
The Functional Relationship between NADPH Thioredoxin Reductase C, 2-Cys Peroxiredoxins, and m-Type Thioredoxins in the Regulation of Calvin–Benson Cycle and Malate-Valve Enzymes in Arabidopsis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
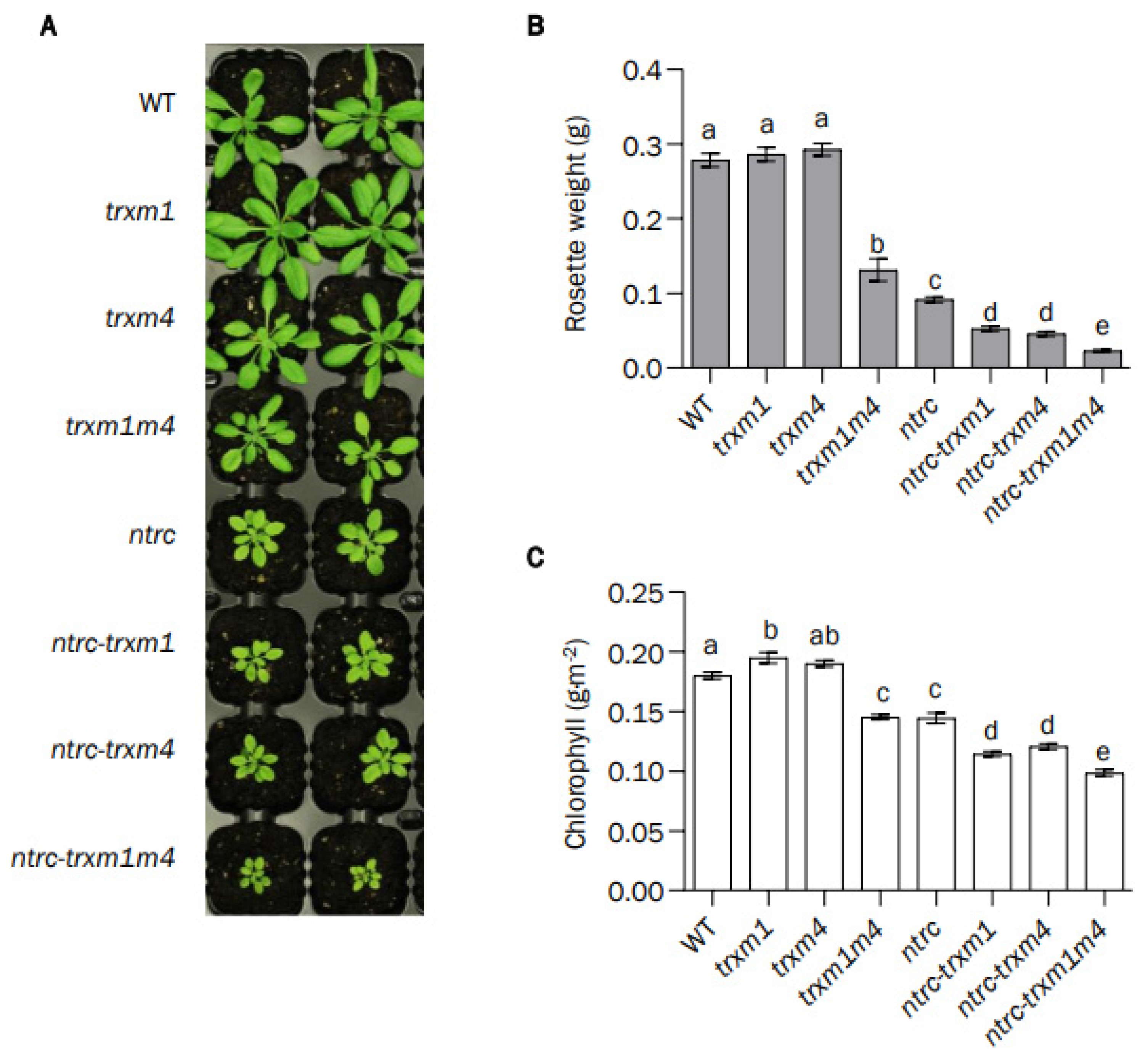
2.1. Biological Materials and Growth Conditions
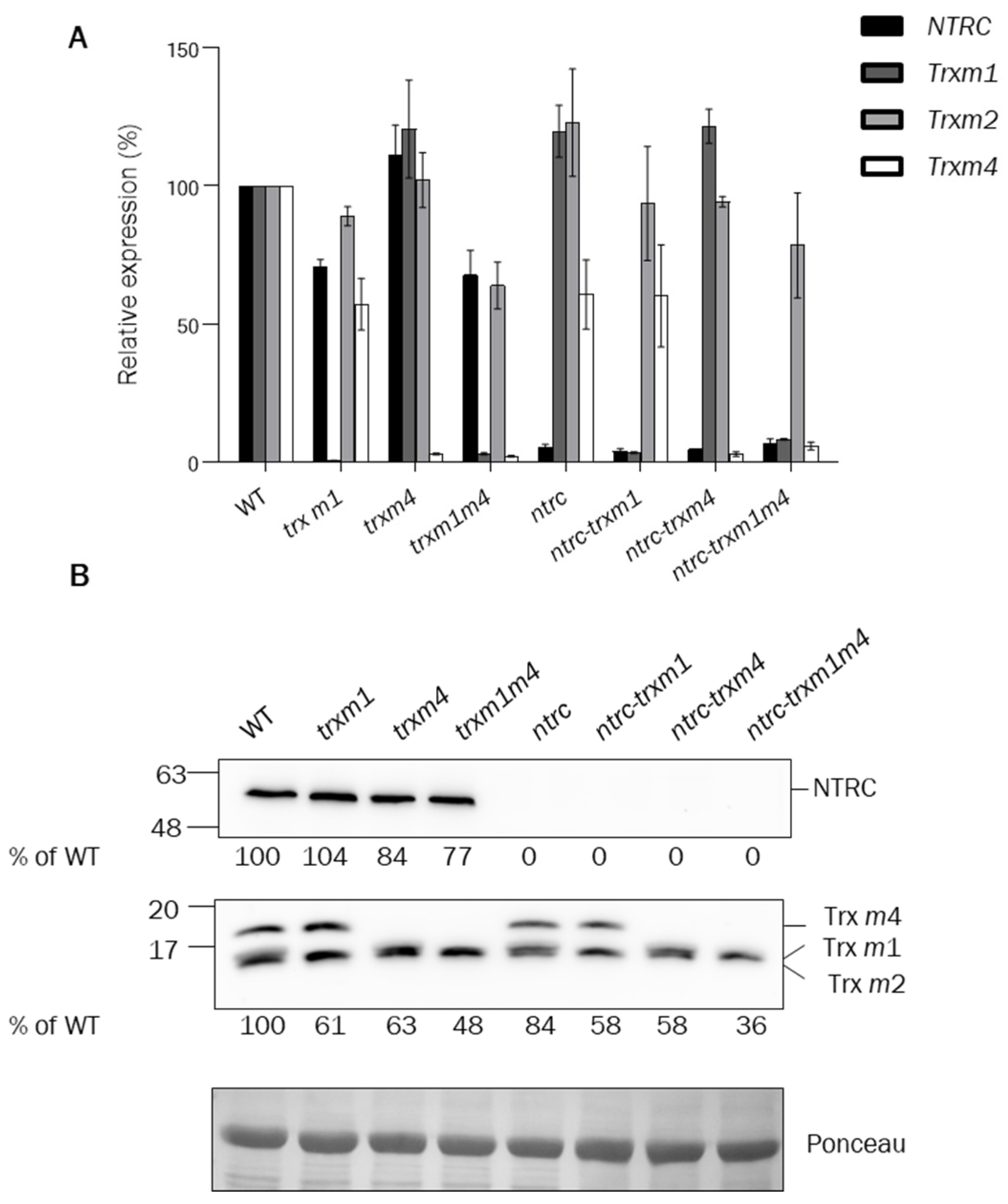
2.2. RNA Extraction, cDNA Synthesis, and Quantitative RT-PCR Analysis
2.3. Protein Extraction, Alkylation Assays, and Western-Blot Analysis
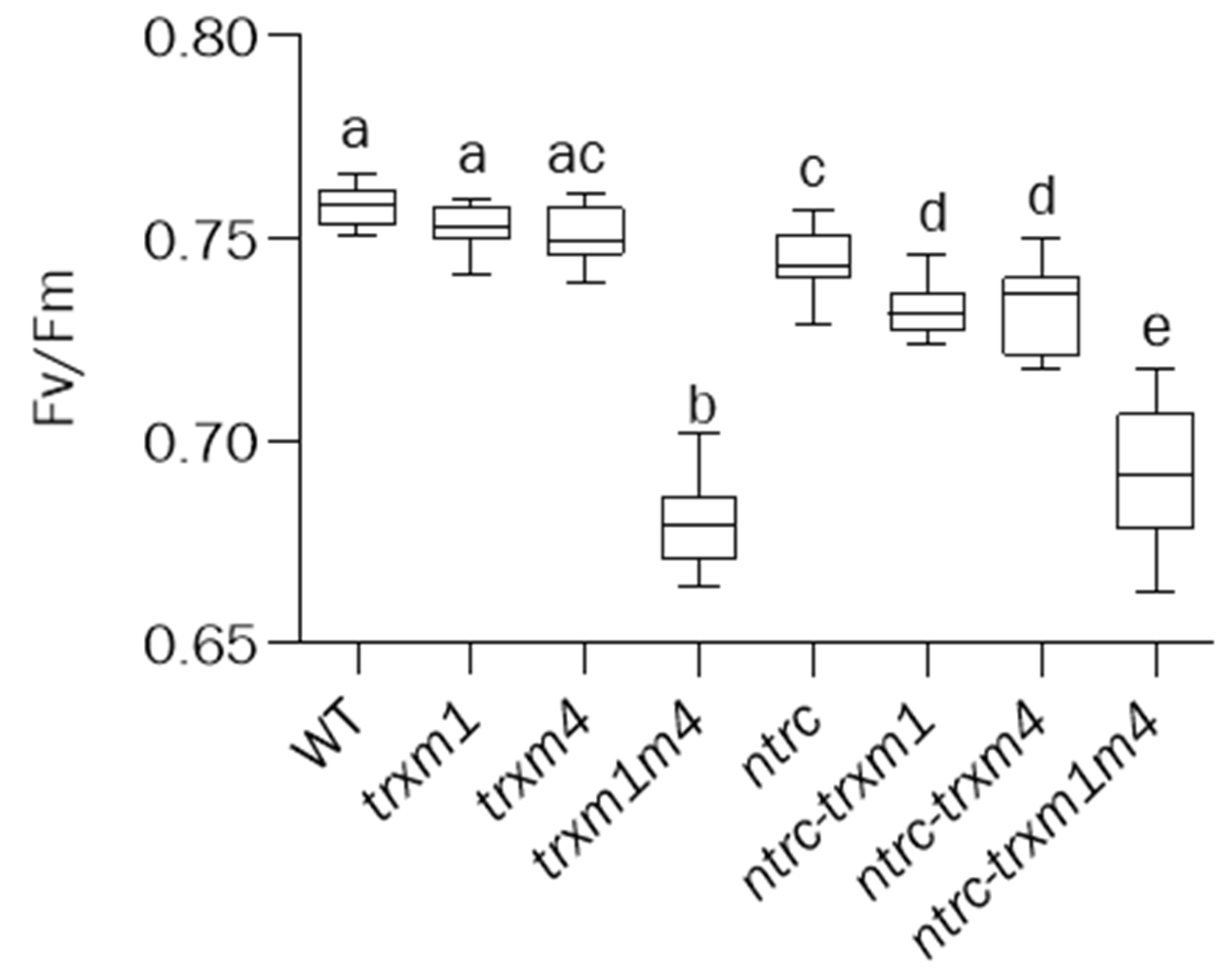
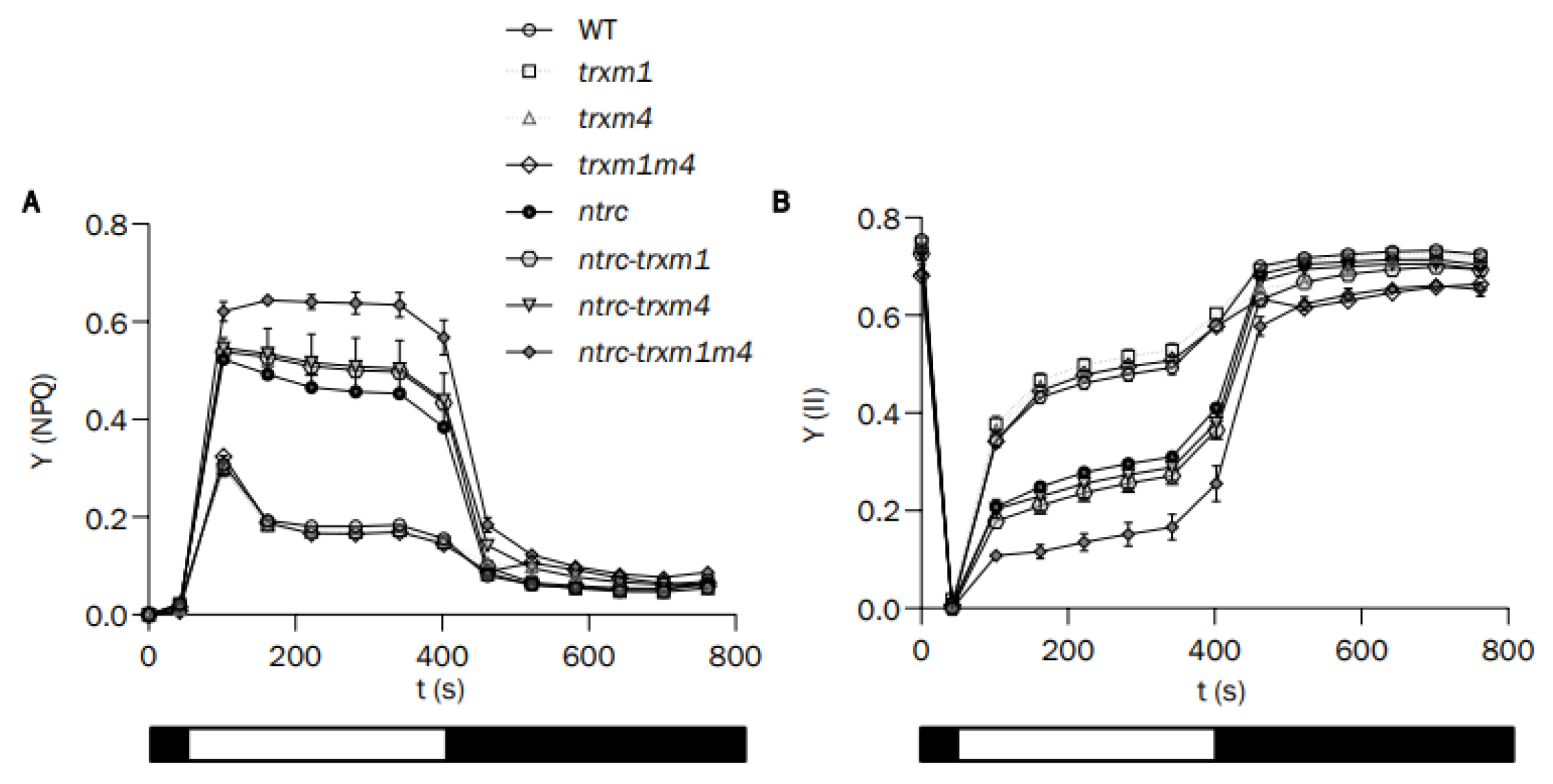
2.4. Analysis of Chlorophyll Content and Measurements of Chlorophyll a Fluorescence
2.5. Expression of NADP-MDH, Trxs m1, m2, and m4 in E. coli, Purification of the Recombinant Proteins, and In Vitro Activity Assays
3. Results
3.1. The Combined Deficiency in NTRC and Trxs m Aggravates the Growth-Inhibition Phenotype Caused by the Deficiency in NTRC
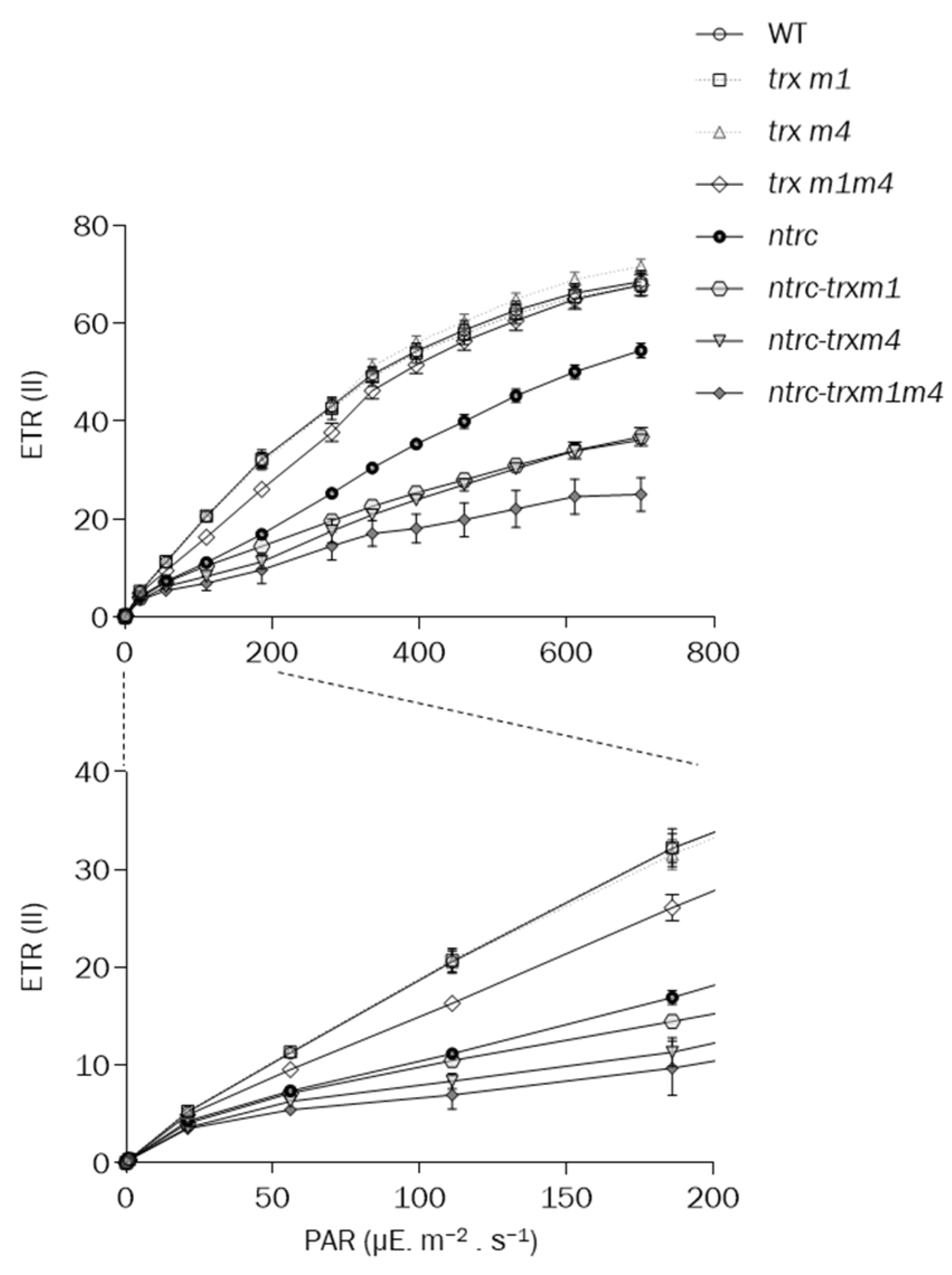
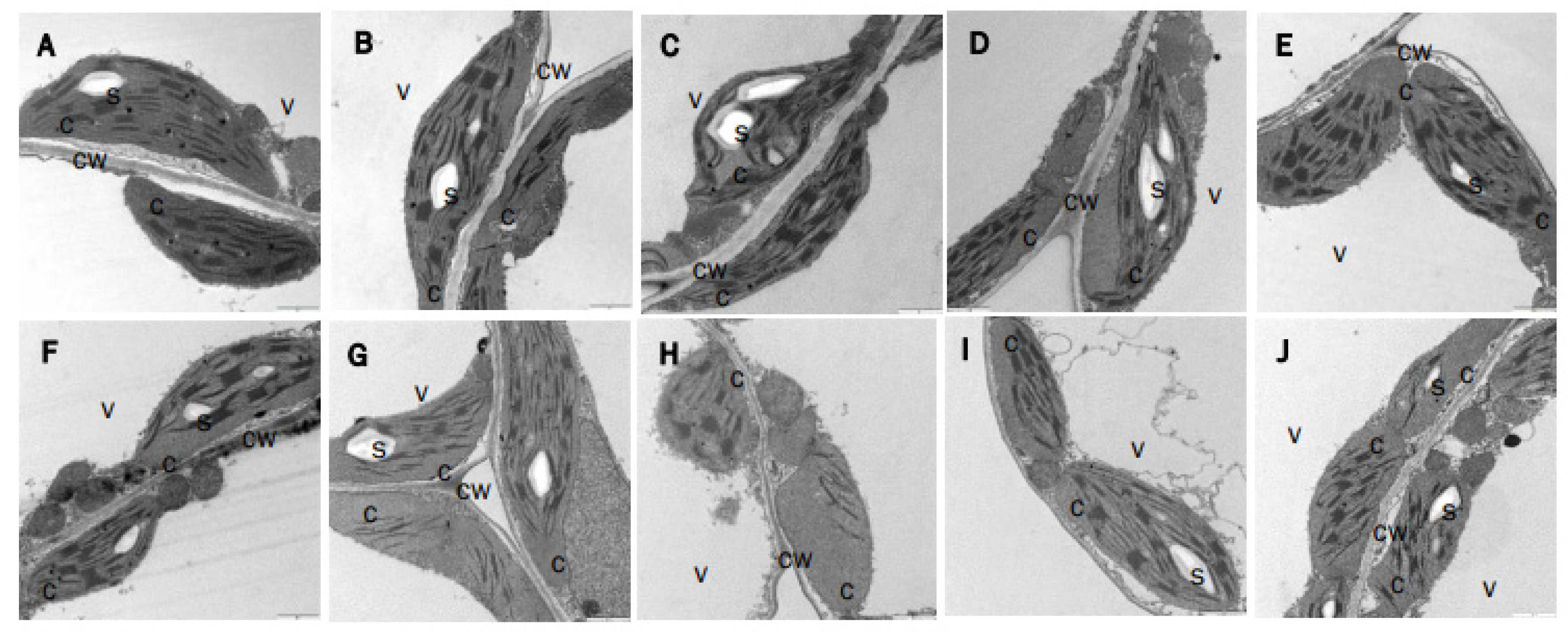
3.2. The Simultaneous Deficiency in Trxs m1 and m4 and NTRC Has an Additive Effect on Photosynthetic Performance and Chloroplast Ultrastructure
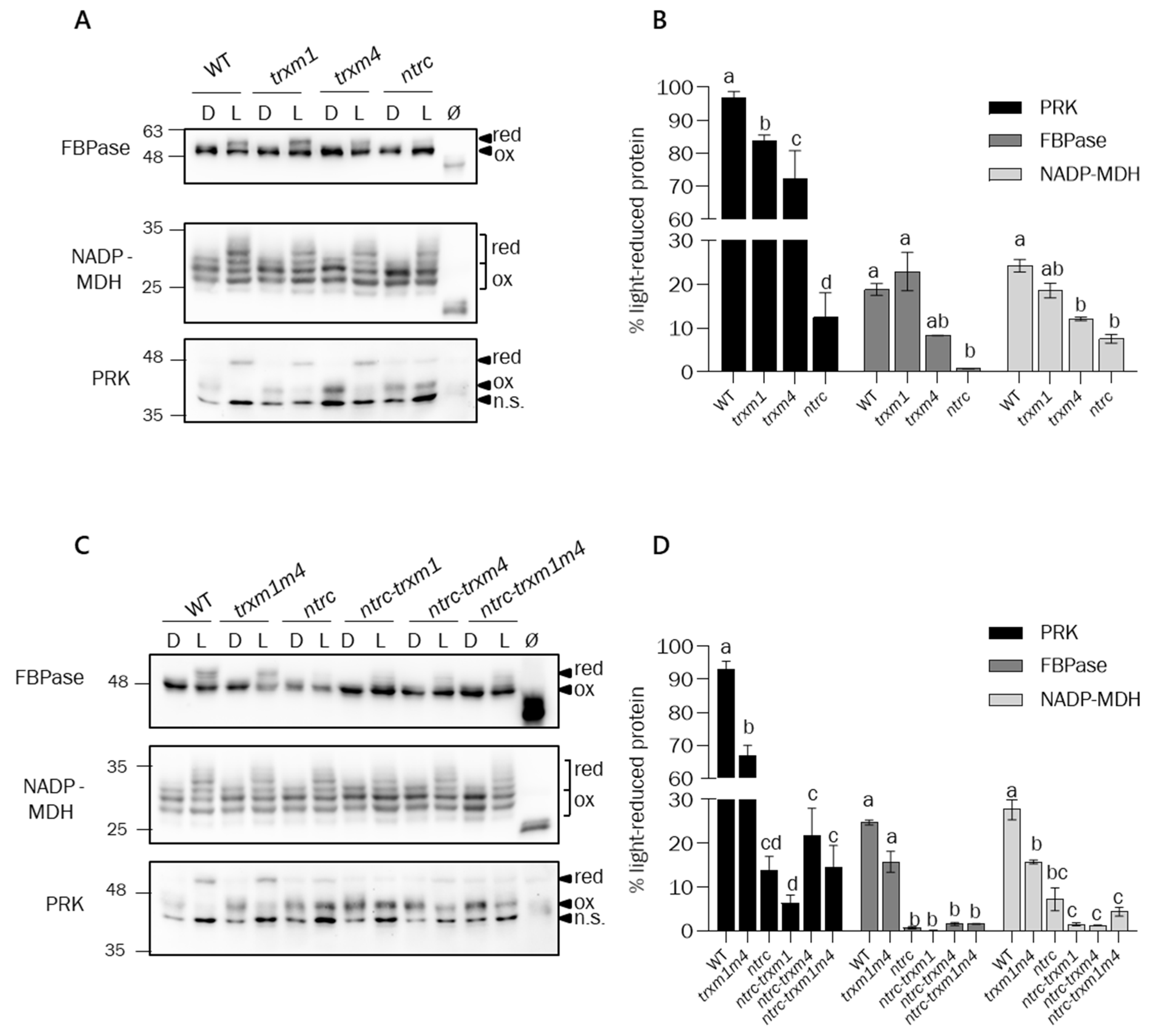
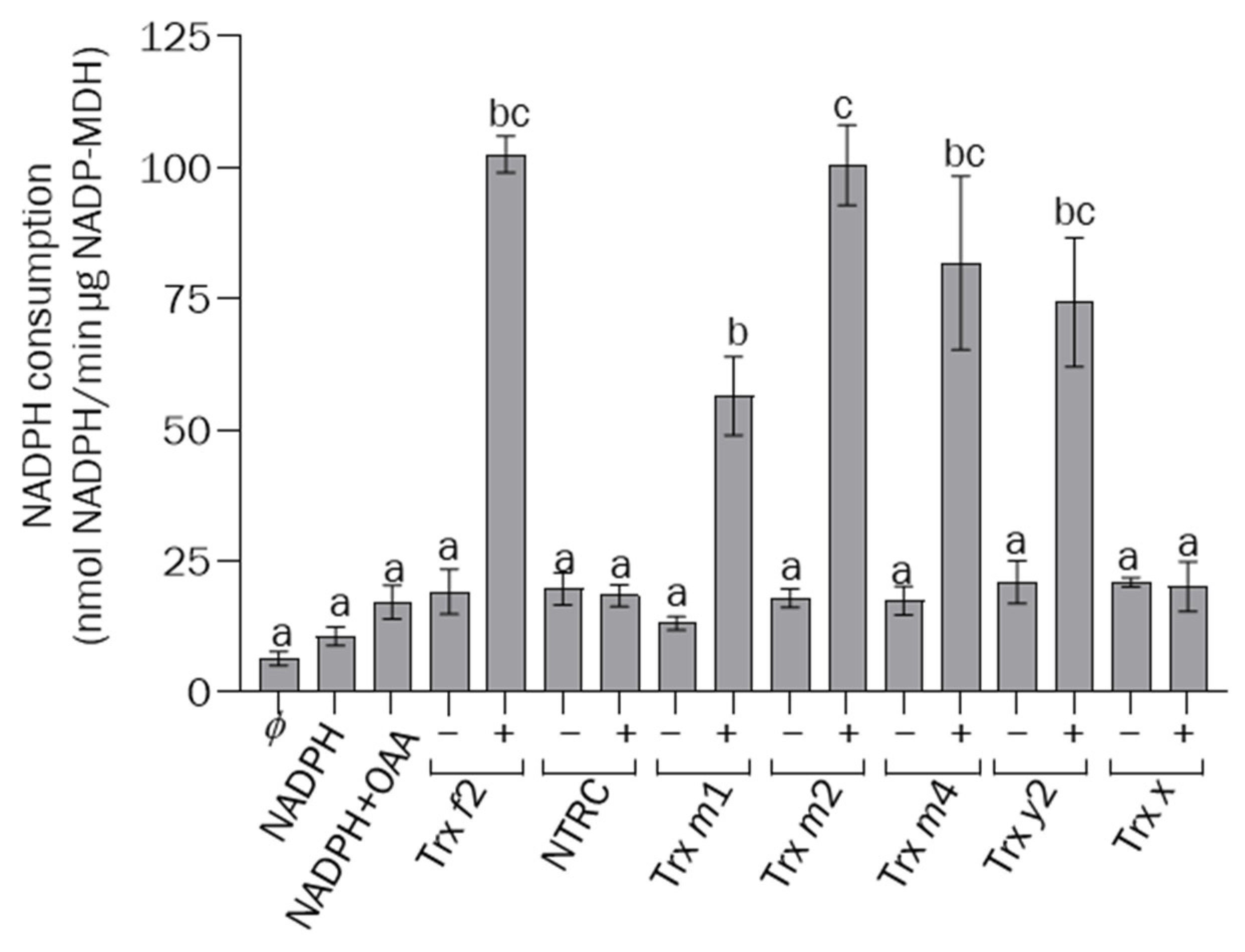
3.3. The Function of Trxs m1 and m4 in the Light-Dependent Regulation of Calvin–Benson Cycle and Malate-Valve Enzymes Requires NTRC Activity
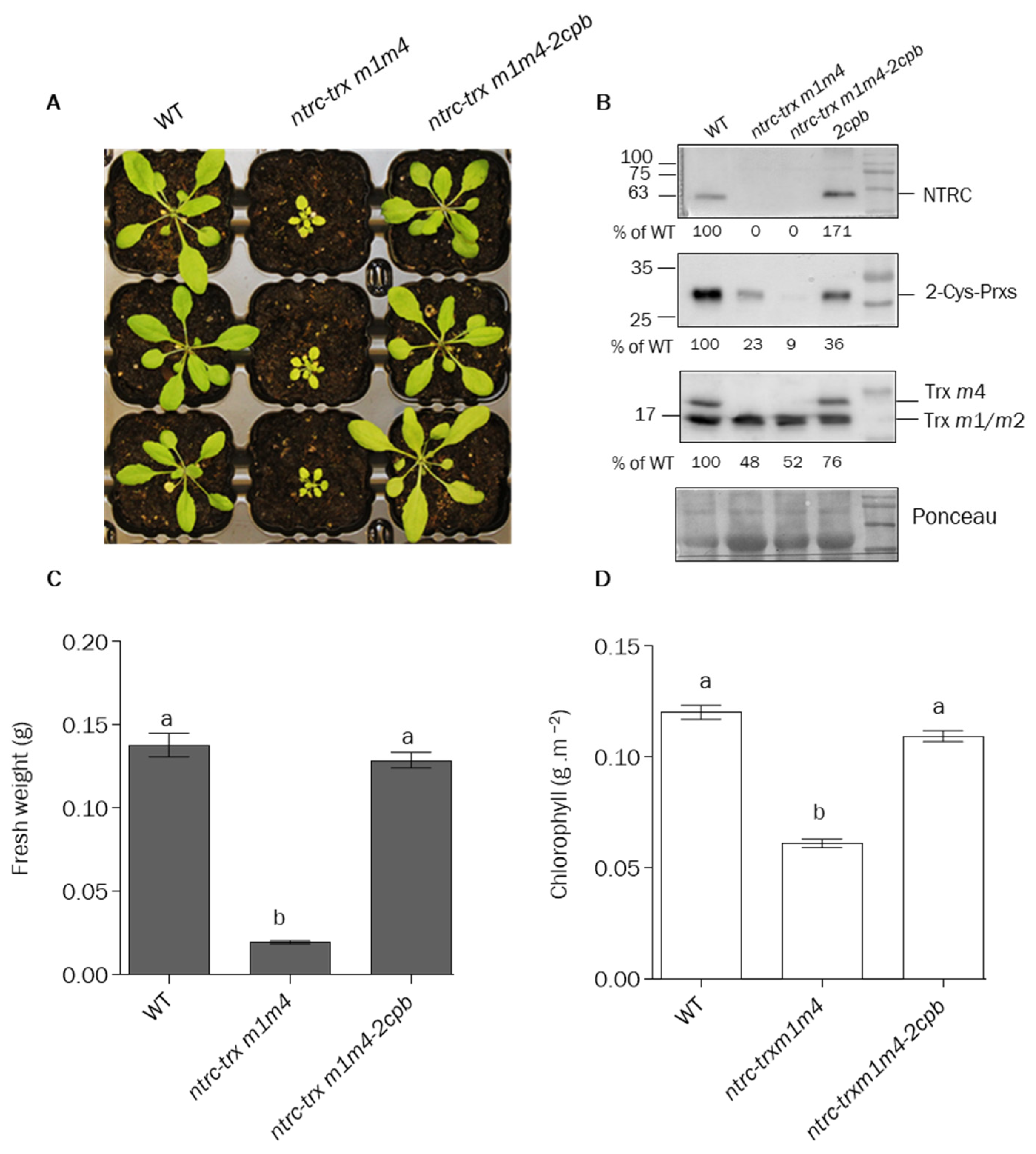
3.4. The Lack of 2-Cys Prx B Supresses the Phenotype of ntrc-trxm1m4 Mutant
4. Discussion
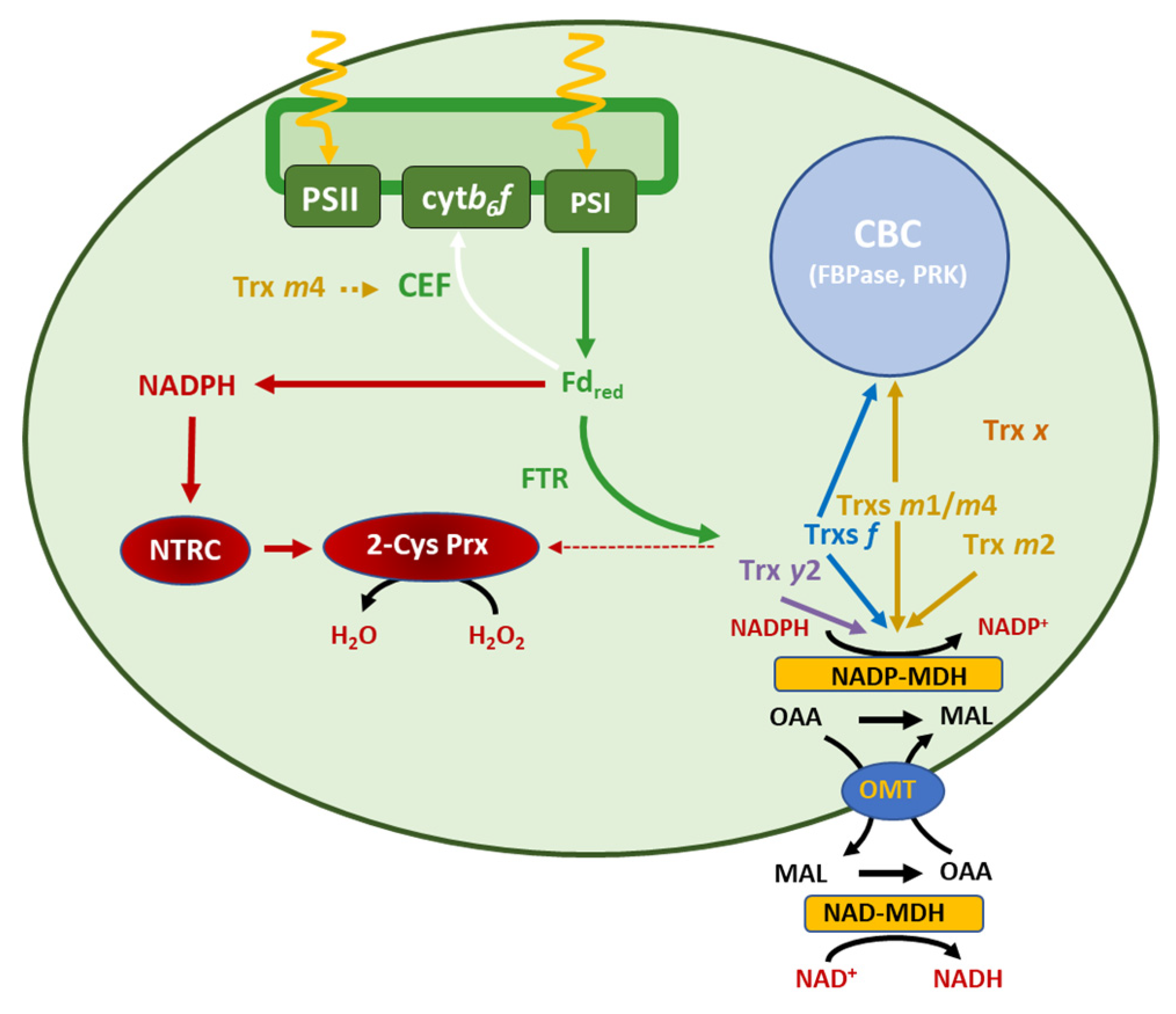
4.1. NTRC Is Required for the Trxs-m-Dependent Regulation of CBC and Malate-Valve Enzymes
4.2. The Function of NTRC and m-Type Trxs in Chloroplast Performance Depends on the Levels of 2-Cys Prxs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fürst-Jansen, J.M.R.; de Vries, S.; Lorenz, M.; von Schwartzenberg, K.; Archibald, J.M.; de Vries, J. Submergence of the filamentous Zygnematophyceae Mougeotia induces differential gene expression patterns associated with core metabolism and photosynthesis. Protoplasma 2022, 259, 1157–1174. [Google Scholar] [CrossRef]
