
Combination Therapy of Radiation and Hyperthermia, Focusing on the Synergistic Anti-Cancer Effects and Research Trends


Abstract
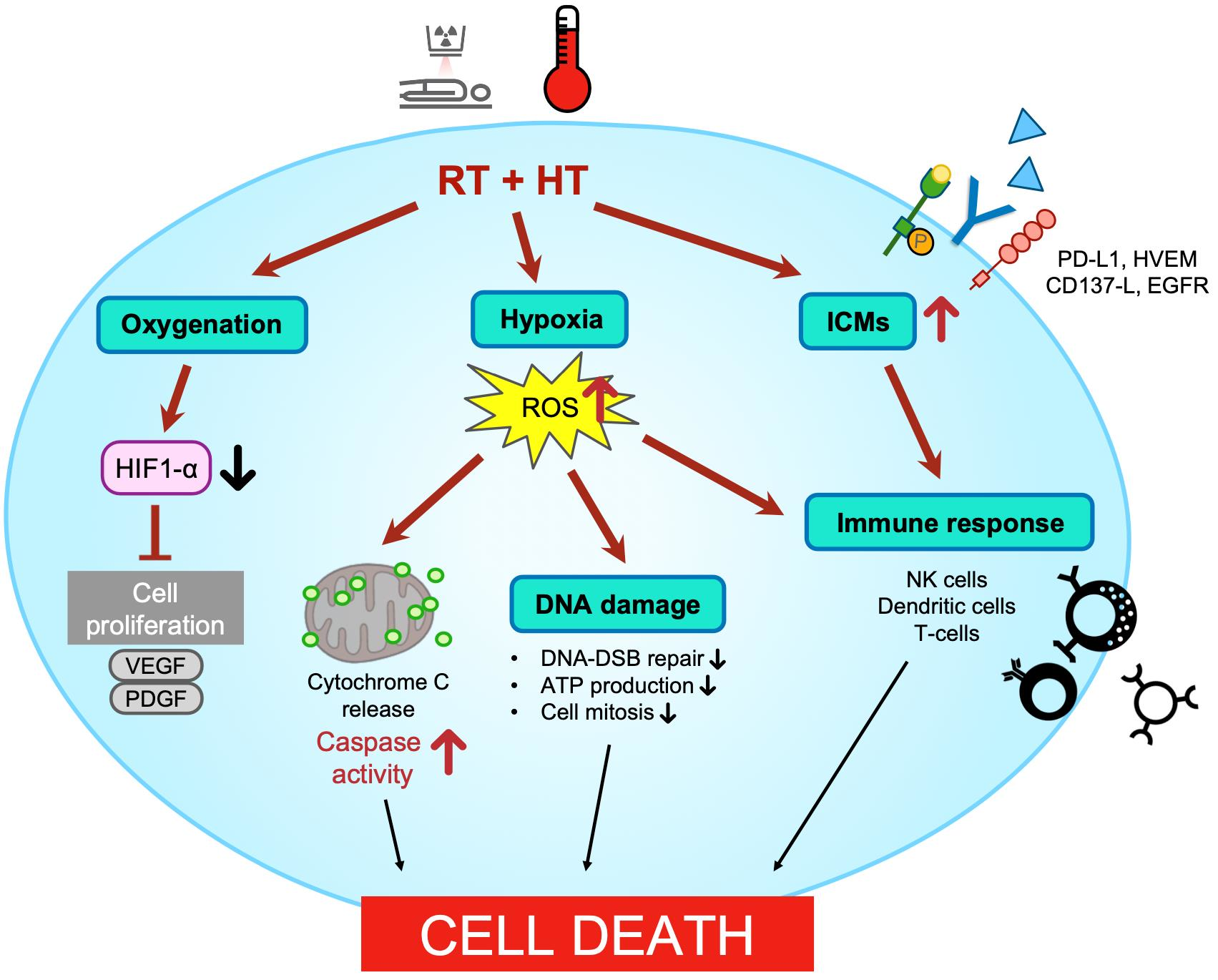
:
1. Introduction
2. Anti-Cancer Effects of RT + HT Treatment
3. Physiological Changes Induced by RT + HT
3.1. Oxygenation
3.2. DNA Damage
3.3. Cell Cycle Arrest
3.4. Apoptosis
3.4.1. Intrinsic Apoptosis
3.4.2. Extrinsic Apoptosis and Other Pathways
4. Immune Response
4.1. Cytokines and Antibodies
4.2. NK Cells
4.3. Immune Checkpoint Molecules (ICMs)
5. Discussion
5.1. Discovery of Effects by the Combination of RT + HT (Phase 1)
5.2. Exploration of the Physiological Changes Following the Demonstration of Effects (Phase 2)
5.3. Immune Response (Phase 3)
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Beyer, A.M.; Bonini, M.G.; Moslehi, J. Cancer therapy-induced cardiovascular toxicity: Old/new problems and old drugs. Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H164–H167. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.; Zaidi, S.F. Treating cancer with heat: Hyperthermia as promising strategy to enhance apoptosis. J. Pak. Med. Assoc. 2013, 63, 504–508. [Google Scholar] [PubMed]
- Ba, M.; Long, H.; Wang, S.; Wu, Y.; Zhang, B.; Yan, Z.; Yu, F.; Cui, S. Hyperthermia enhances radiosensitivity of colorectal cancer cells through ROS inducing autophagic cell death. J. Cell. Biochem. 2018, 119, 3763–3774. [Google Scholar] [CrossRef] [PubMed]
- Dunne, M.; Regenold, M.; Allen, C. Hyperthermia can alter tumor physiology and improve chemo- and radio-therapy efficacy. Adv. Drug Deliv. Rev. 2020, 163–164, 98–124. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.R.; Puric, E.; Klingbiel, D.; Gomez, S.; Bodis, S. Hyperthermia and Radiation Therapy in Locoregional Recurrent Breast Cancers: A Systematic Review and Meta-analysis. Int. J. Radiat. Oncol. Biol. Phys. 2016, 94, 1073–1087. [Google Scholar] [CrossRef]
- Habash, R.W. Therapeutic hyperthermia. Handb. Clin. Neurol. 2018, 157, 853–868. [Google Scholar]
- Yi, G.Y.; Kim, M.J.; Kim, H.I.; Park, J.; Baek, S.H. Hyperthermia Treatment as a Promising Anti-Cancer Strategy: Therapeutic Targets, Perspective Mechanisms and Synergistic Combinations in Experimental Approaches. Antioxidants 2022, 11, 625. [Google Scholar] [CrossRef]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.-W. Cancer and Radiation Therapy: Current Advances and Future Directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Di Lalla, V.; Chaput, G.; Williams, T.; Sultanem, K. Radiotherapy Side Effects: Integrating a Survivorship Clinical Lens to Better Serve Patients. Curr. Oncol. 2020, 27, 107–112. [Google Scholar] [CrossRef]
- Wang, J.; Han, Y.; Li, Y.; Zhang, F.; Cai, M.; Zhang, X.; Chen, J.; Ji, C.; Ma, J.; Xu, F. Targeting Tumor Physical Microenvironment for Improved Radiotherapy. Small Methods 2022, 6, e2200570. [Google Scholar] [CrossRef] [PubMed]
- Jarosz-Biej, M.; Smolarczyk, R.; Cichon, T.; Kulach, N. Tumor Microenvironment as A “Game Changer” in Cancer Radiotherapy. Int. J. Mol. Sci. 2019, 20, 3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huilgol, N.G.; Gupta, S.; Sridhar, C.R. Hyperthermia with radiation in the treatment of locally advanced head and neck cancer: A report of randomized trial. J. Cancer Res. Ther. 2010, 6, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Toraya-Brown, S.; Fiering, S. Local tumour hyperthermia as immunotherapy for metastatic cancer. Int. J. Hyperth. 2014, 30, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Besse, H.C.; Bos, C.; Zandvliet, M.M.J.M.; van der Wurff-Jacobs, K.; Moonen, C.T.W.; Deckers, R. Triggered radiosensitizer delivery using thermosensitive liposomes and hyperthermia improves efficacy of radiotherapy: An in vitro proof of concept study. PLoS ONE 2018, 13, e0204063. [Google Scholar] [CrossRef] [Green Version]
- Le Guevelou, J.; Chirila, M.E.; Achard, V.; Guillemin, P.C.; Lorton, O.; Uiterwijk, J.W.E.; Dipasquale, G.; Salomir, R.; Zilli, T. Combined hyperthermia and radiotherapy for prostate cancer: A systematic review. Int. J. Hyperth. 2022, 39, 547–556. [Google Scholar] [CrossRef]
- Brewer, W.G.; Turrel, J.M. Radiotherapy and hyperthermia in the treatment of fibrosarcomas in the dog. J. Am. Veter Med. Assoc. 1982, 181, 146–150. [Google Scholar]
- Campos, S.N.; Górriz, F.J.M.; Ferrer, R.B.; Vitoria, A.H.; Ramírez, A.C.; Grasa, F.F.; Garrido, J.M.R. Evolution of weight and survival of C3H mice with solid, subcutaneous sarcoma 180, treated with radiotherapy, hyperthermia and a combination of hyperthermia and radiotherapy. Rev. Esp. Oncol. 1983, 30, 379–398. [Google Scholar]
