
An Arylbenzofuran, Stilbene Dimers, and Prenylated Diels–Alder Adducts as Potent Diabetic Inhibitors from Morus bombycis Leaves
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Chemicals and Reagents
2.3. Plant Material
2.4. Extraction, Fractionation, and Isolation
2.5. UPLC-QToF/ESI-MS Analysis
2.6. Determination of Total Phenolic Content (TPC) and Total Flavonoids Content (TFC)
2.7. Assay for Scavenging Activity against ABTS Radical and DPPH Radical
2.8. In Vitro Assay for ONOO− Scavenging Activity
2.9. In Vitro Assay for Inhibitory Activity of α-Glucosidase and PTP1B Enzyme
2.10. Kinetic Parameters of Isolated Compounds for Inhibition of α-Glucosidase and PTP1B Using Lineweaver–Burk and Dixon Plots
2.11. In Silico Molecular Docking Analysis for α-Glucosidase and PTP1B Inhibition
2.12. In Vitro Assay for Inhibitory Activity of HRAR and AGEs Formation
2.13. Statistics
3. Results
3.1. Phytochemical and Bioactivity Analysis of Morus Species
3.1.1. Preliminary Experiment of Three Dominant Morus Species
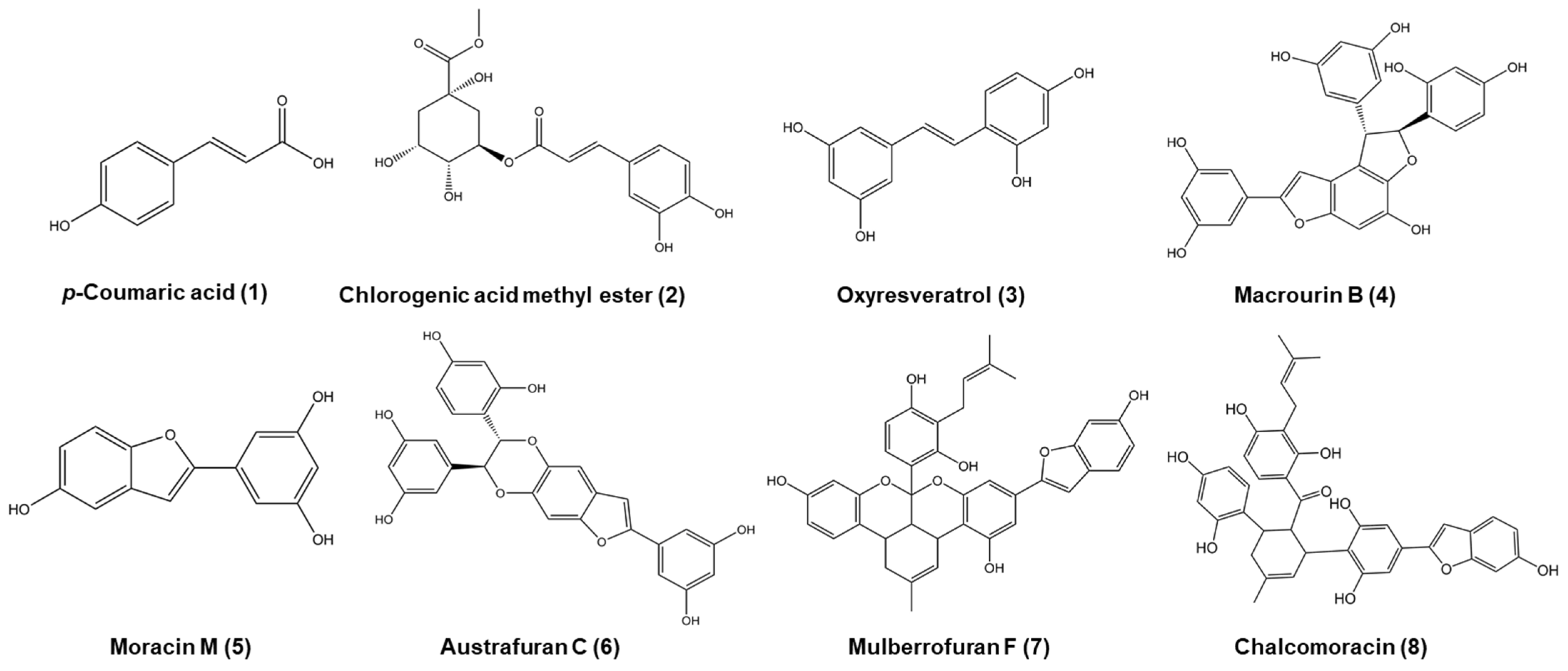
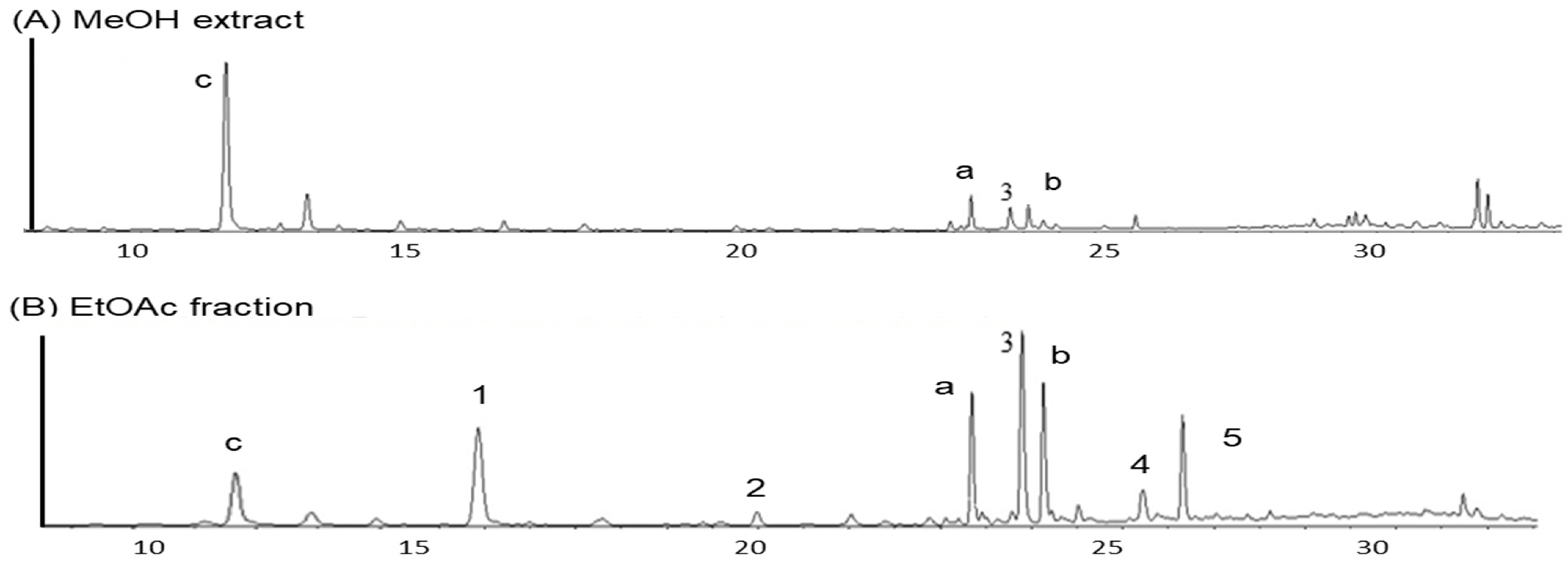
3.1.2. Phytochemical Analysis of the Morus bombycis Leaves
3.1.3. Antioxidant and Anti-Diabetic Activities of the Leaves of Morus bombycis
3.2. Evaluation of Bioactivities of Compounds Derived from the Leaves of Morus bombycis
3.2.1. Antioxidant, Anti-Diabetic, and Anti-Diabetic Complication Activities of Compounds
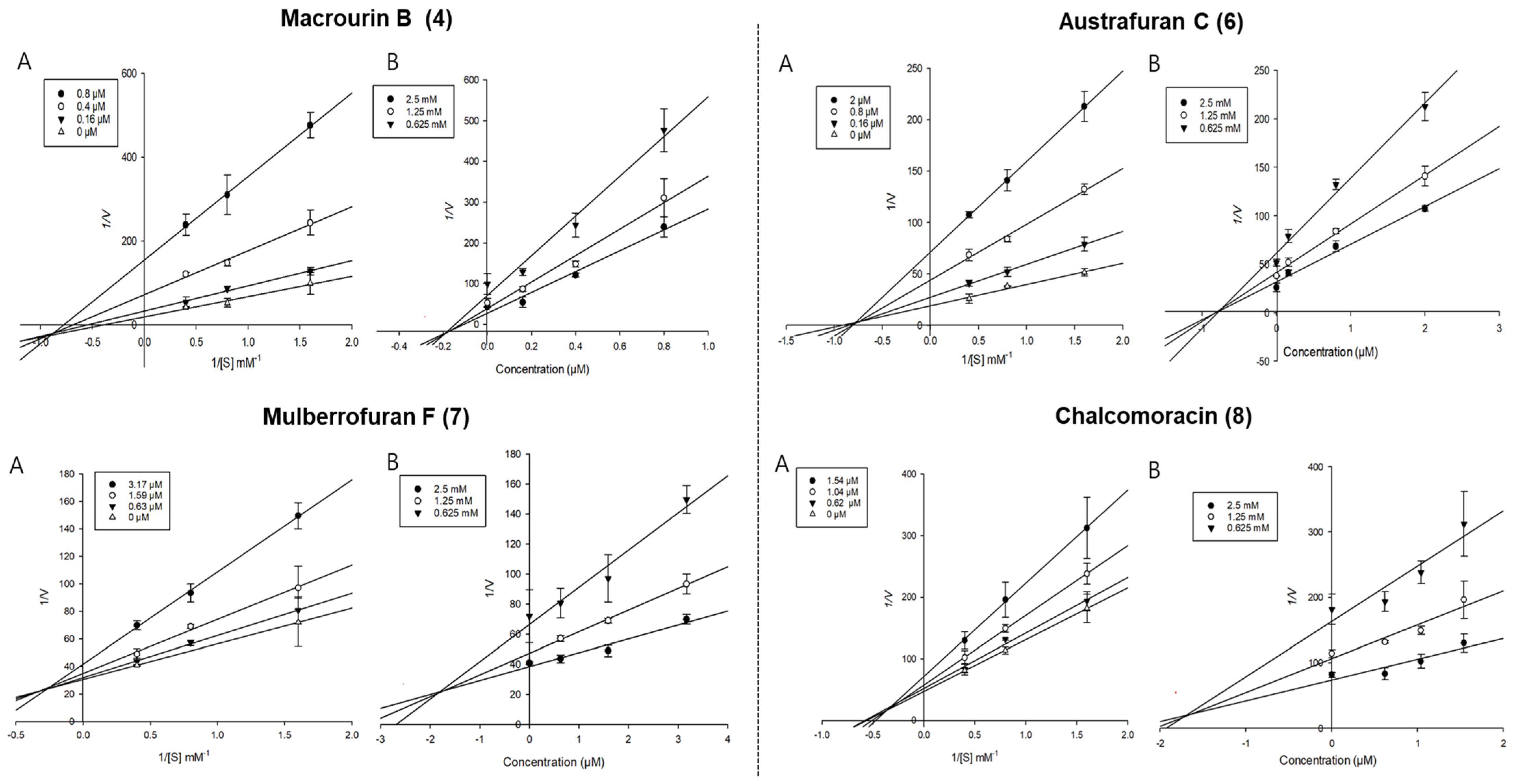
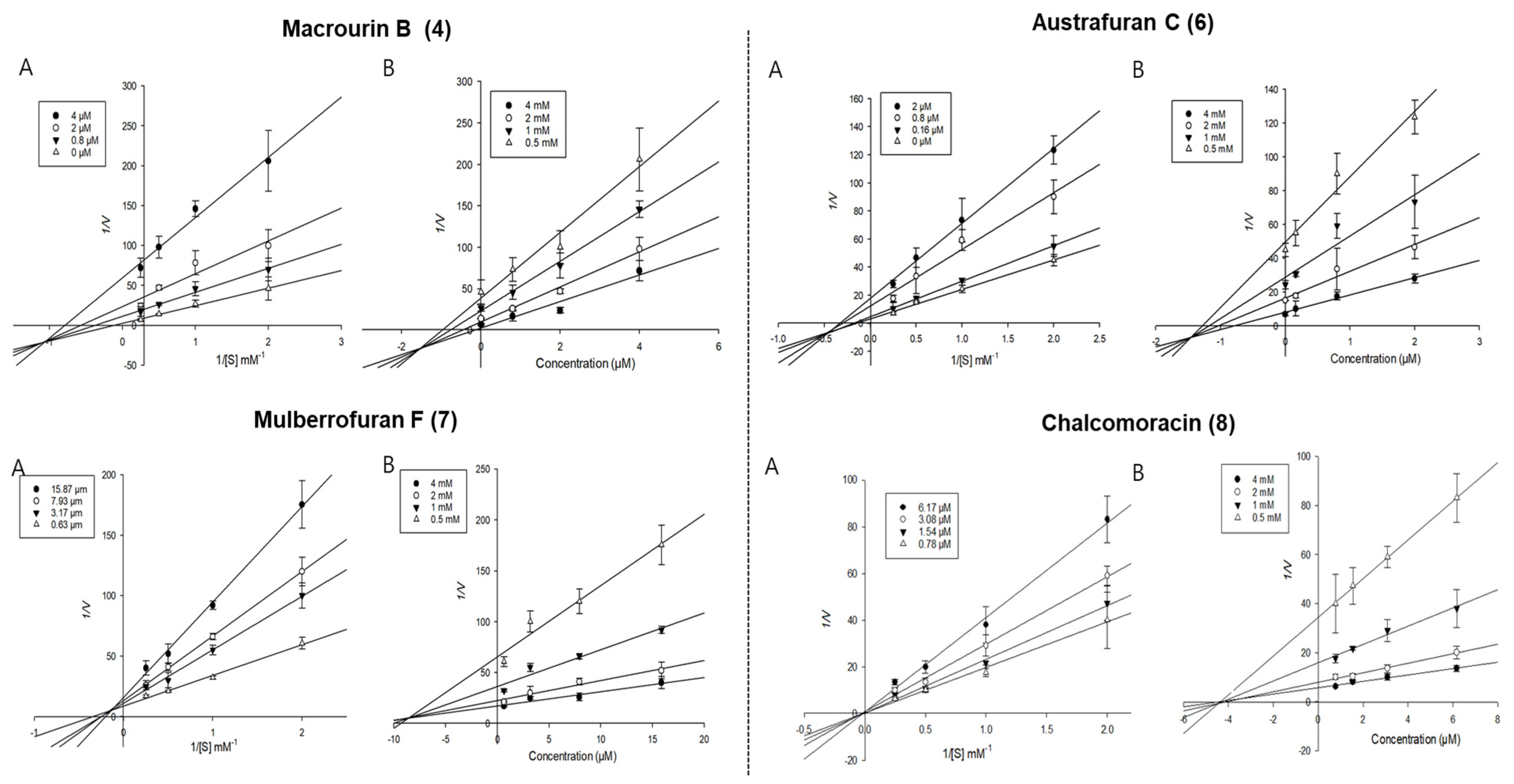
3.2.2. Enzyme Kinetic Study of Isolated Compounds Derived from Morus bombycis Leaves
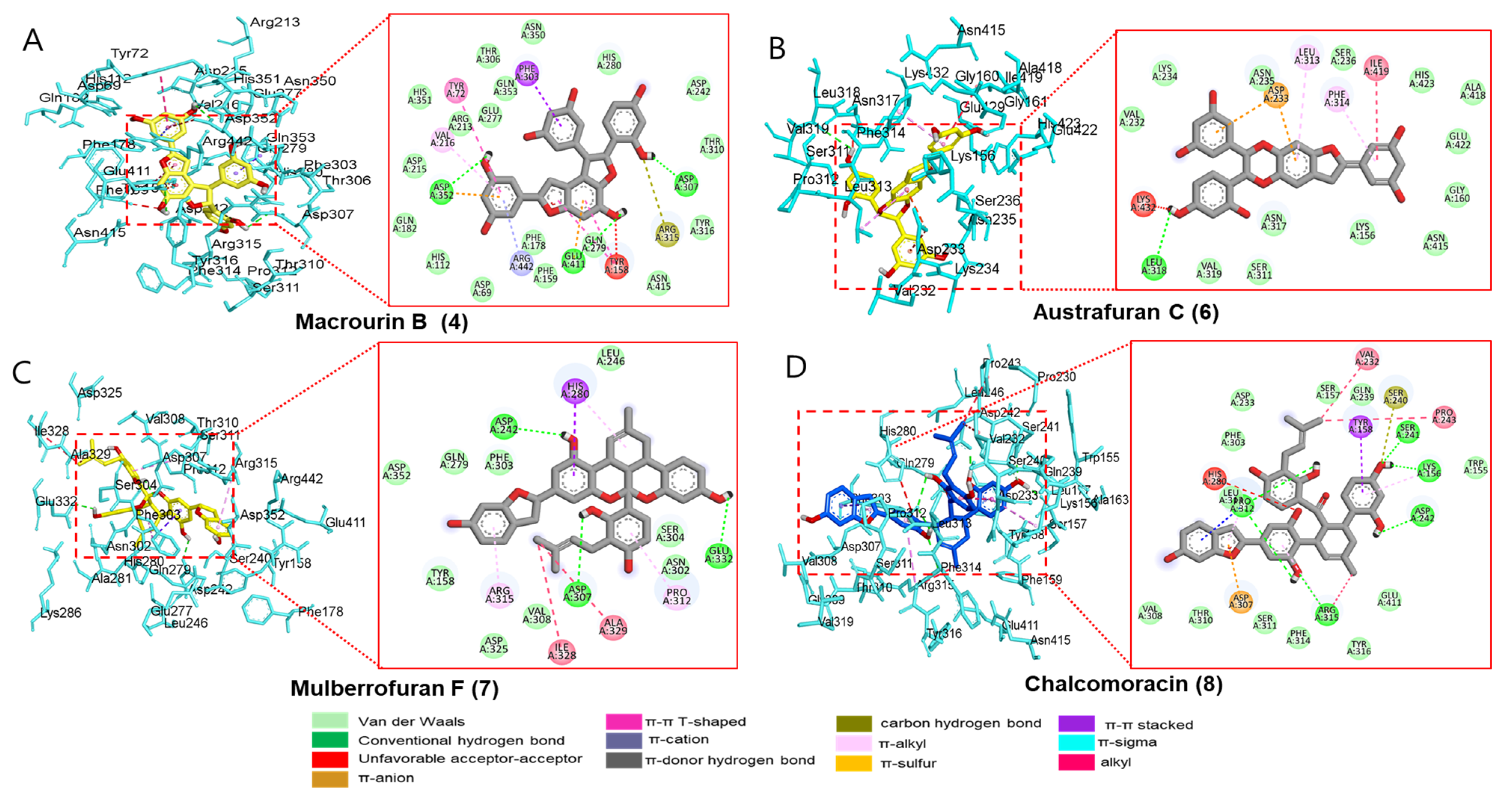
3.2.3. Docking Interaction between Compounds and Key Binding Ligands of α-Glucosidase
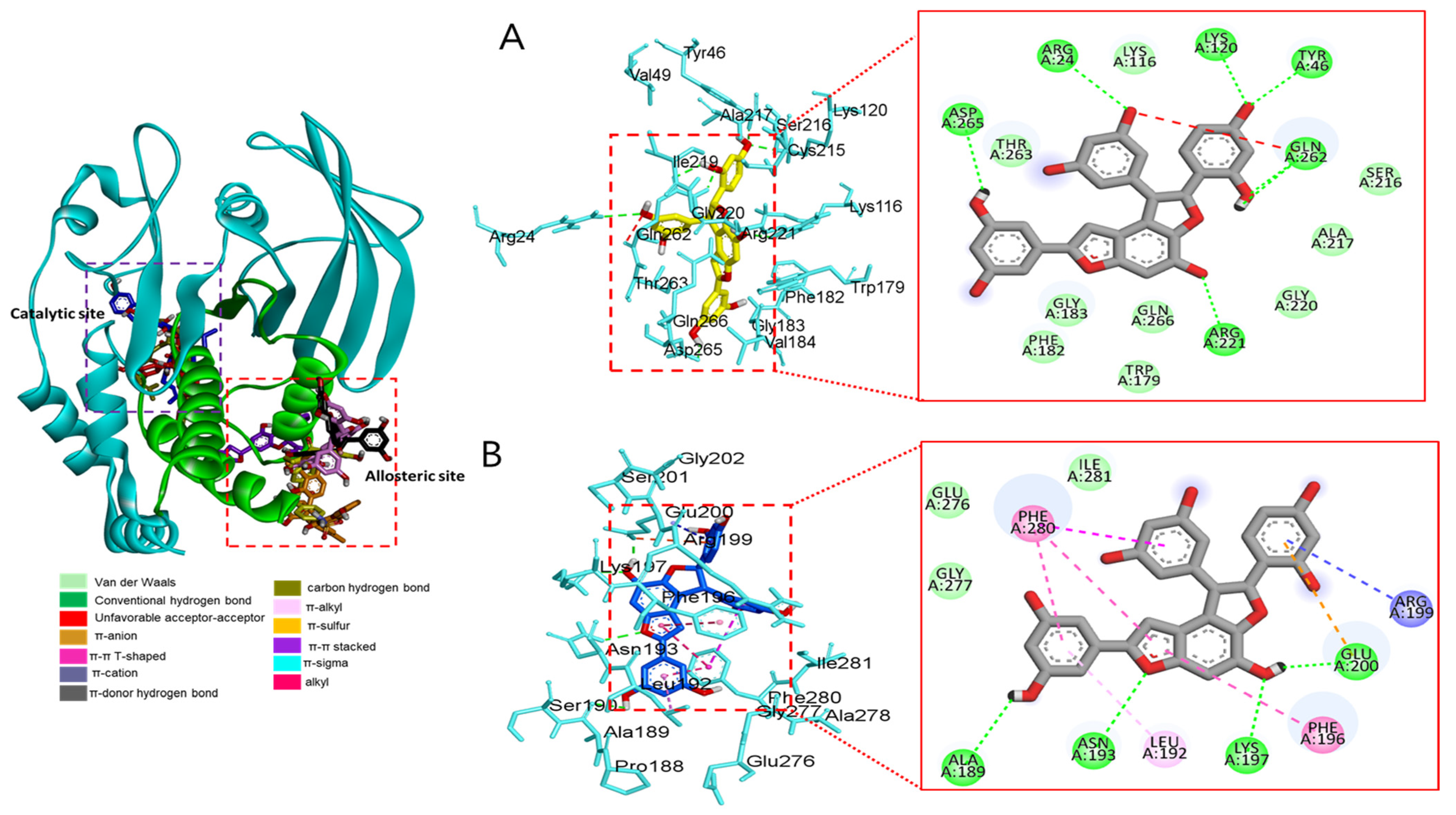
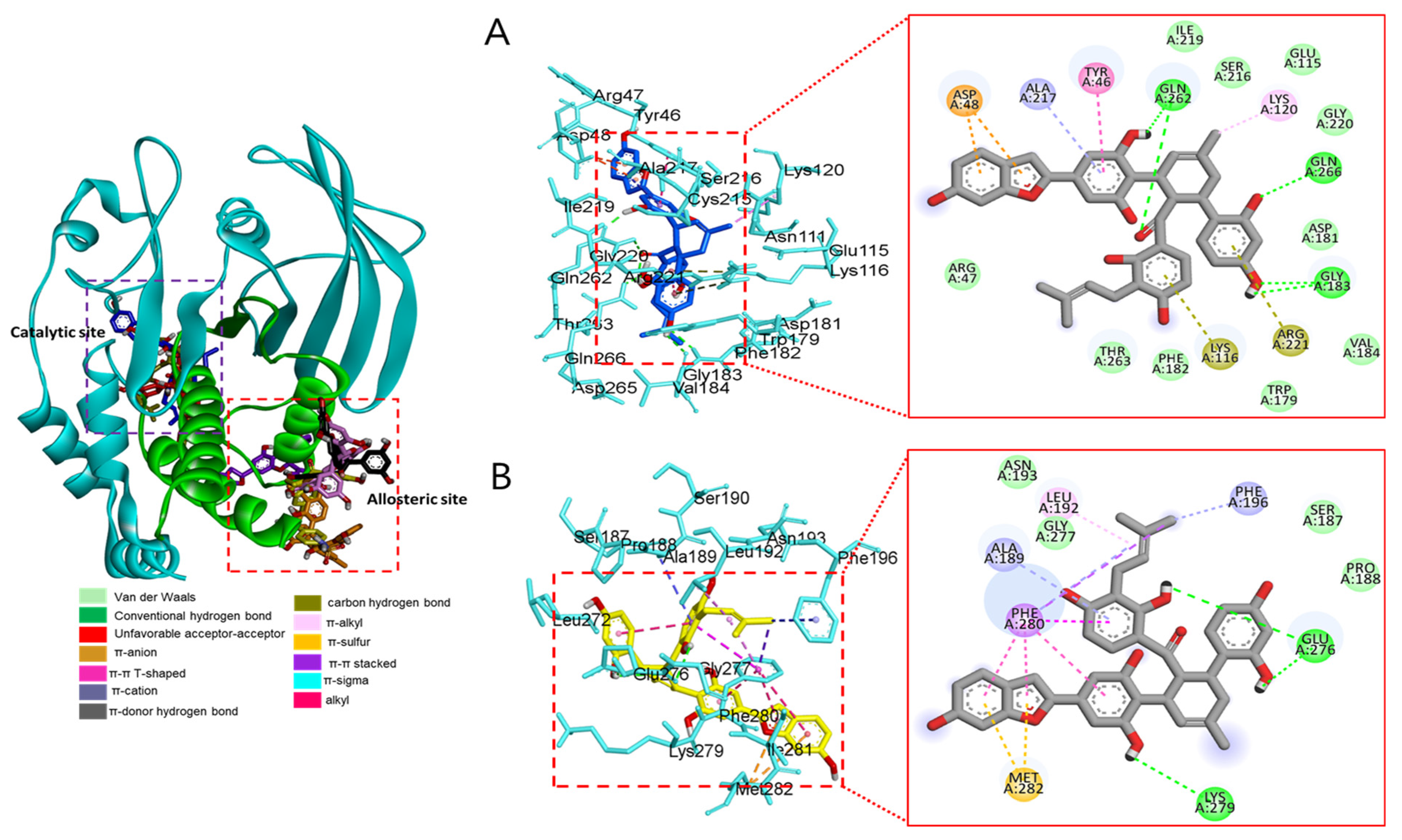
3.2.4. Docking Interaction between Compounds and Key Binding Ligands of PTP1B
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.A.B.; Hashim, M.J.; King, J.K.; Govender, R.D.; Mustafa, H.; Al Kaabi, J. Epidemiology of type 2 diabetes—Global burden of disease and forecasted trends. J. Epidemiol. Glob. Health 2020, 10, 107–111. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association Professional Practice Committee. Prevention or delay of type 2 diabetes and associated comorbidities: Standards of Medical Care in Diabetes-2022. Diabetes Care 2022, 45 (Suppl. S1), S39–S45. [Google Scholar] [CrossRef]
- Ma, Y.M.; Tao, R.Y.; Liu, Q.; Li, J.; Tian, J.Y.; Zhang, X.L.; Xiao, Z.Y.; Ye, F. PTP1B inhibitor improves both insulin resistance and lipid abnormalities in vivo and in vitro. Mol. Cell Biochem. 2011, 357, 65–72. [Google Scholar] [CrossRef]
- Dirir, A.M.; Daou, M.; Yousef, A.F.; Yousef, L.F. A review of alpha-glucosidase inhibitors from plants as potential candidates for the treatment of type-2 diabetes. Phytochem. Rev. 2021, 21, 1049–1079. [Google Scholar] [CrossRef]
