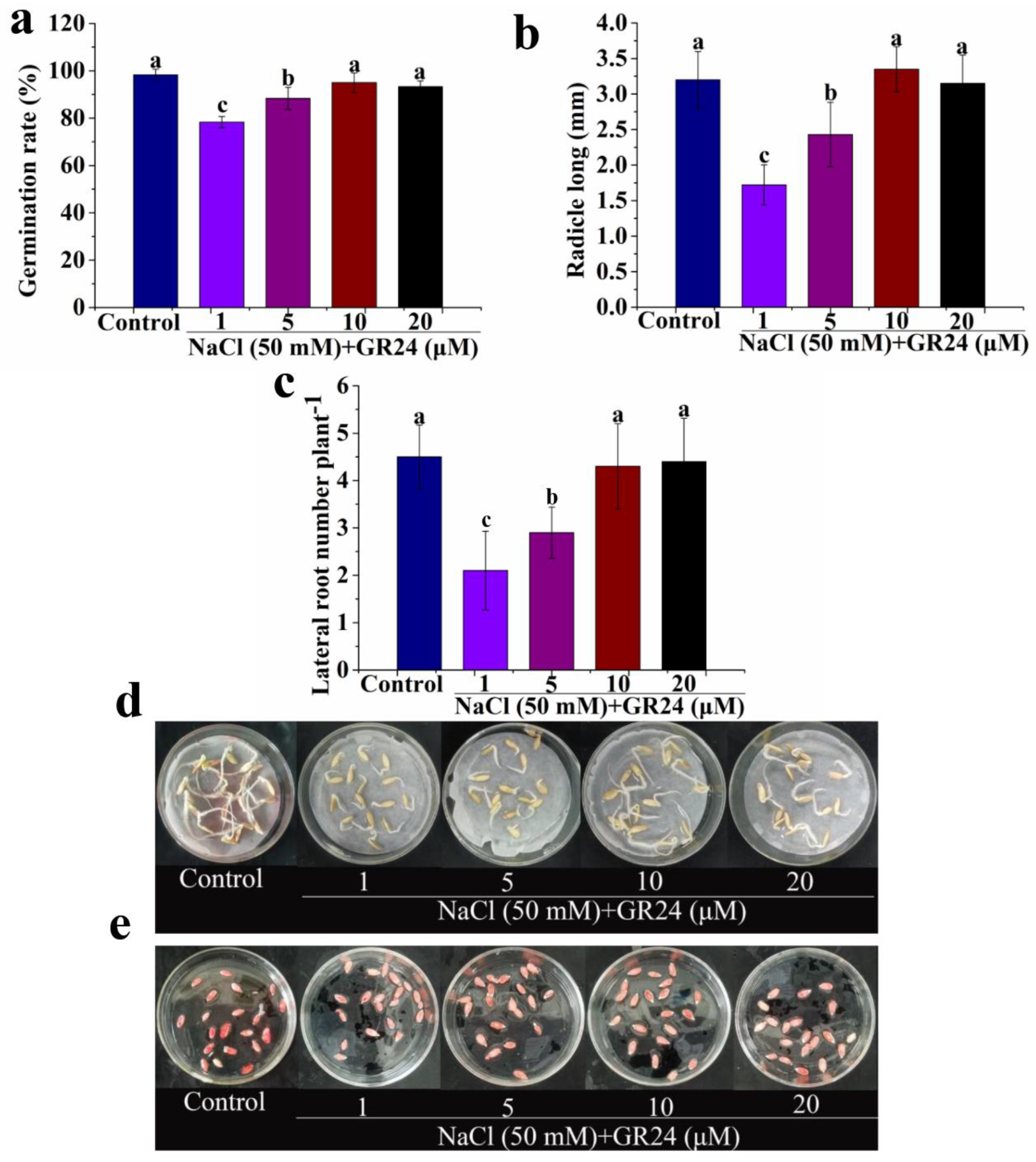
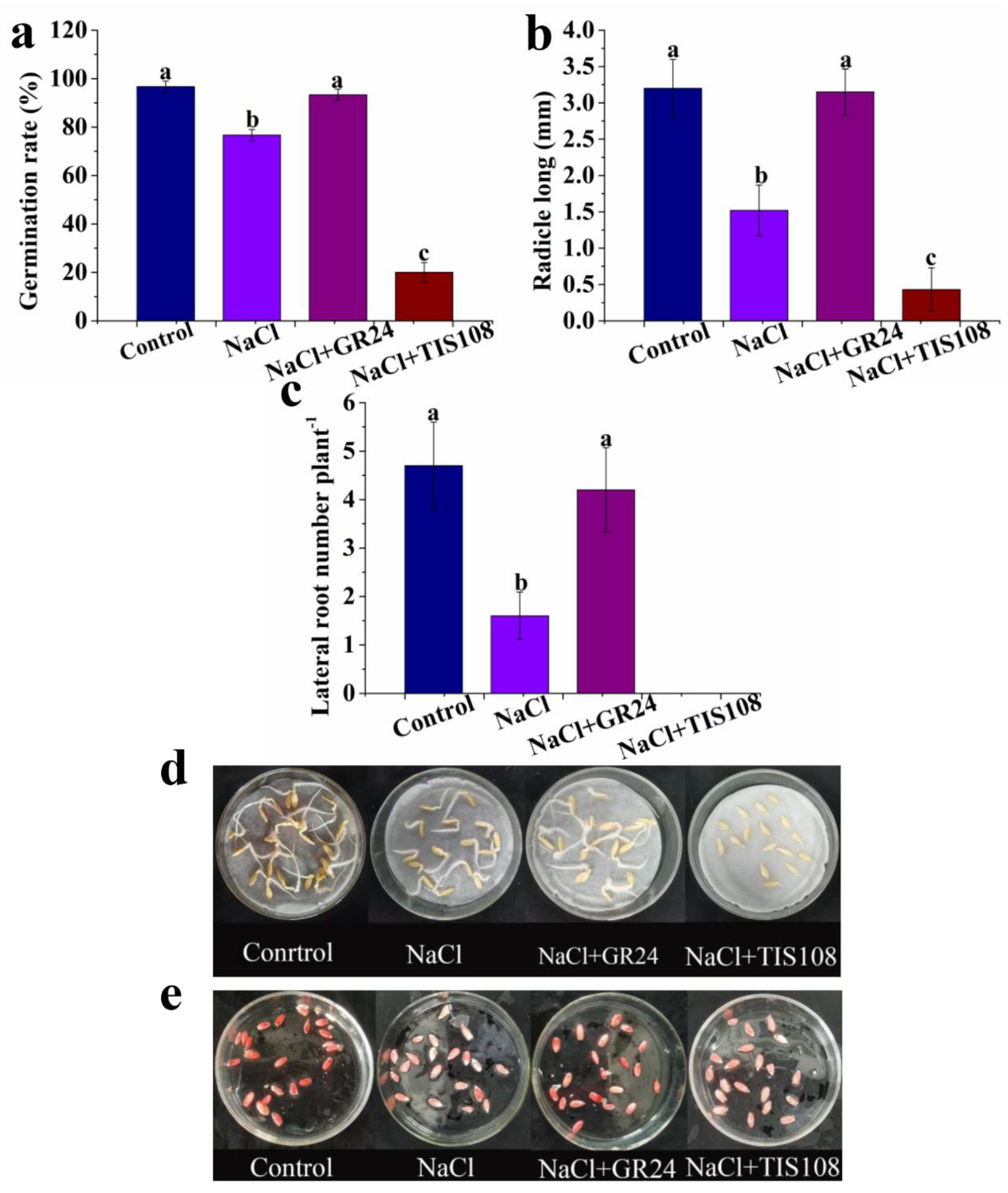
3.2. Effects of Exogenous GR24 on Seed Germination under Salt Stress
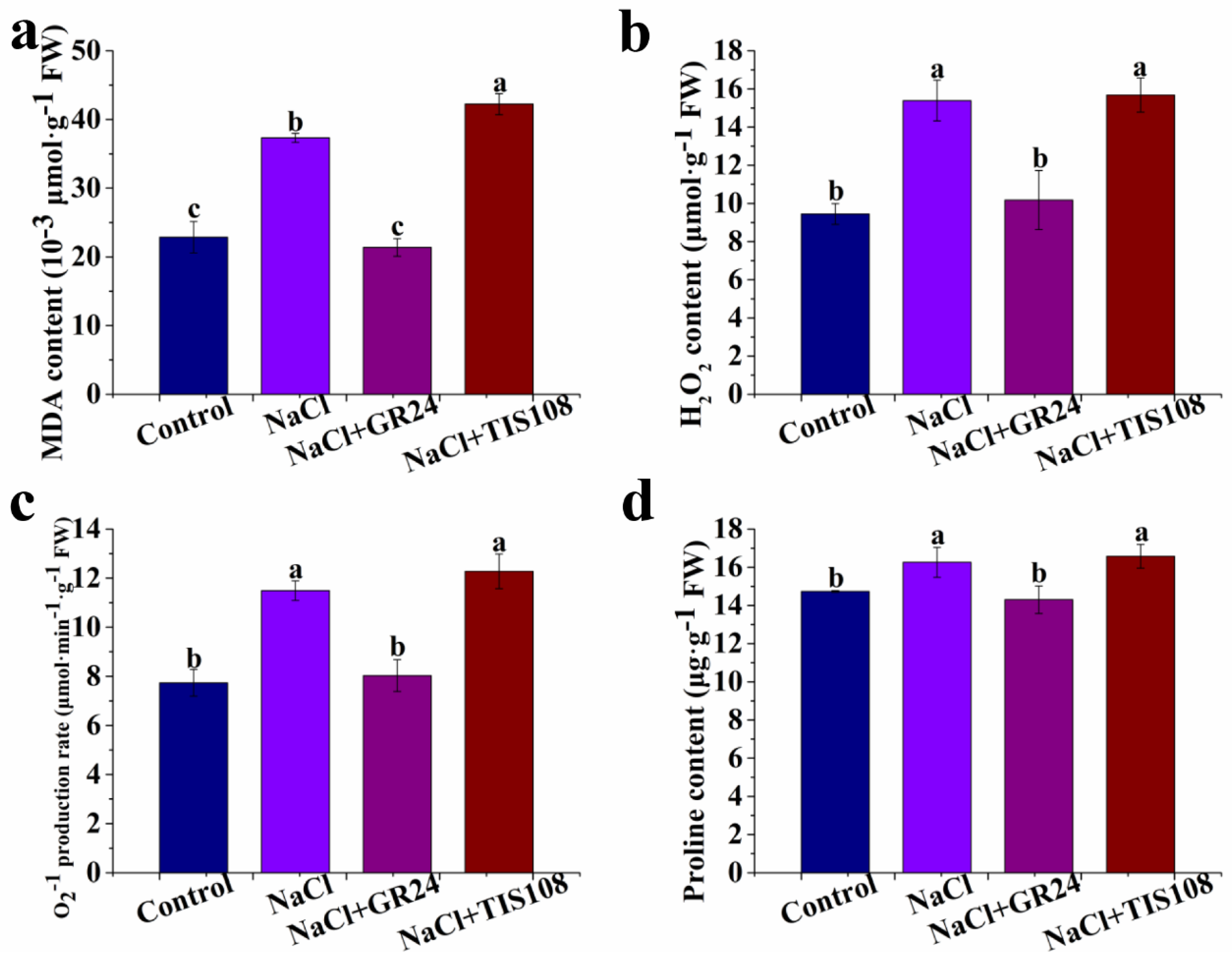
3.3. Effects of GR24 on MDA, H2O2, O2−, and Proline Contents during Seed Germination under Salt Stress
3.4. Effects of GR24 on AsA and GSH Contents during Seed Germination under Salt Stress
3.5. Effects of GR24 on SOD, POD, CAT, and APX Activities during Seed Germination under Salt Stress
3.6. Effects of GR24 on the Gene Expression Level Related to Antioxidant Enzymes during Seed Germination under Salt Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manivannan, P.; Jaleel, C.A.; Sankar, B.; Somasundaram, R.; Murali, P.V.; Sridharan, R.; Panneerselvam, R. Salt stress mitigation by calicium chloride in Vigna radiata (L.) Wilczek. Acta Biol. Cracov. Ser. Bot. 2007, 49, 105–109. [Google Scholar]
- Hussain, S.; Zhang, J.H.; Zhong, C.; Zhu, L.F.; Cao, X.C.; Yu, S.M.; Bohr, J.A.; Hu, J.J.; Jin, Q.Y. Effects of salt stress on rice growth, development characteristics, and the regulating ways: A review. J. Integr. Agric. 2017, 16, 2357–2374. [Google Scholar] [CrossRef]
- Sakr, M.T.; El-Metwally, M.A. Alleviation of the harmful effects of soil salt stress on growth, yield and endogenous antioxidant content of wheat plant by application of antioxidants. Pak. J. Biol. Sci. 2009, 12, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef]
- Hussain, S.; Khaliq, A.; Tanveer, M.; Matloob, A.; Hussain, H.A. Aspirin priming circumvents the salinity-induced effects on wheat emergence and seedling growth by regulating starch metabolism and antioxidant enzyme activities. Acta Physiol. Plant 2018, 40, 68. [Google Scholar] [CrossRef]
