
Milk Fat Globule Membrane Relieves Fatigue via Regulation of Oxidative Stress and Gut Microbiota in BALB/c Mice
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Amino Acids of MFGM
2.1.2. Phospholipids of MFGM
2.2. Animal Experiment
2.3. Exhaustive Swimming Exercise Test
2.4. Free Swimming Test
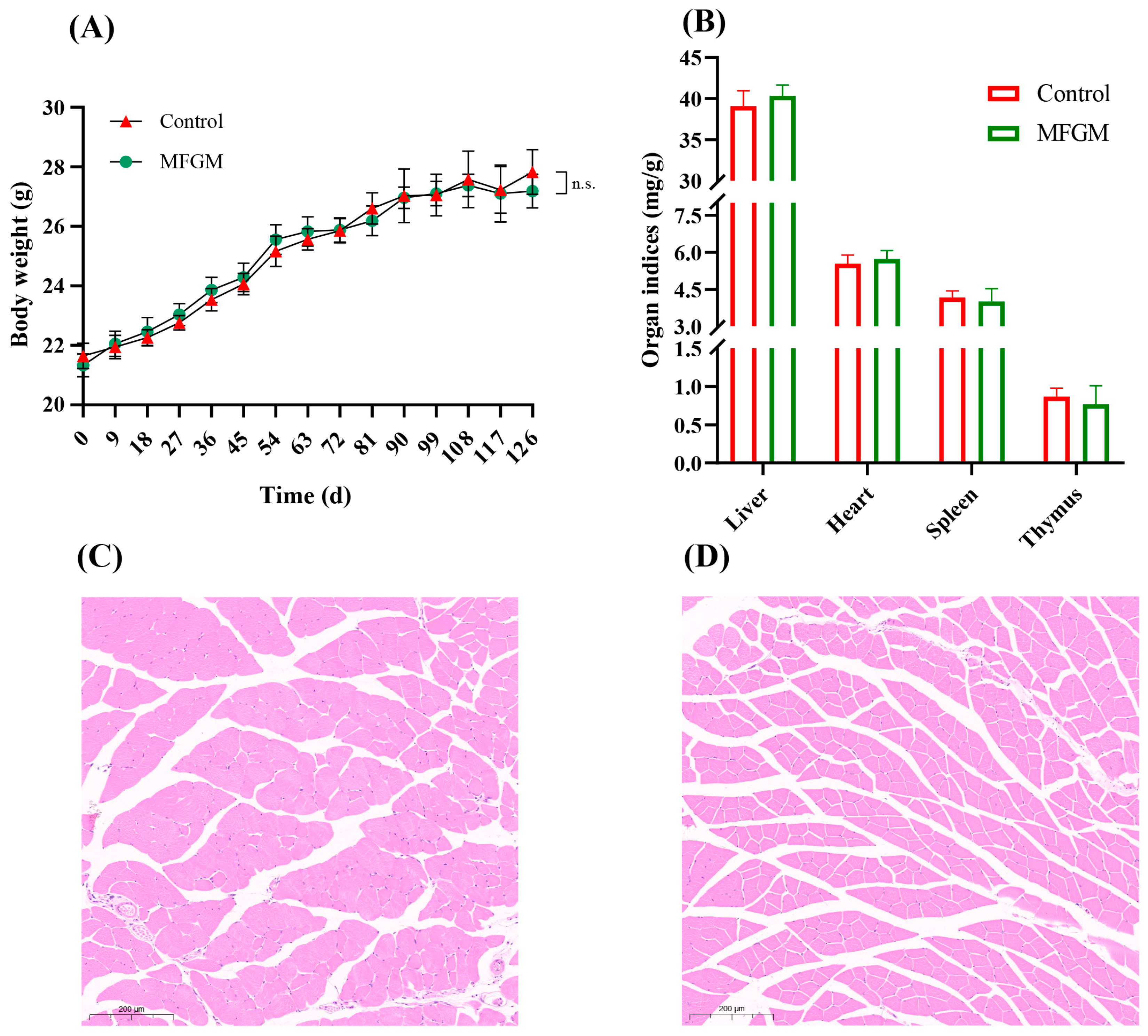
2.5. Organ and Tissue Collection
2.6. Serum Biochemical Indices
2.7. Histological Analysis
2.8. Immunohistochemical Analysis
2.9. Western Blot Analysis
2.10. 16S rRNA Gene Sequencing and Bioinformatic Analysis
2.11. Statistical Analysis
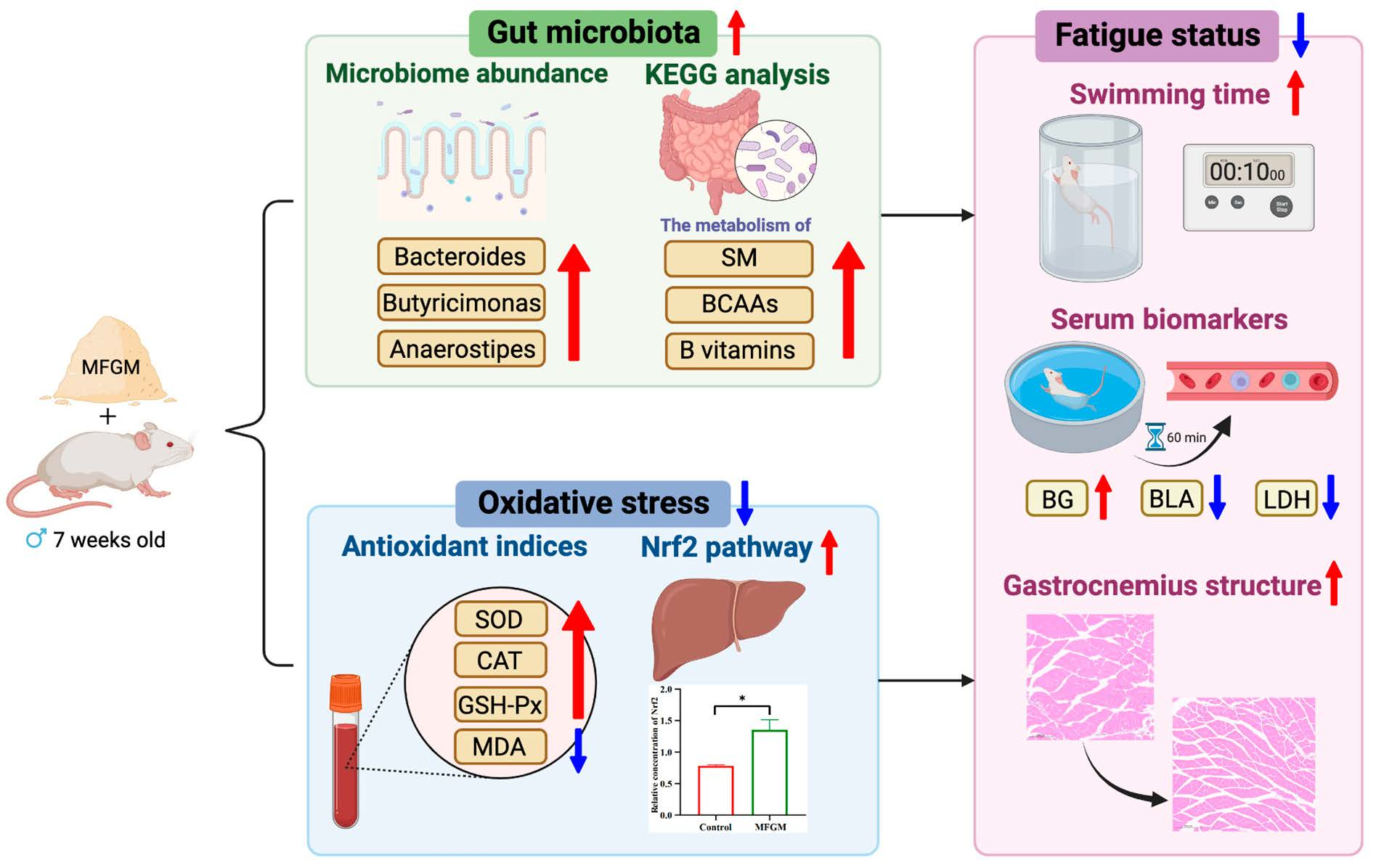
3. Results and Discussion
3.1. Animal Metrics and Structure of Muscle Tissue
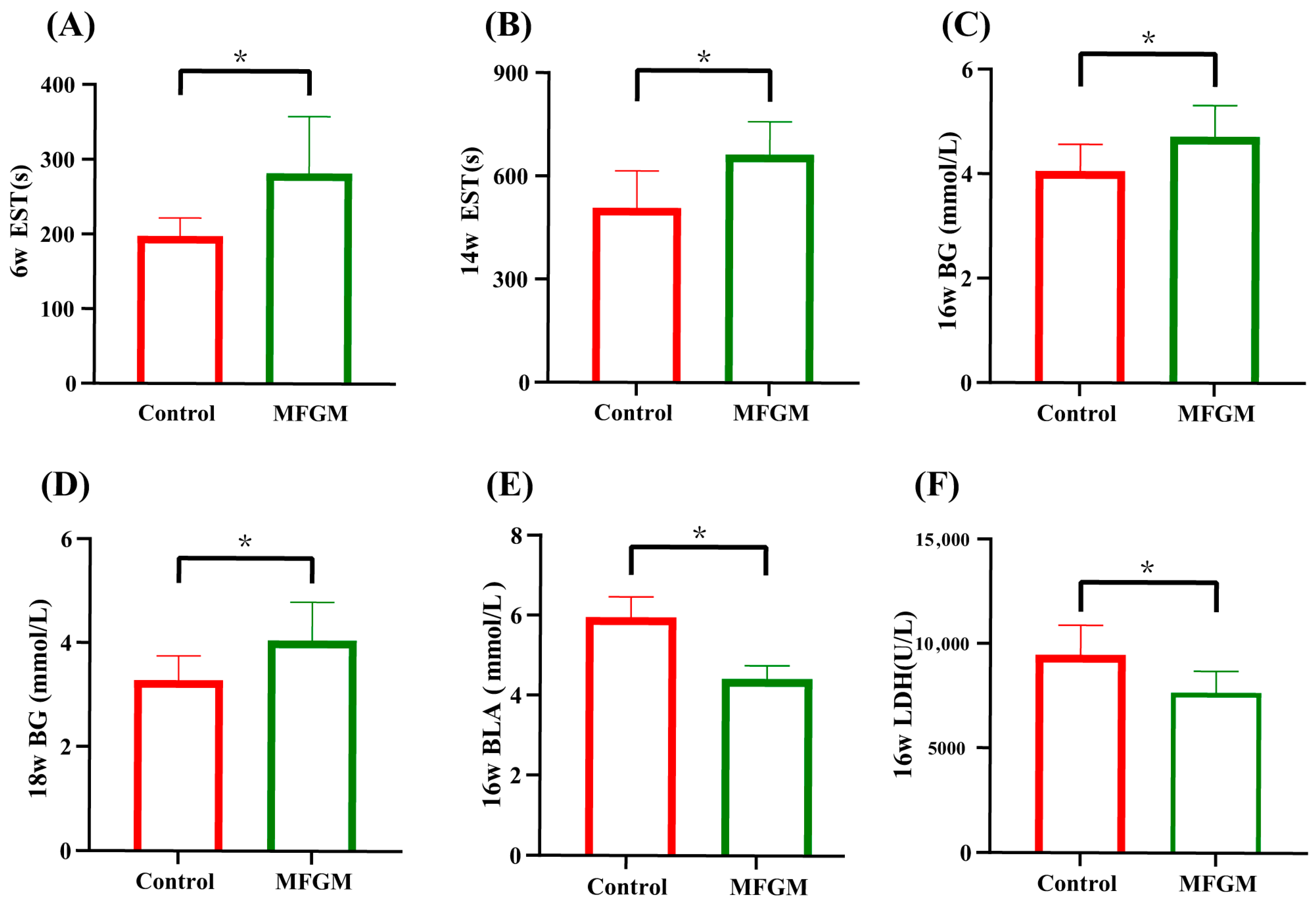
3.2. Swimming Test and Serum Biomarkers of Antifatigue Status
3.3. Serum Biomarkers of Antioxidant Enzymes
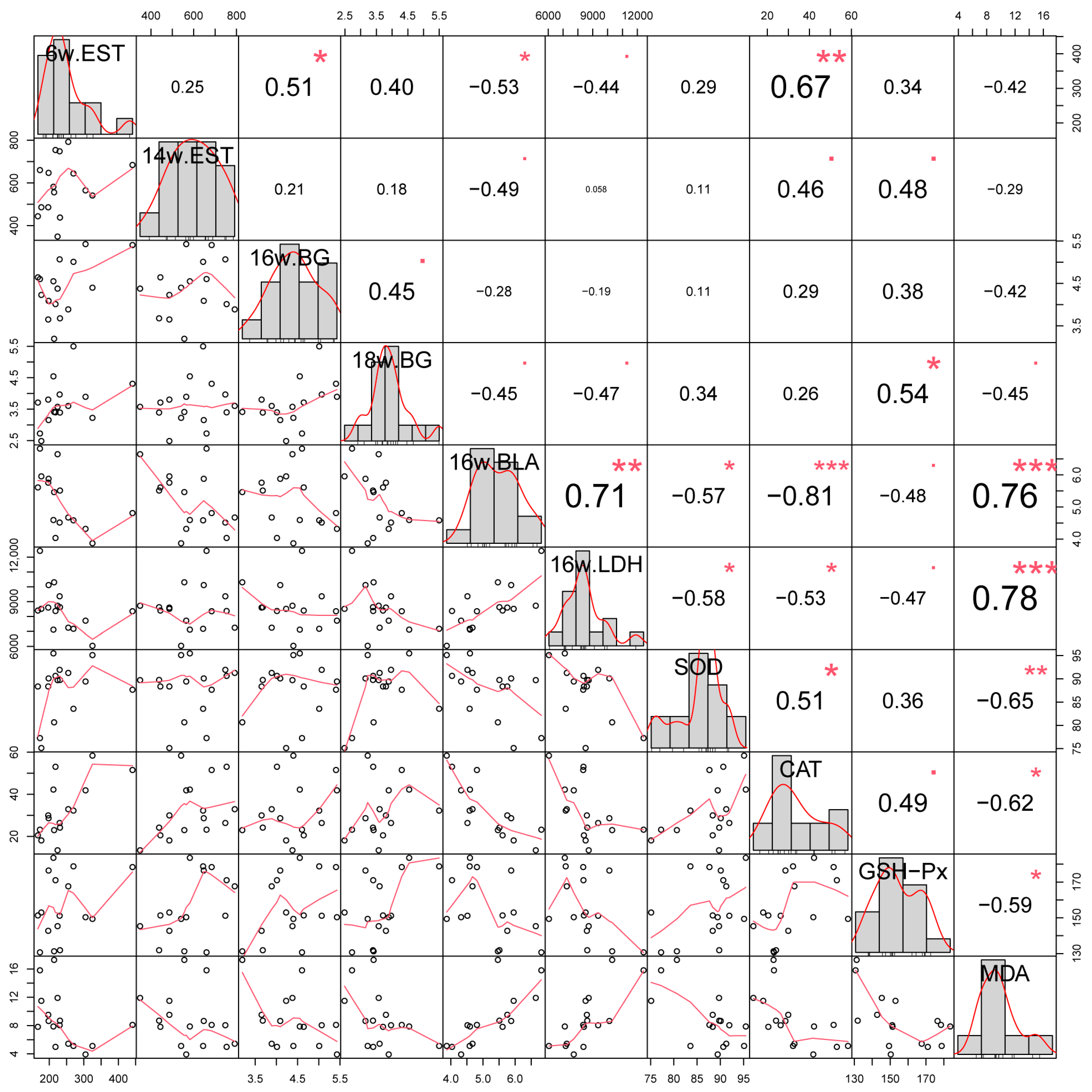
3.4. Pearson Correlation Analysis of Different Indicators
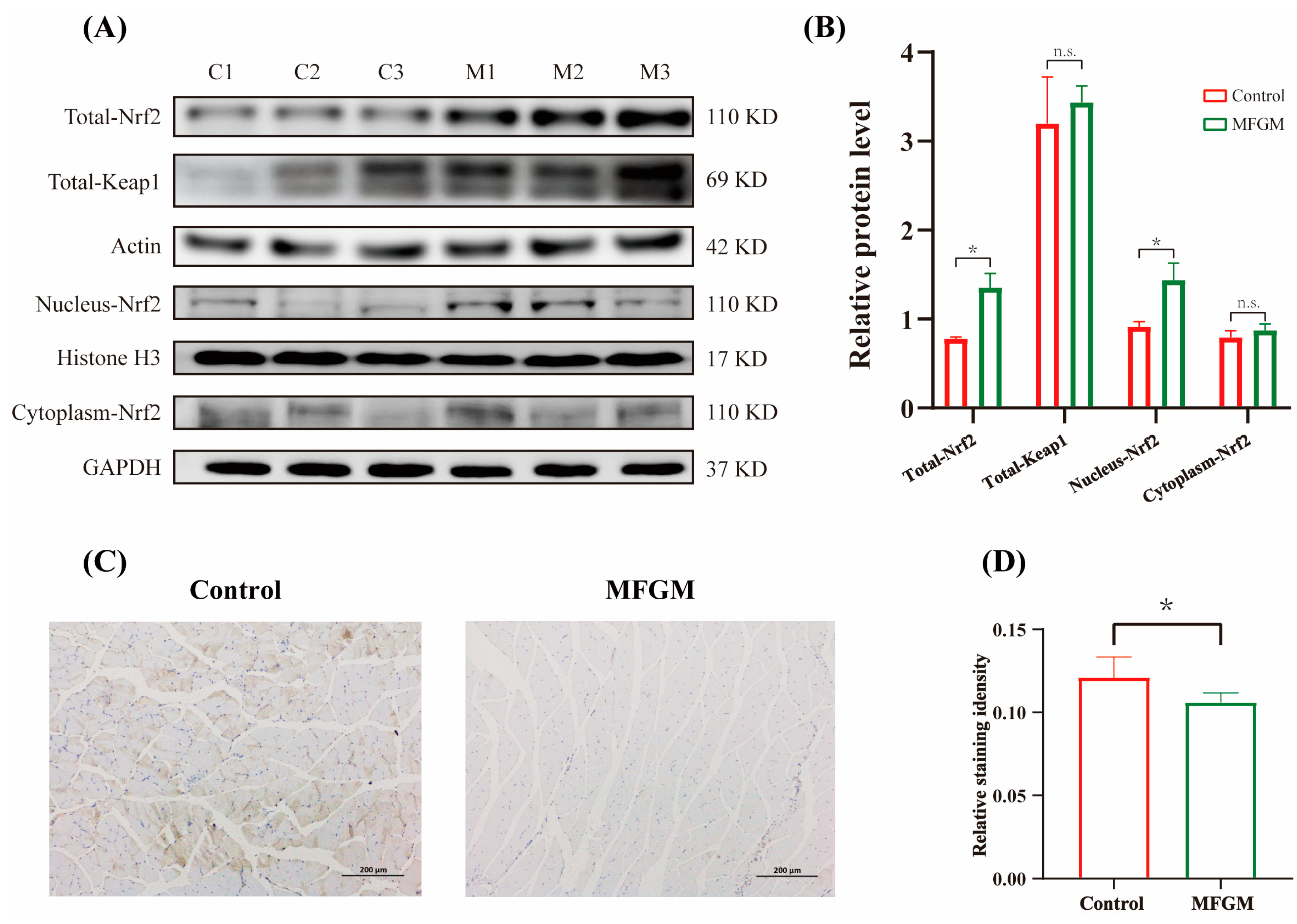
3.5. Effects of MFGM on Protein Expressions in Muscle and Liver
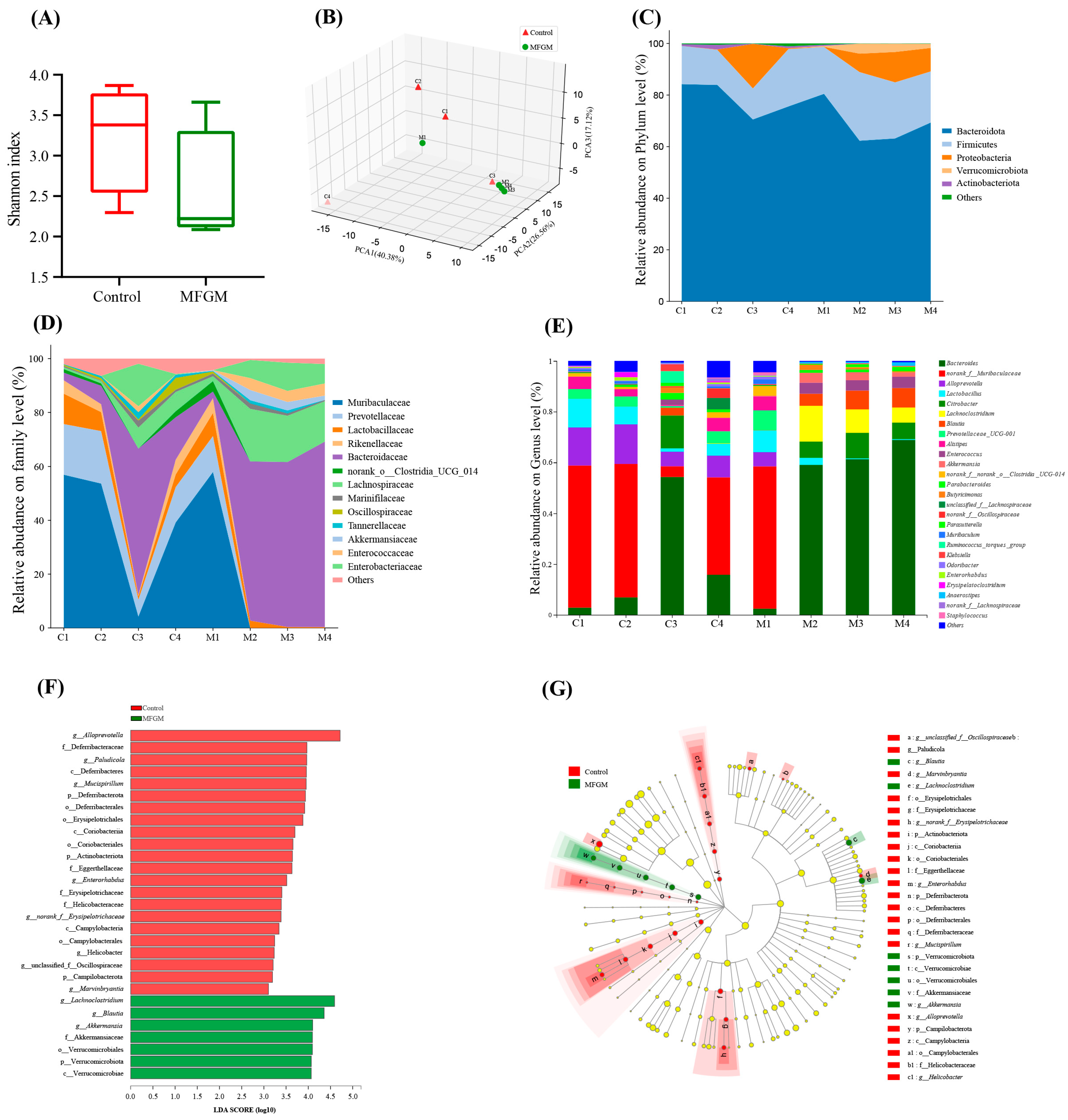
3.6. Effects of MFGM on the Relative Abundance and Composition of Gut Microbiota
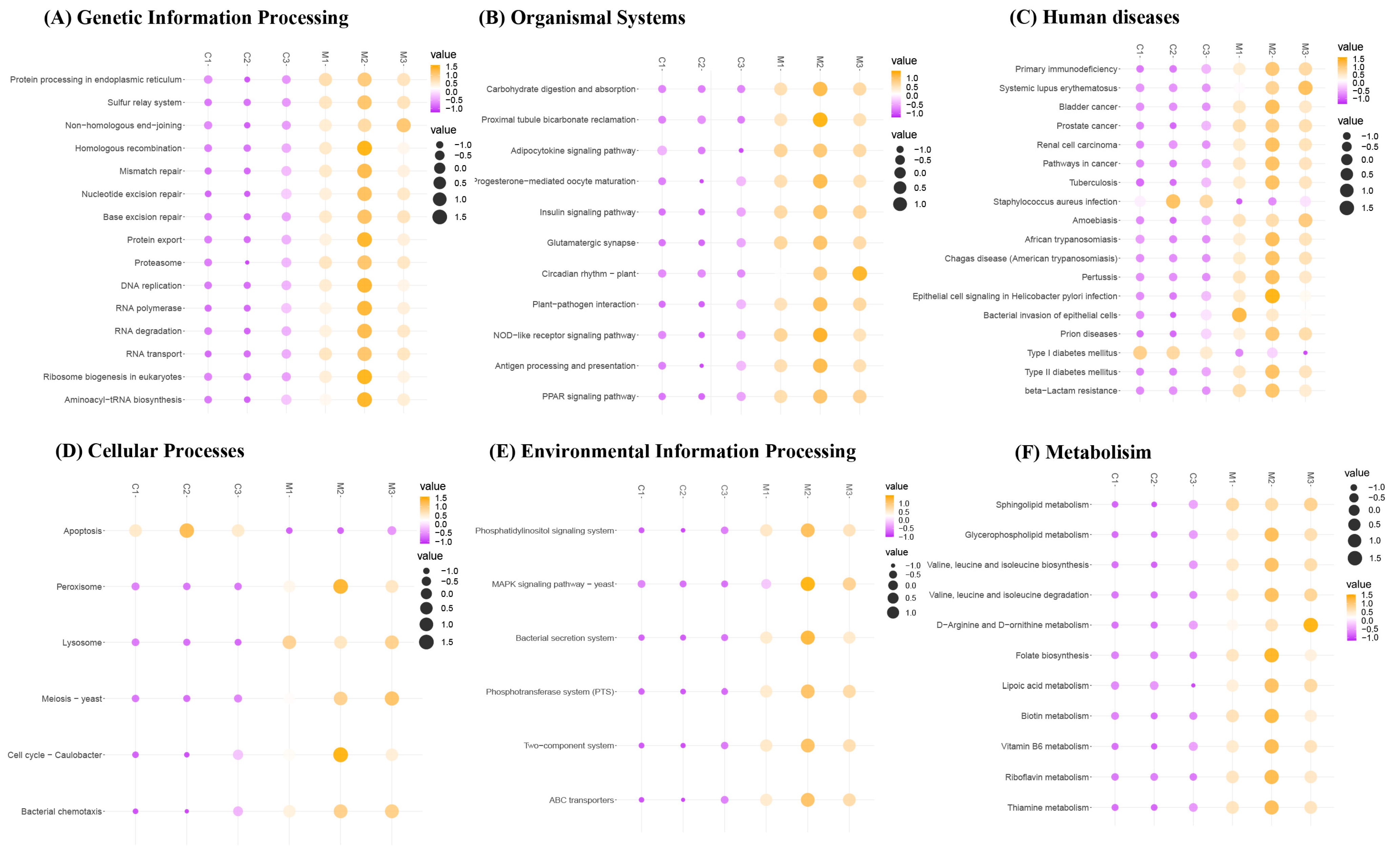
3.7. Effects of MFGM on the Functional Changes in Microbial Communities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wan, J.; Qin, Z.; Wang, P.; Sun, Y.; Liu, X. Muscle Fatigue: General Understanding and Treatment. Exp. Mol. Med. 2017, 49, e384. [Google Scholar] [CrossRef]
- Barclay, J.K.; Hansel, M. Free Radicals May Contribute to Oxidative Skeletal Muscle Fatigue. Can. J. Physiol. Pharmacol. 1991, 69, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.-M.; Wei, P. Anti-Fatigue Properties of Tartary Buckwheat Extracts in Mice. Int. J. Mol. Sci. 2011, 12, 4770–4780. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xing, R.; Chen, Z.; Yu, H.; Li, R.; Li, P. Effect and Mechanism of Mackerel (Pneumatophorus Japonicus) Peptides for Anti-Fatigue. Food Funct. 2014, 5, 2113. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. In Vitro Antioxidant Activity and in Vivo Anti-Fatigue Effect of Loach (Misgurnus Anguillicaudatus) Peptides Prepared by Papain Digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