- Yamashita, M. Radiosensitizing effect of combined radiotherapy, hyperthermia and misonidazole on C3H mouse FM3A tumor. Nihon Igaku Hoshasen Gakkai Zasshi 1984, 44, 1181–1188. [Google Scholar]
- Dewhirst, M.W.; Sim, D.A.; Forsyth, K.; Grochowski, K.J.; Wilson, S.; Bicknell, E. Local control and distant metastases in primary canine malignant melanomas treated with hyperthermia and/or radiotherapy. Int. J. Hyperth. 1985, 1, 219–234. [Google Scholar] [CrossRef]
- Legorreta, R.A.; Denman, D.L.; Kelley, M.C.; Lewis, G.C. Use of hyperthermia and radiotherapy in treatment of a large mast cell sarcoma in a dog. J. Am. Veter Med. Assoc. 1988, 193, 1545–1548. [Google Scholar]
- Patrício, M.B.; Vilhena, M.; Soares, J. Morphologic and morphometric studies on tumor necrosis produced by radiotherapy, and hyperthermia singly and in combination. J. Surg. Oncol. 1989, 42, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, K.; Watanabe, T. Effects of Hyperthermia, Radiotherapy and Thermoradiotherapy on Tumor Microvascular Permeability. Acta Pathol. Jpn. 1990, 40, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Ruifrok, A.C.; Levendag, P.C.; Lakeman, R.F.; Deurloo, I.K.; Visser, A.G. Combined treatment with interstitial hyperthermia and interstitial radiotherapy in an animal tumor model. Int. J. Radiat Oncol. Biol. Phys. 1991, 20, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Sougawa, M.; Urano, M. Significance of additive heat effect in the therapeutic gain factor in combined hyperthermia and radiotherapy: Murine tumor response and foot reaction. Int. J. Radiat. Oncol. Biol. Phys. 1991, 21, 1561–1568. [Google Scholar] [CrossRef]
- Nishimura, Y.; Urano, M. Timing and sequence of hyperthermia in fractionated radiotherapy of a murine fibrosarcoma. Int. J. Radiat. Oncol. Biol. Phys. 1993, 27, 605–611. [Google Scholar] [CrossRef]
- Devi, P.U.; Guruprasad, K. Influence of clamping-induced ischemia and reperfusion on the response of a mouse melanoma to radiation and hyperthermia. Int. J. Hyperth. 2001, 17, 357–367. [Google Scholar] [CrossRef]
- Ressel, A.; Schmitt, O.; Weiss, C.; Feyerabend, T. Therapeutic outcome and side-effects after radiotherapy, chemotherapy and/or hyperthermia treatment of head and neck tumour xenografts. Eur. J. Cancer 2002, 38, 594–601. [Google Scholar] [CrossRef]
- Rao, B.S.S.; Devi, P.U. Response of S 180 murine tumor to bleomycin in combination with radiation and hyperthermia using micronucleus assay: A multimodality approach for therapeutic augmentation. Indian J. Exp. Biol. 2005, 43, 596–600. [Google Scholar]
- Kalthur, G.; Pathirissery, U.D. Enhancement of the Response of B16F1 Melanoma to Fractionated Radiotherapy and Prolongation of Survival by Withaferin A and/or Hyperthermia. Integr. Cancer Ther. 2010, 9, 370–377. [Google Scholar] [CrossRef]
- Franken, N.A.P.; Barendsen, G.W. Enhancement of radiation effectiveness by hyperthermia and incorporation of halogenated pyrimidines at low radiation doses as compared with high doses: Implications for mechanisms. Int. J. Radiat. Biol. 2014, 90, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Alya, G.; Ekhtiar, A.; Saour, G. Effects of lethal dose of gamma-radiation and partial body hyperthermia on Wistar rats. Int. J. Hyperth. 2015, 31, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Borasi, G.; Nahum, A.; Paulides, M.M.; Powathil, G.; Russo, G.; Fariselli, L.; Lamia, D.; Cirincione, R.; Forte, G.I.; Borrazzo, C.; et al. Fast and high temperature hyperthermia coupled with radiotherapy as a possible new treatment for glioblastoma. J. Ther. Ultrasound 2016, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masunaga, S.-I.; Tano, K.; Sanada, Y.; Sakurai, Y.; Tanaka, H.; Suzuki, M.; Kondo, N.; Watanabe, T.; Takata, T.; Maruhashi, A.; et al. Effect of Tirapazamine, Metformin or Mild Hyperthermia on Recovery from Radiation-Induced Damage in Pimonidazole-Unlabeled Quiescent Tumor Cells. World J. Oncol. 2017, 8, 137–146. [Google Scholar] [CrossRef] [Green Version]
- IJff, M.; van Oorschot, B.; Oei, A.L.; Krawczyk, P.M.; Rodermond, H.M.; Stalpers, L.J.A.; Kok, H.P.; Crezee, J.; Franken, N.A.P. Enhancement of Radiation Effectiveness in Cervical Cancer Cells by Combining Ionizing Radiation with Hyperthermia and Molecular Targeting Agents. Int. J. Mol. Sci. 2018, 19, 2420. [Google Scholar] [CrossRef] [Green Version]