- Zhao, S.H.; Zhang, Q.K.; Liu, M.Y.; Zhou, H.P.; Ma, C.L.; Wang, P.P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Hao, S.H.; Wang, Y.R.; Yan, Y.X.; Liu, Y.H.; Wang, J.Y.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Wei, L.J.; Zhang, J.; Wei, S.H.; Hu, D.L.; Liu, Y.Y.; Feng, L.; Li, C.X.; Qi, N.N.; Wang, C.L.; Liao, W.B. Nitric oxide enhanced salt stress tolerance in tomato seedlings, involving phytohormone equilibrium and photosynthesis. Int. J. Mol. Sci. 2022, 23, 4539. [Google Scholar] [CrossRef]
- Li, C.X.; Yu, W.J.; Wu, Y.C.; Li, Y.Q. Roles of hydrogen gas in plants under abiotic stress: Current knowledge and perspectives. Antioxidants 2022, 11, 1999. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.C.; Liu, J.H.; Liu, T.; Xue, S.W. Hydrogen sulfide (H2S) signaling in plant development and stress responses. aBIOTECH 2021, 2, 32–63. [Google Scholar] [CrossRef]
- Liu, K.L.; Xu, S.; Xuan, W.; Ling, T.F.; Cao, Z.Y.; Huang, B.K.; Sun, Y.G.; Fang, L.; Liu, Z.Y.; Zhao, N.; et al. Carbon monoxide counteracts the inhibition of seed germination and alleviates oxidative damage caused by salt stress in Oryza sativa. Plant Sci. 2007, 172, 544–555. [Google Scholar] [CrossRef]
- Su, Q.F.; Zheng, X.D.; Tian, Y.K.; Wang, C.H. Exogenous brassinolide alleviates salt stress in Malus hupehensis ehd. by regulating the transcription of NHX-Type Na+ (K+)/H+ antiporters. Front. Plant Sci. 2020, 11, 38–54. [Google Scholar] [CrossRef]
- Barros Junior, U.O.; Lima, M.D.R.; Alsahli, A.A.; Lobato, A.K.S. Unraveling the roles of brassinosteroids in alleviating drought stress in young Eucalyptus urophylla plants: Implications on redox homeostasis and photosynthetic apparatus. Physiol. Plant 2021, 172, 748–761. [Google Scholar] [CrossRef]
- Marusig, D.; Tombesi, S. Abscisic acid mediates drought and salt stress responses in Vitis vinifera-a review. Int. J. Mol. Sci. 2020, 21, 8648. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Zhou, J.H.; Li, Y.X.; Quan, R.D.; Wang, J.; Huang, R.F.; Qin, H. Salt stress promotes abscisic acid accumulation to affect cell proliferation and expansion of primary roots in rice. Int. J. Mol. Sci. 2021, 22, 10892. [Google Scholar] [CrossRef]