- De Vries, J.; de Vries, S.; Curtis, B.A.; Zhou, H.; Penny, S.; Feussner, K.; Pinto, D.M.; Steinert, M.; Cohen, A.M.; von Schwartzenberg, K.; et al. Heat stress response in the closest algal relatives of land plants reveals conserved stress signaling circuits. Plant J. 2020, 103, 1025–1048. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, C.; Selmar, D. Energy dissipation is an essential mechanism to sustain the viability of plants: The physiological limits of improved photosynthesis. J. Plant Physiol. 2011, 168, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B. The path to thioredoxin and redox regulation in chloroplasts. Ann. Rev. Plant Biol. 2016, 67, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Hell, R. Nothing in biology makes sense but in the light of redox regulation. Plant Cell Physiol. 2017, 58, 1823–1825. [Google Scholar] [CrossRef] [PubMed]
- Chibani, K.; Pucker, B.; Dietz, K.-J.; Cavanagh, A. Genome-wide analysis and transcriptional regulation of the typical and atypical thioredoxins in Arabidopsis thaliana. FEBS Lett. 2021, 595, 2715–2730. [Google Scholar] [CrossRef]
- Geigenberger, P.; Thormählen, I.; Daloso, D.M.; Fernie, A.R. The unprecedented versatility of the plant thioredoxin system. Trends Plant Sci. 2017, 22, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Zaffagnini, M.; Fermani, S.; Marchand, C.H.; Costa, A.; Sparla, F.; Rouhier, N.; Geigenberger, P.; Lemaire, S.; Trost, P. Redox homeostasis in photosynthetic organisms: Novel and established thiol-based molecular mechanisms. Antioxid. Redox Signal. 2019, 31, 155–210. [Google Scholar] [CrossRef]
- Cejudo, F.J.; González, M.-C.; Pérez-Ruiz, J.M. Redox regulation of chloroplast metabolism. Plant Physiol. 2021, 186, 9–21. [Google Scholar] [CrossRef]
- Jacquot, J.-P.; Eklund, H.; Rouhier, N.; Schürmann, P. Structural and evolutionary aspects of thioredoxin reductases in photosynthetic organisms. Trends Plant Sci. 2009, 14, 336–343. [Google Scholar] [CrossRef]
- Schürmann, P.; Buchanan, B.B. The ferredoxin/thioredoxin system of oxygenic photosynthesis. Antioxid. Redox Signal. 2008, 10, 1235–1274. [Google Scholar] [CrossRef] [PubMed]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Perez-Perez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox regulation of the Calvin–Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef] [PubMed]
- Serrato, A.J.; Pérez-Ruiz, J.M.; Spinola, M.C.; Cejudo, F.J. A novel NADPH thioredoxin reductase, localized in the chloroplast, which deficiency causes hypersensitivity to abiotic stress in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 43821–43827. [Google Scholar] [CrossRef]
