
Blood Plasma Calorimetric Profiles of Women with Preeclampsia: Effect of Oxidative Stress
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Patients’ Characteristics
2.2. DSC Blood Plasma Profiles of Healthy Nonpregnant Women and Pregnant Women in the Third Trimester of Pregnancy
2.3. DSC Plasma Profiles of Patients with Preeclampsia
2.4. DSC Blood Plasma Profiles of Patients with Severe vs. Non-Severe Preeclampsia
2.5. Protein Fractions of PC and PE Samples
2.6. Oxidative Stress Simulation
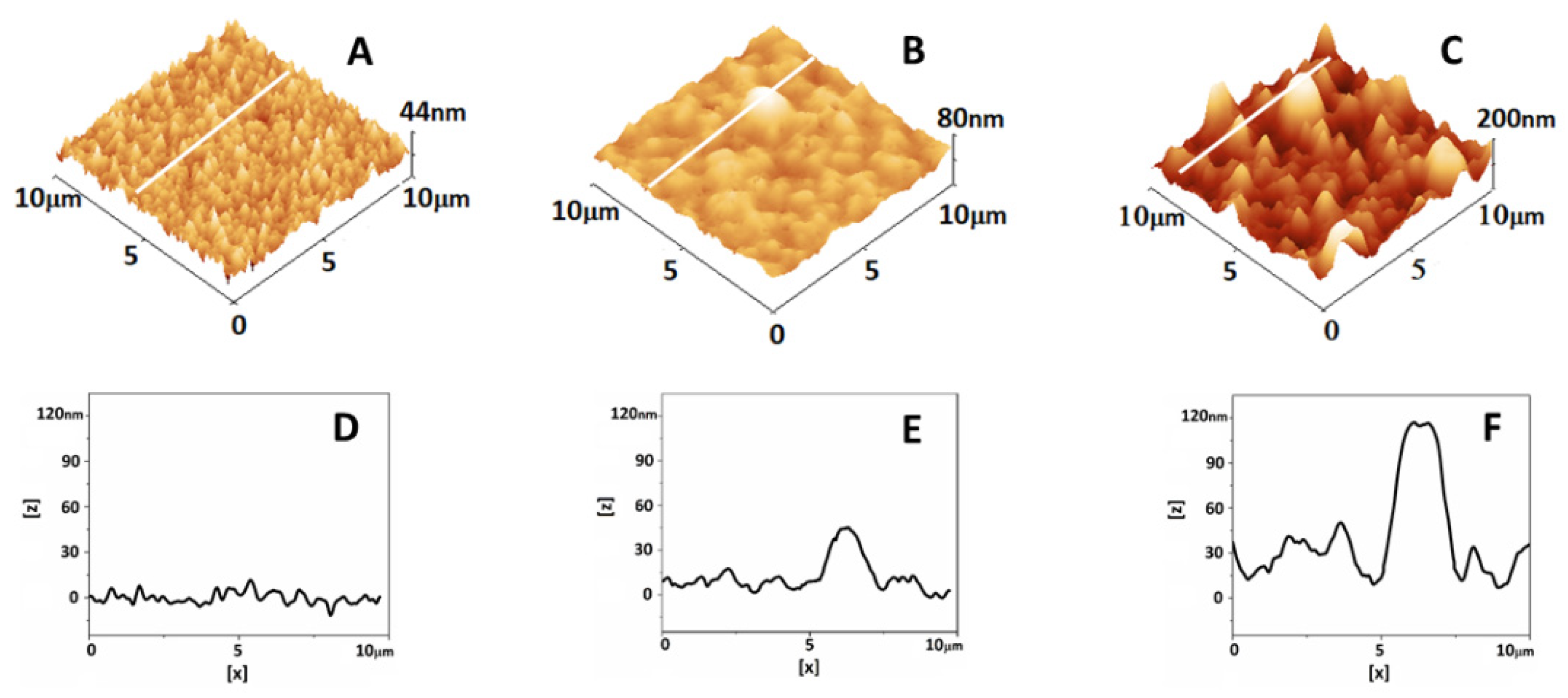
2.7. Atomic Force Microscopy Analysis of Plasma
3. Discussion
3.1. Changes in Blood Plasma Proteome in the Last Trimester of Pregnancy
3.2. Deviations in Calorimetric Profiles and Blood Plasma Proteome in Preeclampsia
3.3. Simulation Experiments on the Effect of Oxidative Stress
Limitations
4. Patients and Methods
4.1. Study Groups and Ethics Statement
4.2. Blood Collection
4.3. Sample Preparation
4.4. Protein Content Analysis
4.5. Plasma and HSA Oxidation
4.5.1. Plasma Oxidation
4.5.2. HSA Oxidation
4.6. DSC Experiments
4.7. AFM Measurements
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- da Silva, W.A.; Pinheiro, A.M.; Lima, P.H.; Malbouisson, L.M.S. Renal and cardiovascular repercussions in preeclampsia and their impact on fluid management: A literature review. Braz. J. Anesthesiol. 2021, 71, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Tanner, M.S.; Davey, M.-A.; Mol, B.W.; Rolnik, D.L. The evolution of the diagnostic criteria of preeclampsia-eclampsia. Am. J. Obstet. Gynecol. 2022, 226, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Gomez, T.; Dominguez, F.; Quiñonero, A.; Diaz-Gimeno, P.; Kapidzic, M.; Gormley, M.; Ona, K.; Padilla-Iserte, P.; McMaster, M.; Genbacev, O.; et al. Defective decidualization during and after severe preeclampsia reveals a possible maternal contribution to the etiology. Proc. Natl. Acad. Sci. USA 2017, 114, E8468–E8477. [Google Scholar] [CrossRef]
- Newhouse, S.; Davidge, S.; Winkler-Lowen, B.; Demianczuk, N.; Guklbert, L. In vitro differentiation of villous trophoblasts from pregnancies complicated by intrauterine growth restriction with and without preeclampsia. Placenta 2007, 28, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Opichka, M.A.; Rappelt, M.W.; Gutterman, D.D.; Grobe, J.L.; McIntosh, J.J. Vascular Dysfunction in Preeclampsia. Cells 2021, 10, 3055. [Google Scholar] [CrossRef]
- Hung, T.H.; Skepper, J.N.; Charnock-Jones, D.S.; Burton, G.J. Hypoxia-reoxygenation: A potent inducer of apoptotic changes in the human placenta and possible etiological factor in preeclampsia. Circ. Res. 2002, 90, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Bokslag, A.; van Weissenbruch, M.; Mol, B.W.; de Groot, C.J.M. Preeclampsia; Short and Long-term Consequences for Mother and Neonate. Early Hum. Dev. 2016, 102, 47–50. [Google Scholar] [CrossRef]
