

Drought and Oxidative Stress in Flax (Linum usitatissimum L.) Entails Harnessing Non-Canonical Reference Gene for Precise Quantification of qRT-PCR Gene Expression

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Identification, Retrieval and Primer Designing
2.2. Plant Material, Total RNA Isolation, and cDNA Synthesis
2.3. qRT-PCR Conditions and Analyses
2.4. Statistical Analyses of RGs Expression, Stability, and Their Validation through Drought-Responsive Genes
3. Results
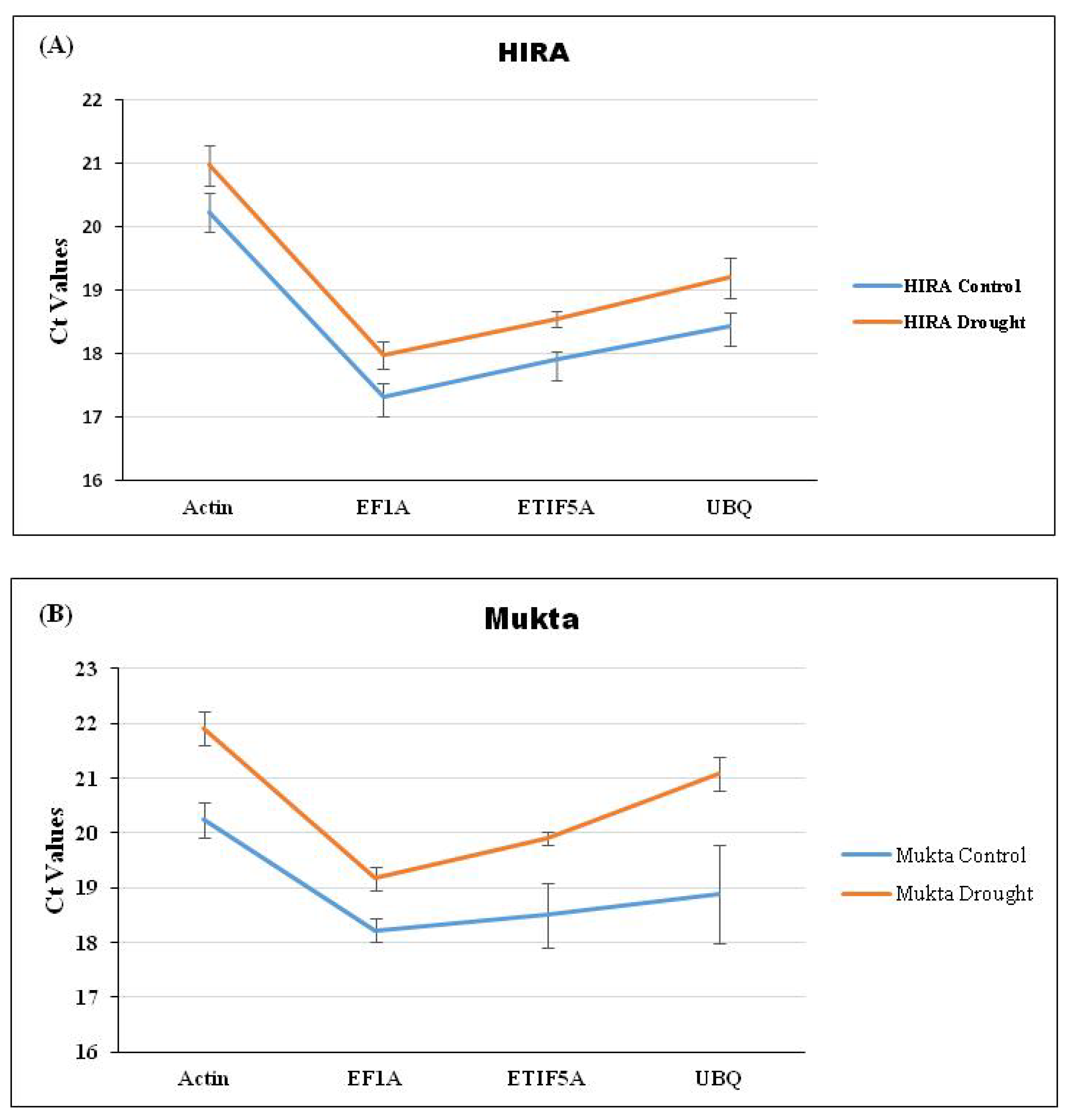
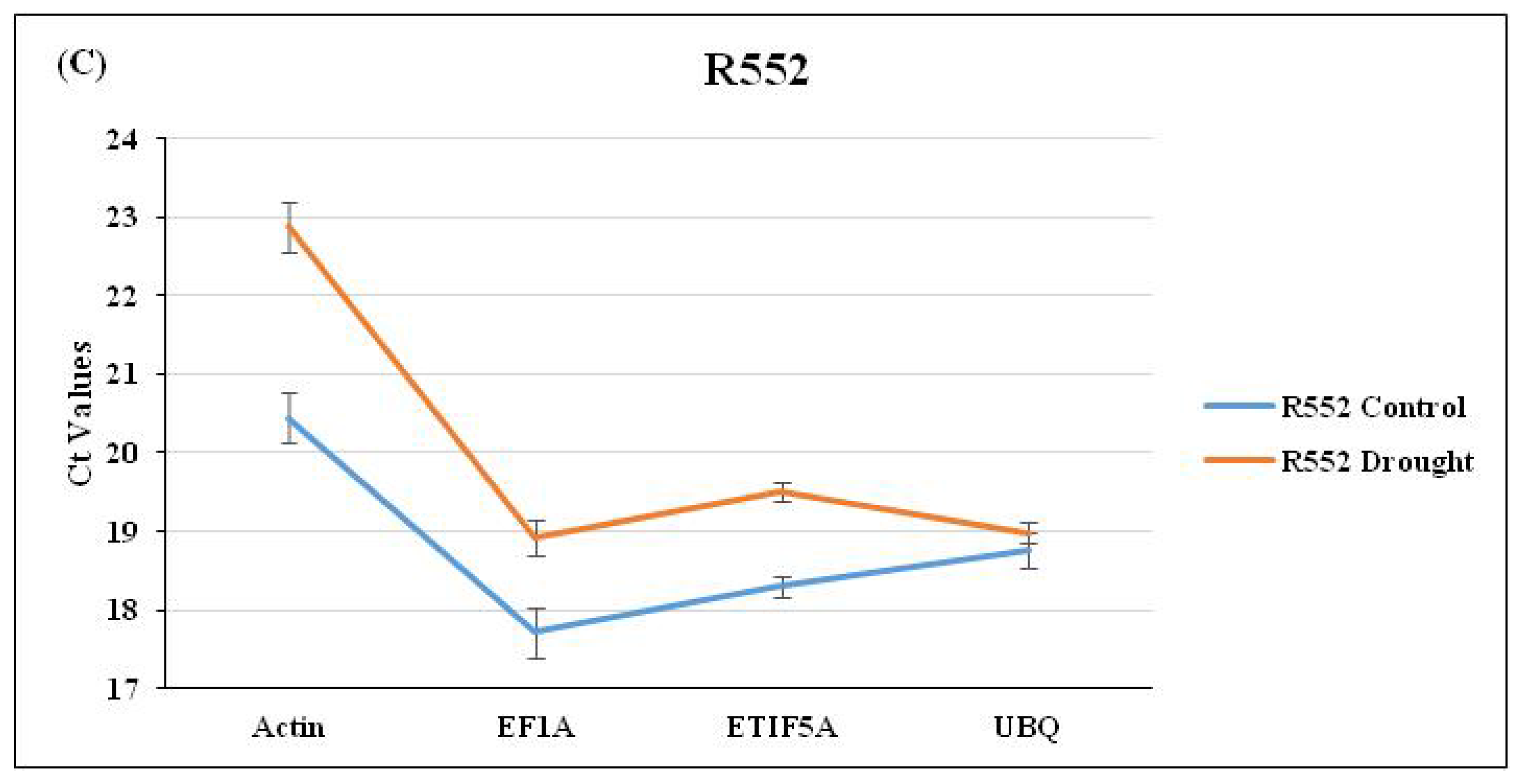
3.1. Selection of Candidate RGs and DRGs and Their Amplification
3.2. Analysis of Stability of Expression of Selected RGs
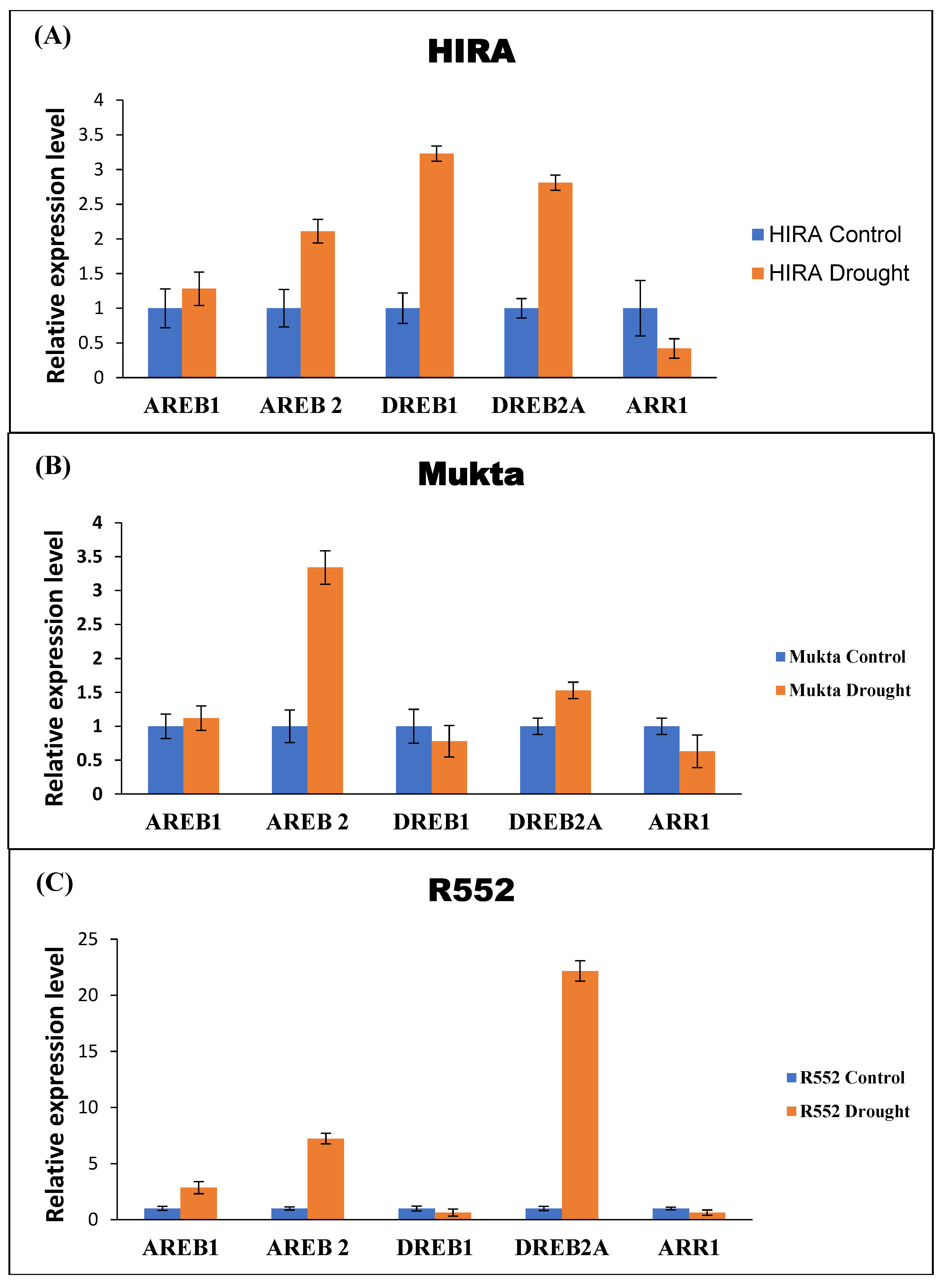
3.3. Validation of RGs through Expression of DRGs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huis, R.; Hawkins, S.; Neutelings, G. Selection of Reference Genes for Quantitative Gene Expression Normalization in Flax (Linum usitatissimum L.). BMC Plant Biol. 2010, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Allaby, R.G.; Peterson, G.W.; Merriwether, D.A.; Fu, Y.-B. Evidence of the Domestication History of Flax (Linum usitatissimum L.) from Genetic Diversity of the Sad2 Locus. Theor. Appl. Genet. 2005, 112, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hobson, N.; Galindo, L.; Zhu, S.; Shi, D.; McDill, J.; Yang, L.; Hawkins, S.; Neutelings, G.; Datla, R.; et al. The Genome of Flax (Linum usitatissimum) Assembled de Novo from Short Shotgun Sequence Reads. Plant J. 2012, 72, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Dash, P.K. Precise Method of in Situ Drought Stress Induction in Flax (Linum usitatissimum) for RNA Isolation towards down-Stream Analysis. Ann. Agric. Res. 2015, 36, 10–17. [Google Scholar]
- Dash, P.K.; Rai, R.; Mahato, A.K.; Gaikwad, K.; Singh, N.K. Transcriptome Landscape at Different Developmental Stages of a Drought Tolerant Cultivar of Flax (Linum usitatissimum). Front. Chem. 2017, 5, 82. [Google Scholar] [CrossRef]
- Singh, K.K.; Mridula, D.; Rehal, J.; Barnwal, P. Flaxseed: A Potential Source of Food, Feed and Fiber. Crit. Rev. Food Sci. Nutr. 2011, 51, 210–222. [Google Scholar] [CrossRef]
- Tawfik, R.S.; Badr, A.; Sammour, R.; Ibrahim, U.; Matter, M.; Sakr, M. Improvement of Flax Drought Tolerance Using Gene Transfer. Plant Tissue Cult. Biotechnol. 2016, 26, 197–207. [Google Scholar] [CrossRef]
- Dash, P.K.; Cao, Y.; Jailani, A.K.; Gupta, P.; Venglat, P.; Xiang, D.; Rai, R.; Sharma, R.; Thirunavukkarasu, N.; Abdin, M.Z.; et al. Genome-Wide Analysis of Drought Induced Gene Expression Changes in Flax (Linum usitatissimum). GM Crop. Food 2014, 5, 106–119. [Google Scholar] [CrossRef]
