
Recent Trends on Mitigative Effect of Probiotics on Oxidative-Stress-Induced Gut Dysfunction in Broilers under Necrotic Enteritis Challenge: A Review
 , , and
, , and
Abstract
:1. Introduction
2. Efficacy of Dietary Probiotics on Enhancement of Gut Health in Conventional Poultry Model
2.1. Intestinal Villi Morphometrics
2.2. Intestinal Epithelial Barrier
2.3. Immune Regulation
2.4. Regulation of Intestinal Inflammation
2.5. Modulation of Intestinal Microbial Composition
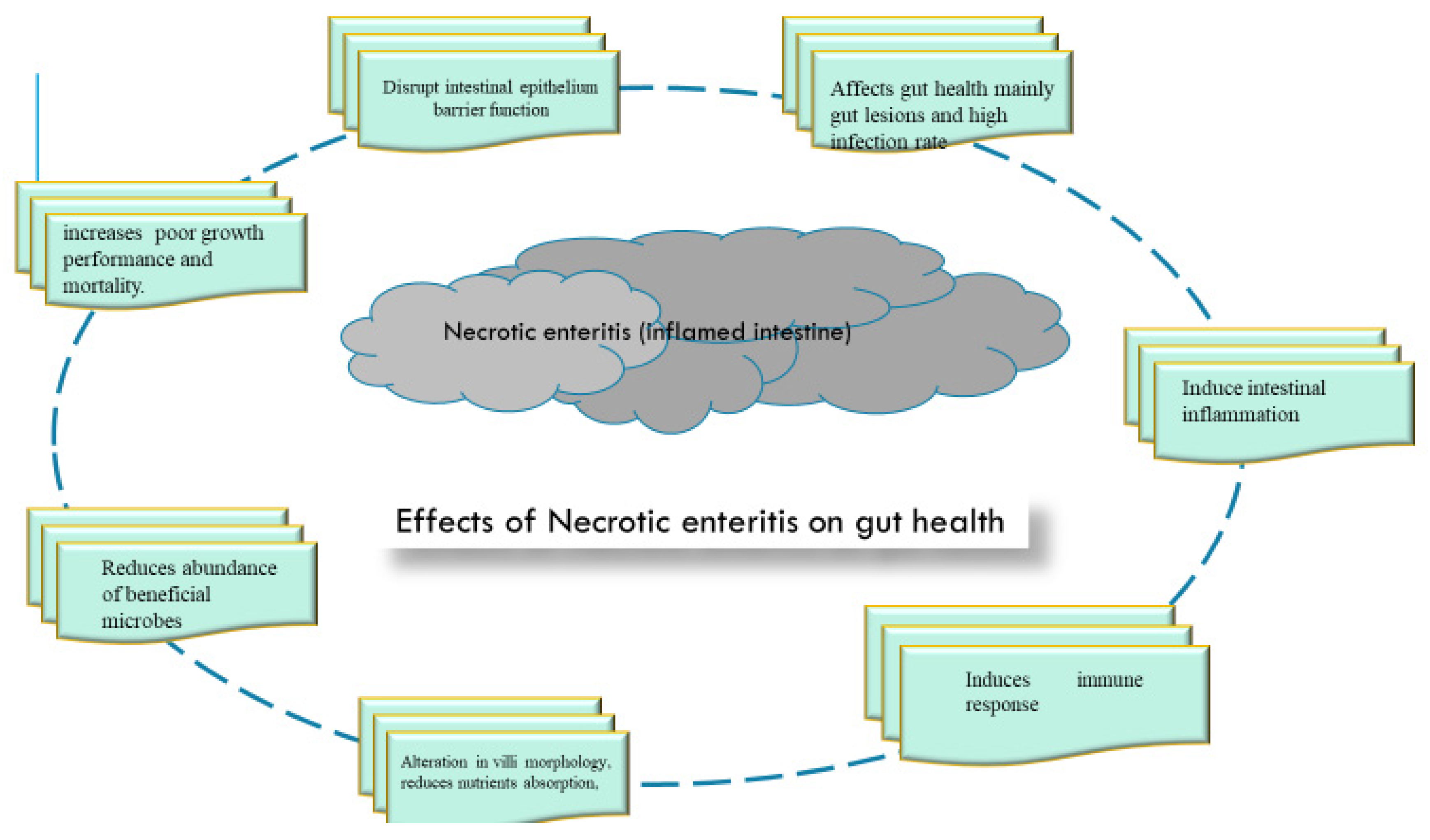
3. Necrotic Enteritis in Broiler Challenged Model
3.1. Effect of Necrotic Enteritis Challenge on Gut Health of Broilers
3.2. Induced Gut Lesions and Pathogen Colonization
3.3. Disruption of Intestinal Epithelial Barrier Function/Integrity
3.4. Alterations in Villi Morphology
3.5. Intestinal Inflammation
3.6. Intestinal Immunity
3.7. Intestinal Microbiota
3.8. Reduced Antioxidant Capacity, Metabolites and Nutrient Transporters
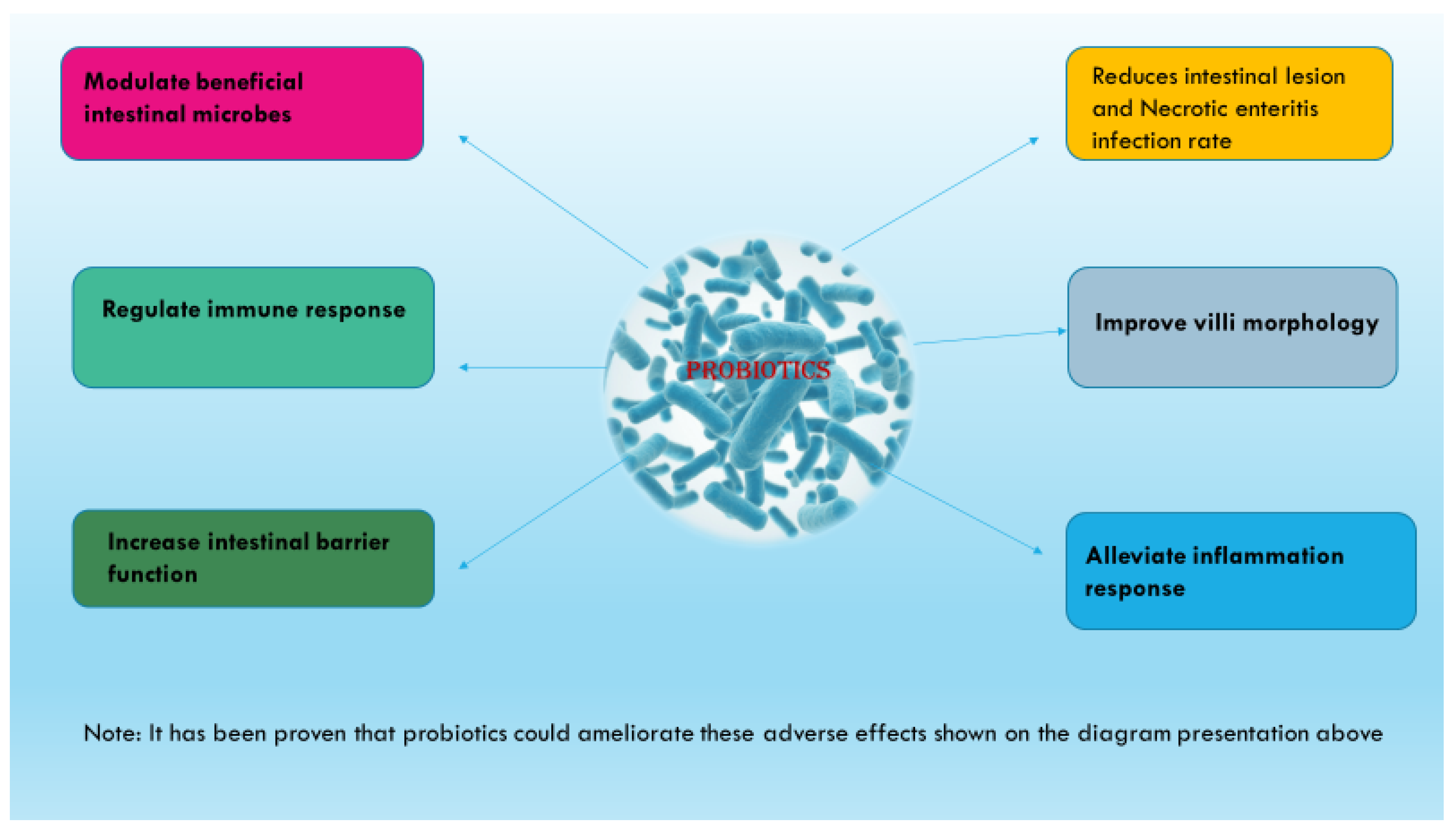
4. Ameliorative Effect of Probiotics Supplementation on Necrotic-Enteritis-Induced Oxidative Stress in the Gut and the Underlying Mechanism of Action
4.1. Pathogen Exclusion and Reduction in Intestinal Lesion Scores
4.2. Improvement in Villi Morphological Structure
4.3. Regulation of Intestinal Epithelial Barrier Function/Integrity
4.4. Regulation of Intestinal Immunity
4.5. Regulation of Intestinal Inflammation
4.6. Antioxidant Capacity
4.7. Modulation of Intestinal Microbiota
4.8. Regulation of Metabolite Synthesis and Nutrient Transporters
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hu, Y.; Cheng, H.; Tao, S. Environmental and human health challenges of industrial livestock and poultry farming in China and their mitigation. Environ. Int. 2017, 107, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Jha, R. Oxidative stress in the poultry gut: Potential challenges and interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyer, H.; Hawken, R.; Murani, E.; Ponsuksili, S.; Wimmers, K. The genetics of feed conversion efficiency traits in a commercial broiler line. Sci. Rep. 2015, 5, 16387. [Google Scholar] [CrossRef] [Green Version]
- Salami, S.A.; Majoka, M.A.; Saha, S.; Garber, A.; Gabarrou, J.-F. Efficacy of dietary antioxidants on broiler oxidative stress, performance and meat quality: Science and market. Avian Biol. Res. 2015, 8, 65–78. [Google Scholar] [CrossRef]
- Jones, D.P. Redefining oxidative stress. Antioxid. Redox Signal. 2006, 8, 1865–1879. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Ballard, S.T.; Hunter, J.H.; Taylor, A.E. Regulation of tight-junction permeability during nutrient absorption across the intestinal epithelium. Annu. Rev. Nutr. 1995, 15, 35–55. [Google Scholar] [CrossRef]
- Min, Y.; Niu, Z.; Sun, T.; Wang, Z.; Jiao, P.; Zi, B.; Chen, P.; Tian, D.; Liu, F. Vitamin E and vitamin C supplementation improves antioxidant status and immune function in oxidative-stressed breeder roosters by up-regulating expression of GSH-Px gene. Poult. Sci. 2018, 97, 1238–1244. [Google Scholar] [CrossRef]
- Liang, F.; Jiang, S.; Mo, Y.; Zhou, G.; Yang, L. Consumption of oxidized soybean oil increased intestinal oxidative stress and affected intestinal immune variables in yellow-feathered broilers. Asian-Australas. J. Anim. Sci. 2015, 28, 1194. [Google Scholar] [CrossRef] [Green Version]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Gharib-Naseri, K.; Dorigam, J.C.; Doranalli, K.; Morgan, N.; Swick, R.A.; Choct, M.; Wu, S.-B. Bacillus amyloliquefaciens CECT 5940 improves performance and gut function in broilers fed different levels of protein and/or under necrotic enteritis challenge. Anim. Nutr. 2021, 7, 185–197. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, Y.; Zeng, D.; Wang, H.; Sun, N.; Xin, J.; Zhou, M.; Yang, H.; Lei, L.; Ling, H. Dietary Probiotic Supplementation Suppresses Subclinical Necrotic Enteritis in Broiler Chickens in a Microbiota-Dependent Manner. Front. Immunol. 2022, 13, 855426. [Google Scholar] [CrossRef] [PubMed]
- Shojadoost, B.; Alizadeh, M.; Boodhoo, N.; Astill, J.; Karimi, S.H.; Shoja Doost, J.; Taha-Abdelaziz, K.; Kulkarni, R.; Sharif, S. Effects of treatment with lactobacilli on necrotic enteritis in broiler chickens. Prob. Antimicrob. Proteins 2022, 14, 1110–1129. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Fu, A.; Gong, L.; Li, W.; Li, Y. Bacillus amyloliquefaciens SC06 alleviates the oxidative stress of IPEC-1 via modulating Nrf2/Keap1 signaling pathway and decreasing ROS production. Appl. Microbiol. Biotechnol. 2017, 101, 3015–3026. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, H.; Yu, Y.; Zhang, R.; Wu, Y.; Yue, M.; Yang, C. Effects of Bacillus coagulans on growth performance, antioxidant capacity, immunity function, and gut health in broilers. Poult. Sci. 2021, 100, 101168. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Al-Sadi, R.; Said, H.M.; Ma, T.Y. Lipopolysaccharide causes an increase in intestinal tight junction permeability in vitro and in vivo by inducing enterocyte membrane expression and localization of TLR-4 and CD14. Am. J. Pathol. 2013, 182, 375–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramlucken, U.; Ramchuran, S.O.; Moonsamy, G.; Lalloo, R.; Thantsha, M.S.; van Rensburg, C.J. A novel Bacillus based multi-strain probiotic improves growth performance and intestinal properties of Clostridium perfringens challenged broilers. Poult. Sci. 2020, 99, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Sandvang, D.; Skjoet-Rasmussen, L.; Cantor, M.D.; Mathis, G.F.; Lumpkins, B.S.; Blanch, A. Effects of feed supplementation with 3 different probiotic Bacillus strains and their combination on the performance of broiler chickens challenged with Clostridium perfringens. Poult. Sci. 2021, 100, 100982. [Google Scholar] [CrossRef]
- Kan, L.; Guo, F.; Liu, Y.; Pham, V.H.; Guo, Y.; Wang, Z. Probiotics Bacillus licheniformis improves intestinal health of subclinical necrotic enteritis-challenged broilers. Front. Microbiol. 2021, 12, 623739. [Google Scholar] [CrossRef]
- Musa, B.B.; Duan, Y.; Khawar, H.; Sun, Q.; Ren, Z.; Elsiddig Mohamed, M.A.; Abbasi, I.H.R.; Yang, X. Bacillus subtilis B21 and Bacillus licheniformis B26 improve intestinal health and performance of broiler chickens with Clostridium perfringens-induced necrotic enteritis. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1039–1049. [Google Scholar]
- Hernandez-Patlan, D.; Solis-Cruz, B.; Pontin, K.P.; Hernandez-Velasco, X.; Merino-Guzman, R.; Adhikari, B.; López-Arellano, R.; Kwon, Y.M.; Hargis, B.M.; Arreguin-Nava, M.A. Impact of a Bacillus direct-fed microbial on growth performance, intestinal barrier integrity, necrotic enteritis lesions, and ileal microbiota in broiler chickens using a laboratory challenge model. Front. Vet. Sci. 2019, 6, 108. [Google Scholar] [CrossRef]
- Hussein, E.O.; Ahmed, S.H.; Abudabos, A.M.; Aljumaah, M.R.; Alkhlulaifi, M.M.; Nassan, M.A.; Suliman, G.M.; Naiel, M.A.; Swelum, A.A. Effect of antibiotic, phytobiotic and probiotic supplementation on growth, blood indices and intestine health in broiler chicks challenged with Clostridium perfringens. Animals 2020, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Emami, N.K.; Calik, A.; White, M.B.; Kimminau, E.A.; Dalloul, R.A. Effect of probiotics and multi-component feed additives on microbiota, gut barrier and immune responses in broiler chickens during subclinical necrotic enteritis. Front. Vet. Sci. 2020, 7, 572142. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic Lactobacillus plantarum promotes intestinal barrier function by strengthening the epithelium and modulating gut microbiota. Front. Microbiol. 2018, 9, 1953. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Shao, Y.; Song, B.; Zhen, W.; Wang, Z.; Guo, Y.; Shahid, M.S.; Nie, W. Effects of Bacillus coagulans supplementation on the growth performance and gut health of broiler chickens with Clostridium perfringens-induced necrotic enteritis. J. Anim. Sci. Biotechnol. 2018, 9, 9. [Google Scholar] [CrossRef]
- Huang, T.; Peng, X.-Y.; Gao, B.; Wei, Q.-L.; Xiang, R.; Yuan, M.-G.; Xu, Z.-H. The effect of Clostridium butyricum on gut microbiota, immune response and intestinal barrier function during the development of necrotic enteritis in chickens. Front. Microbiol. 2019, 10, 2309. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Yang, J.; Ju, Z.; Wu, J.; Wang, L.; Lin, H.; Sun, S. Clostridium butyricum ameliorates Salmonella enteritis induced inflammation by enhancing and improving immunity of the intestinal epithelial barrier at the intestinal mucosal level. Front. Microbiol. 2020, 11, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, S.; Luo, Z.; Liu, D. Supplemental Bacillus subtilis PB6 improves growth performance and gut health in broilers challenged with Clostridium perfringens. J. Immunol. Res. 2021, 2021, 2549541. [Google Scholar] [CrossRef]