- Hung, T.H.; Burton, G.J. Hypoxia and reoxygenation: A possible mechanism for placental oxidative stress in preeclampsia. Taiwan J. Obs. Gynecol. 2006, 45, 189–200. [Google Scholar] [CrossRef]
- Powe, C.E.; Levine, R.J.; Karumanchi, S.A. Preeclampsia, a disease of the maternal endothelium: The role of antiangiogenic factors and implications for later cardiovascular disease. Circulation 2011, 123, 2856–2869. [Google Scholar] [CrossRef]
- Venkatesha, S.; Toporsian, M.; Lam, C.; Hanai, J.; Mammoto, T.; Kim, Y.M.; Bdolah, Y.; Lim, K.H.; Yuan, H.T.; Libermann, T.A.; et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat. Med. 2006, 12, 642–649. [Google Scholar] [CrossRef]
- Levine, R.J.; Lam, C.; Qian, C.; Yu, K.F.; Maynard, S.E.; Sachs, B.P.; Sibai, B.M.; Epstein, F.H.; Romero, R.; Thadhani, R.; et al. CPEP Study Group. Soluble endoglin and other circulating antiangiogenic factors in preeclampsia. N. Engl. J. Med. 2006, 355, 992–1005. [Google Scholar] [CrossRef]
- Myers, J.; Mires, G.; Macleod, M.; Baker, P. In preeclampsia, the circulating factors capable of altering in vitro endothelial function precede clinical disease. Hypertension 2005, 45, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Raijmakers, M.T.M.; Dechend, R.; Poston, L. Oxidative Stress and Preeclampsia. Hypertension 2004, 44, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Khankin, E.V.; Royle, C.; Karumanchi, S.A. Placental vasculature in health and disease. Semin. Thromb. Hemost. 2010, 36, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Banerjee, S.; Daiello, L.A.; Nakashima, A.; Jash, S.; Huang, Z.; Drake, J.D.; Ernerudh, J.; Berg, G.; Padbury, J.; et al. Novel blood test for early biomarkers of preeclampsia and Alzheimer’s disease. Sci. Rep. 2021, 11, 15934. [Google Scholar] [CrossRef] [PubMed]
- Kalkunte, S.S.; Neubeck, S.; Norris, W.E.; Cheng, S.-B.; Kostadinov, S.; Hoang, D.V.; Ahmed, A.; von Eggeling, F.; Shaikh, Z.; Padbury, J.; et al. Transthyretin is dysregulated in preeclampsia, and its native form prevents the onset of disease in a preclinical mouse model. Am. J. Pathol. 2013, 183, 1425–1436. [Google Scholar] [CrossRef] [PubMed]
- Buhimschi, I.A.; Nayeri, U.A.; Zhao, G.; Shook, L.L.; Pensalfini, A.; Funai, E.F.; Bernstein, I.M.; Glabe, C.G.; Buhimschi, C.S. Protein misfolding, congophilia, oligomerization, and defective amyloid processing in preeclampsia. Sci. Transl. Med. 2014, 6, 245ra92. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Cheng, S.; Chen, Q.; DeSousa, J.; Stone, P.R.; James, J.L.; Chamley, L.W.; Sharma, S. Aggregated transthyretin is specifically packaged into placental nano-vesicles in preeclampsia. Sci. Rep. 2017, 7, 6694. [Google Scholar] [CrossRef]