- de Carvalho, M.H.C. Drought Stress and Reactive Oxygen Species: Production, Scavenging and Signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription Factors and Plants Response to Drought Stress: Current Understanding and Future Directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef]
- Rabbani, M.A.; Maruyama, K.; Abe, H.; Khan, M.A.; Katsura, K.; Ito, Y.; Yoshiwara, K.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Monitoring Expression Profiles of Rice Genes under Cold, Drought, and High-Salinity Stresses and Abscisic Acid Application Using CDNA Microarray and RNA Gel-Blot Analyses. Plant Physiol. 2003, 133, 1755–1767. [Google Scholar] [CrossRef]
- Dean, G.; Cao, Y.; Xiang, D.; Provart, N.J.; Ramsay, L.; Ahad, A.; White, R.; Selvaraj, G.; Datla, R.; Haughn, G. Analysis of Gene Expression Patterns during Seed Coat Development in Arabidopsis. Mol. Plant 2011, 4, 1074–1091. [Google Scholar] [CrossRef]
- Aprile, A.; Mastrangelo, A.M.; De Leonardis, A.M.; Galiba, G.; Roncaglia, E.; Ferrari, F.; De Bellis, L.; Turchi, L.; Giuliano, G.; Cattivelli, L. Transcriptional Profiling in Response to Terminal Drought Stress Reveals Differential Responses along the Wheat Genome. BMC Genom. 2009, 10, 279. [Google Scholar] [CrossRef]
- Pereira, S.S.; Guimarães, F.C.M.; Carvalho, J.F.C.; Stolf-Moreira, R.; Oliveira, M.C.N.; Rolla, A.A.P.; Farias, J.R.B.; Neumaier, N.; Nepomuceno, A.L. Transcription Factors Expressed in Soybean Roots under Drought Stress. Genet. Mol. Res. 2011, 10, 3689–3701. [Google Scholar] [CrossRef]
- Hayano-Kanashiro, C.; Calderón-Vázquez, C.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Simpson, J. Analysis of Gene Expression and Physiological Responses in Three Mexican Maize Landraces under Drought Stress and Recovery Irrigation. PLoS ONE 2009, 4, e7531. [Google Scholar] [CrossRef]
- Lamblin, F.; Saladin, G.; Dehorter, B.; Cronier, D.; Grenier, E.; Lacoux, J.; Bruyant, P.; Lainé, E.; Chabbert, B.; Girault, F.; et al. Overexpression of a Heterologous Sam Gene Encoding S-Adenosylmethionine Synthetase in Flax (Linum usitatissimum) Cells: Consequences on Methylation of Lignin Precursors and Pectins. Physiol. Plant. 2001, 112, 223–232. [Google Scholar] [CrossRef]
- Kesiraju, K.; Tyagi, S.; Mukherjee, S.; Rai, R.; Singh, N.K.; Sreevathsa, R.; Dash, P.K. An Apical Meristem-Targeted in Planta Transformation Method for the Development of Transgenics in Flax (Linum usitatissimum): Optimization and Validation. Front. Plant Sci. 2021, 11, 562056. [Google Scholar] [CrossRef]