- Mach, N.; Fuster-Botella, D. Endurance Exercise and Gut Microbiota: A Review. J. Sport Health Sci. 2017, 6, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, J.; Xu, F.; Liu, G.; Pang, B.; Liao, N.; Li, H.; Shi, J. Gut Microbiota as a Potential Target for Developing Anti-Fatigue Foods. In Critical Reviews in Food Science and Nutrition; Taylor & Francis: New York, NY, USA, 2021; pp. 1–16. [Google Scholar] [CrossRef]
- Li, D.; Ren, J.; Zhang, T.; Liu, R.; Wu, L.; Du, Q.; Li, Y. Anti-Fatigue Effects of Small-Molecule Oligopeptides Isolated from Panax Quinquefolium L. in Mice. Food Funct. 2018, 9, 4266–4273. [Google Scholar] [CrossRef] [PubMed]
- Contarini, G.; Povolo, M. Phospholipids in Milk Fat: Composition, Biological and Technological Significance, and Analytical Strategies. Int. J. Mol. Sci. 2013, 14, 2808–2831. [Google Scholar] [CrossRef] [PubMed]
- Moukarzel, S.; Dyer, R.A.; Garcia, C.; Wiedeman, A.M.; Boyce, G.; Weinberg, J.; Keller, B.O.; Elango, R.; Innis, S.M. Milk Fat Globule Membrane Supplementation in Formula-Fed Rat Pups Improves Reflex Development and May Alter Brain Lipid Composition. Sci. Rep. 2018, 8, 15277. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, X.; Han, D.; Pi, Y.; Tao, S.; Zhang, S.; Wang, S.; Zhao, J.; Chen, L.; Wang, J. Early Life Administration of Milk Fat Globule Membrane Promoted SCFA-Producing Bacteria Colonization, Intestinal Barriers and Growth Performance of Neonatal Piglets. Anim. Nutr. 2021, 7, 346–355. [Google Scholar] [CrossRef]
- Yuan, Q.; Zhan, B.; Du, M.; Chang, R.; Li, T.; Mao, X. Dietary Milk Fat Globule Membrane Regulates JNK and PI3K/Akt Pathway and Ameliorates Type 2 Diabetes in Mice Induced by a High-Fat Diet and Streptozotocin. J. Funct. Foods 2019, 60, 103435. [Google Scholar] [CrossRef]
- Haramizu, S.; Ota, N.; Otsuka, A.; Hashizume, K.; Sugita, S.; Hase, T.; Murase, T.; Shimotoyodome, A. Dietary Milk Fat Globule Membrane Improves Endurance Capacity in Mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R1009–R1017. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 19th ed.; Association of Official Analytical Chemists: Rockville, MA, USA, 2012. [Google Scholar]
