
Rehmannia glutinosa Polysaccharides: Optimization of the Decolorization Process and Antioxidant and Anti-Inflammatory Effects in LPS-Stimulated Porcine Intestinal Epithelial Cells
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of RGP
2.2. Calculation of Decolorization Index
2.2.1. Decolorization Rate (DR)
2.2.2. Retention Rate (RR)
2.2.3. Composite Score (CS)
2.3. Optimization of RGP Decolorization Using AB-8 Macroporous Resin
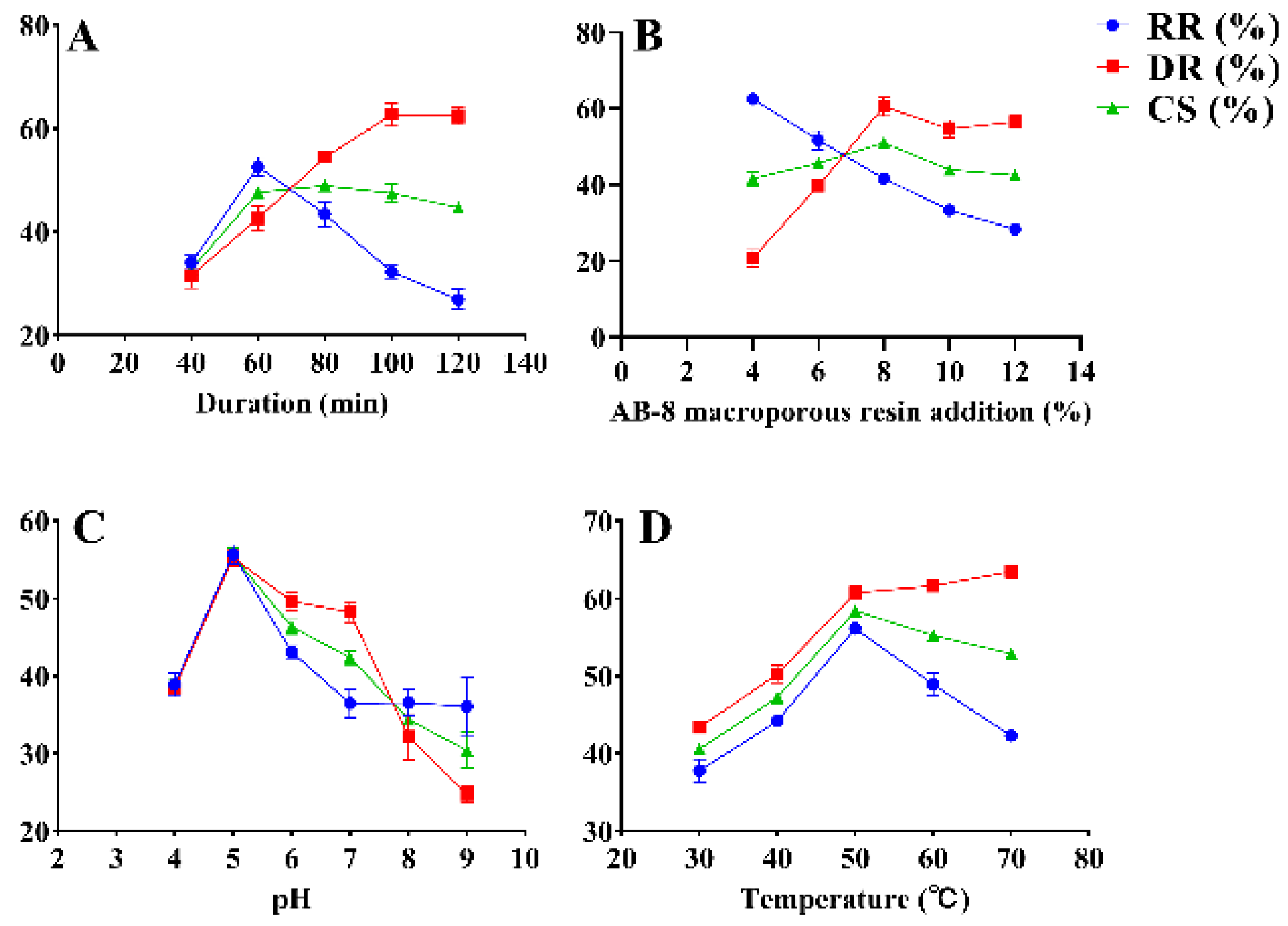
2.3.1. Single-Factor Experiment (SFE) for RGP Decolorization Using AB-8 Macroporous Resin
2.3.2. RSM for RGP Decolorization Using AB-8 Macroporous Resins
2.4. Optimization of RGP Decolorization Using H2O2
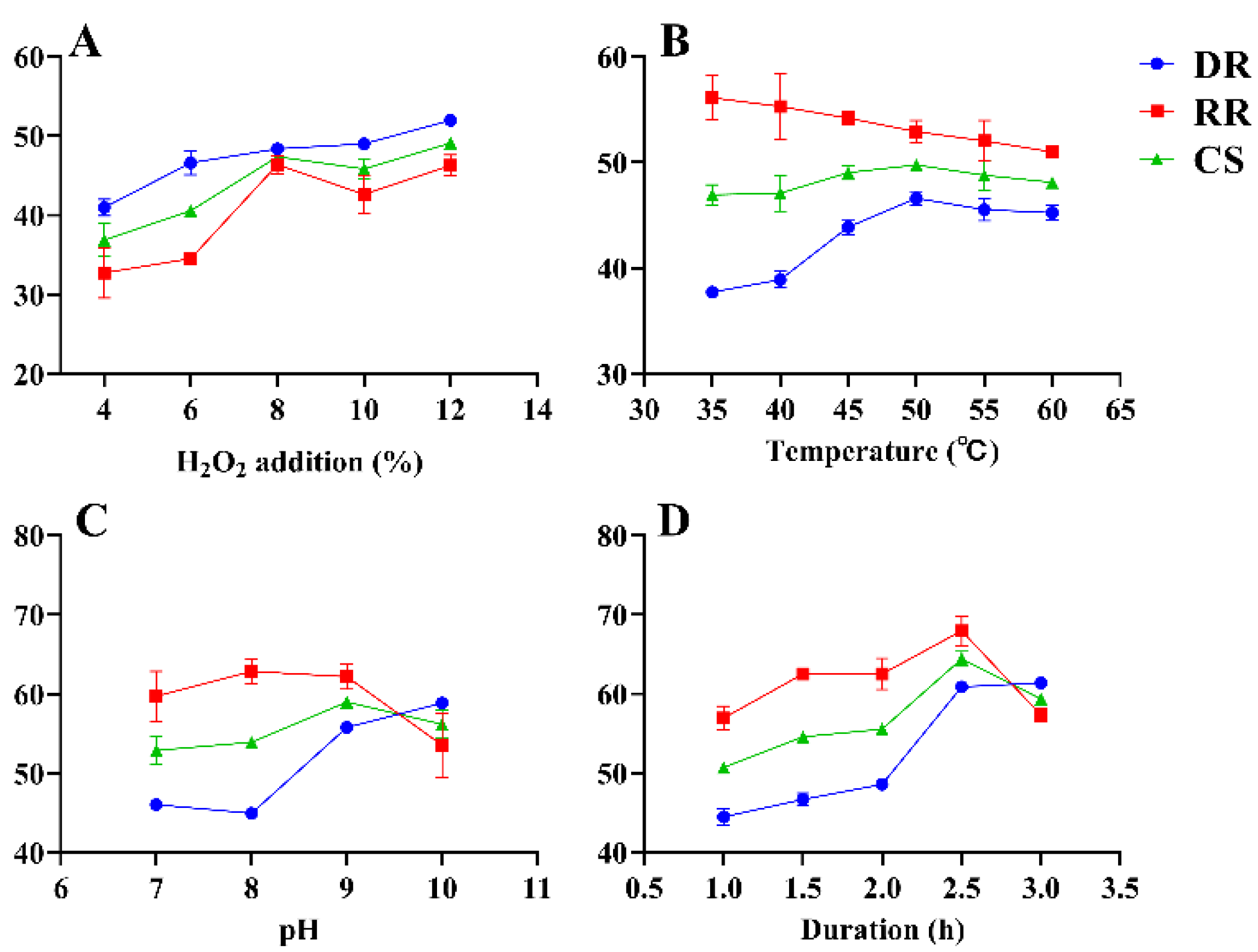
2.4.1. SFE for RGP Decolorization Using H2O2
2.4.2. RSM for RGP Decolorization Using H2O2
2.5. Purification of RGP
2.6. Protective Effect of RGP on IPEC-J2 Cells
2.6.1. Cell Culture
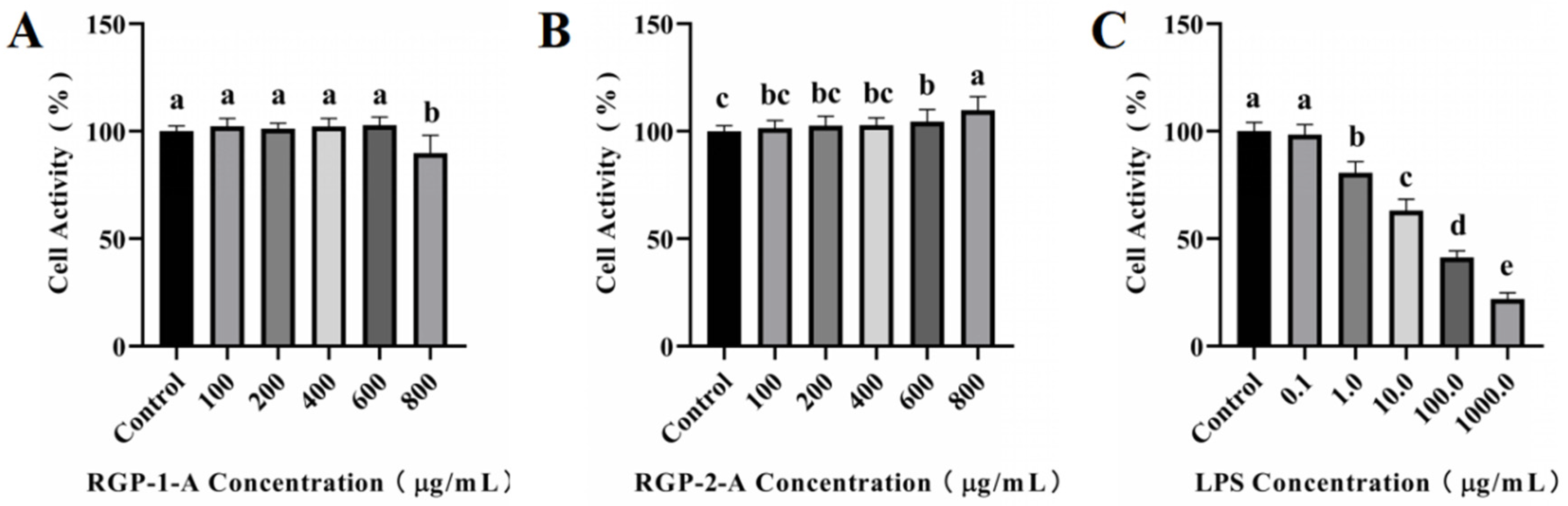
2.6.2. Viability Assay
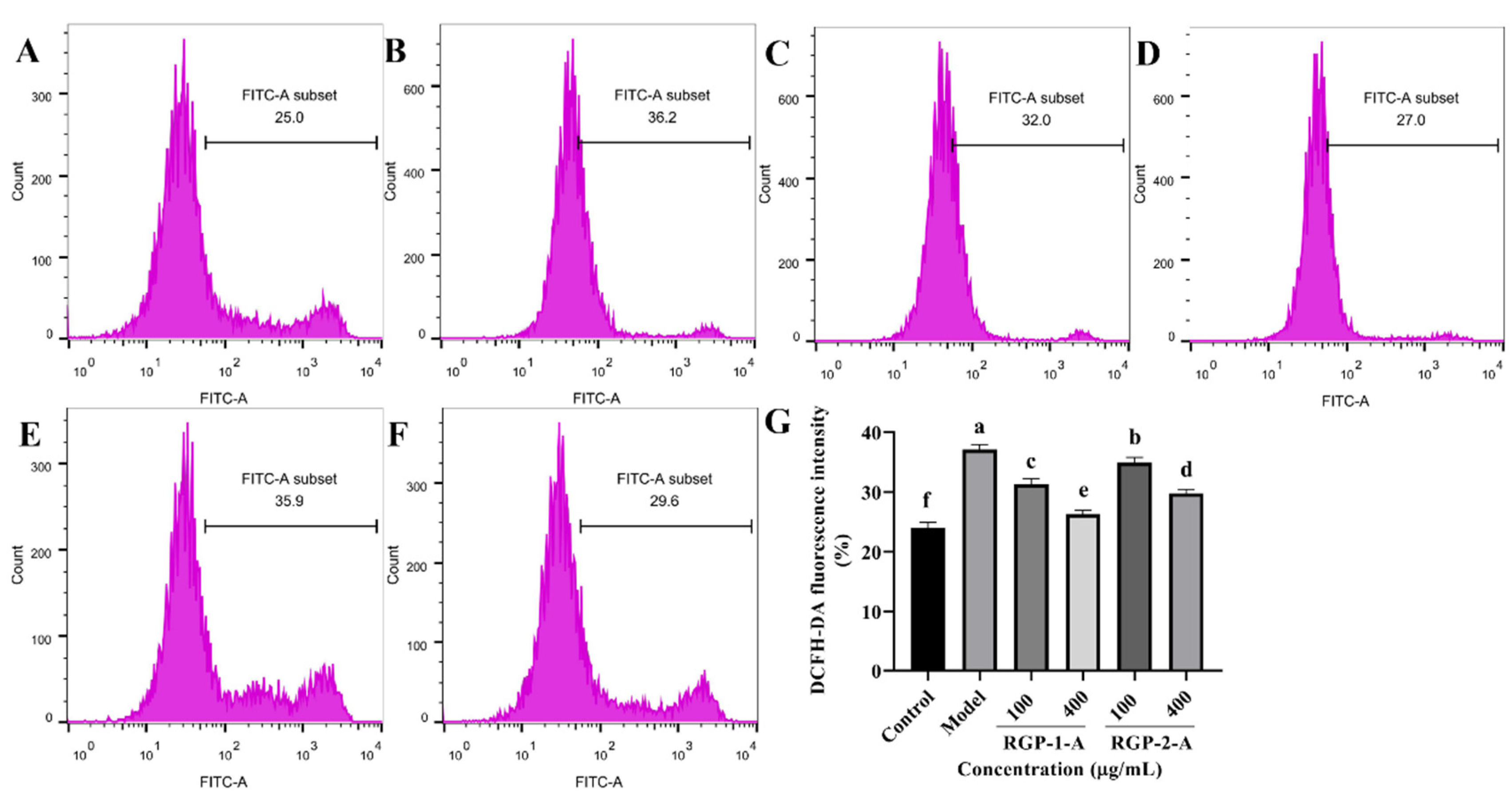
2.6.3. Determination of ROS Levels
2.6.4. Determination of Mitochondrial Membrane Potential (MMP)
2.6.5. Apoptosis Assay
2.6.6. Evaluation of Oxidation-Related Indexes
2.6.7. RT-PCR Analysis
2.7. Statistical Analysis
3. Results
3.1. Optimization of RGP Decolorization Using AB-8 Macroporous Resin
3.1.1. Model Fitting
3.1.2. Optimization of Decolorization Conditions
3.2. Optimization of RGP Decolorization Using H2O2
3.2.1. Model Fitting
3.2.2. Optimization of Decolorization Conditions
3.3. Protection of IPEC-J2 Cells from LPS-Induced Inflammatory Damage
3.3.1. Cytotoxicity Assay
3.3.2. RGP Treatment Reduces the Levels of ROS in LPS-Induced IPEC-J2 Cells
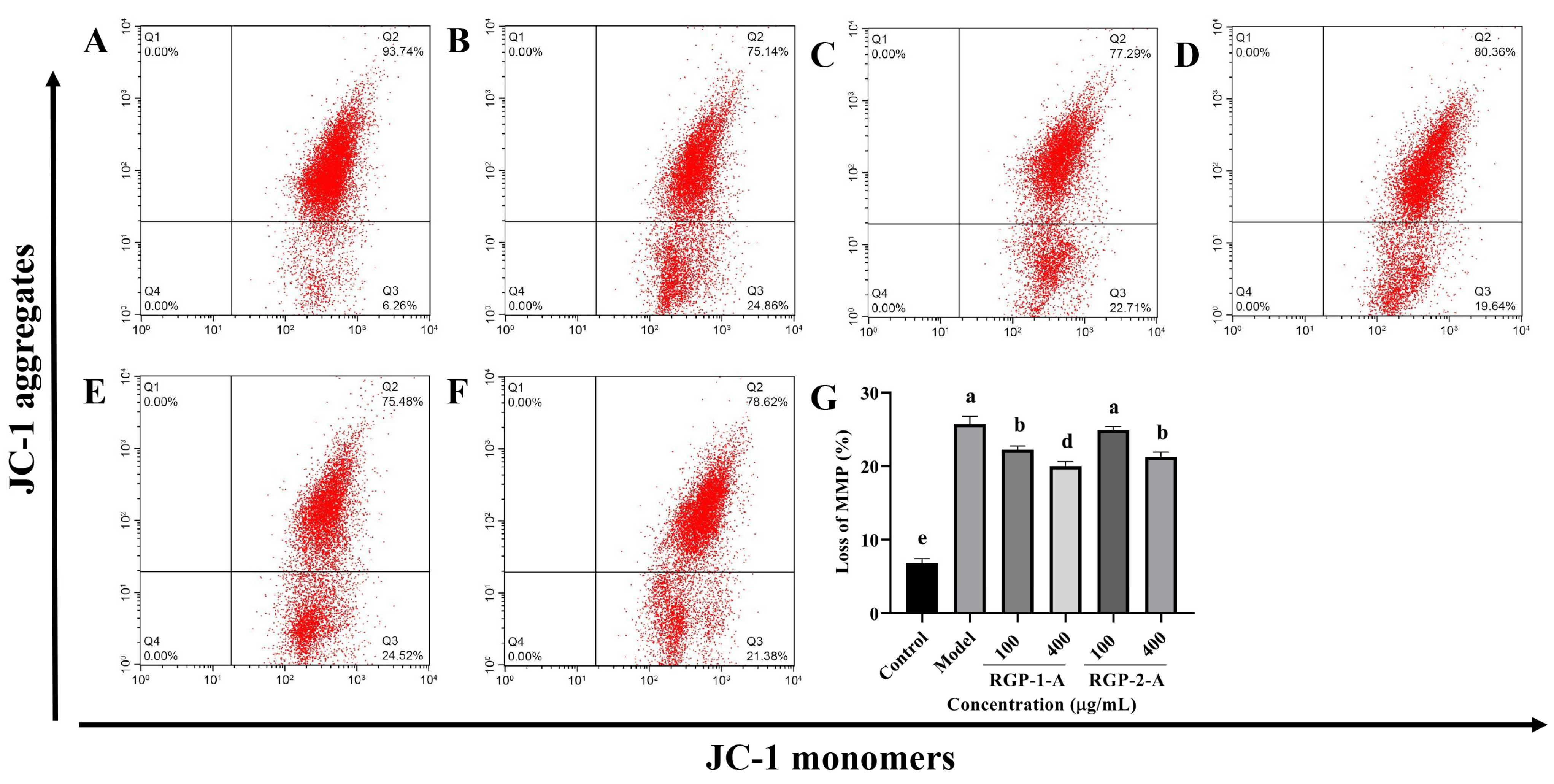
3.3.3. RGP Treatment Attenuates the LPS-Induced Reduction in MMP
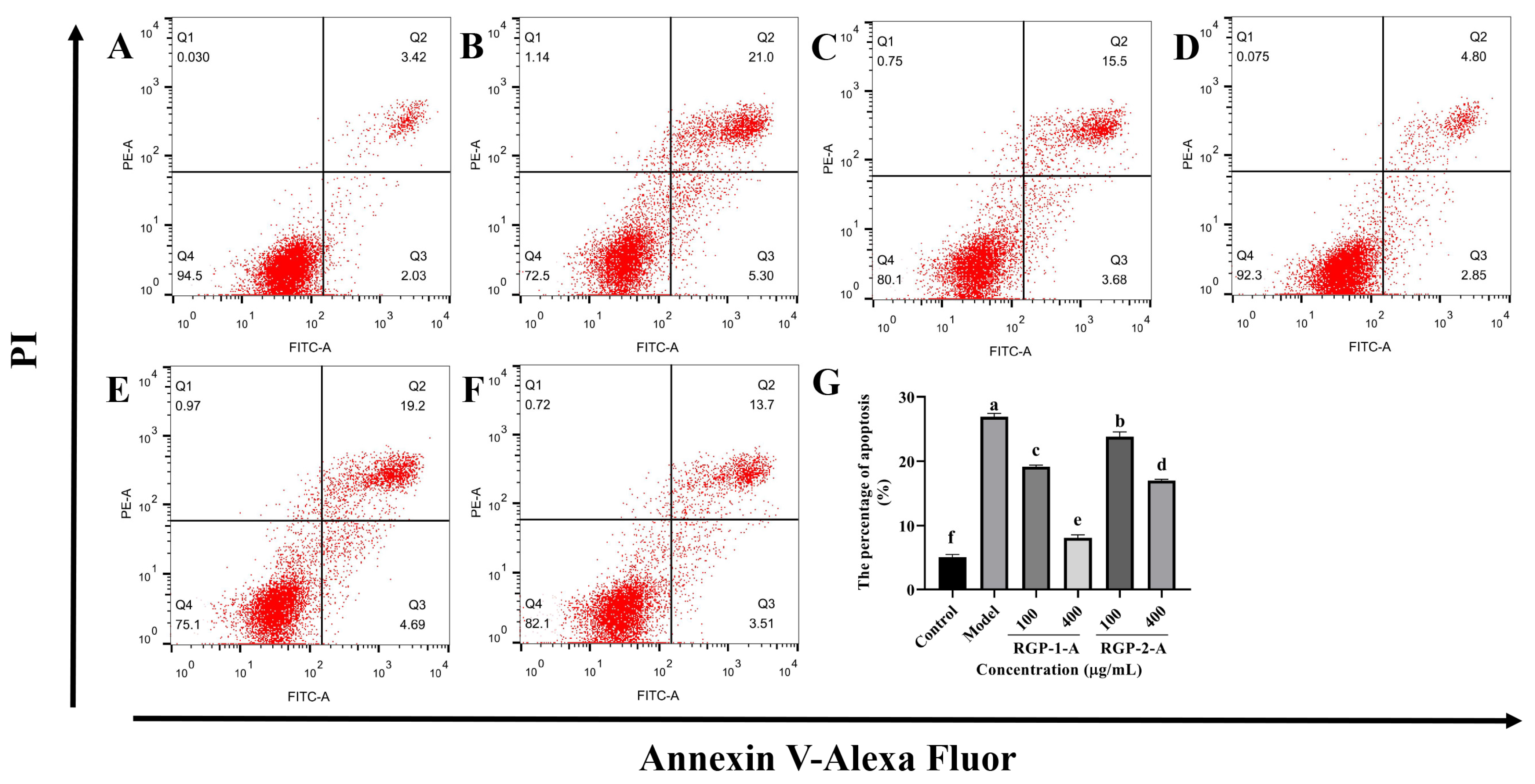
3.3.4. RGP Treatment Ameliorates LPS-Induced Apoptosis
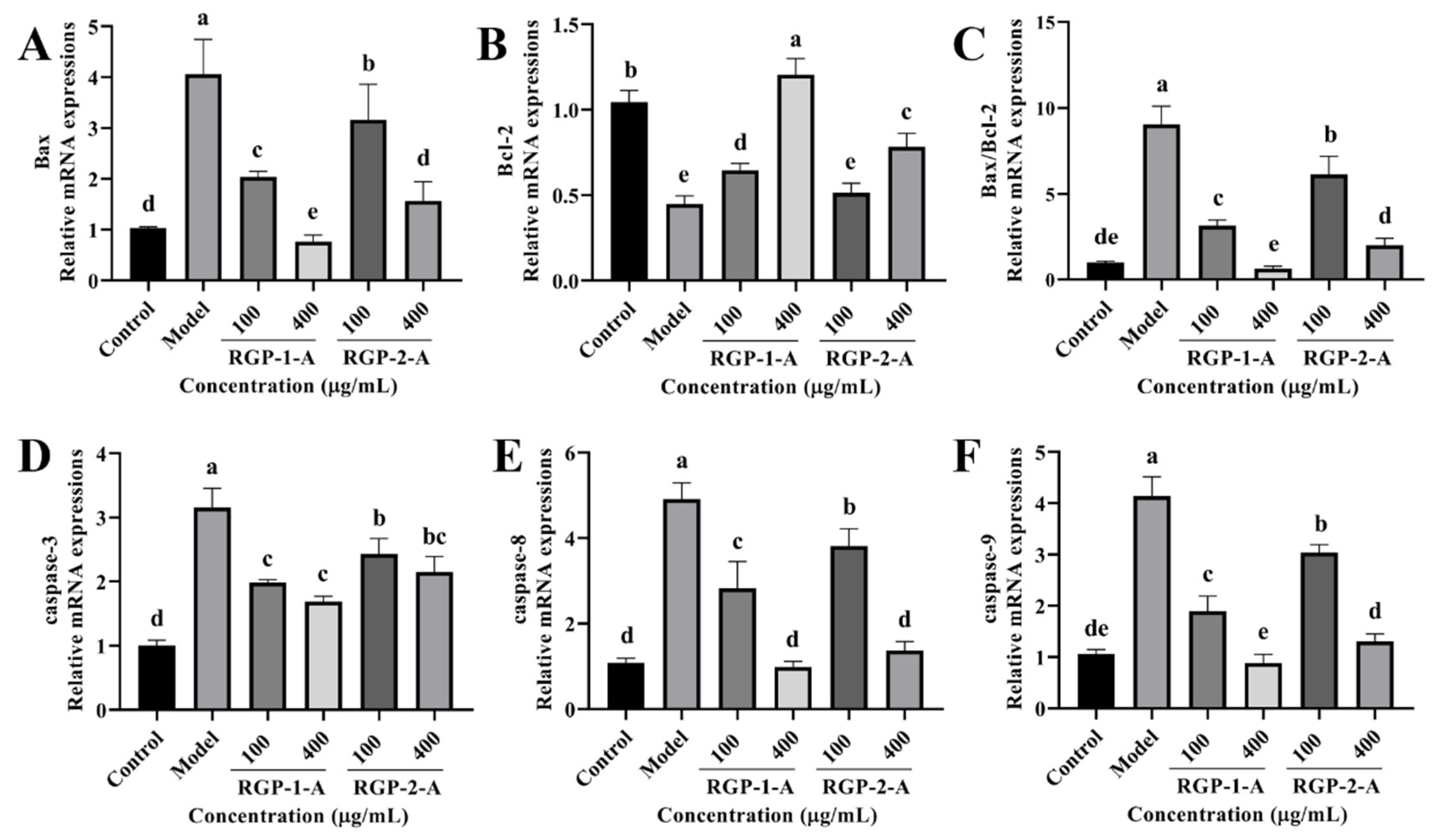
3.3.5. RGP Treatment Ameliorates the Expression of Apoptosis-Related Genes Induced by LPS
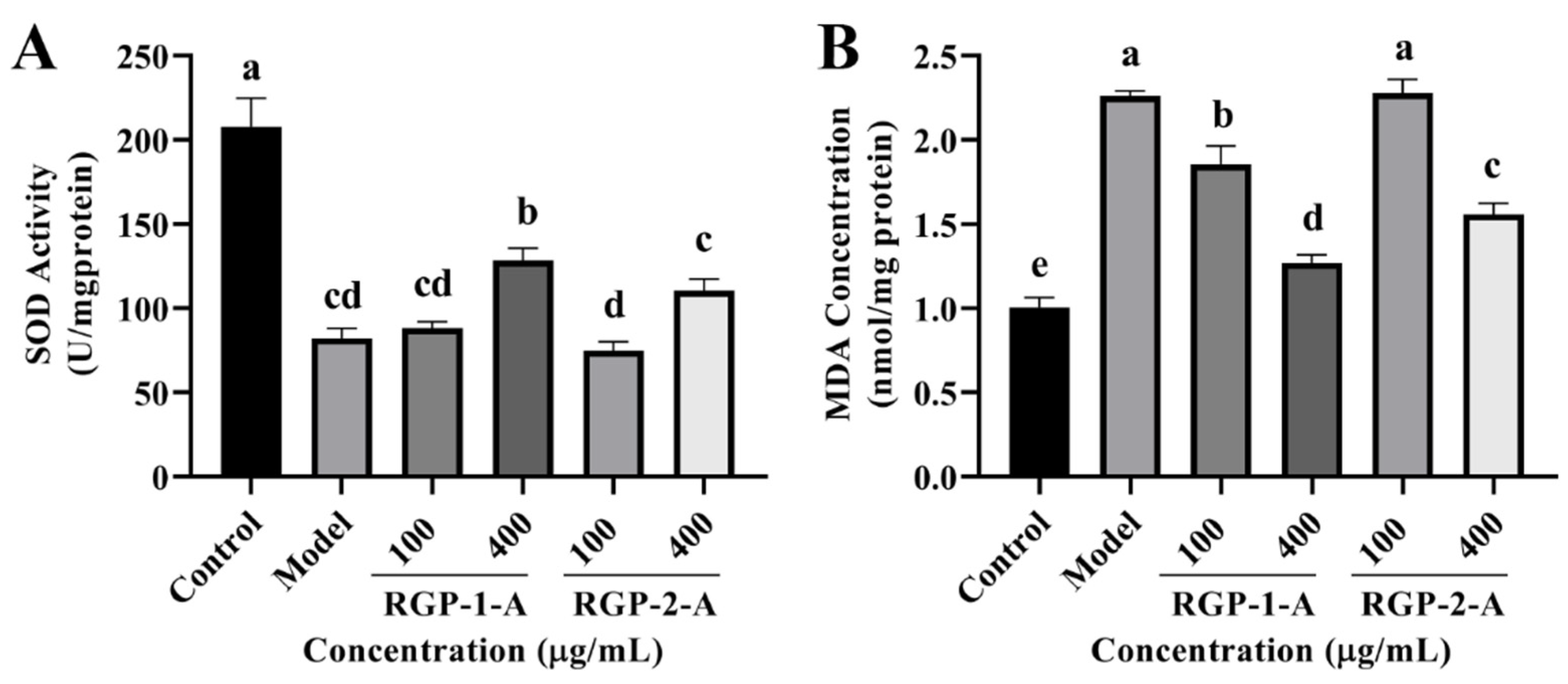
3.3.6. RGP Treatment Increases Intracellular Antioxidant Enzyme Activity
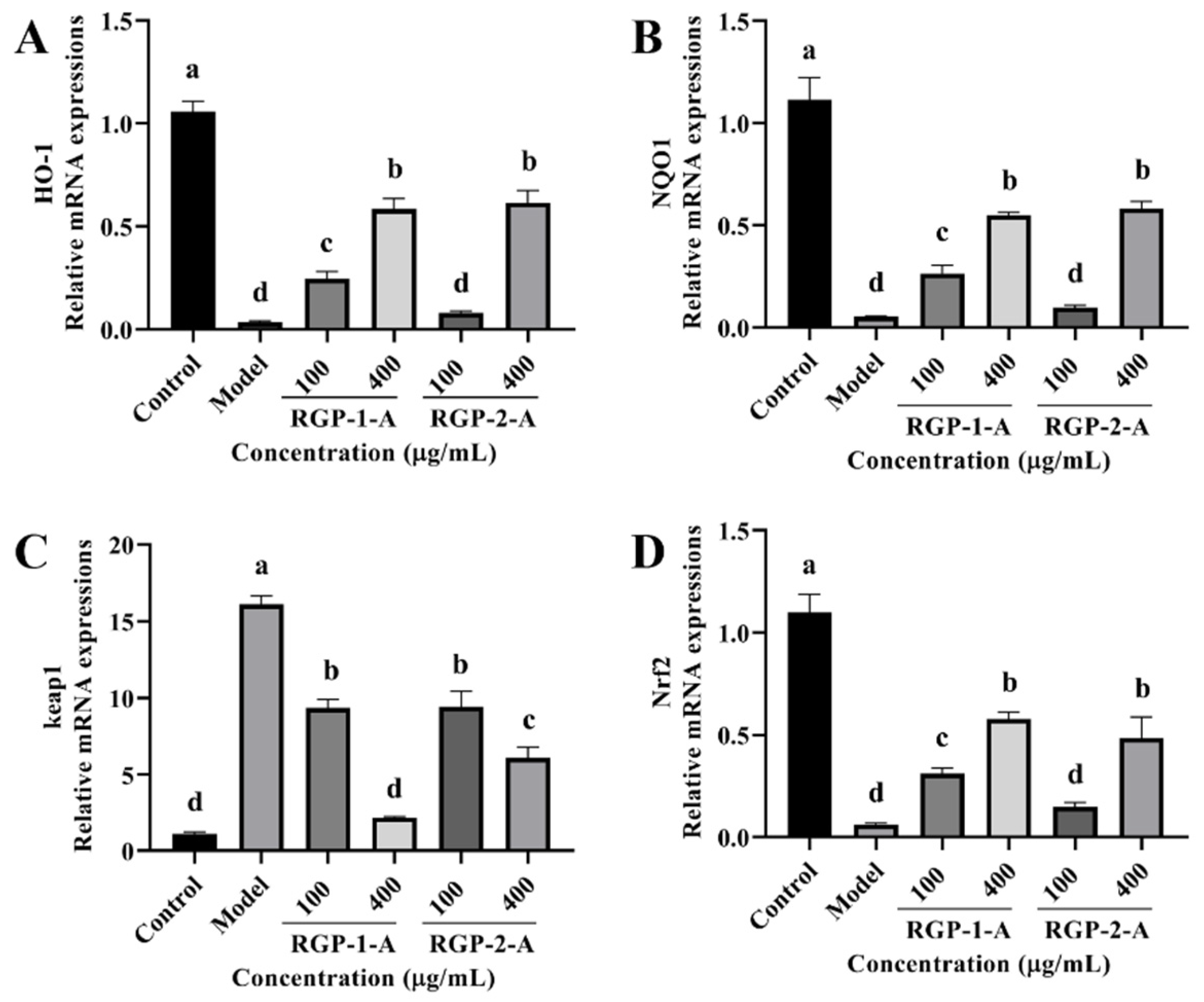
3.3.7. RGP Treatment Promotes Antioxidant Signaling Pathways
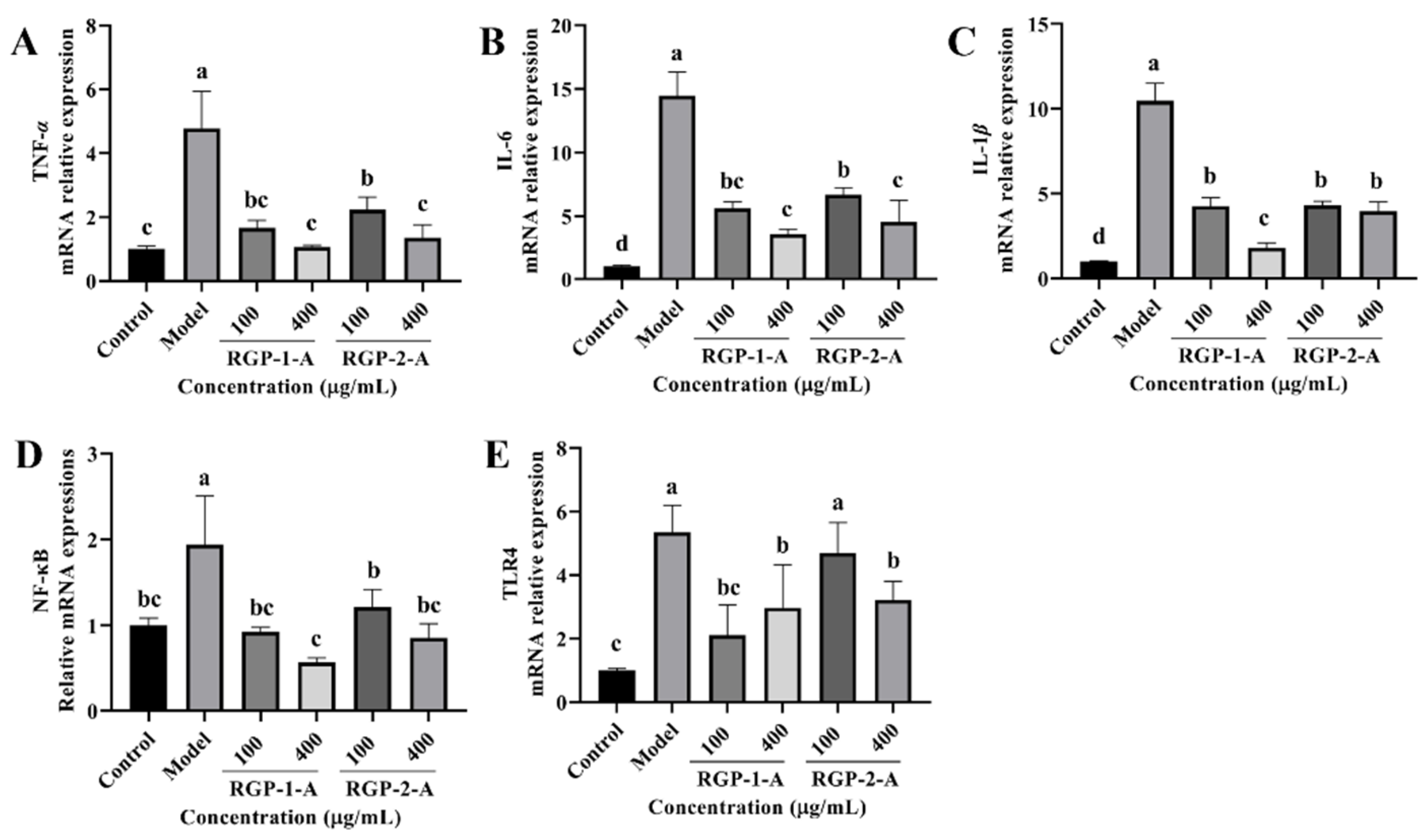
3.3.8. RGP Treatment Inhibits Inflammatory Cytokines and Signaling Pathways
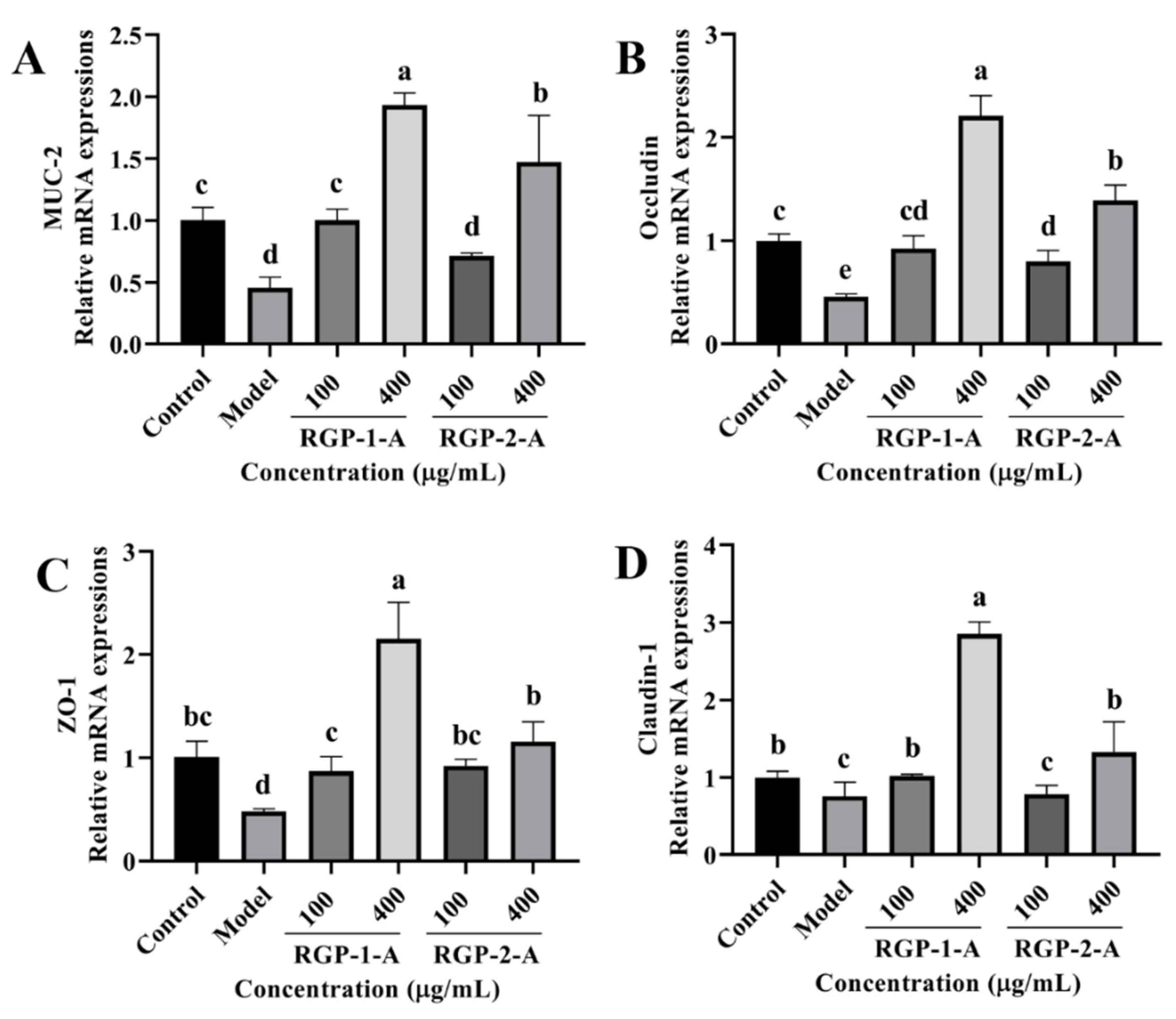