- Caillot, S.; Rosiau, E.; Laplace, C.; Thomasset, B. Influence of Light Intensity and Selection Scheme on Regeneration Time of Transgenic Flax Plants. Plant Cell Rep. 2009, 28, 359–371. [Google Scholar] [CrossRef]
- Dash, P.K.; Gupta, P.; Jailani, A.K.; Rai, R. Hydropenia induces expression of drought responsive genes (DRGs) erd1, hat, plD-δ, and zfa in Linum usitatissimum L. Ind. J. Exp. Biol. 2018, 56, 743–749. [Google Scholar]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to Reference Gene Selection for Quantitative Real-Time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef]
- Gachon, C.; Mingam, A.; Charrier, B. Real-Time PCR: What Relevance to Plant Studies? J. Exp. Bot. 2004, 55, 1445–1454. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yang, H.; Chen, M.; Song, X.; Yu, C.; Zhao, Y.; Wu, Y. Reference Gene Selection for Quantitative Real-Time PCR of Mycelia from Lentinula edodes under High-Temperature Stress. BioMed Res. Int. 2018, 2018, 1670328. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Stürzenbaum, S.R.; Kille, P. Control Genes in Quantitative Molecular Biological Techniques: The Variability of Invariance. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 130, 281–289. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034. [Google Scholar] [CrossRef]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.-R. Genome-Wide Identification and Testing of Superior Reference Genes for Transcript Normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef]
- González-Aguilera, K.L.; Saad, C.F.; Montes, R.A.C.; Alves-Ferreira, M.; de Folter, S. Selection of Reference Genes for Quantitative Real-Time RT-PCR Studies in Tomato Fruit of the Genotype MT-Rg1. Front. Plant Sci. 2016, 7, 1386. [Google Scholar] [CrossRef]
- Magneschi, L.; Kudahettige, R.L.; Alpi, A.; Perata, P. Expansin Gene Expression and Anoxic Coleoptile Elongation in Rice Cultivars. J. Plant Physiol. 2009, 166, 1576–1580. [Google Scholar] [CrossRef]
- Day, A.; Dehorter, B.; Neutelings, G.; Czeszak, X.; Chabbert, B.; Belingheri, L.; David, H. Caffeoyl-Coenzyme A 3-O-Methyltransferase Enzyme Activity, Protein and Transcript Accumulation in Flax (Linum usitatissimum) Stem during Development. Physiol. Plant. 2001, 113, 275–284. [Google Scholar] [CrossRef]
- Wróbel, M.; Zebrowski, J.; Szopa, J. Polyhydroxybutyrate Synthesis in Transgenic Flax. J. Biotechnol. 2004, 107, 41–54. [Google Scholar] [CrossRef]
- Rao, S.; Abdel-Reheem, M.; Bhella, R.; McCracken, C.; Hildebrand, D. Characteristics of High α-Linolenic Acid Accumulation in Seed Oils. Lipids 2008, 43, 749–755. [Google Scholar] [CrossRef]