- Rajaee, Z.; Khoei, S.; Mahdavi, S.R.; Ebrahimi, M.; Shirvalilou, S.; Mahdavian, A. Evaluation of the effect of hyperthermia and electron radiation on prostate cancer stem cells. Radiat. Environ. Biophys. 2018, 57, 133–142. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.; Corde, S.; Lerch, M.; Rosenfeld, A.; Jackson, M.; Tehei, M. First in vitro evidence of modulated electro-hyperthermia treatment performance in combination with megavoltage radiation by clonogenic assay. Sci. Rep. 2018, 8, 16608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, B.; Kim, S.; Cho, W.; Kim, J.K.; Kim, Y.A.; Kim, S.; Wu, H.G. Quantitative Estimation of the Equivalent Radiation Dose Escalation using Radiofrequency Hyperthermia in Mouse Xenograft Models of Human Lung Cancer. Sci. Rep. 2019, 9, 3942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brüningk, S.C.; Rivens, I.; Box, C.; Oelfke, U.; ter Haar, G. 3D tumour spheroids for the prediction of the effects of radiation and hyperthermia treatments. Sci. Rep. 2020, 10, 1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunti, S.; Hoke, A.T.K.; Vu, K.; London, N.R., Jr. Organoid and Spheroid Tumor Models: Techniques and Applications. Cancers 2021, 13, 874. [Google Scholar] [CrossRef]
- Ishiguro, T.; Ohata, H.; Sato, A.; Yamawaki, K.; Enomoto, T.; Okamoto, K. Tumor-derived spheroids: Relevance to cancer stem cells and clinical applications. Cancer Sci. 2017, 108, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Zhang, X.; Unger, M.; Patties, I.; Melzer, A.; Landgraf, L. Focused Ultrasound-Induced Cavitation Sensitizes Cancer Cells to Radiation Therapy and Hyperthermia. Cells 2020, 9, 2595. [Google Scholar] [CrossRef] [PubMed]
- Elming, P.B.; Sorensen, B.S.; Spejlborg, H.; Overgaard, J.; Horsman, M.R. Does the combination of hyperthermia with low LET (linear energy transfer) radiation induce anti-tumor effects equivalent to those seen with high LET radiation alone? Int. J. Hyperth. 2021, 38, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol. Cancer 2019, 18, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elming, P.B.; Sørensen, B.S.; Oei, A.L.; Franken, N.A.P.; Crezee, J.; Overgaard, J.; Horsman, M.R. Hyperthermia: The Optimal Treatment to Overcome Radiation Resistant Hypoxia. Cancers 2019, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, Y.; Urano, M. The effect of hyperthermia on reoxygenation during the fractionated radiotherapy of two murine tumors, FSa-II and MCa. Int. J. Radiat. Oncol. Biol. Phys. 1994, 29, 141–148. [Google Scholar] [CrossRef]
- Vujaskovic, Z.; Poulson, J.M.; Gaskin, A.A.; Thrall, D.E.; Page, R.L.; Charles, H.C.; MacFall, J.R.; Brizel, D.M.; Meyer, R.E.; Prescott, D.M.; et al. Temperature-dependent changes in physiologic parameters of spontaneous canine soft tissue sarcomas after combined radiotherapy and hyperthermia treatment. Int. J. Radiat. Oncol. Biol. Phys. 2000, 46, 179–185. [Google Scholar] [CrossRef]
- Ressel, A.; Weiss, C.; Feyerabend, T. Tumor oxygenation after radiotherapy, chemotherapy, and/or hyperthermia predicts tumor free survival. Int. J. Radiat. Oncol. Biol. Phys. 2001, 49, 1119–1125. [Google Scholar] [CrossRef]
- Thrall, D.E.; LaRue, S.M.; Pruitt, A.F.; Case, B.; Dewhirst, M.W. Changes in tumour oxygenation during fractionated hyperthermia and radiation therapy in spontaneous canine sarcomas. Int. J. Hyperth. 2006, 22, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, N.; Zarei, L.; Galeh, H.E.G.; Motlagh, B.M. Assessment of synergistic effect of combining hyperthermia with irradiation and calcium carbonate nanoparticles on proliferation of human breast adenocarcinoma cell line (MCF-7 cells). Artif. Cells Nanomed. Biotechnol. 2018, 46, 364–372. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Kim, M.-S.; Kim, H.-J.; Lee, E.; Jeong, J.-H.; Park, I.; Jeong, Y.K.; Jang, W.I. Role of HIF-1α in response of tumors to a combination of hyperthermia and radiation in vivo. Int. J. Hyperth. 2018, 34, 276–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeghi, N.; Kok, R.J.; Bos, C.; Zandvliet, M.; Geerts, W.J.C.; Storm, G.; Moonen, C.T.W.; Lammers, T.; Deckers, R. Hyperthermia-triggered release of hypoxic cell radiosensitizers from temperature-sensitive liposomes improves radiotherapy efficacy in vitro. Nanotechnology 2019, 30, 264001. [Google Scholar] [CrossRef] [PubMed]
- Sijens, P.E.; Bovée, W.M.; Seijkens, D.; Koole, P.; Los, G.; Van Rijssel, R.H. Murine mammary tumor response to hyperthermia and radiotherapy evaluated by in vivo 31P-nuclear magnetic resonance spectroscopy. Cancer Res. 1987, 47 Pt 1, 6467–6473. [Google Scholar] [PubMed]
- Multhoff, G.; Habl, G.; Combs, S.E. Rationale of hyperthermia for radio(chemo)therapy and immune responses in patients with bladder cancer: Biological concepts, clinical data, interdisciplinary treatment decisions and biological tumour imaging. Int. J. Hyperth. 2016, 32, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, L.-P.; Sun, H.-T.; Wang, P.; Wang, J.; Zhou, J.-H.; Cao, R.-Q.; Yue, L.; Chen, Y.-G.; Shen, F.-R. Hyperthermia enhances the efficacy of chemotherapeutic drugs in heat-sensitive cells through interfering with DNA damage repair. Ann. Transl. Med. 2022, 10, 463. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Stavridi, E.; Leeper, D.B.; Iliakis, G. Effects of hyperthermia on p53 protein expression and activity. J. Cell. Physiol. 2002, 190, 365–374. [Google Scholar] [CrossRef]