3.3.9. RGP Treatment Attenuates LPS-Induced Barrier Dysfunction in IPEC-J2 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef]
- Kim, S.W.; Holanda, D.M.; Gao, X.; Park, I.; Yiannikouris, A. Efficacy of a yeast cell wall extract to mitigate the effect of naturally co-occurring mycotoxins contaminating feed ingredients fed to young pigs: Impact on gut health, microbiome, and growth. Toxins 2019, 11, 633. [Google Scholar] [CrossRef] [Green Version]
- Uyehara, C.F.T.; Ho, D.H.; Murata, L.M.; Piaggione, J.M.; Ichimura, W.M.; Hernandez, C.A.; Blythe, J.W.; Sherman, W.J. Vasopressin’s role in stress response and glucose homeostasis in a pig model of endotoxin-induced inflammation. FASEB J. 2018, 32, 901–905. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of two mycotoxins deoxynivalenol and fumonisin on pig intestinal health. Porc. Health Manag. 2016, 2, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluske, J.; Miller, D.; Sterndale, S.; Turpin, D. Associations between gastrointestinal-tract function and the stress response after weaning in pigs. Anim. Prod. Sci. 2019, 59, 2015–2022. [Google Scholar] [CrossRef]
- Xiong, L.; Zhang, W.; Zhao, H.; Tian, Z.; Ren, M.; Chen, F.; Guan, W.; Zhang, S. Dietary supplementation of enzymatically treated artemisia annua l. Improves lactation performance, alleviates inflammatory response of sows reared under heat stress, and promotes gut development in preweaning offspring. Front. Vet. Sci. 2022, 9, 843673. [Google Scholar] [CrossRef]
- Taranu, I.; Pistol, G.C.; Anghel, A.C.; Marin, D.; Bulgaru, C. Yeast-fermented rapeseed meal extract is able to reduce inflammation and oxidative stress caused by Escherichia coli lipopolysaccharides and to replace zno in Caco-2/Htx29 co-culture cells. Int. J. Mol. Sci. 2022, 23, 11640. [Google Scholar] [CrossRef]
- Adedokun, S.A.; Olojede, O.C. Optimizing gastrointestinal integrity in poultry: The role of nutrients and feed additives. Front. Vet. Sci. 2018, 5, 348. [Google Scholar] [CrossRef] [Green Version]
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, M.; Jia, Z. Rehmannia glutinosa: Review of botany, chemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, C.; Xu, N.; Li, J.; Meng, F.; Zhai, H. Sorting and identification of Rehmannia glutinosa germplasm resources based on EST-SSR, scanning electron microscopy micromorphology, and quantitative taxonomy. Ind. Crops Prod. 2018, 123, 303–314. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, G.; Huang, H. Extraction, derivatization and antioxidant activities of onion polysaccharide. Food Chem. 2022, 388, 133000. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Chen, J.; Yuan, Y.; Zhang, X.; Li, L.; Zhai, Y.; Gong, X.; Li, B. Structural characterization, antioxidant, and anti-inflammatory activity of polysaccharides from plumula nelumbinis. Int. J. Biol. Macromol. 2022, 212, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, W.; Zeng, L.; Jin, J. Rehmannia glutinosa polysaccharide induced an anti-cancer effect by activating natural killer cells. Int. J. Biol. Macromol. 2017, 105, 680–685. [Google Scholar] [CrossRef]
- Lu, M.; Chang, C.; Chao, C.; Hsu, Y. Structural changes, and anti-inflammatory, anti-cancer potential of polysaccharides from multiple processing of rehmannia glutinosa. Int. J. Biol. Macromol. 2022, 206, 621–632. [Google Scholar] [CrossRef]
- Wang, T.; Wang, X.; Zhuo, Y.; Si, C.; Yang, L.; Meng, L.; Zhu, B. Antiviral activity of a polysaccharide from Radix Isatidis (Isatis indigotica Fortune) against hepatitis B virus (HBV) in vitro via activation of JAK/STAT signal pathway. J. Ethnopharmacol. 2020, 257, 112782. [Google Scholar] [CrossRef]
- Huang, Y.; Nan, L.; Xiao, C.; Su, F.; Li, K.; Ji, Q.; Wei, Q.; Liu, Y.; Bao, G. Pegylated nano-rehmannia glutinosa polysaccharide induces potent adaptive immunity against bordetella bronchiseptica. Int. J. Biol. Macromol. 2021, 168, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, G.; Yan, J.; Li, K.; Bai, Z.; Cheng, W.; Huang, K. Rehmannia glutinosa (gaertn.) Dc. Polysaccharide ameliorates hyperglycemia, hyperlipemia and vascular inflammation in streptozotocin-induced diabetic mice. J. Ethnopharmacol. 2015, 164, 229–238. [Google Scholar] [CrossRef]
- Tomoda, M.; Miyamoto, H.; Shimizu, N.; Gonda, R.; Ohara, N. Characterization of two polysaccharides having activity on the reticuloendothelial system from the root of rehmannia glutinosa. Chem. Pharm. Bull. 1994, 42, 625–629. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Li, Z.; Gao, R.; Zhao, T.; Luo, D.; Yu, Z.; Zhang, S.; Qi, C.; Wang, Y.; Qiao, H.; et al. Structural characteristics of rehmannia glutinosa polysaccharides treated using different decolorization processes and their antioxidant effects in intestinal epithelial cells. Foods 2022, 11, 3449. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, S.; Feng, W.; Zhang, Z.; Li, H. Structural characterization and immunomodulatory activities of two polysaccharides from rehmanniae radix praeparata. Int. J. Biol. Macromol. 2021, 186, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Shen, M.; Nie, S.; Li, C.; Xie, M. Decolorization of polysaccharides solution from cyclocarya paliurus (batal.) Iljinskaja using ultrasound/H2O2 process. Carbohyd. Polym. 2011, 84, 255–261. [Google Scholar] [CrossRef]
- Hu, Z.; Zhou, H.; Li, Y.; Wu, M.; Yu, M.; Sun, X. Optimized purification process of polysaccharides from Carex meyeriana kunth by macroporous resin, its characterization and immunomodulatory activity. Int. J. Biol. Macromol. 2019, 132, 76–86. [Google Scholar] [CrossRef]
- Shao, L.; Sun, Y.; Liang, J.; Li, M.; Li, X. Decolorization affects the structural characteristics and antioxidant activity of polysaccharides from Thesium Chinense turcz: Comparison of activated carbon and hydrogen peroxide decolorization. Int. J. Biol. Macromol. 2020, 155, 1084–1091. [Google Scholar] [CrossRef]
- Sun, L. Protective effects of rehmannia glutinosa polysaccharides on cadiocytes of neonatal rat impaired by hypoxia-reoxygenation. J. Med. Res. 2016, 45, 146–150. [Google Scholar]
- Peng, H. Effects of rehmannia glutinosa polysaccha-rides on myocardial apoptosis of rats after; Myocardial ischemia-reperfusion injury. Int. J. Tradit. Chin. Med. 2016, 10, 918–921. [Google Scholar]
- Meng, J. Protective effects and mechanism of rehmannia polysaccharide on cadiocytes of neonatal rat impaired by H2O2. Pharm. Clin. Chin. Mater. Med. 2016, 32, 90–95. [Google Scholar]
- Rudtanatip, T.; Pariwatthanakun, C.; Somintara, S.; Sakaew, W.; Wongprasert, K. Structural characterization, antioxidant activity, and protective effect against hydrogen peroxide-induced oxidative stress of chemically degraded gracilaria fisheri sulfated galactans. Int. J. Biol. Macromol. 2022, 206, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.; Lee, Y.C. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, M.; Sun, K.; Su, S.; Geng, T.; Sun, H. Hippophae rhamnoides polysaccharides protect IPEC-J2 cells from lps-induced inflammation, apoptosis and barrier dysfunction in vitro via inhibiting TLR4/NF-κB signaling pathway. Int. J. Biol. Macromol. 2020, 155, 1202–1215. [Google Scholar] [CrossRef]