- Chen, C.; Xie, T.; Ye, S.; Jensen, A.B.; Eilenberg, J. Selection of Reference Genes for Expression Analysis in the Entomophthoralean Fungus Pandora Neoaphidis. Braz. J. Microbiol. 2016, 47, 259–265. [Google Scholar] [CrossRef]
- Brentner, L.B.; Mukherji, S.T.; Merchie, K.M.; Yoon, J.M.; Schnoor, J.L.; Aken, B.V. Expression of Glutathione S-Transferases in Poplar Trees (Populus Trichocarpa) Exposed to 2,4,6-Trinitrotoluene (TNT). Chemosphere 2008, 73, 657–662. [Google Scholar] [CrossRef]
- Bracha-Drori, K.; Shichrur, K.; Lubetzky, T.C.; Yalovsky, S. Functional Analysis of Arabidopsis Postprenylation CaaX Processing Enzymes and Their Function in Subcellular Protein Targeting. Plant Physiol. 2008, 148, 119–131. [Google Scholar] [CrossRef]
- Bomal, C.; Bedon, F.; Caron, S.; Mansfield, S.D.; Levasseur, C.; Cooke, J.E.K.; Blais, S.; Tremblay, L.; Morency, M.-J.; Pavy, N.; et al. Involvement of Pinus Taeda MYB1 and MYB8 in Phenylpropanoid Metabolism and Secondary Cell Wall Biogenesis: A Comparative in Planta Analysis. J. Exp. Bot. 2008, 59, 3925–3939. [Google Scholar] [CrossRef]
- Aquea, F.; Gutiérrez, F.; Medina, C.; Arce-Johnson, P. A Novel Otubain-like Cysteine Protease Gene Is Preferentially Expressed during Somatic Embryogenesis in Pinus Radiata. Mol. Biol. Rep. 2008, 35, 567–573. [Google Scholar] [CrossRef]
- Gutierrez, L.; Conejero, G.; Castelain, M.; Guénin, S.; Verdeil, J.-L.; Thomasset, B.; Van Wuytswinkel, O. Identification of New Gene Expression Regulators Specifically Expressed during Plant Seed Maturation. J. Exp. Bot. 2006, 57, 1919–1932. [Google Scholar] [CrossRef]
- Roach, M.J.; Deyholos, M.K. Microarray Analysis of Flax (Linum usitatissimum L.) Stems Identifies Transcripts Enriched in Fibre-Bearing Phloem Tissues. Mol. Genet. Genom. 2007, 278, 149–165. [Google Scholar] [CrossRef]
- Mallona, I.; Lischewski, S.; Weiss, J.; Hause, B.; Egea-Cortines, M. Validation of Reference Genes for Quantitative Real-Time PCR during Leaf and Flower Development in Petunia Hybrida. BMC Plant Biol. 2010, 10, 4. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-S.; Han, N.-N.; Li, J.-H.; Huang, G.-H.; Wan, H. Selection of Reference Genes for Expression Analysis Using RT-QPCR in the Dissemination System of Heliothis Virescens Ascovirus 3 h (HvAV-3h). Sci. Rep. 2017, 7, 7045. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene Networks Involved in Drought Stress Response and Tolerance. J. Exp. Bot. 2006, 58, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Ito, Y.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Networks in Response to Abiotic Stresses in Arabidopsis and Grasses. Plant Physiol. 2009, 149, 88–95. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The Transcriptional Regulatory Network in the Drought Response and Its Crosstalk in Abiotic Stress Responses Including Drought, Cold, and Heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef]