- Li, M.; Li, W.; Kong, F.; Kang, S.; Liang, X.; Han, H.; Wu, J.; Zheng, Y.; Li, Q.; Yue, X.; et al. Metabolomics Methods to Analyze Full Spectrum of Amino Acids in Different Domains of Bovine Colostrum and Mature Milk. Eur. Food Res. Technol. 2020, 246, 213–224. [Google Scholar] [CrossRef]
- Varache, M.; Ciancone, M.; Couffin, A.-C. Development and Validation of a Novel UPLC-ELSD Method for the Assessment of Lipid Composition of Nanomedicine Formulation. Int. J. Pharm. 2019, 566, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Yuen, H.-M.; Lin, C.-C.; Hsu, C.-C.; Bernard, J.R.; Chen, L.-N.; Liao, Y.-H.; Tsai, S.-C. Anti-Fatigue Effects of Santé Premium Silver Perch Essence on Exhaustive Swimming Exercise Performance in Rats. Front. Physiol. 2021, 12, 651972. [Google Scholar] [CrossRef]
- Sun, B.; Luo, H.; Deng, L.; Zhang, S.; Chen, Z. The Study on Mechanism of the Modified Chinese Herbal Compound, Jianpijiedu, on a Mouse Model of Hepatic Carcinoma Cachexia. Mol. Med. Rep. 2016, 14, 3113–3121. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Gao, J.; Du, M.; Mao, X. Milk Fat Globule Membrane Supplementation Modulates the Gut Microbiota and Attenuates Metabolic Endotoxemia in High-Fat Diet-Fed Mice. J. Funct. Foods 2018, 47, 56–65. [Google Scholar] [CrossRef]
- Li, H.; Xu, W.; Ma, Y.; Zhou, S.; Xiao, R. Milk Fat Globule Membrane Protein Promotes C2C12 Cell Proliferation through the PI3K/Akt Signaling Pathway. Int. J. Biol. Macromol. 2018, 114, 1305–1314. [Google Scholar] [CrossRef]
- Noakes, T.D. What Is the Evidence That Dietary Macronutrient Composition Influences Exercise Performance? A Narrative Review. Nutrients 2022, 14, 862. [Google Scholar] [CrossRef]
- Zhao, Y.-Q.; Zeng, L.; Yang, Z.-S.; Huang, F.-F.; Ding, G.-F.; Wang, B. Anti-Fatigue Effect by Peptide Fraction from Protein Hydrolysate of Croceine Croaker (Pseudosciaena Crocea) Swim Bladder through Inhibiting the Oxidative Reactions Including DNA Damage. Mar. Drugs 2016, 14, 221. [Google Scholar] [CrossRef]