- Ekim, B.; Calik, A.; Ceylan, A.; Saçaklı, P. Effects of Paenibacillus xylanexedens on growth performance, intestinal histomorphology, intestinal microflora, and immune response in broiler chickens challenged with Escherichia coli K88. Poult. Sci. 2020, 99, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Wu, X.; Bai, Y.; Wu, X.; Gu, S. Anti-inflammatory and antioxidant activities of probiotic powder containing Lactobacillus plantarum 1.2567 in necrotic enteritis model of broiler chickens. Livest. Sci. 2019, 223, 157–163. [Google Scholar] [CrossRef]
- Shini, S.; Zhang, D.; Aland, R.; Li, X.; Dart, P.; Callaghan, M.; Speight, R.; Bryden, W. Probiotic Bacillus amyloliquefaciens H57 ameliorates subclinical necrotic enteritis in broiler chicks by maintaining intestinal mucosal integrity and improving feed efficiency. Poult. Sci. 2020, 99, 4278–4293. [Google Scholar] [CrossRef] [PubMed]
- Aljumaah, M.R.; Alkhulaifi, M.M.; Abudabos, A.M.; Aljumaah, R.S.; Alsaleh, A.N.; Stanley, D. Bacillus subtilis PB6 based probiotic supplementation plays a role in the recovery after the necrotic enteritis challenge. PLoS ONE 2020, 15, e0232781. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Xi, Y.; Xia, Y.; Wu, T.; Zhao, D.; Zhang, Z.; Ding, B. Dietary Lactobacillus fermentum and Bacillus coagulans supplementation modulates intestinal immunity and microbiota of broiler chickens challenged by Clostridium perfringens. Front. Vet. Sci. 2021, 8, 680742. [Google Scholar] [CrossRef] [PubMed]
- Landoni, M.F.; Albarellos, G. The use of antimicrobial agents in broiler chickens. Vet. J. 2015, 205, 21–27. [Google Scholar] [CrossRef]
- Ding, S.; Yan, W.; Ma, Y.; Fang, J. The impact of probiotics on gut health via alternation of immune status of monogastric animals. Anim. Nutr. 2021, 7, 24–30. [Google Scholar] [CrossRef]
- Gao, J.; Wang, R.; Liu, J.; Wang, W.; Chen, Y.; Cai, W. Effects of novel microecologics combined with traditional Chinese medicine and probiotics on growth performance and health of broilers. Poult. Sci. 2022, 101, 101412. [Google Scholar] [CrossRef]
- Ebeid, T.; Al-Homidan, I.; Fathi, M.; Al-Jamaan, R.; Mostafa, M.; Abou-Emera, O.; El-Razik, M.A.; Alkhalaf, A. Impact of probiotics and/or organic acids supplementation on growth performance, microbiota, antioxidative status, and immune response of broilers. Ital. J. Anim. Sci. 2021, 20, 2263–2273. [Google Scholar] [CrossRef]
- Qiu, K.; Wang, X.; Zhang, H.; Wang, J.; Qi, G.; Wu, S. Dietary supplementation of a new probiotic compound improves the growth performance and health of broilers by altering the composition of cecal microflora. Biology 2022, 11, 633. [Google Scholar] [CrossRef]
- Bao, C.; Zhang, W.; Wang, J.; Liu, Y.; Cao, H.; Li, F.; Liu, S.; Shang, Z.; Cao, Y.; Dong, B. The Effects of Dietary Bacillus amyloliquefaciens TL106 Supplementation, as an Alternative to Antibiotics, on Growth Performance, Intestinal Immunity, Epithelial Barrier Integrity, and Intestinal Microbiota in Broilers. Animals 2022, 12, 3085. [Google Scholar] [CrossRef]
- Gyawali, I.; Zeng, Y.; Zhou, J.; Li, J.; Wu, T.; Shu, G.; Jiang, Q.; Zhu, C. Effect of novel Lactobacillus paracaesi microcapsule on growth performance, gut health and microbiome community of broiler chickens. Poult. Sci. 2022, 101, 101912. [Google Scholar] [CrossRef]
- He, T.; Long, S.; Mahfuz, S.; Wu, D.; Wang, X.; Wei, X.; Piao, X. Effects of probiotics as antibiotics substitutes on growth performance, serum biochemical parameters, intestinal morphology, and barrier function of broilers. Animals 2019, 9, 985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, S.; Sun, Y.; Wang, Y.; Sa, R.; Zhang, H. Bacillus amyloliquefaciens spray improves the growth performance, immune status, and respiratory mucosal barrier in broiler chickens. Poult. Sci. 2019, 98, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Gao, S.; Hou, L.; Li, A.; Zhu, L.-Q.; Dong, J.; Chen, F. Selenium-enriched Bacillus subtilis Improves Growth Performance, Antioxidant Capacity, Immune Status and Gut Health of Broiler Chickens. Biol. Trace Elem. Res. 2022, PPR575593. [Google Scholar] [CrossRef]
- Xiao, D.; Wang, Z.; Dai, X.; Hu, Y.; Zhong, M.; Xiong, L.; Jiang, C.; Khalique, A.; Ni, X.; Zeng, D. Effects of Bacillus methylotrophicus SY200 Supplementation on Growth Performance, Antioxidant Status, Intestinal Morphology, and Immune Function in Broiler Chickens. Probiotics Antimicrob. Proteins 2022, 1–16. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, Y.; Shen, Y.; Li, Q.; Lan, J.; Wu, Y.; Zhang, R.; Cao, G.; Yang, C. Effects of Bacillus subtilis and Bacillus licheniformis on growth performance, immunity, short chain fatty acid production, antioxidant capacity, and cecal microflora in broilers. Poult. Sci. 2021, 100, 101358. [Google Scholar] [CrossRef]
- Deraz, S.F.; Elkomy, A.E.; Khalil, A.A. Assessment of probiotic-supplementation on growth performance, lipid peroxidation, antioxidant capacity, and cecal microflora in broiler chickens. J. Appl. Pharm. Sci. 2019, 9, 030–039. [Google Scholar]
- Bai, K.; Feng, C.; Jiang, L.; Zhang, L.; Zhang, J.; Zhang, L.; Wang, T. Dietary effects of Bacillus subtilis fmbj on growth performance, small intestinal morphology, and its antioxidant capacity of broilers. Poult. Sci. 2018, 97, 2312–2321. [Google Scholar] [CrossRef] [PubMed]
- Al-Khalaifa, H.; Al-Nasser, A.; Al-Surayee, T.; Al-Kandari, S.; Al-Enzi, N.; Al-Sharrah, T.; Ragheb, G.; Al-Qalaf, S.; Mohammed, A. Effect of dietary probiotics and prebiotics on the performance of broiler chickens. Poult. Sci. 2019, 98, 4465–4479. [Google Scholar] [CrossRef]
- Ashraf, R.; Shah, N.P. Immune system stimulation by probiotic microorganisms. Crit. Rev. Food Sci. Nutr. 2014, 54, 938–956. [Google Scholar] [CrossRef]
- Jazi, V.; Foroozandeh, A.; Toghyani, M.; Dastar, B.; Koochaksaraie, R.R. Effects of Pediococcus acidilactici, mannan-oligosaccharide, butyric acid and their combination on growth performance and intestinal health in young broiler chickens challenged with Salmonella typhimurium. Poult. Sci. 2018, 97, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Horng, Y.-B.; Yu, Y.-H.; Dybus, A.; Hsiao, F.S.-H.; Cheng, Y.-H. Antibacterial activity of Bacillus species-derived surfactin on Brachyspira hyodysenteriae and Clostridium perfringens. AMB Express 2019, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Bodinga, B.M.; Hayat, K.; Liu, X.; Zhou, J.; Yang, X.; Ismaila, A.; Soomro, R.; Ren, Z.; Zhang, W. Effects of Bacillus subtilis DSM 32315 on Immunity, nutrient transporters and functional diversity of cecal microbiome of broiler chickens in necrotic enteritis challenge. J. World’s Poult. Res. 2020, 10, 527–544. [Google Scholar] [CrossRef]
- Emami, N.K.; White, M.B.; Calik, A.; Kimminau, E.A.; Dalloul, R.A. Managing broilers gut health with antibiotic-free diets during subclinical necrotic enteritis. Poult. Sci. 2021, 100, 101055. [Google Scholar] [CrossRef]
- Qiu, K.; Li, C.; Wang, J.; Qi, G.-H.; Gao, J.; Zhang, H.; Shugeng, W. Effects of dietary supplementation with Bacillus subtilis, as an alternative to antibiotics, on growth performance, serum immunity, and intestinal health in broiler chickens. Front. Nutr. 2021, 940. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Li, R.; Liu, Y.; Ma, L.; Zha, J.; Qiao, X.; Chai, T.; Wu, B. Benefit of dietary supplementation with Bacillus subtilis BYS2 on growth performance, immune response, and disease resistance of broilers. Probiotics Antimicrob. Proteins 2020, 12, 1385–1397. [Google Scholar] [CrossRef]
- Liu, L.; Ling, H.; Zhang, W.; Zhou, Y.; Li, Y.; Peng, N.; Zhao, S. Functional comparison of Clostridium butyricum and sodium butyrate supplementation on growth, intestinal health, and the anti-inflammatory response of broilers. Front. Microbiol. 2022, 13, 914212. [Google Scholar] [CrossRef]
- Liu, C.; Shih, Y.; Tang, P.; Lin, L.; Lee, T. Effects of dietary supplementation with Bacillus spp. and Debaryomyces spp. on broiler’s growth performance, serum characteristics, intestinal microflora and antioxidant activity. Ital. J. Anim. Sci. 2022, 21, 717–728. [Google Scholar] [CrossRef]
- Lin, J.; Comi, M.; Vera, P.; Alessandro, A.; Qiu, K.; Wang, J.; Wu, S.-G.; Qi, G.-H.; Zhang, H.-J. Effects of Saccharomyces cerevisiae hydrolysate on growth performance, immunity function, and intestinal health in broilers. Poult. Sci. 2023, 102, 102237. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, W.; Zhang, H.; Wang, J.; Zhang, W.; Gao, J.; Wu, S.; Qi, G. Supplemental Bacillus subtilis DSM 32315 manipulates intestinal structure and microbial composition in broiler chickens. Sci. Rep. 2018, 8, 15358. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, T.M.; Sun, W.; Bumbie, G.Z.; Elokil, A.A.; Mohammed, K.A.F.; Zebin, R.; Hu, P.; Wu, L.; Tang, Z. Feeding Bacillus subtilis ATCC19659 to broiler chickens enhances growth performance and immune function by modulating intestinal morphology and cecum microbiota. Front. Microbiol. 2022, 12, 4152. [Google Scholar] [CrossRef]