- Li, X.; Heyer, W.-D. Homologous recombination in DNA repair and DNA damage tolerance. Cell Res. 2008, 18, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Miki, Y. Role of BRCA1 and BRCA2 as regulators of DNA repair, transcription, and cell cycle in response to DNA damage. Cancer Sci. 2004, 95, 866–871. [Google Scholar] [CrossRef]
- Ohnishi, T. The role of the p53 molecule in cancer therapies with radiation and/or hyperthermia. J. Cancer Res. Ther. 2005, 1, 147–150. [Google Scholar] [CrossRef]
- Masunaga, S.-I.; Nagata, K.; Suzuki, M.; Kashino, G.; Kinashi, Y.; Ono, K. Inhibition of repair of radiation-induced damage by mild temperature hyperthermia, referring to the effect on quiescent cell populations. Radiat. Med. 2007, 25, 417–425. [Google Scholar] [CrossRef]
- Masunaga, S.-I.; Tano, K.; Sanada, Y.; Suzuki, M.; Takahashi, A.; Ohnishi, K.; Ono, K. Effects of p53 Status of Tumor Cells and Combined Treatment with Mild Hyperthermia, Wortmannin or Caffeine on Recovery from Radiation-Induced Damage. World J. Oncol. 2019, 10, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genet, S.C.; Fujii, Y.; Maeda, J.; Kaneko, M.; Genet, M.D.; Miyagawa, K.; Kato, T.A. Hyperthermia inhibits homologous recombination repair and sensitizes cells to ionizing radiation in a time- and temperature-dependent manner. J. Cell. Physiol. 2013, 228, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Bergs, J.; Oei, A.L.; Cate, R.T.; Rodermond, H.M.; Stalpers, L.J.; Barendsen, G.W.; Franken, N.A. Dynamics of chromosomal aberrations, induction of apoptosis, BRCA2 degradation and sensitization to radiation by hyperthermia. Int. J. Mol. Med. 2016, 38, 243–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Oorschot, B.; Granata, G.; Di Franco, S.; Ten Cate, R.; Rodermond, H.M.; Todaro, M.; Medema, J.P.; Franken, N.A. Targeting DNA double strand break repair with hyperthermia and DNA-PKcs inhibition to enhance the effect of radiation treatment. Oncotarget 2016, 7, 65504–65513. [Google Scholar] [CrossRef] [Green Version]
- Son, B.; Jeon, J.; Lee, S.; Kim, H.; Kang, H.; Youn, H.; Jo, S.; Youn, B. Radiotherapy in combination with hyperthermia suppresses lung cancer progression via increased NR4A3 and KLF11 expression. Int. J. Radiat. Biol. 2019, 95, 1696–1707. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Johansson, P.; Torchinsky, D.; Lin, Y.-L.; Öz, R.; Ebenstein, Y.; Hammarsten, O.; Westerlund, F. Quantifying DNA damage induced by ionizing radiation and hyperthermia using single DNA molecule imaging. Transl. Oncol. 2020, 13, 100822. [Google Scholar] [CrossRef]
- Khurshed, M.; Prades-Sagarra, E.; Saleh, S.; Sminia, P.; Wilmink, J.W.; Molenaar, R.J.; Crezee, H.; van Noorden, C.J.F. Hyperthermia as a Potential Cornerstone of Effective Multimodality Treatment with Radiotherapy, Cisplatin and PARP Inhibitor in IDH1-Mutated Cancer Cells. Cancers 2022, 14, 6228. [Google Scholar] [CrossRef]
- Lee, Y.S.; Ahmed, M.B.; Alghamdi, A.A.A.; Islam, S.U.; Ahsan, H. The Complex Roles of DNA Repair Pathways, Inhibitors, Hyperthermia, and Contact Inhibition in Cell Cycle Halts. Mini Rev. Med. Chem. 2022, 23, 514–529. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Wu, Q.; Wu, Z.B.; Zhang, J.J.; Zhu, L.C.; Yang, Y.; Ma, S.L.; Zhang, S.R. Microwave hyperthermia promotes caspase-3-dependent apoptosis and induces G2/M checkpoint arrest via the ATM pathway in non-small cell lung cancer cells. Int. J. Oncol. 2018, 53, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Teyssier, F.; Bay, J.O.; Dionet, C.; Verrelle, P. Cell cycle regulation after exposure to ionizing radiation. Bull. Cancer 1999, 86, 345–357. [Google Scholar]
- Yuguchi, T.; Saito, M.; Yokoyama, Y.; Saito, T.; Nagata, T.; Sakamoto, T.; Tsukada, K. Combined use of hyperthermia and irradiation cause antiproliferative activity and cell death to human esophageal cell carcinoma cells--mainly cell cycle examination. Hum. Cell 2002, 15, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as anticancer mechanism: Function and dysfunction of its modulators and targeted therapeutic strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamas-Din, A.; Kale, J.; Leber, B.; Andrews, D.W. Mechanisms of Action of Bcl-2 Family Proteins. Cold Spring Harb. Perspect. Biol. 2013, 5, a008714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plati, J.; Bucur, O.; Khosravi-Far, R. Dysregulation of apoptotic signaling in cancer: Molecular mechanisms and therapeutic opportunities. J. Cell. Biochem. 