- Zhao, X.-N.; Liang, J.-L.; Chen, H.-B.; Liang, Y.-E.; Guo, H.-Z.; Su, Z.-R.; Li, Y.-C.; Zeng, H.-F.; Zhang, X.-J. Anti-Fatigue and Antioxidant Activity of the Polysaccharides Isolated from Millettiae Speciosae Champ. Leguminosae. Nutrients 2015, 7, 8657–8669. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, E.A.; Blomstrand, E. Branched-Chain Amino Acids and Central Fatigue. J. Nutr. 2006, 136, 274S–276S. [Google Scholar] [CrossRef]
- Tsuda, Y.; Yamaguchi, M.; Noma, T.; Okaya, E.; Itoh, H. Combined Effect of Arginine, Valine, and Serine on Exercise-Induced Fatigue in Healthy Volunteers: A Randomized, Double-Blinded, Placebo-Controlled Crossover Study. Nutrients 2019, 11, 862. [Google Scholar] [CrossRef]
- Falavigna, G.; Junior, J.A. de A.; Rogero, M.M.; Pires, I.S. de O.; Pedrosa, R.G.; Junior, E.M.; Castro, I.A. de; Tirapegui, J. Effects of Diets Supplemented with Branched-Chain Amino Acids on the Performance and Fatigue Mechanisms of Rats Submitted to Prolonged Physical Exercise. Nutrients 2012, 4, 1767–1780. [Google Scholar] [CrossRef]
- Powers, S.K.; Jackson, M.J. Exercise-Induced Oxidative Stress: Cellular Mechanisms and Impact on Muscle Force Production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef]
- Liang, X.; Ye, Q.; Yang, M.; Wu, J.; Wu, R.; Liu, B.; Yue, X. Milk Fat Globule Membrane Proteins in Milk Extraction Conditions Optimization and Enzymolysis Products Antioxidant Activity Research. Mater. Sci. Eng. C. 2016, 205–288. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Guan, K.; Liu, M.; Liu, D.; Wang, W. Novel Antioxidant Peptides from MFGM Protein Hydrolysates: Separation, Identification and Effects on Dexamethasone-Induced Mitochondrial Dysfunction. Food Chem. 2023, 403, 134473. [Google Scholar] [CrossRef]