- Blahova, J.; Martiniakova, M.; Babikova, M.; Kovacova, V.; Mondockova, V.; Omelka, R. Pharmaceutical drugs and natural therapeutic products for the treatment of type 2 diabetes mellitus. Pharmaceuticals 2021, 14, 806. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, Z.Y. PTP1B as a drug target: Recent developments in PTP1B inhibitor discovery. Drug Discov. Today 2007, 12, 373–381. [Google Scholar] [CrossRef]
- Liu, R.; Mathieu, C.; Berthelet, J.; Zhang, W.; Dupret, J.M.; Rodrigues Lima, F. Human protein tyrosine phosphatase 1B (PTP1B): From structure to clinical inhibitor perspectives. Int. J. Mol. Sci. 2022, 23, 7027. [Google Scholar] [CrossRef]
- Ottanà, R.; Paoli, P.; Cappiello, M.; Nguyen, T.N.; Adornato, I.; Del Corso, A.; Genovese, M.; Nesi, I.; Moschini, R.; Naß, A.; et al. In search for multi-target ligands as potential agents for diabetes mellitus and its complications-A structure-activity relationship study on inhibitors of aldose reductase and protein tyrosine phosphatase 1B. Molecules 2021, 26, 330. [Google Scholar] [CrossRef]
- Kumar, S.; Mittal, A.; Babu, D.; Mittal, A. Herbal medicines for diabetes management and its secondary complications. Curr. Diabetes Rev. 2021, 17, 437–456. [Google Scholar] [CrossRef] [PubMed]
- Safi, S.Z.; Qvist, R.; Kumar, S.; Batumalaie, K.; Ismail, I.S. Molecular mechanisms of diabetic retinopathy, general preventive strategies, and novel therapeutic targets. Biomed. Res. Int. 2014, 2014, 801269. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlop, M. Aldose reductase and the role of the polyol pathway in diabetic nephropathy. Kidney Int. Suppl. 2000, 58, S3–S12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, W.H.; Martin, K.A.; Hwa, J. Aldose reductase, oxidative stress, and diabetic mellitus. Front. Pharmacol. 2012, 3, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayir, H. Reactive oxygen species. Crit. Care Med. 2005, 33, S498–S501. [Google Scholar] [CrossRef]
- Drew, B.; Leeuwenburgh, C. Aging and the role of reactive nitrogen species. Ann. N. Y. Acad. Sci. 2002, 959, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Szabó, C. Role of peroxynitrite in the pathogenesis of cardiovascular complications of diabetes. Curr. Opin. Pharmacol. 2006, 6, 136–141. [Google Scholar] [CrossRef]
- Laube, H. Acarbose. Clin. Drug Investig. 2002, 22, 141–156. [Google Scholar] [CrossRef]
- Yang, Y.; Tan, Y.X.; Chen, R.Y.; Kang, J. The latest review on the polyphenols and their bioactivities of Chinese Morus plants. J. Asian Nat. Prod. Res. 2014, 16, 690–702. [Google Scholar] [CrossRef]
- Ramappa, V.K.; Seema, C. Mulberry: Life enhancer. J. Med. Plants Res. 2008, 2, 271–278. [Google Scholar]