- Yu, Z.P.; Duan, X.B.; Luo, L.; Dai, S.J.; Ding, Z.J.; Xia, G.M. How plant hormones mediate salt stress responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Khan, M.A. Tolerance and recovery responses of playa halophytes to light, salinity and temperature stresses during seed germination. Flora-Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 764–771. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, Y.; Ye, X.Y.; Li, Z.G. Signaling molecule hydrogen sulfide improves seed germination and seedling growth of maize (Zea mays L.) under high temperature by inducing antioxidant system and osmolyte biosynthesis. Front. Plant Sci. 2018, 9, 1288. [Google Scholar] [CrossRef]
- Cheng, M.J.; Guo, Y.L.; Liu, Q.; Nan, S.W.; Xue, Y.X.; Wei, C.H.; Zhang, Y.; Luan, F.S.; Zhang, X.; Li, H. H2O2 and Ca2+ signaling crosstalk counteracts ABA to induce seed germination. Antioxidants 2022, 11, 1594. [Google Scholar] [CrossRef]
- Pandey, S.; Kumari, A.; Shree, M.; Kumar, V.; Singh, P.; Bharadwaj, C.; Loake, G.J.; Parida, S.K.; Masakapalli, S.K.; Gupta, K.J. Nitric oxide accelerates germination via the regulation of respiration in chickpea. J. Exp. Bot. 2019, 70, 4539–4555. [Google Scholar] [CrossRef]
- Jia, Y.J.; Li, R.J.; Yang, W.J.; Chen, Z.; Hu, X.Y. Carbon monoxide signal regulates light-initiated seed germination by suppressing SOM expression. Plant Sci. 2018, 272, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.F.; Wang, W.; He, J.Y.; Zhang, L.Y.; Wei, Y.J.; Yang, M. Nitric oxide alleviates salt stress in seed germination and early seedling growth of pakchoi (Brassica chinensis L.) by enhancing physiological and biochemical parameters. Ecotoxicol. Environ. Saf. 2020, 187, 109785. [Google Scholar] [CrossRef] [PubMed]
- Vishal, B.; Kumar, P.P. Regulation of seed germination and abiotic stresses by gibberellins and abscisic acid. Front. Plant Sci. 2018, 9, 838. [Google Scholar] [CrossRef] [PubMed]