- Serarols, J.; Poch, J.; Villaescusa, I. Determination of the effective diffusion coefficient of Zn (ii) on a macroporous resin XAD-2 impregnated with di-2-ethylhexyl phosphoric acid (DEHPA): Influence of metal concentration and particle size. React. Funct. Polym. 2001, 48, 53–63. [Google Scholar] [CrossRef]
- Scordino, M.; Di Mauro, A.; Passerini, A.; Maccarone, E. Adsorption of flavonoids on resins: Hesperidin. J. Agric. Food Chem. 2003, 51, 6998–7004. [Google Scholar] [CrossRef]
- Vončina, D.B.; Majcen-Le-Marechal, A. Reactive dye decolorization using combined ultrasound/H2O2. Dye. Pigment. 2003, 59, 173–179. [Google Scholar] [CrossRef]
- Halpern, M.D.; Denning, P.W. The role of intestinal epithelial barrier function in the development of NEC. Tissue Barriers 2015, 3, e1000707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, J.; Wu, M.; Xiao, H.; Ren, W.; Duan, J.; Yang, G.; Li, T.; Yin, Y. Development of an antioxidant system after early weaning in piglets. J. Anim. Sci. 2014, 92, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, M.; Wang, S.; Chu, X.; Ji, H. Identification of phytogenic compounds with antioxidant action that protect porcine intestinal epithelial cells from hydrogen peroxide induced oxidative damage. Antioxidants 2022, 11, 2134. [Google Scholar] [CrossRef]
- Gultekin, F.; Delibas, N.; Yasar, S.; Kilinc, I. In vivo changes in antioxidant systems and protective role of melatonin and a combination of vitamin C and vitamin E on oxidative damage in erythrocytes induced by chlorpyrifos-ethyl in rats. Arch. Toxicol. 2001, 75, 88–96. [Google Scholar] [CrossRef]
- Vasantharaja, R.; Stanley Abraham, L.; Gopinath, V.; Hariharan, D.; Smita, K.M. Attenuation of oxidative stress induced mitochondrial dysfunction and cytotoxicity in fibroblast cells by sulfated polysaccharide from padina gymnospora. Int. J. Biol. Macromol. 2019, 124, 50–59. [Google Scholar] [CrossRef]
- Chen, J.; Huang, Z.; Cao, X.; Chen, X.; Zou, T.; You, J. Plant-derived polyphenols as Nrf2 activators to counteract oxidative stress and intestinal toxicity induced by deoxynivalenol in swine: An emerging research direction. Antioxidants 2022, 11, 2379. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta. Mol. Cell Res. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A. The keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Qu, J.; Huang, P.; Zhang, L.; Qiu, Y.; Qi, H.; Leng, A.; Shang, D. Hepatoprotective effect of plant polysaccharides from natural resources: A review of the mechanisms and structure-activity relationship. Int. J. Biol. Macromol. 2020, 161, 24–34. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Wang, L.; Sanjeewa, K.K.A.; Kang, S.I.; Lee, J.; Jeon, Y. Antioxidant potential of sulfated polysaccharides from Padina boryana; Protective effect against oxidative stress in in vitro and in vivo zebrafish model. Mar. Drugs 2020, 18, 212. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Zhao, M.; Ouyang, K.; Chen, S.; Zhang, Y.; Liu, X.; An, Q.; Zhao, Z.; Wang, W. Sulfated modification, structures, antioxidant activities and mechanism of cyclocarya paliurus polysaccharides protecting dendritic cells against oxidant stress. Ind. Crops Prod. 2021, 164, 113353. [Google Scholar] [CrossRef]
- Zhang, C.; Li, C.; Zhao, P.; Shao, Q.; Ma, Y.; Bai, D.; Liao, C.; He, L.; Huang, S.; Wang, X. Effects of dietary glycyrrhiza polysaccharide supplementation on growth performance, intestinal antioxidants, immunity and microbiota in weaned piglets. Anim. Biotechnol. 2022, 1–12. [Google Scholar] [CrossRef]
- Xie, C.; Zhang, Y.; Niu, K.; Liang, X.; Wang, H.; Shan, J.; Wu, X. Enteromorpha polysaccharide-zinc replacing prophylactic antibiotics contributes to improving gut health of weaned piglets. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2021, 7, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Kantari, C.; Walczak, H. Caspase-8 and bid: Caught in the act between death receptors and mitochondria. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2011, 1813, 558–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amelio, M.; Cavallucci, V.; Cecconi, F. Neuronal caspase-3 signaling: Not only cell death. Cell Death Differ. 2010, 17, 1104–1114. [Google Scholar] [CrossRef]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Johnson, T.S.; Thomas, G.L.; Watson, P.F.; Wagner, B.; Furness, P.N.; El Nahas, A.M. A shift in the bax/bcl-2 balance may activate caspase-3 and modulate apoptosis in experimental glomerulonephritis. Kidney Int. 2002, 62, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Aviello, G.; Knaus, U.G. Ros in gastrointestinal inflammation: Rescue or sabotage? Brit. J. Pharmacol. 2017, 174, 1704–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, A.; Turner, J.R. Cell biology of tight junction barrier regulation and mucosal disease. Csh. Perspect. Biol. 2018, 10, a029314. [Google Scholar] [CrossRef] [PubMed]