- Piehler, A.P.; Grimholt, R.M.; Øvstebø, R.; Berg, J.P. Gene Expression Results in Lipopolysaccharide-Stimulated Monocytes Depend Significantly on the Choice of Reference Genes. BMC Immunol. 2010, 11, 21. [Google Scholar] [CrossRef]
- Gupta, P.; Dash, P.K.; Sanjay, T.D.; Pradhan, S.K.; Sreevathsa, R.; Rai, R. Cloning and Molecular Characterization of the PhlD Gene Involved in the Biosynthesis of “Phloroglucinol”, a Compound with Antibiotic Properties from Plant Growth Promoting Bacteria Pseudomonas Spp. Antibiotics 2023, 12, 260. [Google Scholar] [CrossRef]
- Sahu, S.; Gupta, P.; Gowtham, T.P.; Yogesh, K.S.; Sanjay, T.D.; Singh, A.; Duong, H.V.; Pradhan, S.K.; Bisht, D.S.; Singh, N.K.; et al. Generation of High-Value Genomic Resource in Rice: A “Sub-Genomic Library” of Low-Light-Tolerant Rice Cultivar Swarnaprabha. Biology 2023, 12, 428. [Google Scholar] [CrossRef]
- Gupta, P.; Rai, R.; Dash, P.K. Isolation, Cloning and Characterization of PhlA Gene from an Indigenous Pseudomonas Strain from Indian Soil. Int. J. Trop. Agric. 2015, 33, 3195–3200. [Google Scholar]
- Dash, P.K.; Gupta, P.; Panwar, B.S.; Rai, R. Isolation, Cloning and Characterization of PhlB Gene from an Indian Strain of Gram Negative Soil Bacteria Pseudomonas Fluorescens. Indian J. Exp. Biol. 2020, 58, 412–419. [Google Scholar] [CrossRef]
- Gupta, P.; Karthik, K.; Sreevathsa, R.; Rai, R.; Dash, P.K. Cloning and Characterization of Phloroglucinol Biosynthetic Gene PhlC from AnIndian Strain of Pseudomonas Fluorescens. Indian J. Exp. Biol. 2022, 60, 607–614. [Google Scholar] [CrossRef]
- Rai, R. Isolation, Characterization and Evaluation of the Biocontrol Potential of Pseudomonas protegens RS-9 against Ralstonia solanacearum in Tomato. Indian J. Exp. Biol. 2017, 55, 595–603. [Google Scholar]
- Dash, P.K.; Gupta, P.; Pradhan, S.K.; Shasany, A.K.; Rai, R. Analysis of Homologous Regions of Small RNAs MIR397 and MIR408 Reveals the Conservation of Microsynteny among Rice Crop-Wild Relatives. Cells 2022, 11, 3461. [Google Scholar] [CrossRef]
- Pradhan, K.C.; Pandit, E.; Mohanty, S.P.; Moharana, A.; Sanghamitra, P.; Meher, J.; Jena, B.K.; Dash, P.K.; Behera, L.; Mohapatra, P.M.; et al. Development of Broad Spectrum and Durable Bacterial Blight Resistant Variety through Pyramiding of Four Resistance Genes in Rice. Agronomy 2022, 12, 1903. [Google Scholar] [CrossRef]