- Cater, J.H.; Kumita, J.R.; Abdallah, R.Z.; Zhao, G.; Bernardo-Gancedo, A.; Henry, A.; Winata, W.; Chi, M.; Grenyer, B.S.F.; Townsend, M.L.; et al. Human pregnancy zone protein stabilizes misfolded proteins including preeclampsia- and Alzheimer’s-associated amyloid beta peptide. Proc. Natl. Acad. Sci. USA 2019, 116, 6101–6110. [Google Scholar] [CrossRef]
- Cheng, S.B.; Nakashima, A.; Sharma, S. Understanding Pre-Eclampsia Using Alzheimer’s Etiology: An Intriguing Viewpoint. Am. J. Reprod. Immunol. 2016, 75, 372–381. [Google Scholar] [CrossRef]
- Gerasimova, E.M.; Fedotov, S.A.; Kachkin, D.V.; Vashukova, E.S.; Glotov, A.S.; Chernoff, Y.O.; Rubel, A.A. Protein Misfolding during Pregnancy: New Approaches to Preeclampsia Diagnostics. Int. J. Mol. Sci. 2019, 20, 6183. [Google Scholar] [CrossRef]
- Meads, C.A.; Cnossen, J.S.; Meher, S.; Juarez-Garcia, A.; ter Riet, G.; Duley, L.; Roberts, T.E.; Mol, B.W.; van der Post, J.A.; Leeflang, M.M.; et al. Methods of prediction and prevention of pre-eclampsia: Systematic reviews of accuracy and effectiveness literature with economic modelling. Health Technol. Assess. 2008, 12, 1–270. [Google Scholar] [CrossRef] [PubMed]
- Kirbas, A.; Ersoy, A.O.; Daglar, K.; Dikici, T.; Biberoglu, E.H.; Kirbas, O.; Danisman, N. Prediction of Preeclampsia by First Trimester Combined Test and Simple Complete Blood Count Parameters. J. Clin. Diagn. Res. 2015, 9, QC20-3. [Google Scholar] [CrossRef]
- Garrido-Gómez, T.; Castillo-Marco, N.; Cordero, T.; Simón, C. Decidualization resistance in the origin of preeclampsia. Am. J. Obstet. Gynecol. 2022, 226, S886–S894. [Google Scholar] [CrossRef] [PubMed]
- Fantone, S.; Mazzucchelli, R.; Giannubilo, S.R.; Ciavattini, A.; Marzioni, D.; Tossetta, G. AT-rich interactive domain 1A protein expression in normal and pathological pregnancies complicated by preeclampsia. Histochem. Cell Biol. 2020, 154, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Garbett, N.; Brock, G. Differential scanning calorimetry as a complementary diagnostic tool for the evaluation of biological samples. Biochim. Biophys. Acta BBA Gen. Subj. 2016, 1860, 981–989. [Google Scholar] [CrossRef]
- Todinova, S.; Krumova, S.; Kurtev, P.; Dimitrov, V.; Djongov, L.; Dudunkov, Z.; Taneva, S.G. Calorimetry-based profiling of blood plasma from colorectal cancer patients. Biochim. Biophys. Acta BBA Gen. Subj. 2012, 1820, 1879–1885. [Google Scholar] [CrossRef]
- Michnik, A.; Drzazga, Z.; Michalik, K.; Barczyk, A.; Santura, I.; Sozańska, E.; Pierzchała, W. Differential scanning calorimetry study of blood serum in chronic obstructive pulmonary disease. J. Therm. Anal. Calorim. 2010, 102, 57–60. [Google Scholar] [CrossRef]