- Allam-Ndoul, B.; Castonguay-Paradis, S.; Veilleux, A. Gut microbiota and intestinal trans-epithelial permeability. Int. J. Mol. Sci. 2020, 21, 6402. [Google Scholar] [CrossRef] [PubMed]
- Mizrahy, S.; Peer, D. Polysaccharides as building blocks for nanotherapeutics. Chem. Soc. Rev. 2012, 41, 2623–2640. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Huang, M.; Shen, M.; Wen, P.; Wu, T.; Hong, Y.; Yu, Q.; Chen, Y.; Xie, J. Sulfated modification enhanced the antioxidant activity of mesona chinensis benth polysaccharide and its protective effect on cellular oxidative stress. Int. J. Biol. Macromol. 2019, 136, 1000–1006. [Google Scholar] [CrossRef]
- Wang, X.; Wang, S.; Li, Y.; Wang, F.; Yang, X.; Yao, J. Sulfated astragalus polysaccharide can regulate the inflammatory reaction induced by lps in Caco2 cells. Int. J. Biol. Macromol. 2013, 60, 248–252. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, H.; Han, Q.; Lan, J.; Chen, G.; Cao, G.; Yang, C. Effects of astragalus and ginseng polysaccharides on growth performance, immune function and intestinal barrier in weaned piglets challenged with lipopolysaccharide. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1096–1105. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Coded | Coded Level | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| AB-8 macroporous resin addition (%) | A | 6 | 8 | 10 |
| Temperature (°C) | B | 40 | 50 | 60 |
| Duration (min) | C | 60 | 80 | 100 |
| pH | D | 4 | 5 | 6 |
| Factors | Coded | Coded Level | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| Temperature (°C) | A | 45 | 50 | 55 |
| Duration (h) | B | 2 | 2.5 | 3 |
| pH | C | 8 | 9 | 10 |
| H2O2 addition (%) | D | 6 | 8 | 10 |
| Source | Sum of Squares | Mean Square | F-Value | p-Value |
|---|---|---|---|---|
| Model | 67.19 | 4.80 | 36.07 | <0.0001 |
| A | 3.01 | 3.01 | 22.65 | 0.0005 |
| B | 0.9039 | 0.9039 | 6.79 | 0.0230 |
| C | 0.7928 | 0.7928 | 5.96 | 0.0311 |
| D | 4.41 | 4.41 | 33.13 | <0.0001 |
| AB | 0.0050 | 0.0050 | 0.0379 | 0.8490 |
| AC | 0.4099 | 0.4099 | 3.08 | 0.1047 |
| AD | 4.77 | 4.77 | 35.85 | <0.0001 |
| BC | 1.57 | 1.57 | 11.77 | 0.0050 |
| BD | 0.0776 | 0.0776 | 0.5834 | 0.4597 |
| CD | 9.28 | 9.28 | 69.71 | <0.0001 |