- Toh, S.; Kamiya, Y.; Kawakami, N.; Nambara, E.; McCourt, P.; Tsuchiya, Y. Thermoinhibition uncovers a role for strigolactones in Arabidopsis seed germination. Plant Cell Physiol. 2012, 53, 107–117. [Google Scholar] [CrossRef]
- Liu, H.W.; Li, C.X.; Yan, M.; Zhao, Z.X.; Huang, P.P.; Wei, L.J.; Wu, X.T.; Wang, C.L.; Liao, W.B. Strigolactone is involved in nitric oxide-enhanced the salt resistance in tomato seedlings. J. Plant Res. 2022, 135, 337–350. [Google Scholar] [CrossRef]
- Huang, P.; Li, C.; Liu, H.; Zhao, Z.; Liao, W. Hydrogen gas improves seed germination in cucumber by regulating sugar and starch metabolisms. Horticulturae 2021, 7, 456. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, B.; Xu, Z.; Shi, Z.; Chen, S.; Huang, X.; Chen, S.; Wang, X. Involvement of reactive oxygen species in endosperm cap weakening and embryo elongation growth during lettuce seed germination. J. Exp. Bot. 2014, 65, 3189–3200. [Google Scholar] [CrossRef]
- Ma, Q.; Zhang, T.; Zhang, P.; Wang, Z.Y. Melatonin attenuates postharvest physiological deterioration of cassava storage roots. J. Pineal. Res. 2016, 60, 424–434. [Google Scholar] [CrossRef]
- Prochazkova, D.; Sairam, R.K.; Srivastava, G.C. Oxidative stress and antioxidant activity as the basis of senescence in maize leaves. Plant. Sci. 2001, 161, 765–771. [Google Scholar] [CrossRef]
- Hu, H.; Liu, D.; Li, P. Methane delays the senescence and browning in daylily buds by restablished redox homeostasis. J. Sci. Food Agric. 2018, 98, 1977–1987. [Google Scholar] [CrossRef]
- Storey, R.; Jones, R.G.W. Betaine and choline levels in plants and their relationship to NaCl stress. Plant Sci. Lett. 1975, 4, 161–168. [Google Scholar] [CrossRef]
- Contreras-Calderón, J.; Calderón-Jaimes, L.; Guerra-Hernández, E.; García-Villanova, B. Antioxidant capacity, phenolic content and vitamin c in pulp, peel and seed from 24 exotic fruits from Colombia. Food Res. Int. 2011, 44, 2047–2053. [Google Scholar] [CrossRef]