- Moon, J.C.; Jang, H.H.; Chae, H.B.; Lee, J.R.; Lee, S.Y.; Jung, Y.J.; Shin, M.R.; Lim, H.S.; Chung, W.S.; Yun, D.-J.; et al. The C-type Arabidopsis thioredoxin reductase ANTR-C acts as an electron donor to 2-Cys peroxiredoxins in chloroplasts. Biochem. Biophys. Res. Commun. 2006, 348, 478–484. [Google Scholar] [CrossRef]
- Pérez-Ruiz, J.M.; Spinola, M.C.; Kirchsteiger, K.; Moreno, J.; Sahrawy, M.; Cejudo, F.J. Rice NTRC is a high-efficiency redox system for chloroplast protection against oxidative damage. Plant Cell 2006, 18, 2356–2368. [Google Scholar] [CrossRef] [PubMed]
- Alkhalfioui, F.; Renard, M.; Montrichard, F. Unique properties of NADP-thioredoxin reductase C in legumes. J. Exp. Bot. 2007, 58, 969–978. [Google Scholar] [CrossRef]
- Bernal-Bayard, P.; Hervas, M.; Cejudo, F.J.; Navarro, J.A. Electron transfer pathways and dynamics of chloroplast NADPH-dependent thioredoxin reductase C (NTRC). J. Biol. Chem. 2012, 287, 33865–33872. [Google Scholar] [CrossRef]
- Pascual, M.B.; Mata-Cabana, A.; Florencio, F.J.; Lindahl, M.; Cejudo, F.J. A comparative analysis of the NADPH thioredoxin reductase C-2-Cys peroxiredoxin system from plants and cyanobacteria. Plant Phys. 2011, 155, 1806–1816. [Google Scholar] [CrossRef] [PubMed]
- Michalska, J.; Zauber, H.; Buchanan, B.B.; Cejudo, F.J.; Geigenberger, P. NTRC links built-in thioredoxin to light and sucrose in regulating starch synthesis in chloroplasts and amyloplasts. Proc. Natl. Acad. Sci. USA 2009, 106, 9908–9913. [Google Scholar] [CrossRef]
- Lepistö, A.; Pakula, E.; Toivola, J.; Krieger-Liszkay, A.; Vignols, F.; Rintamäki, E. Deletion of chloroplast NADPH-dependent thioredoxin reductase results in inability to regulate starch synthesis and causes stunted growth under short-day photoperiods. J. Exp. Bot. 2013, 64, 3843–3854. [Google Scholar] [CrossRef]
- Stenbaek, A.; Hanson, A.; Wulff, R.P.; Hansson, M.; Dietz, K.-J.; Jensen, P.E. NADPH-dependent thioredoxin reductase and 2-Cys peroxiredoxins are needed for the protection of Mg-protoporphyrin monomethyl ester cyclase. FEBS Lett. 2008, 582, 2773–2778. [Google Scholar] [CrossRef]