- Deng, S.; Hu, S.; Xue, J.; Yang, K.; Zhuo, R.; Xiao, Y.; Fang, R. Productive performance, serum antioxidant status, tissue selenium deposition, and gut health analysis of broiler chickens supplemented with selenium and probiotics—A pilot study. Animals 2022, 12, 1086. [Google Scholar] [CrossRef]
- Ahmat, M.; Cheng, J.; Abbas, Z.; Cheng, Q.; Fan, Z.; Ahmad, B.; Hou, M.; Osman, G.; Guo, H.; Wang, J. Effects of Bacillus amyloliquefaciens LFB112 on Growth Performance, Carcass Traits, Immune, and Serum Biochemical Response in Broiler Chickens. Antibiotics 2021, 10, 1427. [Google Scholar] [CrossRef]
- Soumeh, E.A.; Cedeno, A.D.R.C.; Niknafs, S.; Bromfield, J.; Hoffman, L.C. The efficiency of probiotics administrated via different routes and doses in enhancing production performance, meat quality, gut morphology, and microbial profile of broiler chickens. Animals 2021, 11, 3607. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, Y.; Liu, M.; Li, J.; Lai, W.; Geng, S.; Yuan, T.; Liu, Y.; Di, Y.; Zhang, W. Effects of dietary Bacillus amyloliquefaciens CECT 5940 supplementation on growth performance, antioxidant status, immunity, and digestive enzyme activity of broilers fed corn-wheat-soybean meal diets. Poult. Sci. 2022, 101, 101585. [Google Scholar] [CrossRef]
- Sun, X.; Chen, D.; Deng, S.; Zhang, G.; Peng, X.; Sa, R. Using combined Lactobacillus and quorum quenching enzyme supplementation as an antibiotic alternative to improve broiler growth performance, antioxidative status, immune response, and gut microbiota. Poult. Sci. 2022, 101, 101997. [Google Scholar] [CrossRef]
- Abeddargahi, F.; Darmani Kuhi, H.; Rafiei, F.; Roostaie-Alimehr, M.; Takalu, Z.; Sajedi, R.H.; Mohammadpour, F. The effect of probiotic and fermented soybean meal based on Bacillus subtilis spore on growth performance, gut morphology, immune response and dry matter digestibility in broiler chickens. Ital. J. Anim. Sci. 2022, 21, 1642–1650. [Google Scholar] [CrossRef]
- Wang, B.; Zhou, Y.; Tang, L.; Zeng, Z.; Gong, L.; Wu, Y.; Li, W.-F. Effects of Bacillus amyloliquefaciens instead of antibiotics on growth performance, intestinal health, and intestinal microbiota of broilers. Front. Vet. Sci. 2021, 8, 679368. [Google Scholar] [CrossRef]
- Meyer, M.M.; Fries-Craft, K.A.; Bobeck, E.A. Composition and inclusion of probiotics in broiler diets alter intestinal permeability and spleen immune cell profiles without negatively affecting performance. J. Anim. Sci. 2020, 98, skz383. [Google Scholar] [CrossRef]
- Zhang, L.; Ben Said, L.; Hervé, N.; Zirah, S.; Diarra, M.S.; Fliss, I. Effects of drinking water supplementation with Lactobacillus reuteri, and a mixture of reuterin and microcin J25 on the growth performance, caecal microbiota and selected metabolites of broiler chickens. J. Anim. Sci. Biotechnol. 2022, 13, 34. [Google Scholar] [CrossRef]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Lai, J.; Khalique, A.; Li, G.; Pan, K.; Jing, B.; Zeng, D. Probiotic enhanced intestinal immunity in broilers against subclinical necrotic enteritis. Front. Immunol. 2017, 8, 1592. [Google Scholar] [CrossRef] [Green Version]
- Kai, Y. Intestinal villus structure contributes to even shedding of epithelial cells. Biophys. J. 2021, 120, 699–710. [Google Scholar] [CrossRef]
- Ye, Y.; Li, Z.; Wang, P.; Zhu, B.; Zhao, M.; Huang, D.; Ye, Y.; Ding, Z.; Li, L.; Wan, G. Effects of probiotic supplements on growth performance and intestinal microbiota of partridge shank broiler chicks. Peer J. 2021, 9, e12538. [Google Scholar] [CrossRef]
- Uni, Z. Early development of small intestinal function. In Avian Gut Function in Health and Disease; CABI: Wallingford, UK, 2006; pp. 29–42. [Google Scholar]
- Celi, P.; Verlhac, V.; Calvo, E.P.; Schmeisser, J.; Kluenter, A.-M. Biomarkers of gastrointestinal functionality in animal nutrition and health. Anim. Feed Sci. Technol. 2019, 250, 9–31. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.-D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Bibi, S.; Du, M.; Suzuki, T.; Zhu, M.-J. Regulation of the intestinal tight junction by natural polyphenols: A mechanistic perspective. Crit. Rev. Food Sci. Nutr. 2017, 57, 3830–3839. [Google Scholar] [CrossRef]
- Pandey, U.; Aich, P. Postnatal intestinal mucosa and gut microbial composition develop hand in hand: A mouse study. Biomed. J. 2022. [Google Scholar] [CrossRef]
- Broom, L.J.; Kogut, M.H. Inflammation: Friend or foe for animal production? Poult. Sci. 2018, 97, 510–514. [Google Scholar] [CrossRef]
- Dongarrà, M.L.; Rizzello, V.; Muccio, L.; Fries, W.; Cascio, A.; Bonaccorsi, I.; Ferlazzo, G. Mucosal immunology and probiotics. Curr. Allergy Asthma Rep. 2013, 13, 19–26. [Google Scholar] [CrossRef]
- Bar-Shira, E.; Friedman, A. Development and adaptations of innate immunity in the gastrointestinal tract of the newly hatched chick. Dev. Comp. Immunol. 2006, 30, 930–941. [Google Scholar] [CrossRef]
- Hoffman, W.; Lakkis, F.G.; Chalasani, G. B cells, antibodies, and more. Clin. J. Am. Soc. Nephrol. 2016, 11, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Zhu, M.-J. AMP-activated protein kinase: A therapeutic target in intestinal diseases. Open Biol. 2017, 7, 170104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Watford, W.T.; Moriguchi, M.; Morinobu, A.; O’Shea, J.J. The biology of IL-12: Coordinating innate and adaptive immune responses. Cytokine Growth Factor Rev. 2003, 14, 361–368. [Google Scholar] [CrossRef]
- Fasina, Y.O.; Lillehoj, H.S. Characterization of intestinal immune response to Clostridium perfringens infection in broiler chickens. Poult. Sci. 2019, 98, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.W.; Blase, J.R.; Zhang, X.; Eickhoff, C.S.; Hoft, D.F. Th17 cells are more protective than Th1 cells against the intracellular parasite Trypanosoma cruzi. PLoS Pathog. 2016, 12, e1005902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef]
- Kogut, M.H.; Arsenault, R.J. Gut health: The new paradigm in food animal production. Front. Vet. Sci. 2016, 3, 71. [Google Scholar] [CrossRef] [Green Version]
- Plöger, S.; Stumpff, F.; Penner, G.B.; Schulzke, J.D.; Gäbel, G.; Martens, H.; Shen, Z.; Günzel, D.; Aschenbach, J.R. Microbial butyrate and its role for barrier function in the gastrointestinal tract. Ann. N. Y. Acad. Sci. 2012, 1258, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Xu, S.; Zeng, D.; Ni, X.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K. Disruption in the cecal microbiota of chickens challenged with Clostridium perfringens and other factors was alleviated by Bacillus licheniformis supplementation. PLoS ONE 2017, 12, e0182426. [Google Scholar] [CrossRef] [Green Version]
- Calik, A.; Omara, I.I.; White, M.B.; Evans, N.P.; Karnezos, T.P.; Dalloul, R.A. Dietary non-drug feed additive as an alternative for antibiotic growth promoters for broilers during a necrotic enteritis challenge. Microorganisms 2019, 7, 257. [Google Scholar] [CrossRef] [Green Version]
- Rajput, D.S.; Zeng, D.; Khalique, A.; Rajput, S.S.; Wang, H.; Zhao, Y.; Sun, N.; Ni, X. Pretreatment with probiotics ameliorate gut health and necrotic enteritis in broiler chickens, a substitute to antibiotics. AMB Express 2020, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Shini, S.; Bryden, W. Probiotics and gut health: Linking gut homeostasis and poultry productivity. Anim. Prod. Sci. 2021, 62, 1090–1112. [Google Scholar] [CrossRef]
- Emami, N.K.; Calik, A.; White, M.B.; Young, M.; Dalloul, R.A. Necrotic enteritis in broiler chickens: The role of tight junctions and mucosal immune responses in alleviating the effect of the disease. Microorganisms 2019, 7, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, Y.; Xu, S.; Yang, J.; Wang, K.; Zhan, X. Bacillus subtilis DSM29784 alleviates negative effects on growth performance in broilers by improving the intestinal health under necrotic enteritis challenge. Front. Microbiol. 2021, 12, 723187. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The successful experimental induction of necrotic enteritis in chickens by Clostridium perfringens: A critical review. Vet. Res. 2012, 43, 74. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.J. Necrotic enteritis predisposing factors in broiler chickens. Avian Pathol. 2016, 45, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Smyth, J.A. Pathology and diagnosis of necrotic enteritis: Is it clear-cut? Avian Pathol. 2016, 45, 282–287. [Google Scholar] [CrossRef]