- Zhao, Z.X.; Li, C.X.; Liu, H.W.; Yang, J.J.; Huang, P.P.; Liao, W. The involvement of glucose in hydrogen gas-medicated adventitious rooting in cucumber. Plants 2021, 10, 1937. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-11 CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- He, J.; Ren, Y.; Chen, X.; Chen, H. Protective roles of nitric oxide on seed germination and seedling growth of rice (Oryza sativa L.) under cadmium stress. Ecotoxicol. Environ. Saf. 2014, 108, 114–119. [Google Scholar] [CrossRef]
- Kumar, A.; Rodrigues, V.; Verma, S.; Singh, M.; Sundaresan, V. Effect of salt stress on seed germination, morphology, biochemical parameters, genomic template stability, and bioactive constituents of Andrographis paniculata Nees. Acta Physiol. Plant. 2021, 43, 68. [Google Scholar] [CrossRef]
- Kataria, S.; Baghel, L.; Guruprasad, K.N. Pre-treatment of seeds with static magnetic field improves germination and early growth characteristics under salt stress in maize and soybean. Biocatalysis Agric. Biotechnol. 2017, 10, 83–90. [Google Scholar] [CrossRef]
- Ali, Q.; Daud, M.K.; Haider, M.Z.; Ali, S.; Rizwan, M.; Aslam, N.; Noman, A.; Iqbal, N.; Shahzad, F.; Deeba, F.; et al. Seed priming by sodium nitroprusside improves salt tolerance in wheat (Triticum aestivum L.) by enhancing physiological and biochemical parameters. Plant Physiol. Bioch. 2017, 119, 50–58. [Google Scholar] [CrossRef]
- Li, Z.; Pei, X.; Yin, S.; Lang, X.; Zhao, X.; Qu, G.Z. Plant hormone treatments to alleviate the effects of salt stress on germination of Betula platyphylla seeds. J. For. Res. 2019, 30, 779–787. [Google Scholar] [CrossRef]
- Aroca, R.; Ruiz-Lozano, J.M.; Zamarreño, Á.M.; Paz, J.A.; García-Mina, J.M.; Pozo, M.J.; López-Ráez, J.A. Arbuscular mycorrhizal symbiosis influences strigolactone production under salinity and alleviates salt stress in lettuce plants. J. Plant Physiol. 2013, 170, 47–55. [Google Scholar] [CrossRef]