- Chen, X.; Liang, D.; Huang, Z.; Jia, G.; Zhao, H.; Liu, G. Anti-Fatigue Effect of Quercetin on Enhancing Muscle Function and Antioxidant Capacity. J. Food Biochem. 2021, 45, e13968. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zeng, H.; Lin, S.; Zhang, Z.; Zhang, Y.; Hu, J. Anti-Fatigue Activities of Hairtail (Trichiurus Lepturus) Hydrolysate in an Endurance Swimming Mice Model. J. Funct. Foods 2020, 74, 104207. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, R.; Hua, H.; Cheng, Y.; Guo, Y.; Qian, H.; Du, P. The Macamide Relieves Fatigue by Acting as Inhibitor of Inflammatory Response in Exercising Mice: From Central to Peripheral. Eur. J. Pharmacol. 2022, 917, 174758. [Google Scholar] [CrossRef]
- Oh, S.; Komine, S.; Warabi, E.; Akiyama, K.; Ishii, A.; Ishige, K.; Mizokami, Y.; Kuga, K.; Horie, M.; Miwa, Y.; et al. Nuclear Factor (Erythroid Derived 2)-like 2 Activation Increases Exercise Endurance Capacity via Redox Modulation in Skeletal Muscles. Sci. Rep. 2017, 7, 12902. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Shi, J.; Zhong, H.; Abdullah; Zhuang, J.; Zhang, J.; Wang, J.; Zhang, X.; Feng, F. High-Degree Hydrolysis Sea Cucumber Peptides Improve Exercise Performance and Exert Antifatigue Effect via Activating the NRF2 and AMPK Signaling Pathways in Mice. J. Funct. Foods 2021, 86, 104677. [Google Scholar] [CrossRef]
- Motohashi, H.; Yamamoto, M. Nrf2–Keap1 Defines a Physiologically Important Stress Response Mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Gu, Y.; Yang, Y.; Cao, X.; Zhao, Y.; Gao, X.; Sun, C.; Zhang, F.; Yuan, Y.; Xu, Y.; Zhang, J.; et al. Plin3 Protects against Alcoholic Liver Injury by Facilitating Lipid Export from the Endoplasmic Reticulum. J. Cell. Biochem. 2019, 120, 16075–16087. [Google Scholar] [CrossRef]
- Ott, M.; Gogvadze, V.; Orrenius, S.; Zhivotovsky, B. Mitochondria, Oxidative Stress and Cell Death. Apoptosis 2007, 12, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Su, L.; Ye, Q.; Zhang, S.; Kung, H.; Jiang, F.; Jiang, G.; Miao, J.; Zhao, B. Discovery of a Novel Nrf2 Inhibitor That Induces Apoptosis of Human Acute Myeloid Leukemia Cells. Oncotarget 2017, 8, 7625–7636. [Google Scholar] [CrossRef]