- Bastia, R.; Pandit, E.; Sanghamitra, P.; Barik, S.R.; Nayak, D.K.; Sahoo, A.; Moharana, A.; Meher, J.; Dash, P.K.; Raj, R.; et al. Association Mapping for Quantitative Trait Loci Controlling Superoxide Dismutase, Flavonoids, Anthocyanins, Carotenoids, γ-Oryzanol and Antioxidant Activity in Rice. Agronomy 2022, 12, 3036. [Google Scholar] [CrossRef]
- Mohapatra, S.; Barik, S.R.; Dash, P.K.; Lenka, D.; Pradhan, K.C.; Raj, K.R.R.; Mohanty, S.P.; Mohanty, M.R.; Sahoo, A.; Jena, B.K.; et al. Molecular Breeding for Incorporation of Submergence Tolerance and Durable Bacterial Blight Resistance into the Popular Rice Variety ‘Ranidhan’. Biomolecules 2023, 13, 198. [Google Scholar] [CrossRef]
- Nguyen, K.H.; Ha, C.V.; Nishiyama, R.; Watanabe, Y.; Leyva-González, M.A.; Fujita, Y.; Tran, U.T.; Li, W.; Tanaka, M.; Seki, M.; et al. Arabidopsis Type B Cytokinin Response Regulators ARR1, ARR10, and ARR12 Negatively Regulate Plant Responses to Drought. Proc. Natl. Acad. Sci. USA 2016, 113, 3090–3095. [Google Scholar] [CrossRef]
- Kozera, B.; Rapacz, M. Reference Genes in Real-Time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar] [CrossRef]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An Optimized Grapevine RNA Isolation Procedure and Statistical Determination of Reference Genes for Real-Time RT-PCR during Berry Development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef]
- Nicot, N.; Hausman, J.-F.; Hoffmann, L.; Evers, D. Housekeeping Gene Selection for Real-Time RT-PCR Normalization in Potato during Biotic and Abiotic Stress. J. Exp. Bot. 2005, 56, 2907–2914. [Google Scholar] [CrossRef]
- Jain, M.; Nijhawan, A.; Tyagi, A.K.; Khurana, J.P. Validation of Housekeeping Genes as Internal Control for Studying Gene Expression in Rice by Quantitative Real-Time PCR. Biochem. Biophys. Res. Commun. 2006, 345, 646–651. [Google Scholar] [CrossRef]
- Jian, B.; Liu, B.; Bi, Y.; Hou, W.; Wu, C.; Han, T. Validation of Internal Control for Gene Expression Study in Soybean by Quantitative Real-Time PCR. BMC Mol. Biol. 2008, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R.; Le, H.; Caldwell, C.; Browning, K.S. Analysis of Translation Elongation Factors from Wheat during Development and Following Heat Shock. Biochem. Biophys. Res. Commun. 1998, 245, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Bonato, M.; Corrà, F.; Bellio, M.; Guidolin, L.; Tallandini, L.; Irato, P.; Santovito, G. PFAS Environmental Pollution and Antioxidant Responses: An Overview of the Impact on Human Field. Int. J. Environ. Res. Public Health 2020, 17, 8020. [Google Scholar] [CrossRef] [PubMed]