- Koynova, R.; Antonova, B.; Sezanova, B.; Tenchov, B. Beneficial effect of sequential chemotherapy treatments of lung cancer patients revealed by calorimetric monitoring of blood plasma proteome denaturation. Thermochim. Acta 2018, 659, 1–7. [Google Scholar] [CrossRef]
- Kim, N.A.; Jin, J.H.; Kim, K.H.; Lim, D.G.; Cheong, H.; Kim, Y.H.; Ju, W.; Kim, S.C.; Jeong, S.H. Investigation of early and advanced stages in ovarian cancer using human plasma by differential scanning calorimetry and mass spectrometry. Arch. Pharm. Res. 2016, 39, 668–676. [Google Scholar] [CrossRef]
- Zapf, I.; Moezzi, M.; Fekecs, T.; Nedvig, K.; Lorinczy, D.; Ferencz, A. Influence of oxidative injury and monitoring of blood plasma by DSC on breast cancer patients. J. Therm. Anal. Calorim. 2016, 123, 2029–2035. [Google Scholar] [CrossRef]
- Dandé, Á.; Kocsis, B.; Lőrinczy, D. Thermal analysis of synovial fluids in different stages of osteoarthritis and after bacterial infections. J. Anal. Calorim. 2020, 142, 797–808. [Google Scholar] [CrossRef]
- Tenchov, B.; Koynova, R.; Antonova, B.; Zaharinova, S.; Abarova, S.; Tsonchev, Z.; Komsa-Penkova, R.; Momchilova, A. Blood plasma thermal behavior and protein oxidation as indicators of multiple sclerosis clinical status and plasma exchange therapy progression. Thermochim. Acta 2019, 671, 193–199. [Google Scholar] [CrossRef]
- Pultrone, L.; Schmid, R.; Waltimo, T.; Braissant, O.; Astasov-Frauenhoffer, M. Saliva profiling with differential scanning calorimetry: A feasibility study with ex vivo samples. PLoS ONE 2022, 17, e0269600. [Google Scholar] [CrossRef] [PubMed]
- Kiserud, T.; Piaggio, G.; Carroli, G.; Widmer, M.; Carvalho, J.; Neerup Jensen, L.; Giordano, D.; Cecatti, J.G.; Abdel Aleem, H.; Talegawkar, S.A.; et al. The World Health Organization Fetal Growth Charts: A Multinational Longitudinal Study of Ultrasound Biometric Measurements and Estimated Fetal Weight. PLoS Med. 2021, 18, e1003526. [Google Scholar] [CrossRef]
- Garbett, N.; Mekmaysy, C.; Helm, C.; Jenson, A.; Chaires, J. Differential scanning calorimetry of blood plasma for clinical diagnosis and monitoring. Exp. Mol. Pathol. 2009, 86, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Komsa-Penkova, R.; Danailova, A.; Krumova, S.; Georgieva, G.; Giosheva, I.; Gartcheva, L.; Iliev, I.; Gartchev, E.; Kercheva, K.; Savov, A.; et al. Altered Thermal Behavior of Blood Plasma Proteome Related to Inflammatory Cytokines in Early Pregnancy Loss. Int. J. Mol. Sci. 2022, 23, 8764. [Google Scholar] [CrossRef] [PubMed]
- Garbett, N.C.; Miller, J.J.; Jenson, A.B.; Chaires, J.B. Calorimetry outside the box: A new window into the plasma proteome. Biophys. J. 2008, 94, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Fish, D.; Brewood, G.; Kim, J.; Garbett, N.; Chaires, J.; Benight, A. Statistical analysis of plasma thermograms measured by differential scanning calorimetry. Biophys. Chem. 2010, 152, 184–190. [Google Scholar] [CrossRef]