2008, 104, 1124–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Zheng, K.; Fan, Q.; Jiang, X.; Xiong, D. Hyperthermia exposure induces apoptosis and inhibits proliferation in HCT116 cells by upregulating miR-34a and causing transcriptional activation of p53. Exp. Ther. Med. 2017, 14, 5379–5386. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.; Reyna, D.E.; Gitego, N.; Kopp, F.; Zhou, H.; Miranda-Roman, M.A.; Nordstrøm, L.U.; Narayanagari, S.-R.; Chi, P.; Vilar, E.; et al. Co-targeting of BAX and BCL-XL proteins broadly overcomes resistance to apoptosis in cancer. Nat. Commun. 2022, 13, 1199. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ding, Y.; Ye, N.; Wild, C.; Chen, H.; Zhou, J. Direct Activation of Bax Protein for Cancer Therapy. Med. Res. Rev. 2016, 36, 313–341. [Google Scholar] [CrossRef] [Green Version]
- Liang, H. Change in expression of apoptosis genes after hyperthermia, chemotherapy and radiotherapy in human colon cancer transplanted into nude mice. World J. Gastroenterol. 2007, 13, 4365–4371. [Google Scholar] [CrossRef]
- Talaat, R.M.; Abo-Zeid, T.M.; Abo-Elfadl, M.T.; El Maadawy, E.; Hassanin, M.M. Combined Hyperthermia and Radiation Therapy for Treatment of Hepatocellular Carcinoma. Asian Pac. J. Cancer Prev. 2019, 20, 2303–2310. [Google Scholar] [CrossRef]
- Singh, P.; Eley, J.; Saeed, A.; Bhandary, B.; Mahmood, N.; Chen, M.; Dukic, T.; Mossahebi, S.; Rodrigues, D.B.; Mahmood, J.; et al. Effect of hyperthermia and proton beam radiation as a novel approach in chordoma cells death and its clinical implication to treat chordoma. Int. J. Radiat. Biol. 2021, 97, 1675–1686. [Google Scholar] [CrossRef]
- Sharifnia, T.; Wawer, M.J.; Chen, T.; Huang, Q.-Y.; Weir, B.A.; Sizemore, A.; Lawlor, M.A.; Goodale, A.; Cowley, G.S.; Vazquez, F.; et al. Small-molecule targeting of brachyury transcription factor addiction in chordoma. Nat. Med. 2019, 25, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Shariati, M.; Meric-Bernstam, F. Targeting AKT for cancer therapy. Expert Opin. Investig. Drugs 2019, 28, 977–988. [Google Scholar] [CrossRef]
- Man, J.; Shoemake, J.D.; Ma, T.; Rizzo, A.E.; Godley, A.R.; Wu, Q.; Mohammadi, A.M.; Bao, S.; Rich, J.N.; Yu, J.S. Hyperthermia Sensitizes Glioma Stem-like Cells to Radiation by Inhibiting AKT Signaling. Cancer Res. 2015, 75, 1760–1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skitzki, J.J.; Repasky, E.A.; Evans, S.S. Hyperthermia as an immunotherapy strategy for cancer. Curr. Opin. Investig. Drugs 2009, 10, 550–558. [Google Scholar] [PubMed]
- Wan Mohd Zawawi, W.F.A.; Hibma, M.H.; Salim, M.I.; Jemon, K. Hyperthermia by near infrared radiation induced immune cells activation and infiltration in breast tumor. Sci. Rep. 2021, 11, 10278. [Google Scholar] [CrossRef] [PubMed]
- Khandia, R.; Munjal, A.; Iqbal, H.M.N.; Dhama, K. Heat Shock Proteins: Therapeutic Perspectives in Inflammatory Disorders. Recent Patents Inflamm. Allergy Drug Discov. 2017, 10, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.; Ruffell, B. Dendritic Cells and Cancer Immunity. Trends Immunol. 2016, 37, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Schildkopf, P.; Holmer, R.; Sieber, R.; Ott, O.J.; Janko, C.; Mantel, F.; Frey, B.; Fietkau, R.; Gaipl, U.S. Hyperthermia in combination with X-irradiation induces inflammatory forms of cell death. Autoimmunity 2009, 42, 311–313. [Google Scholar] [CrossRef] [PubMed]
- Schildkopf, P.; Frey, B.; Mantel, F.; Ott, O.J.; Weiss, E.-M.; Sieber, R.; Janko, C.; Sauer, R.; Fietkau, R.; Gaipl, U.S. Application of hyperthermia in addition to ionizing irradiation fosters necrotic cell death and HMGB1 release of colorectal tumor cells. Biochem. Biophys. Res. Commun. 2010, 391, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Schildkopf, P.; Frey, B.; Ott, O.J.; Rubner, Y.; Multhoff, G.; Sauer, R.; Fietkau, R.; Gaipl, U.S. Radiation combined with hyperthermia induces HSP70-dependent maturation of dendritic cells and release of pro-inflammatory cytokines by dendritic cells and macrophages. Radiother. Oncol. 2011, 101, 109–115. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Xi, X.; Hu, B.; Zhao, L.; Liao, Y.; Tang, J. Effects of magnetic induction hyperthermia and radiotherapy alone or combined on a murine 4T1 metastatic breast cancer model. Int. J. Hyperth. 2011, 27, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Werthmöller, N.; Frey, B.; Rückert, M.; Lotter, M.; Fietkau, R.; Gaipl, U.S. Combination of ionising radiation with hyperthermia increases the immunogenic potential of B16-F10 melanoma cells in vitro and in vivo. Int. J. Hyperth. 2016, 32, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, J.; Alexander, A.A.; Samanta, S.; Kamlapurkar, S.; Singh, P.; Saeed, A.; Carrier, F.; Cao, X.; Shukla, H.D.; Vujaskovic, Z. A Combination of Radiotherapy, Hyperthermia, and Immunotherapy Inhibits Pancreatic Tumor Growth and Prolongs the Survival of Mice. Cancers 2020, 12, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayanc, B.E.; Beachy, S.H.; Ostberg, J.R.; Repasky, E.A. Dissecting the role of hyperthermia in natural killer cell mediated anti-tumor responses. Int. J. Hyperth. 2008, 24, 41–56. [Google Scholar] [CrossRef]