- Saitoh, Y.; Suzuki, H.; Tani, K.; Nishikawa, K.; Irie, K.; Ogura, Y.; Tamura, A.; Tsukita, S.; Fujiyoshi, Y. Structural insight into tight junction disassembly by Clostridium perfringens enterotoxin. Science 2015, 347, 775–778. [Google Scholar] [CrossRef]
- Thung, T.; Mahyudin, N.A.; Basri, D.F.; Radzi, C.W.M.; Nakaguchi, Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance of Salmonella Enteritidis and Salmonella Typhimurium in raw chicken meat at retail markets in Malaysia. Poult. Sci. 2016, 95, 1888–1893. [Google Scholar] [CrossRef]
- Brown, S.P.; Cornell, S.J.; Sheppard, M.; Grant, A.J.; Maskell, D.J.; Grenfell, B.T.; Mastroeni, P. Intracellular demography and the dynamics of Salmonella enterica infections. PLoS Biol. 2006, 4, e349. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-W.; Jia, H.-J.; Zhang, H.-J.; Wang, J.; Lv, H.-Y.; Wu, S.-G.; Qi, G.-H. Supplemental plant extracts from flos lonicerae in combination with baikal skullcap attenuate intestinal disruption and modulate gut microbiota in laying hens challenged by Salmonella pullorum. Front. Microbiol. 2019, 10, 1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abudabos, A.M.; Aljumaah, M.R.; Alkhulaifi, M.M.; Alabdullatif, A.; Suliman, G.M.; Sulaiman, A.R.A. Comparative effects of Bacillus subtilis and Bacillus licheniformis on live performance, blood metabolites and intestinal features in broiler inoculated with Salmonella infection during the finisher phase. Microb. Pathog. 2020, 139, 103870. [Google Scholar] [CrossRef]
- Tarabees, R.; Gafar, K.M.; El-Sayed, M.S.; Shehata, A.A.; Ahmed, M. Effects of dietary supplementation of probiotic mix and prebiotic on growth performance, cecal microbiota composition, and protection against Escherichia coli O78 in broiler chickens. Probiotics Antimicrob. Proteins 2019, 11, 981–989. [Google Scholar] [CrossRef]
- Tan, J.; Liu, S.; Guo, Y.; Applegate, T.J.; Eicher, S.D. Dietary L-arginine supplementation attenuates lipopolysaccharide-induced inflammatory response in broiler chickens. Br. J. Nutr. 2014, 111, 1394–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Necrotic enteritis in broilers: An updated review on the pathogenesis. Avian Pathol. 2011, 40, 341–347. [Google Scholar] [CrossRef]
- LaRock, D.L.; Chaudhary, A.; Miller, S.I. Salmonellae interactions with host processes. Nat. Rev. Microb. 2015, 13, 191–205. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, D.; Wang, H.; Qing, X.; Sun, N.; Xin, J.; Luo, M.; Khalique, A.; Pan, K.; Shu, G. Dietary probiotic Bacillus licheniformis H2 enhanced growth performance, morphology of small intestine and liver, and antioxidant capacity of broiler chickens against Clostridium perfringens–induced subclinical necrotic enteritis. Probiotics Antimicrob. Proteins 2020, 12, 883–895. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Y.; Xu, H.; Liang, C.; Zhai, Z. Bacillus amyloliquefaciens BLCC1-0238 Alone or in Combination with Mannan-Oligosaccharides Alleviates Subclinical Necrotic Enteritis in Broilers. Probiotics Antimicrob. Proteins 2022, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-Y.; Chou, C.-H.; Wang, C. The effects of feed supplementing Akkemansia muciniphila on incidence, severity, and gut microbiota of necrotic enteritis in chickens. Poult. Sci. 2022, 101, 101751. [Google Scholar] [CrossRef]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Pretreatment with probiotic Enterococcus faecium NCIMB 11181 ameliorates necrotic enteritis-induced intestinal barrier injury in broiler chickens. Sci. Rep. 2019, 9, 10256. [Google Scholar] [CrossRef] [Green Version]
- Whelan, R.A.; Doranalli, K.; Rinttilä, T.; Vienola, K.; Jurgens, G.; Apajalahti, J. The impact of Bacillus subtilis DSM 32315 on the pathology, performance, and intestinal microbiome of broiler chickens in a necrotic enteritis challenge. Poult. Sci. 2019, 98, 3450–3463. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Xin, J.; Luo, M.; Khalique, A.; Dan, Y.; Pan, K.; Jing, B. Probiotic Lactobacillus johnsonii BS15 improves blood parameters related to immunity in broilers experimentally infected with subclinical necrotic enteritis. Front. Microbiol. 2018, 9, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, M.; McCartney, E.; Knox, A.; Francesch, M.; Oka, K.; Wada, K.; Ideno, M.; Uno, K.; Kozłowski, K.; Jankowski, J. Effects of the butyric acid-producing strain Clostridium butyricum MIYAIRI 588 on broiler and piglet zootechnical performance and prevention of necrotic enteritis. Anim. Sci. J. 2018, 89, 895–905. [Google Scholar] [CrossRef]
- Sun, N.; Xue, Y.; Wei, S.; Wu, B.; Wang, H.; Zeng, D.; Zhao, Y.; Khalique, A.; Pan, K.; Zeng, Y. Compound Probiotics Improve Body Growth Performance by Enhancing Intestinal Development of Broilers with Subclinical Necrotic Enteritis. Probiotics Antimicrob. Proteins 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sokale, A.; Menconi, A.; Mathis, G.; Lumpkins, B.; Sims, M.; Whelan, R.; Doranalli, K. Effect of Bacillus subtilis DSM 32315 on the intestinal structural integrity and growth performance of broiler chickens under necrotic enteritis challenge. Poult. Sci. 2019, 98, 5392–5400. [Google Scholar] [CrossRef]
- Bae, D.; Lee, J.-W.; Chae, J.-P.; Kim, J.-W.; Eun, J.-S.; Lee, K.-W.; Seo, K.-H. Characterization of a novel bacteriophage φCJ22 and its prophylactic and inhibitory effects on necrotic enteritis and Clostridium perfringens in broilers. Poult. Sci. 2021, 100, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.H.; Zhang, N.; Han, J.C.; Chang, C.W.; Hsiao, F.S.H.; Yu, Y.H. Optimization of surfactin production from Bacillus subtilis in fermentation and its effects on Clostridium perfringens-induced necrotic enteritis and growth performance in broilers. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1232–1244. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-H.; Horng, Y.-B.; Dybus, A.; Yu, Y.-H. Bacillus licheniformis-fermented products improve growth performance and intestinal gut morphology in broilers under Clostridium perfringens challenge. J. Poult. Sci 2021, 58, 30–39. [Google Scholar] [CrossRef]
- Eeckhaut, V.; Wang, J.; Van Parys, A.; Haesebrouck, F.; Joossens, M.; Falony, G.; Raes, J.; Ducatelle, R.; Van Immerseel, F. The probiotic Butyricicoccus pullicaecorum reduces feed conversion and protects from potentially harmful intestinal microorganisms and necrotic enteritis in broilers. Front. Microbiol. 2016, 7, 1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keerqin, C.; Rhayat, L.; Zhang, Z.-H.; Gharib-Naseri, K.; Kheravii, S.; Devillard, E.; Crowley, T.; Wu, S.-B. Probiotic Bacillus subtilis 29,784 improved weight gain and enhanced gut health status of broilers under necrotic enteritis condition. Poult. Sci. 2021, 100, 100981. [Google Scholar] [CrossRef]
- Qing, X.; Zeng, D.; Wang, H.; Ni, X.; Liu, L.; Lai, J.; Khalique, A.; Pan, K.; Jing, B. Preventing subclinical necrotic enteritis through Lactobacillus johnsonii BS15 by ameliorating lipid metabolism and intestinal microflora in broiler chickens. AMB Express 2017, 7, 139. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.J.K.; Sakomura, N.K.; de Paula Dorigam, J.C.; Doranalli, K.; Soares, L.; da Silva Viana, G. Bacillus amyloliquefaciens CECT 5940 alone or in combination with antibiotic growth promoters improves performance in broilers under enteric pathogen challenge. Poult. Sci. 2019, 98, 4391–4400. [Google Scholar] [CrossRef]
- Li, Z.; Wang, W.; Liu, D.; Guo, Y. Effects of Lactobacillus acidophilus on the growth performance and intestinal health of broilers challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2018, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, W.; Liu, D.; Guo, Y. Effects of Lactobacillus acidophilus on gut microbiota composition in broilers challenged with Clostridium perfringens. PLoS ONE 2017, 12, e0188634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.; Smith, A.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult. Sci. 2020, 99, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Rafii, F. Effects of bile acids and nisin on the production of enterotoxin by Clostridium perfringens in a nutrient-rich medium. Int. J. Microbiol. 2018, 2018, 7276523. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Yang, K.; Zhang, A.; Chang, W.; Zheng, A.; Chen, Z.; Cai, H.; Liu, G. Effects of Lactobacillus acidophilus on the growth performance, immune response, and intestinal barrier function of broiler chickens challenged with Escherichia coli O157. Poult. Sci. 2021, 100, 101323. [Google Scholar] [CrossRef] [PubMed]