- Paudel, P.; Seong, S.H.; Wagle, A.; Min, B.S.; Jung, H.A.; Choi, J.S. Antioxidant and anti-browning property of 2-arylbenzofuran derivatives from Morus alba Linn root bark. Food Chem. 2020, 309, 125739. [Google Scholar] [CrossRef]
- Dat, N.T.; Jin, X.; Lee, K.; Hong, Y.S.; Kim, Y.H.; Lee, J.J. Hypoxia-inducible factor-1 inhibitory benzofurans and chalcone-derived diels-alder adducts from Morus species. J. Nat. Prod. 2009, 72, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.W.; Park, J.H.; Lee, Y.G.; Seo, K.H.; Oh, E.J.; Lee, D.Y.; Lim, D.W.; Han, D.; Baek, N.I. Three new isoprenylated flavonoids from the root bark of Morus alba. Molecules 2016, 21, 1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.T.; Seong, S.H.; Nguyen, T.D.; Cho, W.K.; Ah, K.J.; Ma, J.Y.; Woo, M.H.; Choi, J.S.; Min, B.S. Chalcone derivatives from the root bark of Morus alba L. act as inhibitors of PTP1B and α-glucosidase. Phytochemistry 2018, 155, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.H.; Ha, M.T.; Min, B.S.; Jung, H.A.; Choi, J.S. Moracin derivatives from Morus Radix as dual BACE1 and cholinesterase inhibitors with antioxidant and anti-glycation capacities. Life Sci. 2018, 210, 20–28. [Google Scholar] [CrossRef]
- Hong, S.; Kwon, J.; Kim, D.W.; Lee, H.J.; Lee, D.; Mar, W. Mulberrofuran G protects ischemic injury-induced cell death via inhibition of NOX4-mediated ROS generation and ER stress. Phytother. Res. 2017, 31, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Koirala, P.; Seong, S.H.; Zhou, Y.; Shrestha, S.; Jung, H.A.; Choi, J.S. Structure–activity relationship of the tyrosinase inhibitors kuwanon G, mulberrofuran G, and albanol B from Morus species: A kinetics and molecular docking study. Molecules 2018, 23, 1413. [Google Scholar] [CrossRef] [Green Version]
- Paudel, P.; Yu, T.; Seong, S.H.; Kuk, E.B.; Jung, H.A.; Choi, J.S. Protein tyrosine phosphatase 1B inhibition and glucose uptake potentials of mulberrofuran G, albanol B, and kuwanon G from root bark of Morus alba L. in insulin-resistant HepG2 cells: An in vitro and in silico study. Int. J. Mol. Sci. 2018, 19, 1542. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, Y.; Wang, Y.; Zhang, Y. Bioassay-guided screening and isolation of α-glucosidase and tyrosinase inhibitors from leaves of Morus alba. Food Chem. 2012, 131, 617–625. [Google Scholar] [CrossRef]
- Jeong, S.H.; Ryu, Y.B.; Curtis Long, M.J.; Ryu, H.W.; Baek, Y.S.; Kang, J.E.; Lee, W.S.; Park, K.H. Tyrosinase inhibitory polyphenols from roots of Morus lhou. J. Agric. Food Chem. 2009, 57, 1195–1203. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, W.S.; Kim, Y.S.; Curtis-Long, M.J.; Lee, B.W.; Ryu, Y.B.; Park, K.H. Isolation of cholinesterase-inhibiting flavonoids from Morus lhou. J. Agric. Food Chem. 2011, 59, 4589–4596. [Google Scholar] [CrossRef]