- Ma, C.; Deng, Y.; Xiao, R.; Xu, F.; Li, M.; Gong, Q.; Gao, J. Anti-Fatigue Effect of Phlorizin on Exhaustive Exercise-Induced Oxidative Injury Mediated by Nrf2/ARE Signaling Pathway in Mice. Eur. J. Pharmacol. 2022, 918, 174563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, F. Reactive Oxygen Species (ROS), Troublemakers between Nuclear Factor-ΚB (NF-ΚB) and c-Jun NH2-Terminal Kinase (JNK). Cancer Res. 2004, 64, 1902–1905. [Google Scholar] [CrossRef] [PubMed]
- Digaleh, H.; Kiaei, M.; Khodagholi, F. Nrf2 and Nrf1 Signaling and ER Stress Crosstalk: Implication for Proteasomal Degradation and Autophagy. Cell. Mol. Life Sci. 2013, 70, 4681–4694. [Google Scholar] [CrossRef]
- Yu, C.; Minemoto, Y.; Zhang, J.; Liu, J.; Tang, F.; Bui, T.N.; Xiang, J.; Lin, A. JNK Suppresses Apoptosis via Phosphorylation of the Proapoptotic Bcl-2 Family Protein BAD. Mol. Cell 2004, 13, 329–340. [Google Scholar] [CrossRef]
- Varì, R.; Scazzocchio, B.; Santangelo, C.; Filesi, C.; Galvano, F.; D’Archivio, M.; Masella, R.; Giovannini, C. Protocatechuic Acid Prevents OxLDL-Induced Apoptosis by Activating JNK/Nrf2 Survival Signals in Macrophages. Oxid. Med. Cell. Longev. 2015, 2015, 351827. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Lu, Y.; Wang, H.; Feng, Y.; Jiang, S.; Gao, X.-H.; Qi, R.; Wu, Y.; Chen, H.-D. Paeoniflorin Resists H2O2-Induced Oxidative Stress in Melanocytes by JNK/Nrf2/HO-1 Pathway. Front. Pharmacol. 2020, 11, 536. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Oh, H.-J.; Cho, S.; Lee, O.-H.; Lee, B.-Y. Okra (Abelmoschus Esculentus L. Moench) Prevents Obesity by Reducing Lipid Accumulation and Increasing White Adipose Browning in High-Fat Diet-Fed Mice. Food Funct. 2022, 33, 11840–11852. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Inoue, R.; Tsukahara, T.; Ushida, K.; Chiji, H.; Matsubara, N.; Hara, H. Voluntary Running Exercise Alters Microbiota Composition and Increases N-Butyrate Concentration in the Rat Cecum. Biosci. Biotechnol. Biochem. 2008, 72, 572–576. [Google Scholar] [CrossRef]
- Rajili, M. The First 1000 Cultured Species of the Human Gastrointestinal Microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar]