- Voloshanenko, O.; Erdmann, G.; Dubash, T.D.; Augustin, I.; Metzig, M.; Moffa, G.; Hundsrucker, C.; Kerr, G.; Sandmann, T.; Anchang, B. Wnt secretion is required to maintain high levels of Wnt activity in colon cancer cells. Nat. Comm. 2013, 4, 2610. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Luo, L.; Zhang, Y.; Wang, Z.; Xia, Z. Effects of the dietary probiotic, Enterococcus faecium NCIMB11181, on the intestinal barrier and system immune status in Escherichia coli O78-challenged broiler chickens. Probiotics Antimicrob. Proteins 2019, 11, 946–956. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.H.; Teng, P.Y.; Lee, T.T.; Yu, B. Effects of multi-strain probiotic supplementation on intestinal microbiota, tight junctions, and inflammation in young broiler chickens challenged with Salmonella enterica subsp. enterica. Asian-Australas. J. Anim. Sci. 2020, 33, 1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Khan, M.; Ghosh, C.; Bank, S.; Maiti, S. The role of Dermcidin isoform-2 in the occurrence and severity of Diabetes. Sci. Rep. 2017, 7, 8252. [Google Scholar] [CrossRef] [Green Version]
- Enko, D.; Meinitzer, A.; Mangge, H.; Kriegshäuser, G.; Halwachs-Baumann, G.; Reininghaus, E.Z.; Bengesser, S.A.; Schnedl, W.J. Concomitant prevalence of low serum diamine oxidase activity and carbohydrate malabsorption. Can. J. Gastroenterol. Hepatol. 2016, 2016, 4893501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chelakkot, C.; Choi, Y.; Kim, D.-K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.-S.; Jee, Y.-K.; Gho, Y.S. Akkermansia muciniphila-derived extracellular vesicles influence gut permeability through the regulation of tight junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef] [Green Version]
- Lin, E.-R.; Cheng, Y.-H.; Hsiao, F.S.-H.; Proskura, W.S.; Dybus, A.; Yu, Y.-H. Optimization of solid-state fermentation conditions of Bacillus licheniformis and its effects on Clostridium perfringens-induced necrotic enteritis in broilers. Rev. Bras. Zootec. 2019, 48. [Google Scholar] [CrossRef] [Green Version]
- Shanmugasundaram, R.; Markazi, A.; Mortada, M.; Ng, T.; Applegate, T.; Bielke, L.; Syed, B.; Pender, C.; Curry, S.; Murugesan, G. Research Note: Effect of synbiotic supplementation on caecal Clostridium perfringens load in broiler chickens with different necrotic enteritis challenge models. Poult. Sci. 2020, 99, 2452–2458. [Google Scholar] [CrossRef]
- Zhen, W.; Shao, Y.; Gong, X.; Wu, Y.; Geng, Y.; Wang, Z.; Guo, Y. Effect of dietary Bacillus coagulans supplementation on growth performance and immune responses of broiler chickens challenged by Salmonella enteritidis. Poult. Sci. 2018, 97, 2654–2666. [Google Scholar] [CrossRef]
- Tristram, D.A.; Hicks, W.; Hard, R. Respiratory syncytial virus and human bronchial epithelium. Arch. Otolaryngol. Head Neck Surg. 1998, 124, 777–783. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.S.; Akaike, T.; Okamoto, S.; Kubota, T.; Yoshitake, J.; Sawa, T.; Miyamoto, Y.; Tamura, F.; Maeda, H. Role of nitric oxide in host defense in murine salmonellosis as a function of its antibacterial and antiapoptotic activities. Infect. Immun. 2002, 70, 3130–3142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, W.; Lv, Z.; Liu, D.; Guo, Y. Bacillus subtilis and yeast cell wall improve the intestinal health of broilers challenged by Clostridium perfringens. Br. Poult. Sci. 2017, 58, 635–643. [Google Scholar] [CrossRef]
- Wang, W.; Li, Z.; Han, Q.; Guo, Y.; Zhang, B.; D’inca, R. Dietary live yeast and mannan-oligosaccharide supplementation attenuate intestinal inflammation and barrier dysfunction induced by Escherichia coli in broilers. Br. J. Nutr. 2016, 116, 1878–1888. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.T.; Lillehoj, H.S. The role of host genetic factors and host immunity in necrotic enteritis. Avian Pathol. 2016, 45, 313–316. [Google Scholar] [CrossRef]
- Macpherson, A.; McCoy, K.; Johansen, F.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Lin, Y.; Zeng, D.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K.; Jing, B. Bacillus licheniformis normalize the ileum microbiota of chickens infected with necrotic enteritis. Sci. Rep. 2018, 8, 1744. [Google Scholar] [CrossRef] [Green Version]
- Lange, K.; Buerger, M.; Stallmach, A.; Bruns, T. Effects of antibiotics on gut microbiota. Dig. Dis. 2016, 34, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Proctor, A.; Phillips, G.J. Differential effects of bacitracin methylene disalicylate (BMD) on the distal colon and cecal microbiota of young broiler chickens. Front. Vet. Sci. 2019, 6, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.H.; Teng, P.Y.; Lee, T.T.; Yu, B. The effects of the supplementation of multi-strain probiotics on intestinal microbiota, metabolites and inflammation of young SPF chickens challenged with Salmonella enterica subsp. enterica. Anim. Sci. J. 2019, 90, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Rho, J.-H.; Wright, D.P.; Christie, D.L.; Clinch, K.; Furneaux, R.H.; Roberton, A.M. A novel mechanism for desulfation of mucin: Identification and cloning of a mucin-desulfating glycosidase (sulfoglycosidase) from Prevotella strain RS2. J. Bacteriol. 2005, 187, 1543–1551. [Google Scholar] [CrossRef] [Green Version]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Sallam, E.A.; Mohammed, L.S.; Elbasuni, S.S.; Azam, A.E.; Soliman, M.M. Impacts of Microbial based Therapy on Growth Performance, Intestinal Health, Carcass Traits and Economic Efficiency of Clostridium perfringens-Infected Cobb and Arbor Acres Broilers. Vet. Med. Sci. 2021, 7, 773–791. [Google Scholar] [CrossRef]
- Bortoluzzi, C.; Serpa Vieira, B.; de Paula Dorigam, J.C.; Menconi, A.; Sokale, A.; Doranalli, K.; Applegate, T.J. Bacillus subtilis DSM 32315 supplementation attenuates the effects of Clostridium perfringens challenge on the growth performance and intestinal microbiota of broiler chickens. Microorganisms 2019, 7, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, A.-L.; Tan, H.-M. Effect of Bacillus subtilis PB6 (CloSTAT) on broilers infected with a pathogenic strain of Escherichia coli. J. Appl. Poult. Res. 2006, 15, 229–235. [Google Scholar] [CrossRef]
- Khan, R.; Naz, S. The applications of probiotics in poultry production. World’s Poult. Sci. J. 2013, 69, 621–632. [Google Scholar] [CrossRef]
- Awad, W.; Ghareeb, K.; Böhm, J. Effect of addition of a probiotic micro-organism to broiler diet on intestinal mucosal architecture and electrophysiological parameters. J. Anim. Physiol. Anim. Nutr. 2010, 94, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yan, Y.; Wan, P.; Chen, D.; Ding, Y.; Ran, L.; Mi, J.; Lu, L.; Zhang, Z.; Li, X. Gut microbiota modulation and anti-inflammatory properties of anthocyanins from the fruits of Lycium ruthenicum Murray in dextran sodium sulfate-induced colitis in mice. Free Radic. Biol. Med. 2019, 136, 96–108. [Google Scholar] [CrossRef]
- Zhang, H.; Pan, S.; Zhang, K.; Michiels, J.; Zeng, Q.; Ding, X.; Wang, J.; Peng, H.; Bai, J.; Xuan, Y. Impact of dietary manganese on intestinal barrier and inflammatory response in broilers challenged with Salmonella typhimurium. Microorganisms 2020, 8, 757. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, D.; Wang, H.; Sun, N.; Xin, J.; Yang, H.; Lei, L.; Khalique, A.; Rajput, D.S.; Pan, K. Analysis of miRNA expression in the ileum of broiler chickens during Bacillus licheniformis H2 supplementation against subclinical necrotic enteritis. Probiotics Antimicrob. Proteins 2021, 13, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Thangavel, G.; Kurian, H.; Mani, R.; Mukkalil, R.; Chirakkal, H. Bacillus subtilis PB6 improves intestinal health of broiler chickens challenged with Clostridium perfringens-induced necrotic enteritis. Poult. Sci. 2013, 92, 370–374. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Roos, S.; Jonsson, H.; Ahl, D.; Dicksved, J.; Lindberg, J.E.; Lundh, T. Effects of Lactobacillus johnsonii and Lactobacillus reuteri on gut barrier function and heat shock proteins in intestinal porcine epithelial cells. Physiol. Rep. 2015, 3, e12355. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Dicksved, J.; Lundh, T.; Lindberg, J.E. Heat shock proteins: Intestinal gatekeepers that are influenced by dietary components and the gut microbiota. Pathogens 2014, 3, 187–210. [Google Scholar] [CrossRef] [PubMed]
- De, A.K.; Kodys, K.M.; Yeh, B.S.; Miller-Graziano, C. Exaggerated human monocyte IL-10 concomitant to minimal TNF-α induction by heat-shock protein 27 (Hsp27) suggests Hsp27 is primarily an antiinflammatory stimulus. J. Immunol. 2000, 165, 3951–3958. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, T.M.; Leonel, A.J.; Melo, M.A.; Santos, R.R.; Cara, D.C.; Cardoso, V.N.; Correia, M.I.; Alvarez-Leite, J.I. Oral supplementation of butyrate reduces mucositis and intestinal permeability associated with 5-fluorouracil administration. Lipids 2012, 47, 669–678. [Google Scholar] [CrossRef]