- Shim, S.Y.; Sung, S.H.; Lee, M. Anti-inflammatory activity of mulberrofuran K isolated from the bark of Morus bombycis. Int. Immunopharmacol. 2018, 58, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.I.; Jin, Y.S.; Jung, M.J.; Wang, M.H. Antidiabetic properties of 2,5-dihydroxy-4,3′-di-(beta-D-glucopyranosyloxy)-trans-stilbene from mulberry (Morus bombycis koidzumi) root in streptozotocin-induced diabetic rats. J. Med. Food 2007, 10, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.M.; Ngoc, T.M.; Dat, N.T.; Kim, Y.H.; Van Luong, H.; Ahn, J.S.; Bae, K. Protein tyrosine phosphatase 1B inhibitors isolated from Morus bombycis. Bioorg. Med. Chem. Lett. 2009, 19, 6759–6761. [Google Scholar] [CrossRef]
- Kim, Y.S.; Lee, Y.M.; Kim, H.; Kim, J.; Jang, D.S.; Kim, J.H.; Kim, J.S. Anti-obesity effect of Morus bombycis root extract: Anti-lipase activity and lipolytic effect. J. Ethnopharmacol. 2010, 130, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.B.; Kim, S.D.; Kim, T.B.; Jeong, E.J.; Kim, Y.C.; Sung, J.H.; Sung, S.H. Tyrosinase inhibitory constituents of Morus bombycis cortex. Nat. Prod. Sci. 2011, 17, 198–201. [Google Scholar]
- Moon, M.; Choi, J.G.; Kim, S.Y.; Oh, M.S. Bombycis excrementum reduces amyloid-beta oligomer-induced memory impairments, neurodegeneration, and neuroinflammation in mice. J. Alzheimers Dis. 2014, 41, 599–613. [Google Scholar] [CrossRef]
- Tomczyk, M.; Miłek, M.; Sidor, E.; Kapusta, I.; Litwińczuk, W.; Puchalski, C.; Dżugan, M. The effect of adding the leaves and fruits of Morus alba to rape honey on its antioxidant properties, polyphenolic profile, and amylase activity. Molecules 2019, 25, 84. [Google Scholar] [CrossRef] [Green Version]
- Gross, V.; Tran Thi, T.A.; Schwarz, R.; Elbein, A.; Decker, K.; Heinrich, P. Different effects of the glucosidase inhibitors 1-deoxynojirimycin, N-methyl-1-deoxynojirimycin and castanospermine on the glycosylation of rat α1-proteinase inhibitor and α1-acid glycoprotein. Biochem. J. 1986, 236, 853–860. [Google Scholar] [CrossRef]
- Rho, T.; Yoon, K.D. Chemical constituents of Nelumbo nucifera seeds. Nat. Prod. Sci. 2017, 23, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.W.; Kim, K.H.; Lee, I.K.; Choi, S.U.; Ryu, S.Y.; Lee, K.R. Phytochemical constituents of Bistorta manshuriensis. Nat. Prod. Sci. 2009, 15, 234–240. [Google Scholar]
- Ban, J.Y.; Jeon, S.Y.; Nguyen, T.T.; Bae, K.; Song, K.S.; Seong, Y.H. Neuroprotective effect of oxyresveratrol from Smilacis Chinae rhizome on amyloid β protein (25-35)-induced neurotoxicity in cultured rat cortical neurons. Biol. Pharm. Bull. 2006, 29, 2419–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basnet, P.; Kadota, S.; Terashima, S.; Shimizu, M.; Namba, T. Two new 2-arylbenzofuran derivatives from hypoglycemic activity-bearing fractions of Morus insignis. Chem. Pharm. Bull. 1993, 41, 