- Fu, X.J.; Wang, J.; Shangguan, T.W.; Wu, R.; Li, S.J.; Chen, G.; Xu, S.C. SMXLs regulate seed germination under salinity and drought stress in soybean. Plant Growth Regul. 2022, 96, 397–408. [Google Scholar] [CrossRef]
- Ling, F.L.; Su, Q.W.; Jiang, H.; Cui, J.J.; He, X.L.; Wu, Z.H.; Zhang, Z.A.; Liu, J.; Zhao, Y.J. Effects of strigolactone on photosynthetic and physiological characteristics in salt-stressed rice seedlings. Sci. Rep. 2020, 10, 6183. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Yu, H.J.; Li, Q.; Chai, L.; Jiang, W.J. Improving plant growth and alleviating photosynthetic inhibition and oxidative stress from low-light stress with exogenous GR24 in tomato (Solanum lycopersicum L.) seedlings. Front. Plant Sci. 2019, 10, 490. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.Q.; Bian, C.J.; Liu, W.J.; Sun, Z.J.; Xi, X.L.; Guo, D.M.; Liu, X.L.; Tian, Y.K.; Wang, C.H.; Zheng, X.D. Strigolactone alleviates the salinity-alkalinity stress of Malus hupehensis seedlings. Front. Plant Sci. 2022, 13, 901782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Zhang, L.; Ma, C.; Su, M.; Wang, J.; Zheng, S.; Zhang, T.G. Exogenous strigolactones alleviate the photosynthetic inhibition and oxidative damage of cucumber seedlings under salt stress. Sci. Hortic. 2022, 297, 110962. [Google Scholar] [CrossRef]
- Zhou, X.P.; Tan, Z.M.; Zhou, Y.G.; Guo, S.R.; Sang, T.; Wang, Y.; Shu, S. Physiological mechanism of strigolactone enhancing tolerance to low light stress in cucumber seedlings. BMC Plant Biol. 2022, 22, 30. [Google Scholar] [CrossRef]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotoxicol. Environ. Saf. 2017, 137, 64–70. [Google Scholar] [CrossRef]
- Yu, L.X.; Zhang, C.J.; Shang, H.Q.; Wang, X.F.; Wei, M.; Yang, F.J.; Shi, Q.H. Exogenous hydrogen sulfide enhanced antioxidant capacity, amylase activities and salt tolerance of cucumber hypocotyls and radicles. J. Integr. Agric. 2013, 12, 445–456. [Google Scholar] [CrossRef]