- Wei, X.; Luo, L.; Chen, J. Roles of mTOR signaling in tissue regeneration. Cells 2019, 8, 1075. [Google Scholar] [CrossRef] [Green Version]
- Dolezal, T.; Krejcova, G.; Bajgar, A.; Nedbalova, P.; Strasser, P. Molecular regulations of metabolism during immune response in insects. Insect Biochem. Mol. Biol 2019, 109, 31–42. [Google Scholar] [CrossRef]
- Morhardt, T.L.; Hayashi, A.; Ochi, T.; Quirós, M.; Kitamoto, S.; Nagao-Kitamoto, H.; Kuffa, P.; Atarashi, K.; Honda, K.; Kao, J.Y. IL-10 produced by macrophages regulates epithelial integrity in the small intestine. Sci. Rep. 2019, 9, 1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagahama, M.; Kihara, A.; Kintoh, H.; Oda, M.; Sakurai, J. Involvement of tumour necrosis factor-α in Clostridium perfringensβ-toxin-induced plasma extravasation in mice. Br. J. Pharm. 2008, 153, 1296–1302. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Qu, J.-M.; Summah, H.; Zhang, J.; Zhu, Y.-G.; Jiang, H.-N. Protective effects of imipramine in murine endotoxin-induced acute lung injury. Eur. J. Pharm. 2010, 638, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wang, Y.; Li, Z.; Qamar, H.; Mehmood, K.; Zhang, L.; Liu, J.; Zhang, H.; Li, J. Probiotics isolated from yaks improves the growth performance, antioxidant activity, and cytokines related to immunity and inflammation in mice. Microb. Cell Factories 2019, 18, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimao, R.; Muroi, H.; Furukawa, K.; Toyomizu, M.; Kikusato, M. Effects of low-dose oleuropein diet supplementation on the oxidative status of skeletal muscles and plasma hormonal concentration of growing broiler chickens. Br. Poult. Sci. 2019, 60, 784–789. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci. 2009, 122, 3589–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lourdes Pérez-Chabela, M.; Yenizey, M.; Soriano-Santos, J.; Pérez-Hernández, M.A. Los probióticos y sus metabolitos en la acuicultura. Una Revisión The probiotics and their metabolites in aquaculture. A review. Hidrobiológica 2020, 30, 93–105. [Google Scholar]
- Rolfe, R.D. The role of probiotic cultures in the control of gastrointestinal health. J. Nutr. 2000, 130, 396S–402S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaenhammer, T.R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the normal gut microbiota. World J. Gastroenterol. WJG 2015, 21, 8787. [Google Scholar] [CrossRef]
- Zuo, F.; Yu, R.; Feng, X.; Chen, L.; Zeng, Z.; Khaskheli, G.B.; Ma, H.; Chen, S. Characterization and in vitro properties of potential probiotic Bifidobacterium strains isolated from breast-fed infant feces. Ann. Microbiol. 2016, 66, 1027–1037. [Google Scholar] [CrossRef]
- Apajalahti, J.; Vienola, K. Interaction between chicken intestinal microbiota and protein digestion. Anim. Feed Sci. Technol. 2016, 221, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Ohtani, K.; Hirakawa, H.; Ohshima, K.; Yamashita, A.; Shiba, T.; Ogasawara, N.; Hattori, M.; Kuhara, S.; Hayashi, H. Complete genome sequence of Clostridium perfringens, an anaerobic flesh-eater. Proc. Nat. Acad. Sci. USA 2002, 99, 996–1001. [Google Scholar] [CrossRef] [Green Version]
- Dabard, J.; Bridonneau, C.; Phillipe, C.; Anglade, P.; Mollé, D.; Nardi, M.; Ladire, M.; Girardin, H.; Marcille, F.; Gomez, A. Ruminococcin A, a new lantibiotic produced by a Ruminococcus gnavus strain isolated from human feces. Appl. Environ. Microbiol. 2001, 67, 4111–4118. [Google Scholar] [CrossRef] [Green Version]
- Stanojević-Nikolić, S.; Dimić, G.; Mojović, L.; Pejin, J.; Djukić-Vuković, A.; Kocić-Tanackov, S. Antimicrobial activity of lactic acid against pathogen and spoilage microorganisms. J. Food Process. Preserv. 2016, 40, 990–998. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Padgett, C.; Bailey, C.; Gancel, F.; Drider, D. Protective effects of novel Lactobacillaceae strains isolated from chicken Caeca against necrotic enteritis infection: In vitro and in vivo evidences. Microorganisms 2022, 10, 152. [Google Scholar] [CrossRef]
- Lima, E.T.; Andreatti Filho, R.L.; Okamoto, A.S.; Noujaim, J.C.; Barros, M.R.; Crocci, A.J. Evaluation in vitro of the antagonistic substances produced by Lactobacillus spp. isolated from chickens. Can. J. Vet. Res. 2007, 71, 103. [Google Scholar]
- Guo, S.; Liu, D.; Zhang, B.; Li, Z.; Li, Y.; Ding, B.; Guo, Y. Two Lactobacillus species inhibit the growth and α-toxin production of Clostridium perfringens and induced proinflammatory factors in chicken intestinal epithelial cells in vitro. Front. Microbiol. 2017, 8, 2081. [Google Scholar] [CrossRef] [PubMed]
- Shamshirgaran, M.A.; Golchin, M.; Mohammadi, E. Lactobacillus casei displaying Clostridium perfringens NetB antigen protects chickens against necrotic enteritis. Appl. Microbiol. Biotechnol. 2022, 106, 6441–6453. [Google Scholar] [CrossRef]
- Jumpertz, R.; Le, D.S.; Turnbraugh, P.J.; Trinidad, C.; Bogardus, C.; Gordon, J.I. Energy balance studies reveal associations between gut microbes, calorie load, and nutrient absorption in humans. Am. J. Clin. Nutr. 2011, 94, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Qu, A.; Brulc, J.M.; Wilson, M.K.; Law, B.F.; Theoret, J.R.; Joens, L.A.; Konkel, M.E.; Angly, F.; Dinsdale, E.A.; Edwards, R.A. Comparative metagenomics reveals host specific metavirulomes and horizontal gene transfer elements in the chicken cecum microbiome. PLoS ONE 2008, 3, e2945. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Lv, Z.; Li, Z.; Wang, W.; Li, G.; Guo, Y. Dietary L-arginine supplementation alleviates the intestinal injury and modulates the gut microbiota in broiler chickens challenged by Clostridium perfringens. Front. Microbiol. 2018, 9, 1716. [Google Scholar] [CrossRef] [Green Version]
- Lyons, P.; Turnbull, J.; Dawson, K.A.; Crumlish, M. Phylogenetic and functional characterization of the distal intestinal microbiome of rainbow trout Oncorhynchus mykiss from both farm and aquarium settings. J. Appl. Microbiol. 2017, 122, 347–363. [Google Scholar] [CrossRef] [Green Version]
- Davenport, M.; Poles, J.; Leung, J.M.; Wolff, M.J.; Abidi, W.M.; Ullman, T.; Mayer, L.; Cho, I.; Loke, P.N. Metabolic alterations to the mucosal microbiota in inflammatory bowel disease. Inflamm. Bowel Dis. 2014, 20, 723–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, N.; Eom, T.; Gupta, S.K.; Jeong, S.-Y.; Jeong, D.-Y.; Kim, Y.S.; Lee, J.-H.; Sadowsky, M.J.; Unno, T. Genes and gut bacteria involved in luminal butyrate reduction caused by diet and loperamide. Genes 2017, 8, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineda-Quiroga, C.; Borda-Molina, D.; Chaves-Moreno, D.; Ruiz, R.; Atxaerandio, R.; Camarinha-Silva, A.; García-Rodríguez, A. Microbial and functional profile of the ceca from laying hens affected by feeding prebiotics, probiotics, and synbiotics. Microorganisms 2019, 7, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| S/N | Probiotic Strain | GP | VM | IMF | INF | TJs | ANT | GM | Reference |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Bacillus coagulans | NS | NE | P | P | NE | P | P | [15] |
| 2 | Lactobacillus plantarum ZLP001 | NE | NE | P | P | P | NE | P | [24] |
| 3 | Lactobacillus plantarum A37 and L. plantarum MIII | P | P | P | P | P | P | P | [36] |
| 4 | Bacillus subtilis and Bacillus licheniformis | P | P | P | NE | NE | P | NE | [37] |
| 5 | Lactobacillus LP184 and Yeast SC167 | P | P | P | NE | NE | P | P | [38] |
| 6 | Bacillus amyloliquefaciens TL106 | P | P | P | P | P | NE | P | [39] |
| 7 | Lactobacillus paracaesi | NS | P | P | P | P | P | P | [40] |
| 8 | Multi strain | P | P | P | NE | P | P | NE | [41] |
| 9 | Bacillus amyloliquefaciens | P | NE | NE | P | P | NE | NE | [42] |
| 10 | Bacillus subtilis | P | P | P | P | P | P | NE | [43] |
| 11 | Bacillus methylotrophicus SY200 | P | P | NE | NE | P | P | P | [44] |
| 12 | Bacillus subtilis and Bacillus licheniformis | P | NE | P | P | NE | P | P | [45] |
| 13 | Lactococcus and Lactobacillus plantarum | P | P | P | NE | NE | NE | P | [46] |
| 14 | Bacillus subtilis fmbj | P | P | P | NE | NE | P | NE | [47] |
| 15 | Bacillus subtilis | P | P | P | NE | P | NE | P | [54] |
| 16 | Bacillus subtilis (B. subtilis) BYS2 | P | P | P | P | NE | NE | NE | [55] |
| 17 | Clostridium butyricum | P | P | NE | P | P | NE | P | [56] |
| 18 | Bacillus spp. and Debaryomyces spp. | P | P | NS | NE | NE | P | P | [57] |
| 19 | Saccharomyces cerevisiae hydrolysate | P | P | NE | P | P | NE | P | [58] |
| 20 | Bacillus subtilis DSM 32315 | P | P | NS | NS | NE | NE | P | [59] |
| 21 | Bacillus subtilis ATCC19659 | P | P | P | NE | NE | P | P | [60] |
| 22 | Lactobacillus and Yeast | P | P | NE | NE | NE | P | NS | [61] |