- Pacchini, S.; Piva, E.; Schumann, S.; Irato, P.; Pellegrino, D.; Santovito, G. An Experimental Study on Antioxidant Enzyme Gene Expression in Trematomus newnesi (Boulenger, 1902) Experimentally Exposed to Perfluoro-Octanoic Acid. Antioxidants 2023, 12, 352. [Google Scholar] [CrossRef]
- Shivakumara, T.N.; Sreevathsa, R.; Dash, P.K.; Sheshshayee, M.S.; Papolu, P.K.; Rao, U.; Tuteja, N.; UdayaKumar, M. Overexpression of Pea DNA Helicase 45 (PDH45) Imparts Tolerance to Multiple Abiotic Stresses in Chili (Capsicum annuum L.). Sci. Rep. 2017, 7, 2760. [Google Scholar] [CrossRef]
- Tyagi, S.; Rathinam, M.; Dokka, N.; Chaudhary, N.; Satish, L.; Dash, P.K.; Shasany, A.K.; Sreevathsa, R. Cajanus Platycarpus Flavonoid 3′5′ Hydroxylase_2 (CpF3′5′H_2) Confers Resistance to Helicoverpa Armigera by Modulating Total Polyphenols and Flavonoids in Transgenic Tobacco. Int. J. Mol. Sci. 2023, 24, 1755. [Google Scholar] [CrossRef]
- Brunner, A.M.; Yakovlev, I.A.; Strauss, S.H. Validating internal controls for quantitative plant gene expression studies. BMC Plant Biol. 2004, 4, 14. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant Responses to Drought, Salinity and Extreme Temperatures: Towards Genetic Engineering for Stress Tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 Are Master Transcription Factors That Cooperatively Regulate ABRE-Dependent ABA Signaling Involved in Drought Stress Tolerance and Require ABA for Full Activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two Transcription Factors, DREB1 and DREB2, with an EREBP/AP2 DNA Binding Domain Separate Two Cellular Signal Transduction Pathways in Drought- and Low-Temperature-Responsive Gene Expression, Respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. A Novel Cis-Acting Element in an Arabidopsis Gene Is Involved in Responsiveness to Drought, Low-Temperature, or High-Salt Stress. Plant Cell 1994, 6, 251–264. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arabidopsis | Linum usitatissimum | |||||
|---|---|---|---|---|---|---|
| Gene Name | Gene Id | Gene Id | E-Value | Bit Score | %Identity | CDS Length (in bp) |
| AREB1 | AT1G45249 | Lus10006489 | 5.00 × 10−86 | 315 | 50.35 | 1209 |
| AREB2/ABF4 | AT3G19290 | Lus10014066 | 2.00 × 10−104 | 376 | 50.76 | 1266 |
| DREB1/CBF | AT4G25490 | Lus10031657 | 9.00 × 10−59 | 223 | 60.59 | 696 |
| DREB2A | AT5G05410 | Lus10034902 | 2.00 × 10−49 | 193 | 42.29 | 753 |
| ARR1 | AT3G16857 | Lus10037719 | 6.00 × 10−177 | 618 | 52.16 | 2121 |
| Sr. No. | Name | Sequence (5′-3′) |
|---|---|---|