- Walle, T.; Kraske, J.A.; Liao, B.; Lenoir, B.; Timke, C.; von Bohlen Und Halbach, E.; Tran, F.; Griebel, P.; Albrecht, D.; Ahmed, A.; et al. Radiotherapy orchestrates natural killer cell dependent antitumor immune responses through CXCL8. Sci. Adv. 2022, 8, eabh4050. [Google Scholar] [CrossRef] [PubMed]
- Sordo-Bahamonde, C.; Lorenzo-Herrero, S.; Payer, A.R.; Gonzalez, S.; López-Soto, A. Mechanisms of Apoptosis Resistance to NK Cell-Mediated Cytotoxicity in Cancer. Int. J. Mol. Sci. 2020, 21, 3726. [Google Scholar] [CrossRef] [PubMed]
- Hietanen, T.; Kapanen, M.; Kellokumpu-Lehtinen, P.-L. Restoring Natural Killer Cell Cytotoxicity After Hyperthermia Alone or Combined with Radiotherapy. Anticancer. Res. 2016, 36, 555–563. [Google Scholar] [PubMed]
- Hietanen, T.; Kapanen, M.; Kellokumpu-Lehtinen, P.L. Natural Killer Cell Viability After Hyperthermia Alone or Combined with Radiotherapy with or without Cytokines. Anticancer Res. 2018, 38, 655–663. [Google Scholar] [PubMed]
- Finkel, P.; Frey, B.; Mayer, F.; Bösl, K.; Werthmöller, N.; Mackensen, A.; Gaipl, U.S.; Ullrich, E. The dual role of NK cells in antitumor reactions triggered by ionizing radiation in combination with hyperthermia. Oncoimmunology 2016, 5, e1101206. [Google Scholar] [CrossRef]
- Li, B.; Chan, H.L.; Chen, P. Immune Checkpoint Inhibitors: Basics and Challenges. Curr. Med. Chem. 2019, 26, 3009–3025. [Google Scholar] [CrossRef]
- Stoll, E.; Hader, M.; Rückert, M.; Weissmann, T.; Lettmaier, S.; Putz, F.; Hecht, M.; Fietkau, R.; Rosin, A.; Frey, B.; et al. Detailed in vitro analyses of the impact of multimodal cancer therapy with hyperthermia and radiotherapy on the immune phenotype of human glioblastoma cells. Int. J. Hyperth. 2022, 39, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.J.; Puric, E.; Eberle, B.; Datta, N.R.; Bodis, S.B. Radiotherapy for Melanoma: More than DNA Damage. Dermatol. Res. Pract. 2019, 2019, 9435389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hader, M.; Savcigil, D.P.; Rosin, A.; Ponfick, P.; Gekle, S.; Wadepohl, M.; Bekeschus, S.; Fietkau, R.; Frey, B.; Schlücker, E.; et al. Differences of the Immune Phenotype of Breast Cancer Cells after Ex Vivo Hyperthermia by Warm-Water or Microwave Radiation in a Closed-Loop System Alone or in Combination with Radiotherapy. Cancers 2020, 12, 1082. [Google Scholar] [CrossRef] [PubMed]
- Hader, M.; Streit, S.; Rosin, A.; Gerdes, T.; Wadepohl, M.; Bekeschus, S.; Fietkau, R.; Frey, B.; Schlücker, E.; Gekle, S.; et al. In Vitro Examinations of Cell Death Induction and the Immune Phenotype of Cancer Cells Following Radiative-Based Hyperthermia with 915 MHz in Combination with Radiotherapy. Cells 2021, 10, 1436. [Google Scholar] [CrossRef]
- Sengedorj, A.; Hader, M.; Heger, L.; Frey, B.; Dudziak, D.; Fietkau, R.; Ott, O.J.; Scheidegger, S.; Barba, S.M.; Gaipl, U.S.; et al. The Effect of Hyperthermia and Radiotherapy Sequence on Cancer Cell Death and the Immune Phenotype of Breast Cancer Cells. Cancers 2022, 14, 2050. [Google Scholar] [CrossRef]
- Kim, H.; Kim, D.; Kim, W.; Kim, E.; Jang, W.I.; Kim, M.-S. The Efficacy of Radiation is Enhanced by Metformin and Hyperthermia Alone or Combined Against FSaII Fibrosarcoma in C3H Mice. Radiat. Res. 2022, 198, 190–199. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Z. The history and advances in cancer immunotherapy: Understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell. Mol. Immunol. 2020, 17, 807–821. [Google Scholar] [CrossRef]

{kind=link}
| RT | HT | Cell Line and Observation Model | Classification of the Molecular Mechanism | Ref. |
|---|---|---|---|---|
| 32–48 Gy | 50 °C, 30 s 43 °C, 30 min | Oral or external nasal fibrosarcoma, ten dogs/in vivo | Necrosis | [17] |
| - | - | Murine sarcoma, sarcoma 180, C3H mice/in vivo | [18] | |
| 51.08 Gy | 43 °C, 10 min | Murine mammary carcinoma, FM3A, C3H mice/in vivo | [19] | |
| 36.8 Gy (8 × 4.6 Gy) | 43 °C, 20 min | Primary malignant melanoma, 43 dogs/in vivo | [20] | |
| 45.5 Gy (13 × 3.5 Gy) | 44 °C, 30 min | Mast cell sarcoma, a dog/in vivo | Necrosis | [21] |
| 17 Gy (2 × 8.5 Gy) | 43.5 °C, 30 min | Breast carcinoma, Tx; the sarcoma 37, S37, BALB/C male mice/in vivo | Necrosis | [22] |