- Samuel, B.S.; Shaito, A.; Motoike, T.; Rey, F.E.; Backhed, F.; Manchester, J.K.; Hammer, R.E.; Williams, S.C.; Crowley, J.; Yanagisawa, M.; et al. Effects of the Gut Microbiota on Host Adiposity Are Modulated by the Short-Chain Fatty-Acid Binding G Protein-Coupled Receptor, Gpr41. Proc. Natl. Acad. Sci. USA 2008, 105, 16767–16772. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, Y.; Chen, Z.; Zou, C.; Liu, W.; Yang, L.; Fu, L.; Wang, Y.; Liu, G.-M.; Cao, M.-J. Depolymerized Sulfated Galactans from Eucheuma Serra Ameliorate Allergic Response and Intestinal Flora in Food Allergic Mouse Model. Int. J. Biol. Macromol. 2021, 166, 977–985. [Google Scholar] [CrossRef]
- Wexler, H.M. Bacteroides: The Good, the Bad, and the Nitty-Gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef]
- Ghosh, S.; Dai, C.; Brown, K.; Rajendiran, E.; Makarenko, S.; Baker, J.; Ma, C.; Halder, S.; Montero, M.; Ionescu, V.A.; et al. Colonic Microbiota Alters Host Susceptibility to Infectious Colitis by Modulating Inflammation, Redox Status, and Ion Transporter Gene Expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G39–G49. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Wei, L.; Chiu, Y.-S.; Hsu, Y.-J.; Tsai, T.-Y.; Wang, M.-F.; Huang, C.-C. Lactobacillus Plantarum TWK10 Supplementation Improves Exercise Performance and Increases Muscle Mass in Mice. Nutrients 2016, 8, 205. [Google Scholar] [CrossRef]
- Shao, C.; Ren, Y.; Wang, Z.; Kang, C.; Jiang, H.; Chi, A. Detection of Urine Metabolites in a Rat Model of Chronic Fatigue Syndrome before and after Exercise. BioMed Res. Int. 2017, 2017, 8182020. [Google Scholar] [CrossRef] [PubMed]
- Cowart, L.A. A Novel Role for Sphingolipid Metabolism in Oxidant-Mediated Skeletal Muscle Fatigue. Focus on “Sphingomyelinase Stimulates Oxidant Signaling to Weaken Skeletal Muscle and Promote Fatigue”. Am. J. Physiol. Cell Physiol. 2010, 299, C549–C551. [Google Scholar] [CrossRef]
- He, P.; Chen, L.; Qin, X.; Du, G.; Li, Z. Astragali Radix–Codonopsis Radix–Jujubae Fructus Water Extracts Ameliorate Exercise-induced Fatigue in Mice via Modulating Gut Microbiota and Its Metabolites. J. Sci. Food Agric. 2022, 102, 5141–5152. [Google Scholar] [CrossRef]