- Richter, A.S.; Peter, E.; Rothbart, M.; Schlicke, H.; Toivola, J.; Rintamäki, E.; Grimm, B. Posttranslational influence of NADPH-dependent thioredoxin reductase C on enzymes in tetrapyrrole synthesis. Plant Phys. 2013, 162, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ruiz, J.M.; Guinea, M.; Puerto-Galán, L.; Cejudo, F.J. NADPH thioredoxin reductase C is involved in redox regulation of the Mg-chelatase I subunit in Arabidopsis thaliana chloroplasts. Mol. Plant 2014, 7, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Hisabori, T. Two distinct redox cascades cooperatively regulate chloroplast functions and sustain plant viability. Proc. Natl. Acad. Sci. USA 2016, 113, E3967–E3976. [Google Scholar] [CrossRef]
- González, M.; Delgado-Requerey, V.; Ferrández, J.; Serna, A.; Cejudo, F.J. Insights into the function of NADPH thioredoxin reductase C (NTRC) based on identification of NTRC-interacting proteins in vivo. J. Exp. Bot. 2019, 70, 5787–5798. [Google Scholar] [CrossRef]
- Ancín, M.; Fernández-Irigoyen, J.; Santamaría, E.; Larraya, L.; Fernández-San Millán, A.; Veramendi, J.; Farrán, I. New in vivo approach to burden the thioredoxin family interactome in chloroplasts. Antioxidants 2022, 11, 1979. [Google Scholar] [CrossRef]
- Montrichard, F.; Alkhalfioui, F.; Yano, H.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin targets in plants: The first 30 years. J. Proteom. 2009, 72, 452–474. [Google Scholar] [CrossRef] [PubMed]
- Nikkanen, L.; Rintamäki, E. Chloroplast thioredoxin systems dynamically regulate photosynthesis in plants. Biochem. J. 2019, 476, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Guinea Diaz, M.; Nikkanen, L.; Himanen, K.; Toivola, J.; Rintamäki, E. Two chloroplast thioredoxin systems differentially modulate photosynthesis in Arabidopsis depending on light intensity and leaf age. Plant J. 2020, 104, 718–734. [Google Scholar] [CrossRef] [PubMed]
- Collin, V.; Issakidis-Bourguet, E.; Marchand, C.; Hirasawa, M.; Lancelin, J.-M.; Knaff, D.B.; Miginiac-Maslow, M. The Arabidopsis plastidial thioredoxins. J. Biol. Chem. 2003, 278, 23747–23752. [Google Scholar] [CrossRef]
- Collin, V.; Lamkemeyer, P.; Miginiac-Maslow, M.; Hirasawa, M.; Knaff, D.B.; Dietz, K.-J.; Issakidis-Bourguet, E. Characterization of plastidial thioredoxins from Arabidopsis belonging to the new y-type. Plant Phys. 2004, 136, 4088–4095. [Google Scholar] [CrossRef]