| 10 Gy | 46 °C, 60 min | Shope-virus-induced skin papilloma, VX-2, rabbits/in vivo | Necrosis | [23] |
| 30 Gy | 44 °C, 30 min | Rhabdomyosarcoma, R-1, Wag/Rij female rats/in vivo | [24] | |
| 7.2 Gy/min (total dose not reported) | 45.5 °C, 10 min; followed by 41.5 °C, 60 min | Murine fibrosarcoma, FSa-II, C3Hf/Sed mice/in vivo | [25] | |
| 5.5–5.6 Gy/min (86.2–101.7 Gy for five days) | 43.5 °C, 45 min | Murine fibrosarcoma, FSa-II, C3Hf/Sed mice/in vivo | [26] | |
| 10 Gy | 43 °C, 30 min | Murine melanoma, B16F1, C57BL mice/in vivo | Apoptosis | [27] |
| 10 Gy (5 × 2 Gy) | 41.8 °C, 60 min | Human-derived head and neck squamous cell carcinoma, athymic nude mice/in vivo | [28] | |
| 10 Gy | 43 °C, 30 min | Murine sarcoma, Sarcoma 180 (S180), Balb/c mice/in vivo | Necrosis | [29] |
| 30 Gy | 43 °C, 30 min | Murine melanoma, B16F1, C57BL mice/in vivo | [30] | |
| 0.5 Gy/min (total dose not reported) | 41 and 43 °C, 30 min | Human origin cervical carcinoma, SIHA; non-small-cell squamous lung carcinoma, SW-1573; colon cancer, RKO; rodents cell line V79, R1 and RUC/in vitro | [31] | |
| 9 Gy | 43 °C, 60 min | 396 Wistar rats / in vivo | [32] | |
| 36 Gy | 43 °C, 60 min | Human glioblastoma, U-87MG/in vitro | [33] | |
| 2.5 Gy/min (total dose not reported) | 40 °C, 60 min | Lymphoma, EL4, C57BL/6J mouse/in vitro | Apoptosis | [34] |
| 2 Gy | 42 °C, 60 min | Cervical cancer, SiHa and HeLa/in vitro | [35] | |
| 6 Gy | 43 °C, 60 and 90 min | Human prostate cancer stem cells (CSCs), DU145/in vitro | [36] | |
| 5 Gy | 42 °C, 30 min | Rat gliosarcoma, Gs-9L; non-cancerous tissue of canine kidney, MDCK; human-derived breast cancer, MCF-7/in vitro | Apoptosis | [37] |
| 10 Gy (2 × 5 Gy) | 42 °C, 30 min | Lung cancer, A549 and NCI-H1299, BALB/c nude mice/in vitro, in vivo | Apoptosis | [38] |
| 2 and 5 Gy | 47 °C, 0–780 CEM43 | Human colon cancer, HCT116; oral squamous carcinoma, CAL27/in vitro | Apoptosis, Necrosis | [39] |
| 10 Gy | 45 °C, 30 min | Human head and neck cancer, FaDu; human glioblastoma, T98G; human prostate cancer, PC-3/in vitro | [42] | |
| 44 and 46 Gy | 41.5 °C, 60 min | Mammary carcinoma cell line, C3H, CDF1 mice/in vivo | [43] |
| RT | HT | Cell Line and Observation Model | Classification of the Molecular Mechanism | Molecular Mechanism | Ref. |
|---|---|---|---|---|---|
| 20 Gy | 44 °C, 15 min | Murine mammary carcinoma, NU-82, DBA-2 mouse/in vivo | Necrosis/ ATP depletion | Pi ↑, ATP and phosphodiesters ↓ | [53] |
| 102.8 Gy (FSa-II); 40.4 Gy (MCa) | 43.5 °C, 45 min | Spontaneous murine fibrosarcoma, FSa-II; mammary carcinoma, MCa, C3Hf/Sed mice/in vivo | Oxygenation | - | [46] |
| 56.25 Gy (25 × 2.25 Gy) | 44 °C | Spontaneous canine soft tissue sarcomas, 13 dogs/in vivo | Oxygenation | pO2 ↑, tumor perfusion ↑, hypoxic fraction ↓, pHe ↓ | [47] |
| 10 Gy (5 × 2 Gy) | 41.8 °C, 60 min | Human-derived head and neck squamous carcinoma, athymic nude (nu-nu) mice/in vivo | Oxygenation | pO2 ↑ | [48] |
| 10 Gy (5 × 2 Gy) | 43.5 °C, 60 min | Human esophageal carcinoma SGF-3, −4, −5, −7, −8, and −9/in vitro | Cell cycle arrest | Chromosomal aberrations, G2/M phase accumulation | [71] |
| 2 Gy | 42 °C, 20 min | Cancer cell lines carrying a different p53 gene status (wt p53 and m p53)/in vitro | Apoptosis/ DNA damage | p53–dependent apoptosis, Bax and Caspase-3 pathways | [59] |
| 56.25 Gy (25 × 2.25 Gy) | 43 °C, CEM43°CT90 = 10 and 40 min | Canine soft tissue sarcomas/in vitro | Oxygenation | - | [49] |
| 10 Gy | 43 °C, 60 min | Human colon cancer, HT29, nude mice/ in vivo | Apoptosis/ Bax | p53 and Bcl-2 ↓, Bax ↑ | [78] |
| 2.75 Gy/min (total dose not reported) | 40 °C, 9 h | Squamous cell carcinoma, SCC VII, C3H/He mice/in vivo | DNA damage | Change in MN frequency | [60] |
| 1 Gy | 42.5 °C, 60 min | Chinese hamster ovary cells, CHO WT (CHO 10B2); normal human fibroblast cell line, AG1521; DNA repair deficient CHO mutants, V3 (DNA-PKcs), irs1SF (XRCC3), KO40 (FancG), 51D1 (Rad51D), and xrs5 (Ku80); V79 mutants irs1 (XRCC2) and irs3 (Rad51C)/in vitro | DNA damage | Chromosomal aberrations, Rad51 activity at DSBs | [62] |
| 2 Gy | 42.4 °C, 60 min | GSC, patient specimens 3691 and 387, athymic nude mice/in vitro, in vivo | Apoptosis/ AKT | DNA repair ↓, AKT Signaling ↓ | [83] |
| 4 Gy | 41 °C, 60 min | Human lung carcinoma, SW-1573; human colorectal carcinoma, RKO/in vitro | Apoptosis/ DNA damage | Chromosomal aberrations and translocation, BRCA2 degradation, homologous recombination pathway ↓ | [63] |
| 4 Gy | 42 °C, 60 min | Human cervical cancer, HeLa and SiHa; human breast cancer, MCF7, and T47D; primary human breast cancer, BCSC, athymic mice/in vivo (SiHa), in vitro | Apoptosis/ DNA damage | DNA-DSB repair ↓, G2/M phase arrest, Caspase-3 activity ↑ | [64] |