| A2 | 26.53 | 26.53 | 199.40 | <0.0001 |
| B2 | 16.64 | 16.64 | 125.05 | <0.0001 |
| C2 | 6.36 | 6.36 | 47.79 | <0.0001 |
| D2 | 25.50 | 25.50 | 191.64 | <0.0001 |
| Residual | 1.60 | 0.1330 | ||
| Lack of Fit | 1.49 | 0.1487 | 2.71 | 0.2995 |
| Pure Error | 0.1097 | 0.0549 | ||
| Correlation Total | 68.79 | |||
| CV% | 0.5404 | |||
| R2 | 0.9768 | |||
| Adjusted-R2 | 0.9497 |
| Source | Sum of Squares | Mean Square | F-Value | p-Value |
|---|---|---|---|---|
| Model | 284.18 | 20.30 | 10.03 | 0.0001 |
| A | 5.07 | 5.07 | 2.51 | 0.1394 |
| B | 101.73 | 101.73 | 50.27 | <0.0001 |
| C | 2.70 | 2.70 | 1.33 | 0.2705 |
| D | 16.90 | 16.90 | 8.35 | 0.0136 |
| AB | 2.46 | 2.46 | 1.22 | 0.2914 |
| AC | 4.81 | 4.81 | 2.38 | 0.1491 |
| AD | 0.2652 | 0.2652 | 0.1310 | 0.7236 |
| BC | 0.0992 | 0.0992 | 0.0490 | 0.8285 |
| BD | 7.37 | 7.37 | 3.64 | 0.0805 |
| CD | 12.46 | 12.46 | 6.16 | 0.0289 |
| A2 | 81.89 | 81.89 | 40.46 | <0.0001 |
| B2 | 61.05 | 61.05 | 30.17 | 0.0001 |
| C2 | 68.66 | 68.66 | 33.92 | <0.0001 |
| D2 | 37.81 | 37.81 | 18.68 | 0.0010 |
| Residual | 24.29 | 2.02 | ||
| Lack of Fit | 15.09 | 1.68 | 0.5471 | 0.7877 |
| Pure Error | 9.19 | 3.06 | ||
| Correlation Total | 308.47 | |||
| CV% | 1.77 | |||
| R2 | 0.9213 | |||
| Adjusted-R2 | 0.8294 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, H.; Li, K.; Min, Y.; Qiu, B.; Huang, X.; Luo, J.; Qi, L.; Kang, M.; Xia, P.; Qiao, H.; et al. Rehmannia glutinosa Polysaccharides: Optimization of the Decolorization Process and Antioxidant and Anti-Inflammatory Effects in LPS-Stimulated Porcine Intestinal Epithelial Cells. Antioxidants 2023, 12, 914. https://doi.org/10.3390/antiox12040914
Ren H, Li K, Min Y, Qiu B, Huang X, Luo J, Qi L, Kang M, Xia P, Qiao H, et al. Rehmannia glutinosa Polysaccharides: Optimization of the Decolorization Process and Antioxidant and Anti-Inflammatory Effects in LPS-Stimulated Porcine Intestinal Epithelial Cells. Antioxidants. 2023; 12(4):914. https://doi.org/10.3390/antiox12040914
Chicago/Turabian StyleRen, Heng, Kejie Li, Yan Min, Binhang Qiu, Xiaolu Huang, Jingxin Luo, Liwen Qi, Maoli Kang, Peng Xia, Hanzhen Qiao, and et al. 2023. "Rehmannia glutinosa Polysaccharides: Optimization of the Decolorization Process and Antioxidant and Anti-Inflammatory Effects in LPS-Stimulated Porcine Intestinal Epithelial Cells" Antioxidants 12, no. 4: 914. https://doi.org/10.3390/antiox12040914