- Okegawa, Y.; Motohashi, K. Chloroplastic thioredoxin m functions as a major regulator of Calvin cycle enzymes during photosynthesis in vivo. Plant J. 2015, 84, 900–913. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, B.; Diaz-Espejo, A.; Lindahl, M.; Cejudo, F.J. Type-f thioredoxins have a role in the short-term activation of carbon metabolism and their loss affects growth under short-day conditions in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 1951–1964. [Google Scholar] [CrossRef] [PubMed]
- Arsova, B.; Hoja, U.; Wimmelbacher, M.; Greiner, E.; Ustun, S.; Melzer, M.; Petersen, K.; Lein, W.; Bornke, F. Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: Evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell 2010, 22, 1498–1515. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liu, J.; Liu, B.; Da, Q.; Feng, D.; Su, J.; Zhang, Y.; Wang, J.; Wang, H.-B. Ferredoxin:thioredoxin reductase is required for proper chloroplast development and is involved in the regulation of plastid gene expression in Arabidopsis thaliana. Mol. Plant 2014, 7, 1586–1590. [Google Scholar] [CrossRef] [PubMed]
- Thormählen, I.; Meitzel, T.; Groysman, J.; Öchsner, A.B.; von Roepenack-Lahaye, E.; Naranjo, B.; Cejudo, F.J.; Geigenberger, P. Thioredoxin f1 and NADPH-dependent thioredoxin reductase C have overlapping functions in regulating photosynthetic metabolism and plant growth in response to varying light conditions. Plant Phys. 2015, 169, 1766–1786. [Google Scholar] [CrossRef]
- Ojeda, V.; Nájera, V.A.; González, M.; Pérez-Ruiz, J.M.; Cejudo, F.J. Photosynthetic activity of cotyledons is critical during post-germinative growth and seedling establishment. Plant Signal. Behav. 2017, 12, e1347244. [Google Scholar] [CrossRef]
- Ojeda, V.; Pérez-Ruiz, J.M.; Gonzalez, M.; Najera, V.A.; Sahrawy, M.; Serrato, A.J.; Geigenberger, P.; Cejudo, F.J. NADPH thioredoxin reductase C and thioredoxins act concertedly in seedling development. Plant Phys. 2017, 174, 1436–1448. [Google Scholar] [CrossRef]
- Jurado-Flores, A.; Delgado-Requerey, V.; Gálvez-Ramírez, A.; Puerto-Galán, L.; Pérez-Ruiz, J.M.; Cejudo, F.J. Exploring the functional relationship between y-type thioredoxins and 2-Cys peroxiredoxins in Arabidopsis chloroplasts. Antioxidants 2020, 9, 1072. [Google Scholar] [CrossRef]