1238–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.S.; Wang, Y.H.; Li, R.L.; Lin, M. Five new stilbene dimers from the lianas of Gnetum hainanense. J. Nat. Prod. 2000, 63, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.J.; Zheng, Z.F.; Chen, R.Y.; Yu, D.Q. Two new dimeric stilbenes from the stem bark of Morus australis. J. Asian Nat. Prod. Res. 2009, 11, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Liu, L.; Zhang, S.; Yang, C.; Yue, W.; Zhao, H.; Ho, C.T.; Du, J.; Zhang, H.; Bai, N. Chemical characterization of the main bioactive polyphenols from the roots of Morus australis (Mulberry). Food Funct. 2019, 10, 6915–6926. [Google Scholar] [CrossRef]
- Happyana, N.; Hakim, E.H.; Syah, Y.M.; Kayser, O.; Juliawaty, L.D.; Mujahidin, D.; Ermayanti, T.M.; Achmad, S.A. Diels-Alder type adducts from hairy root cultures of Morus macroura. Nat. Prod. Sci. 2019, 25, 233–237. [Google Scholar] [CrossRef] [Green Version]
- National Institute of Agricultural Sciences (NAS); Rural Development Administration (RDA). RDA DB 1.0—Flavonoids; Rural Development Administration (RDA): Wanju, Republic of Korea, 2016. [Google Scholar]
- Jung, H.A.; Jung, Y.J.; Yoon, N.Y.; Jeong, D.M.; Bae, H.J.; Kim, D.W.; Na, D.H.; Choi, J.S. Inhibitory effects of Nelumbo nucifera leaves on rat lens aldose reductase, advanced glycation endproducts formation, and oxidative stress. Food Chem. Toxicol. 2008, 46, 3818–3826. [Google Scholar] [CrossRef]
- Kwon, R.H.; Thaku, N.; Timalsina, B.; Park, S.E.; Choi, J.S.; Jung, H.A. Inhibition mechanism of components isolated from Morus alba branches on diabetes and diabetic complications via experimental and molecular docking analyses. Antioxidants 2022, 11, 383. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Dixon, M. The determination of enzyme inhibitor constants. Biochem. J. 1953, 55, 170. [Google Scholar] [CrossRef]
- Eisenthal, R.; Cornish Bowden, A. The direct linear plot. A new graphical procedure for estimating enzyme kinetic parameters. Biochem. J. 1974, 139, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Y.; Zamponi, G.W.; Seong, S.H.; Jung, H.A.; Choi, J.S. 6-Formyl umbelliferone, a furanocoumarin from Angelica decursiva L., inhibits key diabetes-related enzymes and advanced glycation end-product formation. Molecules 2022, 27, 5720. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. Autodockvina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [PubMed] [Green Version]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.A.; Jin, S.E.; Park, J.S.; Choi, J.S. Antidiabetic complications and anti-Alzheimer activities of sophoflavescenol, a prenylated flavonol from Sophora flavescens, and its structure–activity relationship. Phytother. Res. 2011, 25, 709–715. [Google Scholar] [CrossRef]