| 1 | LuActin_qRTFwd | TCCAGGCCGTTCTTTCTCTA |
| 2 | LuActin_qRTRev | CTGTAAGGTCACGACCAGCA |
| 3 | LuEF1A_qRTFwd | GCTGCCAACTTCACATCTCA |
| 4 | LuEF1A_qRTRev | GATCGCCTGTCAATCTTGGT |
| 5 | LuETIF5A_qRTFwd | TGCCACATGTGAACCGTACT |
| 6 | LuETIF5A_qRTRev | CTTTACCCTCAGCAAATCCG |
| 7 | LuUBQ_qRTFwd | CTCCGTGGAGGTATGCAGAT |
| 8 | LuUBQ_qRTRev | TTCCTTGTCCTGGATCTTCG |
| 9 | LuAREB1_qRTFwd | ATCAGATGGGATTGGGAAGAGC |
| 10 | LuAREB1_qRTRev | GGAGGCAGAAGAGAATGCTCA |
| 11 | LuAREB2_qRTFwd | TGTTGAGAGAAGACACAGAAGG |
| 12 | LuAREB2_qRTRev | GGAGATGAATGAAGAACTGGAG |
| 13 | LuDREB1_qRTFwd | CGGCGGTGGAAGCGACGAC |
| 14 | LuDREB1_qRTRev | GCCGGGGCTTTTGACGAGCA |
| 15 | LuDREB2A_qRTFwd | AGACGTTAAGGACTATGAGTGGC |
| 16 | LuDREB2A_qRTRev | GGCTTGCTGTTAGGGGATAATA |
| 17 | LuARR1_qRTFwd | CAAGGCAATATTGAGGTGGGCTC |
| 18 | LuARR1_qRTRev | CTCTGCTGCTGGCGTGGAACA |
| Group | HMR | HM | MR | HR |
|---|---|---|---|---|
| Gene Name | Stability Score | |||
| Actin | 0.020 | 0.012 | 0.030 | 0.019 |
| ETIF5A | 0.015 | 0.016 | 0.018 | 0.008 |
| EF1A | 0.007 | 0.007 | 0.008 | 0.005 |
| UBQ | 0.025 | 0.016 | 0.033 | 0.023 |
| Group | Best Combination of Two Genes | Stability Score |
|---|---|---|
| HMR | EF1a and ETIF5A | 0.010 |
| EF1a and Actin | 0.016 | |
| ETIF5A and Actin | 0.018 | |
| HM | EF1a and ETIF5A | 0.009 |
| EF1a and Actin | 0.011 | |
| ETIF5A and Actin | 0.019 | |
| MR | EF1a and ETIF5A | 0.012 |
| EF1a and Actin | 0.021 | |
| ETIF5A and Actin | 0.021 | |
| HR | EF1a and ETIF5A | 0.006 |
| EF1a and Actin | 0.014 | |
| ETIF5A and Actin | 0.026 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dash, P.K.; Rai, R.; Pradhan, S.K.; Shivaraj, S.M.; Deshmukh, R.; Sreevathsa, R.; Singh, N.K. Drought and Oxidative Stress in Flax (Linum usitatissimum L.) Entails Harnessing Non-Canonical Reference Gene for Precise Quantification of qRT-PCR Gene Expression. Antioxidants 2023, 12, 950. https://doi.org/10.3390/antiox12040950
Dash PK, Rai R, Pradhan SK, Shivaraj SM, Deshmukh R, Sreevathsa R, Singh NK. Drought and Oxidative Stress in Flax (Linum usitatissimum L.) Entails Harnessing Non-Canonical Reference Gene for Precise Quantification of qRT-PCR Gene Expression. Antioxidants. 2023; 12(4):950. https://doi.org/10.3390/antiox12040950
Chicago/Turabian StyleDash, Prasanta K., Rhitu Rai, Sharat Kumar Pradhan, Sheelavanta Matha Shivaraj, Rupesh Deshmukh, Rohini Sreevathsa, and Nagendra K. Singh. 2023. "Drought and Oxidative Stress in Flax (Linum usitatissimum L.) Entails Harnessing Non-Canonical Reference Gene for Precise Quantification of qRT-PCR Gene Expression" Antioxidants 12, no. 4: 950. https://doi.org/10.3390/antiox12040950