- Pérez-Ruiz, J.M.; Naranjo, B.; Ojeda, V.; Guinea, M.; Cejudo, F.J. NTRC-dependent redox balance of 2-Cys peroxiredoxins is needed for optimal function of the photosynthetic apparatus. Proc. Natl. Acad. Sci. USA 2017, 114, 12069–12074. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, V.; Pérez-Ruiz, J.M.; Cejudo, F.J. The NADPH-dependent thioredoxin reductase C-2-Cys peroxiredoxin redox system modulates the activity of thioredoxin x in Arabidopsis chloroplasts. Plant Cell Physiol. 2018, 59, 2155–2164. [Google Scholar] [CrossRef]
- Benitez-Alfonso, Y.; Cilia, M.; Roman, A.S.; Thomas, C.; Maule, A.; Hearn, S.; Jackson, D. Control of Arabidopsis meristem development by thioredoxin-dependent regulation of intercellular transport. Proc. Natl. Acad. Sci. USA 2009, 106, 3615–3620. [Google Scholar] [CrossRef]
- Da, Q.; Wang, P.; Wang, M.; Sun, T.; Jin, H.; Liu, B.; Wang, J.; Grimm, B.; Wang, H.B. Thioredoxin and NADPH-dependent thioredoxin reductase C regulation of tetrapyrrole biosynthesis. Plant Phys. 2017, 175, 652–666. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Trijueque, J.; Serrato, A.-J.; Sahrawy, M. Proteomic analyses of thioredoxins f and m Arabidopsis thaliana mutants indicate specific functions for these proteins in plants. Antioxidants 2019, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Serrato, A.J.; Rojas González, J.A.; Torres-Romero, D.; Vargas, P.; Mérida, A.; Sahrawy, M. Thioredoxins m are major players in the multifaceted light-adaptive response in Arabidopsis thaliana. Plant J. 2021, 108, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Pulido, P.; Spínola, M.C.; Kirchsteiger, K.; Guinea, M.; Pascual, M.B.; Sahrawy, M.; Sandalio, L.M.; Dietz, K.-J.; González, M.; Cejudo, F.J. Functional analysis of the pathways for 2-Cys peroxiredoxin reduction in Arabidopsis thaliana chloroplasts. J. Exp. Bot. 2010, 61, 4043–4054. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta 1989, 75, 384–394. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New fluorescence parameters for the determination of q(a) redox state and excitation energy fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Wang, P.; Liu, J.; Liu, B.; Feng, D.; Da, Q.; Wang, P.; Shu, S.; Su, J.; Zhang, Y.; Wang, J.; et al. Evidence for a role of chloroplastic m-type thioredoxins in the biogenesis of Photosystem II in Arabidopsis. Plant Phys. 2013, 163, 1710–1728. [Google Scholar] [CrossRef]