- Johnson, T.O.; Ermolieff, J.; Jirousek, M.R. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat. Rev. Drug Discov. 2002, 1, 696–709. [Google Scholar] [CrossRef]
- Singh, S.; Grewal, A.S.; Grover, R.; Sharma, N.; Chopra, B.; Dhingra, A.K.; Arora, S.; Redhu, S.; Lather, V. Recent updates on development of protein-tyrosine phosphatase 1B inhibitors for treatment of diabetes, obesity and related disorders. Bioorg. Chem. 2022, 121, 105626. [Google Scholar] [CrossRef]
- Nilsson, B.O. Biological effects of aminoguanidine: An update. Inflamm. Res. 1999, 48, 509–515. [Google Scholar] [CrossRef]
- Soung, D.Y.; Kim, J.S.; Chung, H.Y.; Jung, H.A.; Park, J.C.; Choi, J.S. Flavonoids and chlorogenic acid from Eriobotrya japonica scavenge peroxynitrite. Nat. Prod. Sci. 1999, 5, 80–84. [Google Scholar]
- Ren, H.; Xu, Q.L.; Zhang, M.; Dong, L.M.; Zhang, Q.; Luo, B.; Luo, Q.W.; Tan, J.W. Bioactive caffeic acid derivatives from Wedelia trilobata. Phytochem. Lett. 2017, 19, 18–22. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, Y.; Su, H.; Wu, L.; Huang, Y.; Zhao, L.; Han, B.; Shu, G.; Xiang, M.; Yang, J.M. Chlorogenic acid methyl ester exerts strong anti-inflammatory effects via inhibiting the COX-2/NLRP3/NF-κB pathway. Food Funct. 2018, 9, 6155–6164. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.X.; Liu, C.; Zhang, T.; Chen, R.Y.; Yu, D.Q. Bioactive constituents of Morus wittiorum. Phytochem. Lett. 2010, 3, 57–61. [Google Scholar] [CrossRef]
- Wang, M.; Gao, L.X.; Wang, J.; Li, J.Y.; Yu, M.H.; Li, J.; Hou, A.J. Diels–Alder adducts with PTP1B inhibition from Morus notabilis. Phytochemistry 2015, 109, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Leach, A.R.; Shoichet, B.K.; Peishoff, C.E. Prediction of protein−ligand interactions. Docking and scoring: Successes and gaps. J. Med. Chem. 2006, 49, 5851–5855. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Cho, Y.S.; Oh, S.H.; Lee, S.; Min, B.S.; Moon, K.H.; Choi, J.S. Kinetics and molecular docking studies of pimarane-type diterpenes as protein tyrosine phosphatase (PTP1B) inhibitors from Aralia continentalis roots. Arch. Pharm.Res. 2013, 36, 957–965. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | TPC (mg GAE/g) a | TFC (mg CE/g) a | ABTS IC50 (μg/mL) a | DPPH IC50 (μg/mL) a | α-Glucosidase IC50 (μg/mL) a |
|---|---|---|---|---|---|
| Morus alba | 47.91 ± 0.08 | 74.80 ± 0.47 | 72.25 ± 0.75 | 49.83 ± 2.87 | 319.26 ± 22.71 |
| Morus lhou | 57.82 ± 0.52 | 112.93 ± 0.88 | 132.49 ± 8.24 | 24.25 ± 0.59 | 148.24 ± 16.29 |
| Morus bombycis | 62.67 ± 0.18 | 87.70 ± 1.67 | 95.74 ± 19.52 | 24.30 ± 0.62 | 26.35 ± 2.98 |
| Trolox b | 2.72 ± 0.13 | ||||
| L-ascorbic acid b | 3.38 ± 0.19 | 2.82 ± 0.57 | |||
| Acarbose b | 352.09 ± 22.27 |
| Species | TPC (mg GAE/g) a | TFC (mg CE/g) a | ABTS IC50 (μg/mL) a | DPPH IC50 (μg/mL) a | α-Glucosidase IC50 (μg/mL) a | PTP1B IC50 (μg/mL) a |