- Haram, K.; Augensen, K.; Elsayed, S. Serum protein pattern in normal pregnancy with special reference to acute-phase reactants. Br. J. Obs. Gynaecol. 1983, 90, 139–145. [Google Scholar] [CrossRef]
- Joseph, J.C.; Baker, C.; Sprang, M.L.; Bermes, E.W. Changes in Plasma Proteins During Pregnancy. Ann. Clin. Lab. Sci. 1978, 8, 130–141. [Google Scholar] [PubMed]
- De Haas, S.; Ghossein-Doha, C.; Van Kuijk, S.M.J.; Van Drongelen, J.; Spaanderman, M.E.A. Physiological adaptation of maternal plasma volume during pregnancy: A systematic review and meta-analysis. Ultrasound Obs. Gynecol. 2017, 49, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef]
- Belinskaia, D.A.; Voronina, P.A.; Shmurak, V.I.; Jenkins, R.O.; Goncharov, N.V. Serum Albumin in Health and Disease: Esterase, Antioxidant, Transporting and Signaling Properties. Int. J. Mol. Sci. 2021, 22, 10318. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Chen, H.; Chen, H. Reduced plasma adiponectin and elevated leptin in pre-eclampsia. Int. J. Gynaecol. Obstet. Off. Organ. Int. Fed. Gynaecol. Obstetrics. 2007, 98, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Muttukrishna, S.; North, R.A.; Morris, J.; Schellenberg, J.C.; Taylor, R.S.; Asselin, J.; Ledger, W.; Groome, N.; Redman, C.W.G. Serum inhibin A and activin A are elevated prior to the onset of pre-eclampsia. Hum. Reprod. 2000, 15, 1640–1645. [Google Scholar] [CrossRef] [PubMed]
- Bersinger, N.A.; Smárason, A.K.; Muttukrishna, S.; Groome, N.P.; Redman, C.W. Women with preeclampsia have increased serum levels of pregnancy-associated plasma protein A (PAPP-A), inhibin A, activin A and soluble E-selectin. Hypertens. Pregnancy 2003, 22, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef]
- Chiarello, D.I.; Abad, C.; Rojas, D.; Toledo, F.; Vázquez, C.M.; Mate, A.; Sobrevia, L.; Marín, R. Oxidative stress: Normal pregnancy versus preeclampsia. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165354. [Google Scholar] [CrossRef] [PubMed]
- Zerón, H.M.; Parada-Flores, A.; Chávez, A.A.; García, M.V.D. Oxidative stress in preeclampsia, more than enzymes. Rev. Latinoam. Hipertens. 2013, 8, 25–28. [Google Scholar]
- Matheson, N.R.; Wong, P.S.; Travis, J. Enzymatic inactivation of human alpha-1-proteinase inhibitor by neutrophil myeloperoxidase. Biochem. Biophys. Res. Commun. 1979, 88, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Gathiram, P.; Moodley, J. Pre-eclampsia: Its pathogenesis and pathophysiolgy. Cardiovasc. J. Afr. 2016, 27, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Hypertension in Pregnancy: Diagnosis and Management|NICE› Guidance› Resources. Available online: https://www.nice.org.uk (accessed on 25 June 2019).