| 2 Gy | 41 °C, 60 min | Human breast adenocarcinoma, MCF-7/in vitro | Apoptosis/ Oxygenation | ROS ↑, Caspase-3 and –9 ↑ | [50] |
| 15 Gy | 41 °C, 30 min | Fibrosarcoma, FSa-II, C3H mice/in vivo | Apoptosis/ Oxygenation | HIF-1a and VEGF ↓ | [51] |
| 4 Gy | 40 °C, 48 h | Liver cancer, HepG2/in vitro | Apoptosis, Necrosis/ Bax | Bax and FasL ↑, VEGF and PDGF ↓ | [79] |
| 8.5, 14, and 21 Gy (LDR); 4, 8.5, 14.5, 20, and 24 Gy (HDR) | 40 °C, 2 h | Human head and neck squamous cell carcinoma, SAS, nude mice/in vivo | Apoptosis/ DNA damage | Tumor sensitivity ↓, p53-dependent recovery | [61] |
| 4 Gy | 42 °C, 5 min | Human hypopharyngeal carcinoma, FaDu/in vitro | Oxygenation | histone γH2AX phosphorylation ↑ | [52] |
| 3 Gy | 44 °C, 60 min | Human non-small-cell lung cancer, A549 and NCI-H292/in vitro, in vivo | Apoptosis/ DNA damage, HSP70 | KLF11 and NR4A3↑, intracellular ROS ↑ | [65] |
| 2 Gy | 42 °C, 30 min | DNA extracts/in vitro | DNA damage | - | [66] |
| - | - | Human chordoma, U-CH2 and Mug-chor1/in vitro | Apoptosis/ gene expression, HSP70 | Brachyury ↓, death receptor activation | [80] |
| 2 Gy | 42 °C, 60 min | Colon cancer, IDH1MUT and IDH1WT HCT116; chondrosarcoma, Hyperthermia1080 cells/in vitro | DNA damage | NADPH↓, homologous recombination repair ↓ | [67] |
| RT | HT | Cell Line and Observation Model | Classification of the Molecular Mechanism | Molecular Mechanism | Ref. |
|---|---|---|---|---|---|
| 5 Gy | 41.5 °C, 60 min | Human colorectal adenocarcinoma, HCT15/in vitro | Necrosis/ Immune response | HMGB1 release, G2/M phase arrest | [88] |
| 5 or 10 Gy | 41.5 °C, 60 min | Human colorectal adenocarcinoma, SW480, and HCT 15/in vitro | Necrosis/ Immune response | HMGB1 release, G2/M phase arrest | [89] |
| 2, 5, and 10 Gy | 41.5 °C, 60 min | Human colorectal tumor, HCT15, and SW480; mouse colon carcinoma tumor, CT26.WT (CRL-2638)/ in vitro, in vivo | Immune response, HSP70 | CD80 and CCR7 ↑, phagocytosis of macrophages and DCs ↑, IL-8, and IL-12 ↑ | [90] |
| 20 Gy (2 × 10 Gy) | 45 °C, 3 min | Human murine breast cancer, 4T1, BALB/C mice/in vivo | Apoptosis/ Immune response | CD4+ T cell and CD4+/CD8+ cell ratio ↑, TNF-α, IFN-γ, and IL-2 ↑, Bax ↑, MMP-9 ↓ | [91] |
| 2 Gy | 41.5 °C, 60 min | Mouse melanoma, B16-F10, C57/BL6 mice/ in vitro, in vivo | Apoptosis, Necrosis/ Immune response, HSP70 | HMGB1 ↑, infiltration of CD8 + T cells, DCs, and NK cells ↑ | [92] |
| 20 Gy | 42 °C, 30 min | Erythroleukemia, K-562/in vitro | Immune response | NK cytotoxicity ↓ | [97] |
| 15 Gy | 41.5 °C, 60 min | Mouse melanoma, B16-F10, C57BL/6 mice/ in vivo, in vitro | Apoptosis, Necrosis/ Immune response | HMGB1 release, NK cell, B cell, and T cell count ↑ | [99] |
| 20 Gy | 42 °C, 0–180 min | NK cell; Erythroleukemia, K-562/in vitro | Immune response | ATP level of NK cell ↓ | [98] |
| 8 Gy | 42.5 °C, 30 min | Murine syngeneic Panc02, Panc02, C57BL/6 mice/in vivo | Immune response | CD8a+ and CD4+ T cells ↑ | [93] |
| 10 Gy (5 × 2 and 2 × 5 Gy) | 39, 41, 44 °C, 60 min | Human breast cancer, MCF-7, and MDA-MB-231/in vitro | Apoptosis, Necrosis/ ICM, HSP70 | PD-L1, PD-L2, HVEM ↑, CD137-L, OX40-L, CD27-L, ICOS-L ↑, EGFR ↑ | [103] |
| 10 Gy (5 × 2 and 2 × 5 Gy) | 39, 41, 44 °C, 60 min | Murine melanoma, B16; human breast cancer, MCF-7, and MDA-MB-231/in vitro | Apoptosis, Necrosis/ ICM | PD-L1, PD-L2, HVEM, and Gal-9 ↑ | [104] |
| 10 Gy (5 × 2 Gy) | 41 and 44 °C, 1 h | Human glioblastoma, U87 and U251/in vitro | Apoptosis, Necrosis/ ICM, HSP70 | PD-L1, PD-L2, HVEM ↑, ICOS-L, CD137-L, and Ox40-L ICMs ↑ | [101] |
| 10 Gy (2 × 5 Gy) | 39, 41, 44 °C, 60 min | Human MCF-7 and MDA-MB-231 breast cancer cells/in vitro | Apoptosis, Necrosis/ICMs | PD-L1, PD-L2, and HVEM ↑ | [105] |
| 15 Gy | 41.0 °C, 30 min | Fibrosarcoma, FSa-II, C3H mice/in vitro | Apoptosis/ ICM, Hypoxia | PD-L1 ↓, VEGF ↓, HIF-1α ↓ | [106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, S.; Jung, S.; Baek, S.H. Combination Therapy of Radiation and Hyperthermia, Focusing on the Synergistic Anti-Cancer Effects and Research Trends. Antioxidants 2023, 12, 924. https://doi.org/10.3390/antiox12040924
Kwon S, Jung S, Baek SH. Combination Therapy of Radiation and Hyperthermia, Focusing on the Synergistic Anti-Cancer Effects and Research Trends. Antioxidants. 2023; 12(4):924. https://doi.org/10.3390/antiox12040924
Chicago/Turabian StyleKwon, Seeun, Sumin Jung, and Seung Ho Baek. 2023. "Combination Therapy of Radiation and Hyperthermia, Focusing on the Synergistic Anti-Cancer Effects and Research Trends" Antioxidants 12, no. 4: 924. https://doi.org/10.3390/antiox12040924