- Fan, H.F.; Du, C.X.; Ding, L.; Xu, Y.L. Effects of nitric oxide on the germination of cucumber seeds and antioxidant enzymes under salinity stress. Acta Physiol. Plant. 2013, 35, 2707–2719. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Alamri, S.A.; Al-Khaishany, M.Y.; Al-Qutami, M.A.; Ali, H.M.; AL-Rabiah, H.; Kalaji, H.M. Exogenous application of nitric oxide and spermidine reduces the negative effects of salt stress on tomato. Hortic. Environ. Biotechnol. 2017, 58, 537–547. [Google Scholar] [CrossRef]
- Ahmadi, F.I.; Karimi, K.; Struik, P.C. Effect of exogenous application of methyl jasmonate on physiological and biochemical characteristics of Brassica napus L. cv. Talaye under salinity stress. S. Afr. J. Bot. 2018, 115, 5–11. [Google Scholar] [CrossRef]
- Kumar, S.; Beena, A.S.; Awana, M.; Singh, A. Physiological, biochemical, epigenetic and molecular analyses of wheat (Triticum aestivum) genotypes with contrasting salt tolerance. Front. Plant Sci. 2017, 8, 1151. [Google Scholar] [CrossRef]
- Chawla, S.; Jain, S.; Jain, V. Salinity induced oxidative stress and antioxidant system in salt-tolerant and salt-sensitive cultivars of rice (Oryza sativa L.). J. Plant Biochem. Biotechnol. 2013, 22, 27–34. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front. Plant Sci. 2016, 7, 276. [Google Scholar] [CrossRef]
- Turan, M.; Ekinci, M.; Kul, R.; Boynueyri, F.G.; Yildirim, E. Mitigation of salinity stress in cucumber seedlings by exogenous hydrogen sulfide. J. Plant Res. 2022, 135, 517–529. [Google Scholar] [CrossRef]
- Raziq, A.; Wang, Y.; Din, A.; Sun, J.; Shu, S.; Guo, S.A. Comprehensive evaluation of salt tolerance in tomato (Var. Ailsa Craig): Responses of physiological and transcriptional changes in RBOH’s and ABA biosynthesis and signalling genes. Int. J. Mol. Sci. 2022, 23, 1603. [Google Scholar] [CrossRef]
- Zhang, R.M.; Dong, Y.L.; Li, Y.Y.; Ren, G.Y.; Chen, C.; Jin, X.X. SLs signal transduction gene CsMAX2 of cucumber positively regulated to salt, drought and ABA stress in Arabidopsis thaliana L. Gene 2023, 864, 147282. [Google Scholar] [CrossRef]