|---|---|---|---|---|---|---|
| MeOH ext. | 62.67 ± 0.18 | 87.70 ± 1.67 | 95.74 ± 19.52 | 24.30 ± 0.62 | 26.35 ± 2.98 | 24.71 ± 2.92 |
| CH2Cl2 fr. | 55.71 ± 1.17 | 208.45 ± 0.98 | 85.16 ± 13.13 | 115.34 ± 25.41 | 57.67 ± 2.01 | 7.09 ± 0.72 |
| EtOAc fr. | 261.59 ± 3.58 | 260.43 ± 9.07 | 12.84 ± 1.23 | 4.66 ± 0.65 | 6.74 ± 1.57 | 25.17 ± 1.66 |
| n-BuOH fr. | 138.18 ± 0.72 | 173.16 ± 0.93 | 18.75 ± 0.94 | 7.60 ± 0.36 | 18.63 ± 1.60 | 46.32 ± 3.39 |
| H2O fr. | 40.45 ± 1.82 | 37.96 ± 1.30 | 98.65 ± 11.42 | 37.12 ± 1.18 | 39.73 ± 9.04 | 115.52 ± 12.44 |
| Trolox b | 2.72 ± 0.13 | |||||
| L-ascorbic acid b | 3.38 ± 0.19 | 2.82 ± 0.57 | ||||
| Acarbose b | 352.09 ± 22.27 | |||||
| Ursolic acid b | 6.39 ± 0.42 |
| Test Compounds | Peroxynitirite | α-Glucosidase | PTP1B | ||||
|---|---|---|---|---|---|---|---|
| IC50 (μM) a | IC50 (μM) a | Inhibition Mode c | Inhibition Constant (Ki) d | IC50 (μM) a | Inhibition Mode c | Inhibition Constant (Ki) d | |
| Oxyresveratrol (3) | 6.24 ± 0.06 | 2.58 ± 0.23 | - | 72.88 ± 1.87 | - | ||
| Macrourin B (4) | 2.61 ± 0.13 | 0.44 ± 0.03 | Mixed | 0.19 | 2.50 ± 0.17 | Mixed | 1.54 |
| Moracin M (5) | 0.92 ± 0.13 | 6.11 ± 0.53 | - | 27.14 ± 4.20 | - | ||
| Austrafuran C (6) | 2.76 ± 0.25 | 1.01 ± 0.17 | Mixed | 0.75 | 1.69 ± 0.02 | Mixed | 1.45 |
| Mulberrofuran F (7) | 8.63 ± 0.02 | 1.22 ± 0.05 | Mixed | 1.84 | 10.53 ± 0.25 | Mixed | 8.90 |
| Chalcomoracin (8) | 3.03 ± 0.53 | 0.98 ± 0.03 | Mixed | 1.71 | 2.06 ± 0.39 | Non-competitive | 4.41 |
| L-Penicillamine b | 0.62 ± 0.17 | ||||||
| Acarbose b | 321.46 ± 21.13 | ||||||
| Ursolic acid b | 13.53 ± 0.18 | ||||||
| Test Compounds | BSA-AGEs IC50 (μM) a | HAS-AGEs IC50 (μM) a | HRAR IC50 (μM) c |
|---|---|---|---|
| Oxyresveratrol (3) | 10.36 ± 0.39 | 5.38 ± 0.26 | 264.8 |
| Macrourin B (4) | 9.44 ± 0.18 | 7.98 ± 0.62 | <4 |
| Moracin M (5) | 2.40 ± 0.18 | 2.07 ± 0.03 | 238.4 |
| Austrafuran C (6) | 13.74 ± 0.55 | 6.15 ± 0.25 | 26.78 |
| Mulberrofuran F (7) | ND | ND | 337.3 |
| Chalcomoracin (8) | 137.60 ± 0.33 | 112.59 ± 3.62 | 265.0 |
| Aminoguanidine b | 581.03 ± 28.67 | 504.07 ± 14.92 | |
| Quercetin bc | 16.67 |
| Compounds | Number of H-Bonds | Binding Energy (kcal/mol) | Hydrogen Bonds Interacting Residues | Hydrophobic Interacting Residues | Electrostatic Interacting Residues |
|---|---|---|---|---|---|
| Macrourin B (4) | 3 | −11.1 | Asp352, Asp307, Glu411 | Arg315 (carbon–hydrogen bond), Val216 (π-alkyl), Tyr158 (unfavorable acceptor–acceptor), Phe303 (π-π stacked), Tyr158 (π-π T-shaped), Tyr72 (π-π T-shaped) | Arg442 (π-cation), Asp352 (π-anion) |
| Austrafuran C (6) | 1 | −9.3 | Leu318 | Lys432 (unfavorable donor–donor), Ile419 (alky), Phe314 (π-alkyl), Leu313 (π-alkyl) | Asp233 (π-anion) |