- Ryan, M.T.; Chopra, R.K. Paradoxical effect of fatty acid on steroid–albumin interaction. Biochim. Biophys. Acta 1976, 427, 337–349. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Reference Values for Pregnant Women | NPC (N = 19) | PC (N = 14) | PE (N = 27) |
|---|---|---|---|---|
| Maternal age (years) | 29.8 ± 5.5 | 27.3 ± 3.4 | 31.2 ± 4.5 | |
| Mean BP (systolic/diastolic) | 110 ± 7/74 ± 5 | 104 ± 6/72 ± 3 | 155 ± 11/104 ± 7 | |
| Gestational week (interval) | - | 27–38 | 27–36 | |
| GA at diagnostic of PE | - | - | 29.3 ± 3.2 | |
| GA at delivery | - | 39.0 ± 1.09 | 33.9 ± 4.6 | |
| Body weight (kg) | 57 ± 8 | 87 ± 7 | 92 ± 5 | |
| BMI (kg/m2) | 21.4 ± 1.5 | 31.1 ± 1.8 | 32.8 ± 1.6 | |
| Newborn weight (g) | - | 3376 ± 349 | 2144 ± 999 | |
| Proteinuria (mg in 24-h urine collection) | - | - | 1090 ± 0.5 | |
| TP (g/L) | 63–84 | 71.45 ± 3.11 | 68.34 ± 3.52 | 65.36 ± 7.42 |
| HAS (g/L) | 35–50 | 47.1 ± 1.77 | 39.54 ± 3.68 | 37.60 ± 4.79 |
| Fg (g/L) | 2.90–6.50 | 3.1 ± 0.56 | 5.32 ± 0.58 | 5.27 ± 1.22 |
| CRP (mg/L) (interval) | 0.5–5.0 | 0.37–1.12 | 5.1–19.9 | 0.93–95.35 |
| Platelet Count × 109/L | 146–429 | 289 ± 73 | 254 ± 60 | 236 ± 99 |
| ASAT (U/L) | 4–32 | 21.1 ± 2.7 | 23.2 ± 3.4 | 19.9 ± 7.3 |
| ALAT (U/L) | 3–30 | 18.1 ± 3.0 | 14.9 ± 1.5 | 16.9 ± 8.3 |
| Groups | cPFg (J·g−1·K−1) | TmHSA (°C) | cPHSA (J·g−1·K−1) | cPIgs (J·g−1·K−1) | cPHSA/cPIgs | ∆Hcal (J·g−1) | TFM (°C) | P | r | ρ |
|---|---|---|---|---|---|---|---|---|---|---|
| NPC (19) | 0.08 ± 0.016 | 61.5 ± 0.6 | 1.54 ± 0.17 | 0.79 ± 0.1 | 1.94 ± 0.2 | 17.6 ± 0.5 | 65.2 ± 0.4 | |||
| PC (14) | 0.15 ± 0.017 * | 63.0 ± 0.3 * | 1.39 ± 0.07 | 1.10 ± 0.11 * | 1.29 ± 0.16 * | 19.7 ± 0.8 * | 66.4 ± 0.3 * | - | - | - |
| PE1 (5) | 0.15 ± 0.01 | 61.6 ± 0.3 ** | 1.44 ± 0.13 | 0.97 ± 0.1 | 1.49 ± 0.17 | 19.0 ± 0.24 | 65.9 ± 0.4 | 0.92 ± 0.04 | 0.86 ± 0.03 | 0.89 ± 0.03 |
| PE2 (11) | 0.14 ± 0.02 | 64.2 ± 0.2 ** | 1.21 ± 0.12 ** | 1.0 ± 0.11 | 1.22 ± 0.07 | 18.0 ± 0.17 ** | 66.8 ± 0.2 | 0.76 ± 0.04 | 0.88 ± 0.04 | 0.79 ± 0.04 |
| PE3 (9) | 0.14 ± 0.03 | 66.3 ± 0.2 ** | 1.14 ± 0.14 ** | 1.2 ± 0.13 | 0.95 ± 0.06 ** | 18.0 ± 0.18 ** | 67.5 ± 0.3 ** | 0.51 ± 0.03 | 0.80 ± 0.02 | 0.71 ± 0.02 |
| Case18 | 0.18 ** | 65.9 | 0.95 | 1.1 | 0.86 | 16.9 ** | 67.4 ** | 0.58 | 0.76 | 0.71 |
| Case27 | 0.13 | - | - | 1.16 | - | 15.6 ** | 66.7 | 0.60 | 0.75 | 0.70 |
| Groups | TmHSA (°C) | cPHSA (J·g−1·K−1) | cPIgs (J·g−1·K−1) | cPHSA/cPIgs | ∆Hcal (J·g−1) | TFM (°C) | P | r | ρ |
|---|---|---|---|---|---|---|---|---|---|