| 23 | Bacillus amyloliquefaciens LFB112 | P | NE | P | NE | NE | NE | NE | [62] |
| 24 | Bacillus spp. | P | P | NE | NE | NE | NE | P | [63] |
| 25 | Bacillus amyloliquefaciens CECT 5940 | P | NE | P | P | NE | P | NE | [64] |
| 26 | Lactobacillus | P | NE | P | NS | NE | NS | P | [65] |
| 27 | Bacillus subtilis | P | P | NE | NE | NE | NE | NE | [66] |
| 28 | Bacillus amyloliquefaciens SCO6 | NS | NE | P | P | P | P | P | [67] |
| 29 | Lactobacillus | NS | NE | P | NE | P | NE | NE | [68] |
| 30 | Lactobacillus reuteri | NS | NE | NE | NE | NE | NE | P | [69] |
| Probiotic Strain | Response to NE Challenge | Response to Dietary Probiotics | Ref | |||||
|---|---|---|---|---|---|---|---|---|
| S/N | Lesion Site | Mortality Rate | GP | Lesion Site | Mortality Rate | GP | ||
| 1 | B. amyloliquefaciens CECT 5940 | Jejunum | Comparable to positive control | Reduced growth performance | No effect | Not significant | Improved performance | [11] |
| 2 | B. licheniformis H2 | NE | High mortality rate | Reduced growth performance | NE | Reduced mortality rate | Enhanced weight gain and reduced FCR | [12] |
| 3 | Multi strains of Lactobacillus | Small intestine | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [13] |
| 4 | B. subtilis DSM29784 | Comparable to control | High mortality rate | Reduced growth performance | NE | Reduced mortality rate | Enhanced weight gain | [17] |
| 5 | B. subtilis | Jejunum and ileum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Reduced FCR | [18] |
| 6 | B. licheniformis | Duodenum, jejunum and ileum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Reduced FCR | [19] |
| 7 | Bacillus | Ileum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Improved weight | [21] |
| 8 | B. licheniformis | Duodenum, jejunum and ileum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Reduced FCR | [23] |
| 9 | B. coagulans | Duodenum and jejunum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [25] |
| 10 | Clostridium. butyricum | Intestine | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Reduced FCR | [26] |
| 11 | L. plantarum 1.2567 | Small intestine | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Improved weight gain | [30] |
| 12 | B. amyloliquefaciens H57. | Small intestine | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [31] |
| 13 | B. subtilis | Duodenum, jejunum and ileum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Reduced FCR | [32] |
| 14 | L. fermentum, B. coagulans | Duodenum and Ileum | High mortality rate | Reduced growth performance | no effect | Reduced mortality rate | Reduced FCR | [33] |
| 15 | B. subtilis DSM 32315. | NE | High mortality rate | Reduced growth performance | NE | Reduced mortality rate | [52] | |
| 16 | Multi strain | Duodenum, jejunum and ileum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Reduced FCR | [53] |
| 17 | Lactobacillus johnsonii BS15 | NE | High mortality rate | Reduced growth performance | NE | Reduced mortality rate | Enhanced weight gain and reduced FCR | [70] |
| 18 | Primlac: multi strain | Duodenum and jejunum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Reduced FCR | [94] |
| 19 | B. subtilis DSM29784 | Jejunum | High mortality rate | Reduced growth performance | Comparable to control | Reduced mortality rate | Enhanced weight gain and reduced FCR | [95] |
| 20 | B. licheniformis H2 | Ileum | High mortality rate | Reduced growth performance | NO | Reduced mortality rate | Enhanced weight gain and reduced FCR | [109] |
| 21 | B. amyloliquefaciens BLCC1-0238 | Duodenum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [110] |
| 22 | Multi strain | Duodenum, jejunum and ileum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [111] |
| 23 | E. faecium | Jejunum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [112] |
| 24 | B. subtilis DSM 32315 | Footpad | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [113] |
| 25 | L. johnsonii BS15 | NE | High mortality rate | Reduced growth performance | NE | Reduced mortality rate | Enhanced weight gain and reduced FCR | [114] |
| 26 | C. butyricum MIYAIRI 588 | Small intestine | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [115] |
| 27 | L. johnsonii BS15, Bacillus | NE | High mortality rate | Reduced growth performance | NE | Reduced mortality rate | Enhanced weight gain and reduced FCR | [116] |
| 28 | B. subtilis DSM 32315 | Mid intestine | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [117] |
| 29 | Bacteriophage | Jejunum | High mortality rate | Reduced growth performance | Reduced lesion scores | Zero mortality in high-dose PRO group | Improved weight gain | [118] |
| 30 | Bacillus. Surfactin (fermented product) | Duodenum, jejunum and ileum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [119] |
| 31 | B. licheniformis | Duodenum and jejunum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [120] |
| 32 | Butyricicoccus pullicaecorum 25-3T | Duodenum and jejunum | High mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Reduced FCR | [121] |
| 33 | B. subtilis DSM29784 | Ileum | Low mortality rate | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | No effect | [122] |
| 34 | L. johnsonii. LB 15 | NE | High mortality rate | Reduced growth performance | NE | Reduced mortality rate | Enhanced weight gain and reduced FCR | [123] |
| 35 | B. amyloliquefaciens CECT 5940 | Footpad | Comparable to positive control | Reduced growth performance | Reduced lesion scores | Reduced mortality rate | Enhanced weight gain and reduced FCR | [124] |
| S/N | Response to NE Challenge | Probiotic Strain | Response to Dietary Probiotics | Ref |
|---|---|---|---|---|
| 1 | Decreased the expression of CLDN-1, CLDN-3, ZO-1 and ZO-2 | B. licheniformis H2 | Increased the expression of these proteins | [12] |
| 2 | Reduced the expression of IGF-1 and EGF in the gut | Multi probiotic strain. B. subtilis | Increased IGF-1 and EGF in the jejunum and ileum | [18] |
| 3 | Increased expression of CLDN3, | Multi strain | Decreased CLDN3; increased CLDN3 and Muc-2 on day 42 | [19] |
| 4 | Reduced the expression of CLDN-3 | B. licheniformis | Increased the expression of CLDN-3 | [23] |
| 5 | Reduced mRNA expression of Muc2; reduced IAP activity | B. coagulans | No effect on mucin expression; increased IAP activity in the jejunum | [25] |
| 6 | Decreased CLDN-1 but had no influence on OCLDN and CLDN-2 | L. fermentum, B. coagulans | Increased CLDN-3 | [26] |
| 7 | Reduced mRNA expression of CLDN-3, CLDN-1 and ZO-2 | Primlac: multi strain probiotics | Increased expression of these proteins | [94] |
| 8 | Reduced expression of CLDN-1 and OCLDN transcripts; increased Muc-2 | B. subtilis DSM29784 | No effect on expression of claudins | [95] |
| 9 | Increased level of serum DAO and reduced CLDN-3 and MUC-2 | B. amyloliquefaciens BLCC1-0238 | Reduced level of serum DAO and D-lactic acid; increased OCLN, ZO-1 and MUC2 | [110] |
| 10 | Reduced CLDN-3 and ZO-1; increased MLCK mRNA expression | E. faecium | Increased expression of CLDN-1 | [112] |
| 11 | Reduced JAM2 | B. licheniformis | Increased JAM2 | [122] |
| S/N | Response to NE Challenge | Probiotic Strains | Response to Dietary Probiotics | Ref |
|---|---|---|---|---|
| 1 | Hyperemia of lamina propria and necrotic intestinal epithelial cells | B. licheniformis | Restored it | [12] |
| 2 | Reduced V/C ratio | Multi strains of Lactobacillus | Increased V/C ratio | [13] |
| 3 | Decreased goblet number and no effect on jejunal VH, CD and V/C ratio | Multi strain | No significant effect on VH, CD and V/CD | [19] |
| 4 | Reduced villi length, necrosis of intestinal villi and hyperplasia | B. subtilis | Restored villi morphology | [22] |
| 5 | Reduced VH/CD, goblet cell number, VH and IAP activity | B. coagulans | Restored the increased negative effect | [25] |
| 6 | Damaged ileal tissue, loss of villi architecture, mucosal damage and decreased density and length of villi enterocytes | L. plantarum 1.2567 powder | Restored it and reduced NO and MPO activity in the ileum mucosa | [30] |
| 7 | Irregular villi, oedma, separation from basement membrane and goblet cell metaplasia | B. subtilis | Restored the villi architecture, and reduced oedema | [32] |
| 8 | Irregular villi structure and shorter jejunal villi length | B. subtilis DSM29784 | Restored it | [95] |
| 9 | Reduced villi length and VH/CD | B. licheniformis | Restored it | [109] |
| 10 | Irregular villi shape, swelling of villus tip and reduced villi length | B. amyloliquefaciens, BLCC1-0238 | Restored it | [110] |
| 11 | Increased intestinal histopathology | E. faecium | Increased PCNA-positive cells and reduced TUNEL -positive cells | [112] |
| 12 | Damaged villi, shedding of epithelial cells and congested lamina propria | L. johnsonii BS15 | Restored it | [114] |