| Mulberrofuran F (7) | 3 | −10.6 | Asp242, Glu332, Asp307 | Ala329 (alkyl), Ile328 (alkyl), Arg315 (π-alkyl), Pro312 (π-alkyl), His280 (π-alkyl), His280 (π-π stacked) | |
| Chalcomoracin (8) | 5 | −11.5 | Arg315, Asp242, Lys156, Ser241, Pro312 | His280 (unfavorable acceptor–acceptor), Ser240 (carbon–hydrogen bond), Tyr158 (π-π stacked), Pro243 (alkyl), Val232 (alkyl), Arg315 (alkyl), Arg315 (π-alkyl), Lys156 (π-alkyl), Pro312 (π-sigma) | Asp307 (π-anion), |
| Acarbose | 7 | −8.2 | Asp352, Asp215, Arg442, Gln279, Pro312, Ser240, Tyr158 | Pro312 (carbon–hydrogen bond), His280 (π-sigma), Glu411 (unfavorable acceptor–acceptor) | |
| Alpha-D-glucose | 8 | −6.8 | Asp69, Arg442, Arg213, Asp352, Asp215, Glu277, His112, His351 | Tyr72 (π-donor–hydrogen bond), Asp69 (carbon–hydrogen bond) |
| Compounds | Number of H-Bonds | Binding Energy (kcal/mol) | Hydrogen Bonds Interacting Residues | Hydrophobic Interacting Residues | Electrostatic Interacting Residues |
|---|---|---|---|---|---|
| Macrourin B (4) | 6 | −8.7 | Arg24, Arg221, Asp265, Gln262, Lys120, Tyr46 | Gln262 (unfavorable acceptor–acceptor) | |
| Austrafuran C (6) | 4 | −9.0 | Ala189, Asn193, Glu200, Lys197 | Leu192 (π-alkyl), Arg199 (amide-π stacked), Phe280 (π-π T-shaped), Phe280 (π-π stacked), Phe196 (π-π stacked) | Glu200 (π-anion) |
| Mulberrofuran F (7) | 3 | −7.9 | Asn193, Glu200, Arg199 | Phe196 (π-π T-shaped), Phe196 (π-π stacked), Phe280 (π-π stacked) | Lys197 (π-cation), |
| Chalcomoracin (8) | 2 | −8.1 | Arg268, Glu186 | Pro180 (π-alkyl), Tyr152 (π-alkyl), Ala189 (alkyl), Val184 (π-sigma), | Glu186 (π-anion), |
| Compound 2 (allosteric inhibitor) | 3 | −8.9 | Gly183, Gln266, Gln262 | Ala217 (π-alkyl), Lys120 (alkyl), Tyr46 (π-π stacked) | Lys116 (π-cation), Arg221 (π-cation), Asp48 (π-anion) |
| Compound 23 (catalytic inhibitor) | 2 | −8.4 | Glu276, Lys279 | Leu192 (alkyl), Ala189 (π-alkyl), Phe196 (π-alkyl), Phe280 (π-alkyl), Met282 (π-sulfur), Phe280 (π-sigma), Phe280 (π-π T-shaped), Phe280 (π-π stacked) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ju, S.M.; Ali, M.Y.; Ko, S.-M.; Ryu, J.-H.; Choi, J.-S.; Jung, H.-A. An Arylbenzofuran, Stilbene Dimers, and Prenylated Diels–Alder Adducts as Potent Diabetic Inhibitors from Morus bombycis Leaves. Antioxidants 2023, 12, 837. https://doi.org/10.3390/antiox12040837
Ju SM, Ali MY, Ko S-M, Ryu J-H, Choi J-S, Jung H-A. An Arylbenzofuran, Stilbene Dimers, and Prenylated Diels–Alder Adducts as Potent Diabetic Inhibitors from Morus bombycis Leaves. Antioxidants. 2023; 12(4):837. https://doi.org/10.3390/antiox12040837
Chicago/Turabian StyleJu, Seon Min, Md Yousof Ali, Seung-Mi Ko, Jung-Hye Ryu, Jae-Sue Choi, and Hyun-Ah Jung. 2023. "An Arylbenzofuran, Stilbene Dimers, and Prenylated Diels–Alder Adducts as Potent Diabetic Inhibitors from Morus bombycis Leaves" Antioxidants 12, no. 4: 837. https://doi.org/10.3390/antiox12040837