- Meneguello, M.O.; Mendonça, J.R.; Lancha, A.H.; Costa Rosa, L.F.B.P. Effect of Arginine, Ornithine and Citrulline Supplementation upon Performance and Metabolism of Trained Rats. Cell Biochem. Funct. 2003, 21, 85–91. [Google Scholar] [CrossRef] [PubMed]
- van de Lagemaat, E.; de Groot, L.; van den Heuvel, E. Vitamin B12 in Relation to Oxidative Stress: A Systematic Review. Nutrients 2019, 11, 482. [Google Scholar] [CrossRef] [PubMed]
- Omotoso, G.O.; Abdulsalam, F.A.; Mutholib, N.Y.; Bature, A.I.; Gbadamosi, I.T. Cortico-Hippocampal Morphology and Behavioural Indices Improved in Maternal Deprivation Model of Schizophrenia Following Vitamin B Complex Supplementation. Neurol. Psychiatry Brain Res. 2020, 38, 74–82. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content | Proportion (%) |
|---|---|
| Phosphatidylcholine (PC) | 1.97 |
| Sphingomyelin (SM) | 1.86 |
| Phosphatidylethanolamine (PE) | 2.61 |
| Phosphatidylinositol (PI) | 0.14 |
| Phosphatidylserine (PS) | 0.88 |
| Total | 7.46 |
| SOD | CAT | GSH-Px | MDA | |
|---|---|---|---|---|
| (U/mL) | (U/mL) | (μmol/L) | (nmol/mL) | |
| Control | 84.92 ± 6.20 | 22.61 ± 5.39 | 145.33 ± 15.40 | 11.40 ± 3.50 |
| MFGM | 90.62 ± 3.89 * | 42.32 ± 11.36 * | 166.28 ± 14.10 * | 6.07 ± 1.66 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, X.; Yokoyama, W.; Liu, X.; Wang, K.; Hong, H.; Luo, Y.; Tan, Y. Milk Fat Globule Membrane Relieves Fatigue via Regulation of Oxidative Stress and Gut Microbiota in BALB/c Mice. Antioxidants 2023, 12, 712. https://doi.org/10.3390/antiox12030712
Zou X, Yokoyama W, Liu X, Wang K, Hong H, Luo Y, Tan Y. Milk Fat Globule Membrane Relieves Fatigue via Regulation of Oxidative Stress and Gut Microbiota in BALB/c Mice. Antioxidants. 2023; 12(3):712. https://doi.org/10.3390/antiox12030712
Chicago/Turabian StyleZou, Xiaoxiao, Wallace Yokoyama, Xiaohui Liu, Kai Wang, Hui Hong, Yongkang Luo, and Yuqing Tan. 2023. "Milk Fat Globule Membrane Relieves Fatigue via Regulation of Oxidative Stress and Gut Microbiota in BALB/c Mice" Antioxidants 12, no. 3: 712. https://doi.org/10.3390/antiox12030712