- Omoarelojie, L.O.; Kulkarni, M.G.; Finnie, J.F.; Pospíšil, T.; Strnad, M.; Van Staden, J. Synthetic strigolactone (rac-GR24) alleviates the adverse effects of heat stress on seed germination and photosystem II function in lupine seedlings. Plant Physiol. Biochem. 2020, 155, 965–979. [Google Scholar] [CrossRef]
- Zhou, Y.; Wen, Z.L.; Zhang, J.W.; Chen, X.J.; Cui, J.X.; Xu, W.; Liu, H.Y. Exogenous glutathione alleviates salt-induced oxidative stress in tomato seedlings by regulating glutathione metabolism, redox status, and the antioxidant system. Sci. Hortic. 2017, 220, 90–101. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Primer Sequence (5′–3′) |
|---|---|---|
| SOD | LOC101215417 | ACGGGTAATGTTTCTGGTCTCAAGC CATGCAGCCATTGGTTGTGTCAC |
| POD | LOC101212957 | AGACGGTGGTGGGAGTGGAATC GAGCAACGATGTCAGCACAAGAAAC |
| CAT | LOC101216662 | GAGCAACGATGTCAGCACAAGAAAC TCCAAGACGGTGCCTCTGAGTATC |
| APX | LOC101212957 | AAAGTGCTACCCTGTTGTGAGTGAG AAAGTGCTACCCTGTTGTGAGTGAG |
| GRX2 | LOC101205064 | TCGGTGGTTGCGATGGTAAGAAC GCTGACCTCTGTGATGCTTCTCTG |
| Actin | LOC101220617 | F: TTCTGGTGATGG TGTGAGTC R: GGCAGTGGTGGTGAACATG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Lu, X.; Liu, Y.; Xu, J.; Yu, W. Strigolactone Alleviates the Adverse Effects of Salt Stress on Seed Germination in Cucumber by Enhancing Antioxidant Capacity. Antioxidants 2023, 12, 1043. https://doi.org/10.3390/antiox12051043
Li C, Lu X, Liu Y, Xu J, Yu W. Strigolactone Alleviates the Adverse Effects of Salt Stress on Seed Germination in Cucumber by Enhancing Antioxidant Capacity. Antioxidants. 2023; 12(5):1043. https://doi.org/10.3390/antiox12051043
Chicago/Turabian StyleLi, Changxia, Xuefang Lu, Yunzhi Liu, Junrong Xu, and Wenjin Yu. 2023. "Strigolactone Alleviates the Adverse Effects of Salt Stress on Seed Germination in Cucumber by Enhancing Antioxidant Capacity" Antioxidants 12, no. 5: 1043. https://doi.org/10.3390/antiox12051043