| PC (14) | 63.0 ± 0.3 | 1.39 ± 0.07 | 1.10 ± 0.11 | 1.29 ± 0.16 | 19.7 ± 0.8 | 66.4 ± 0.3 | |||
| PE31 (5) | 65.6 ± 0.2 * | 1.19 ± 0.08 | 1.23 ± 0.091 * | 0.96 ± 0.05 * | 18.0 ± 0.19 * | 67.2 ± 0.2 * | 0.55 ± 0.05 | 0.81 ± 0.02 | 0.73 ± 0.02 |
| PE32 (4) | 65.6 ± 0.2 * | 0.98 ± 0.07 | 1.14 ± 0.06 * | 0.86 ± 0.06 * | 17.5 ± 0.14 * | 68.4 ± 0.4 * | 0.49 ± 0.03 | 0.78 ± 0.02 | 0.69 ± 0.02 |
| Groups | HAS (%) | α1 (%) | α2 (%) | β1 (%) | β2 (%) | γ (%) |
|---|---|---|---|---|---|---|
| NPC | 62.9 ± 1.8 | 3.8 ± 0.2 | 8.3 ± 1.2 | 6.1 ± 0.6 | 4.5 ± 1.2 | 14.5 ± 1.1 |
| PC | 49.0 ± 2.5 * | 7.5 ± 1.4 * | 15.2 ± 1.7 * | 8.7 ± 1.1 * | 8.6 ± 1.3 * | 10.9 ± 1.7 * |
| PE1 | 49.2 ± 4.0 | 6.7 ± 1.5 | 14.8 ± 2.1 | 9.8 ± 2.0 | 7.2 ± 1.4 | 12.3 ± 2.7 |
| PE2 | 51.7 ± 3.1 | 8.2 ± 1.1 | 12.8 ± 2.2 ** | 8.6 ± 2.1 | 7.7 ± 0.9 | 10.7 ± 2.5 |
| PE31 | 50.3 ± 3.1 | 7.2 ± 0.9 | 15.7 ± 1.8 | 9.4 ± 1.4 | 7.7 ± 1.8 | 6.7 ± 1.6 ** |
| PE32 | 51.5 ± 2.4 | 7.75 ± 0.7 | 13.8 ± 1.9 | 8.5 ± 0.8 | 8.8 ± 1.7 | 10.2 ± 1.7 |
| Case18 Case27 | 35.86 ** 51.34 | 9.59 ** 7.14 | 25.18 ** 15.69 | 8.21 11.39 ** | 8.15 7.72 | 13.01 ** 6.71 ** |
| Samples | ΔH (J·g−1) | TFM (°C) | cPHSA/cPIgs |
|---|---|---|---|
| NPC | 17.4 ± 0.5 | 65.8 ± 0.6 | 1.69 ± 0.06 |
| NPC + 25 mM H2O2 | 14.7 ± 0.3 | 67.7 ± 0.4 | 1.20 ± 0.05 |
| NPC + 100 mM H2O2 | 13.5 ± 0.6 | 67.9 ± 0.7 | 0.91 ± 0.09 |
| Samples | Tm (°C) | ΔH (J·g−1) |
|---|---|---|
| HSA | 62.0 | 9.8 |
| HSA + 25 mM of H2O2 | 61.7 | 8.4 |
| HSA + 100 mM of H2O2 | 61.4 | 4.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komsa-Penkova, R.; Krumova, S.; Langari, A.; Giosheva, I.; Gartcheva, L.; Danailova, A.; Topalova, L.; Stoyanova, T.; Strijkova, V.; Savov, A.; et al. Blood Plasma Calorimetric Profiles of Women with Preeclampsia: Effect of Oxidative Stress. Antioxidants 2023, 12, 1032. https://doi.org/10.3390/antiox12051032
Komsa-Penkova R, Krumova S, Langari A, Giosheva I, Gartcheva L, Danailova A, Topalova L, Stoyanova T, Strijkova V, Savov A, et al. Blood Plasma Calorimetric Profiles of Women with Preeclampsia: Effect of Oxidative Stress. Antioxidants. 2023; 12(5):1032. https://doi.org/10.3390/antiox12051032
Chicago/Turabian StyleKomsa-Penkova, Regina, Sashka Krumova, Ariana Langari, Ina Giosheva, Lidia Gartcheva, Avgustina Danailova, Lora Topalova, Tanya Stoyanova, Velichka Strijkova, Alexey Savov, and et al. 2023. "Blood Plasma Calorimetric Profiles of Women with Preeclampsia: Effect of Oxidative Stress" Antioxidants 12, no. 5: 1032. https://doi.org/10.3390/antiox12051032