| 13 | Reduced villi length | L. johnsonii, Bacillus | Restored it | [116] |
| 14 | Increased cecal mucosal thickness | B. subtilis DSM 32315 | Decreased CD, TLI and EI; increased V/C ratio | [117] |
| 15 | Disruption in villi crypt and lamina propria; reduced villi length | Bacteriophage | Restored it | [118] |
| 16 | Reduced villi length | Bacillus. Surfactin (fermented product) | Restored villi morphology | [119] |
| 17 | Reduced villi length | B. licheniformis fermented product | Restored villi morphology | [120] |
| 18 | Reduced villi length and increased duodenal crypt depth | B. licheniformis | Increased villi length and reduced crypt depth | [136] |
| 19 | Decreased jejunal height | Multi strain | Improved jejunal V/C ratio | [137] |
| Response to NE Challenge | Response to Dietary Probiotics | |||||
|---|---|---|---|---|---|---|
| S/N | Intestinal Immunity | Intestinal Inflammation | Probiotic Strains | Intestinal Immunity | Intestinal Inflammation | Ref |
| 1 | Reduced expression of IgA and IgG; increased PGC-1a expression | NE | B. licheniformis H2 | Increased IgA and IgG; reduced expression of PGC-1a | NE | [12] |
| 2 | Decreased immunoglobulins and T helper: Th cells in the cecal tonsils | Increased IL-1B | Multi strains of Lactobacillus | Increased immunoglobulins and T cells. Reduced sIgA | Increased IFN-γ, IL-13 and IL-2; reduced IL-1B, IL-12p35, IL-17 and TGF-B | [13] |
| 3 | Decreased jejunal mRNA TRIF and NF-KB | Changes in IL-1β, IL-10, IL-17 and TNF-α | B. licheniformis | Increased jejunal mRNA TRIF and NF-KB; no effect on TLR2 or TLR4 | No effect on IL-1β, IL-10, IL-17 or TNF-α; increased GFs and HSP proteins | [19] |
| 4 | Increased FITC and intestinal IgA | NE | B. licheniformis H2 | Increased the contents | NE | [21] |
| 5 | Increased sIgA | Increased IFN-γ | B. licheniformis | Reduced sIgA | Reduced IFN-γ, IL-10 and IL-17 | [23] |
| 6 | Reduced sIgA, TLR2, TLR4 and TNFSF15 Fowlcidin gene | Increased IFN-γ. Decreased TLR2, TLR4 and TNFSF15 gene | B. coagulans | Increased sIgA; no effect on TLRsl only increased fowlcidin-2 | Reduced IFN-γ; no effect on others | [25] |
| 7 | Reduced TLR2 | No effect on TNF or TLR2; increased IL-17 | C. butyricum | No effect on intestinal IgA | Increased TNF-α, IL-10, reduced IL-17A | [26] |
| 8 | NE | Increased (chTNF-α) and IL-1β in the ileum mucosa | L. plantarum 1.2567 | NE | Reduced (chTNF-α) and IL-1β | [30] |
| 9 | Reduced populationof CD3+ | Increased jejunal IL-1β and TGF-β4 by 28. Both increased | L. fermentum, B. coagulans | Increased CD3 | Increased and reduced IL-1β, INF-γ, IL-13,1L-17 and TGF-B. | [33] |
| 10 | NE | Increased IL-6, TNF-a and IFN-γ | B. subtilis DSM 32315. | NE | Reduced IL-6, TNF-α and IFN-γ and increased IL-10 and SIgA | [52] |
| 11 | Reduced ileum IgA and IgG, sIgA and content. Increased MMP-2 | Increased IFN-γ and IL-10 | Lactobacillus johnsonii BS15 | Increased the IgA, IgG and sIgA content; reduced MMP-2 | Reduced IFN-γ and IL-10; increased Nrf-2 and IL-8. | [70] |
| 12 | NE | Reduced IL-10 and IL-17 | Primlac:multi strain | NE | Increased IL-10 and IL-17 | [94] |
| 13 | Reduced serum sIgA and IgG | Increased IL-1β, TNF-α, INF-γ and IL-6 | B. subtilis DSM29784 | No effect on sIgA | Reduced IFN-γ and TNF- α | [95] |
| 14 | Reduced sIgA | Increased IFN-γ, IL-10 and IL-6 | B. amyloliquefaciens BLCC1-0238 | Increased sIgA | Decreased IFN-γ, IL-10 and IL-6 | [110] |
| 15 | NE | Reduced TLR-2, IL-1β, IL-4, IFN-γ, iNOS and IL-10 | E. faecium | Increased MYD88, NFK-B, IL-1β, IL-4 and iNOS | [112] | |
| 16 | Reduced serum IgG and IgA, CD3+, CD4+ and lymphocyte percentage | Reduced IL-2, IL-4 and IFN-γ | L. johnsonii BS15 | Increased the IgG and IgA and CD+ cells | Increased IFN-γ and IL-2 | [114] |
| 17 | Reduced serum IgG and IgM | No effect on IL-6, TNF-α or IFN-γ | L. johnsonii BS15, Bacillus | Increased serum IgG and IgM | No effect on IL-6, TNF-α or IFN-γ but reduced IL-8 | [116] |
| 18 | Reduced TLR21 | Reduced INF-γ, IL-12 and TGF-B4 | B. subtilis 29,784 | Increased TLR21 after infection and TLR 5 | Increased INF-γ, IL-12 and TGF-B4 | [122] |
| S/N | Response to NE Challenge | Probiotic Strain | Response to Dietary Probiotics | Ref |
|---|---|---|---|---|
| 1 | Decreased Actinobacteria, Lactobacillacae and Firmicutes (Clostridia) | Multi strains of Lactobacillus | Increased Actinobacteria, Lactobacillacae and Firmicutes (Clostridia) | [13] |
| 2 | Decreased Lachnospiraceae_UCG_010, Clostridiales_vadinBB60 and Ruminococcaceae_NK4A214_ | B. licheniformis | Increased Lachnospiraceae_UCG_010, Clostridiales_vadinBB60 and Ruminococcaceae_NK4A214 | [19] |
| 3 | Reduced Firmicutes, increased genera; Turicibacter, Streptococcus, Enterococcus and Clostridium | B. licheniformis H2 | Reduced Proteobacteria, increased Lactobacillus and Bacillus | [21] |
| 4 | Increased Clostridium sensu stricto-1 and reduced Lactobacillus | B. licheniformis | Reduced Clostridium sensu stricto 1 and Escherichia-Shigella; increased Lactobacillus | [23] |
| 5 | Reduced cecal Lactobacillus and Bifidobacterium; increased cecal coliform | B. coagulans | Reversed the trend | [25] |
| 6 | Increased Candidatus Arthromitus unclassified Brachybacterium and decreased Lactobacillus sp. KC45b | C. butyricum | Probiotics reversed all; increased Weissella thailandensis and Pediococcus acidilactici | [26] |
| 7 | Increased Dorea, Bacteroides, Eubacterium, Caldanaerocella and Enterococcus | B. subtilis | Decreased Dorea, Ruminococcus and Proteobacteria | [32] |
| 8 | Increased Romboutsia, f_Lachnospiraceae and Ruminococcus_torques group, and decreased Lactobacillus. Lower ileal Bacteriodetes and cecal Proteobacteria on day 28 | L. fermentum, B. coagulans | Probiotics reversed it; decreased Faecalibacterium spp. and f_Peptostreptococcaceae | [33] |
| 9 | Increased proliferation of CP; reduced L. salivarius and B. fidobacterium | Bacillus subtilis DSM 32315. | Reduced proliferation of CP; increased L. Salivarius and B. fidobacterium | [52] |
| 10 | Increased Prevotellacea, Muribaculacea, Rominiclostridium 9, Oscillibacter, RuminococcaceaeUCG_014, ASF356, Clostridium sensu stricto 1, | Multi strain | Decreased the abundance of the pathogens; increased ileum Firmicutes and Lactobacillus | [53] |
| 11 | Increased Enterococcus, Escherichia/Shigella, Barnesiella, Desulfovibrio and Campylobacter; reduced Lactobacillus and Bacteriodes | Primlac: multi strain probiotics | Reduced Enterococcus, Escherichia/Shigella, Barnesiella, Desulfovibrio and Campylobacter; increased Lactobacillus and Bacteriodes | [94] |
| 12 | Reduced Ruminococcaceae and Bifidobacterium | B. subtilis DSM29784 | Increased Ruminococcaceae and Bifidobacterium, | [95] |
| 13 | Reduced Lactobacillus | E. faecium | Reversed the negative effect | [112] |
| 14 | Increased lachnopiraceae and Ruminococcaceae | B. subtilis DSM 32315 | Increased L. johnsonii and Salivarius; reduced CP alpha toxin | [113] |
| 15 | Increased CP coliforms in the ileum and cecum | Bacteriophage | Reduced CP coliforms in the ileum and cecum | [118] |
| 16 | Increase in Escherichia coli, Staphylococcus aureus, Salmonella typhimurium and C. perfringens. | Bacillus. Surfactin (fermented product) | In vitro reduction in Escherichia coli, Staphylococcus aureus, Salmonella typhimurium and C. perfringens | [119] |
| 17 | NE reduced Faecalibacterium | B. subtilis 29784 | Increased Butyricicoccus and Faecalibacterium genera | [122] |
| 18 | Reduced Bacteriodetes, C. cluster IV and C. cluster XIVa, Lactobacillus, increased Streptococcus spp. Enterobacteriaceae | Lactobacillus johnsonii. LB 15 | Increased Lactobacillus, C. cluster IV and C. cluster XIVa, reduced Streptococcus and Enterobacteriaceae | [123] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obianwuna, U.E.; Agbai Kalu, N.; Wang, J.; Zhang, H.; Qi, G.; Qiu, K.; Wu, S. Recent Trends on Mitigative Effect of Probiotics on Oxidative-Stress-Induced Gut Dysfunction in Broilers under Necrotic Enteritis Challenge: A Review. Antioxidants 2023, 12, 911. https://doi.org/10.3390/antiox12040911
Obianwuna UE, Agbai Kalu N, Wang J, Zhang H, Qi G, Qiu K, Wu S. Recent Trends on Mitigative Effect of Probiotics on Oxidative-Stress-Induced Gut Dysfunction in Broilers under Necrotic Enteritis Challenge: A Review. Antioxidants. 2023; 12(4):911. https://doi.org/10.3390/antiox12040911
Chicago/Turabian StyleObianwuna, Uchechukwu Edna, Nenna Agbai Kalu, Jing Wang, Haijun Zhang, Guanghai Qi, Kai Qiu, and Shugeng Wu. 2023. "Recent Trends on Mitigative Effect of Probiotics on Oxidative-Stress-Induced Gut Dysfunction in Broilers under Necrotic Enteritis Challenge: A Review" Antioxidants 12, no. 4: 911. https://doi.org/10.3390/antiox12040911