- Courteille, A.; Vesa, S.; Sanz-Barrio, R.; Cazale, A.C.; Becuwe-Linka, N.; Farran, I.; Havaux, M.; Rey, P.; Rumeau, D. Thioredoxin m4 controls photosynthetic alternative electron pathways in Arabidopsis. Plant Phys. 2013, 161, 508–520. [Google Scholar] [CrossRef]
- Selinski, J.; Scheibe, R. Malate valves: Old shuttles with new perspectives. Plant Biol. 2019, 21, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Hara, S.; Hisabori, T. Thioredoxin selectivity for thiol-based redox regulation of target proteins in chloroplasts. J. Biol. Chem. 2015, 290, 14278–14288. [Google Scholar] [CrossRef] [PubMed]
- Meinke, D.; Sweeney, C.; Muralla, R. Integrating the genetic and physical maps of Arabidopsis thaliana: Identification of mapped alleles of cloned essential (EMB) genes. PLoS ONE 2009, 4, e7386. [Google Scholar] [CrossRef] [PubMed]
- Goulas, E.; Schubert, M.; Kieselbach, T.; Kleczkowski, L.; Gardeström, P.; Schröder, W.; Hurry, V. The chloroplast lumen and stromal proteomes of Arabidopsis thaliana show differential sensitivity to short- and long-term exposure to low temperature. Plant J. 2006, 47, 720–734. [Google Scholar] [CrossRef]
- Zhang, M.; Takano, T.; Liu, S.; Zhang, X. Arabidopsis mitochondrial voltage-dependent anion channel 3 (AtVDAC3) protein interacts with thioredoxin m2. FEBS Lett. 2015, 589, 1207–1213. [Google Scholar] [CrossRef]
- Sahrawy, M.; Fernández-Trijueque, J.; Vargas, P.; Serrato, A.J. Comprehensive expression analyses of plastidial thioredoxins of Arabidopsis thaliana indicate a main role of thioredoxin m2 in roots. Antioxidants 2022, 11, 1365. [Google Scholar] [CrossRef]
- Laugier, E.; Tarrago, L.; Courteille, A.; Innocenti, G.; Eymery, F.; Rumeau, D.; Issakidis-Bourguet, E.; Rey, P. Involvement of thioredoxin y2 in the preservation of leaf methionine sulfoxide reductase capacity and growth under high light. Plant Cell Environ. 2013, 36, 670–682. [Google Scholar] [CrossRef] [PubMed]
- Thormählen, I.; Zupok, A.; Rescher, J.; Leger, J.; Weissenberger, S.; Groysman, J.; Orwat, A.; Chatel-Innocenti, G.; Issakidis-Bourguet, E.; Armbruster, U.; et al. Thioredoxins play a crucial role in dynamic acclimation of photosynthesis in fluctuating light. Mol. Plant 2017, 10, 168–182. [Google Scholar] [CrossRef]
- Rey, P.; Sanz-Barrio, R.; Innocenti, G.; Ksas, B.; Courteille, A.; Rumeau, D.; Issakidis-Bourguet, E.; Farran, I. Overexpression of plastidial thioredoxins f and m differentially alters photosynthetic activity and response to oxidative stress in tobacco plants. Front. Plant Sci. 2013, 4, 390. [Google Scholar] [CrossRef]
- Yokochi, Y.; Yoshida, K.; Hahn, F.; Miyagi, A.; Wakabayashi, A.-I.; Kaway-Yamada, M.; Weber, A.P.M.; Hisabori, T. Redox regulation of NADP-malate dehydrogenase is vital for land plants under fluctuating light environment. Proc. Natl. Acad. Sci. USA 2021, 118, e2016903118. [Google Scholar] [CrossRef] [PubMed]
- Marri, L.; Zaffagnini, M.; Collin, V.; Issakidis-Bourguet, E.; Lemaire, S.D.; Pupillo, P.; Sparla, F.; Miginiac-Maslow, M.; Trost, P. Prompt and easy activation by specific thioredoxins of Calvin Cycle enzymes of Arabidopsis thaliana associated in the GAPDH/CP12/PRK supramolecular complex. Mol. Plant 2009, 2, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Hebbelmann, I.; Selinski, J.; Wehmeyer, C.; Goss, T.; Voss, I.; Mulo, P.; Kangasjärvi, S.; Aro, E.-M.; Oelze, M.-L.; Dietz, K.-J.; et al. Multiple strategies to prevent oxidative stress in Arabidopsis plants lacking the malate valve enzyme NADP-malate dehydrogenase. J. Exp. Bot. 2012, 63, 1445–1459. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Requerey, V.; Cejudo, F.J.; González, M.-C. The Functional Relationship between NADPH Thioredoxin Reductase C, 2-Cys Peroxiredoxins, and m-Type Thioredoxins in the Regulation of Calvin–Benson Cycle and Malate-Valve Enzymes in Arabidopsis. Antioxidants 2023, 12, 1041. https://doi.org/10.3390/antiox12051041
Delgado-Requerey V, Cejudo FJ, González M-C. The Functional Relationship between NADPH Thioredoxin Reductase C, 2-Cys Peroxiredoxins, and m-Type Thioredoxins in the Regulation of Calvin–Benson Cycle and Malate-Valve Enzymes in Arabidopsis. Antioxidants. 2023; 12(5):1041. https://doi.org/10.3390/antiox12051041
Chicago/Turabian StyleDelgado-Requerey, Víctor, Francisco Javier Cejudo, and María-Cruz González. 2023. "The Functional Relationship between NADPH Thioredoxin Reductase C, 2-Cys Peroxiredoxins, and m-Type Thioredoxins in the Regulation of Calvin–Benson Cycle and Malate-Valve Enzymes in Arabidopsis" Antioxidants 12, no. 5: 1041. https://doi.org/10.3390/antiox12051041