
Phytochemicals and Biological Activities of Walnut Septum: A Systematic Review

Abstract
:1. Introduction
2. Search Methodology
2.1. Eligibility Criteria
2.2. Information Sources
2.3. Search Strategy
2.4. Selection Process
2.5. Data Items
3. Phytochemical Composition
3.1. Phenolic Compounds
3.2. Lipidic Compounds
3.3. Carbohydrate Compounds
3.4. Other Compounds
4. Biological Activities
4.1. Antioxidant Activities
4.2. Anti-Inflammatory Effects
4.3. Antidiabetic Activities
4.4. Antimicrobial Properties
4.5. Enzyme Inhibitory Activities
4.6. Antitumor Properties
4.7. Antitussive Effect
4.8. Myelopoiesis Activation
4.9. Anti-Aging Potential
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salem, M.A.; Aborehab, N.M.; Al-Karmalawy, A.A.; Fernie, A.R.; Alseekh, S.; Ezzat, S.M. Potential Valorization of Edible Nuts By-Products: Exploring the Immune-Modulatory and Antioxidants Effects of Selected Nut Shells Extracts in Relation to Their Metabolic Profiles. Antioxidants 2022, 11, 462. [Google Scholar] [CrossRef]
- Slavin, J.L.; Lloyd, B. Health Benefits of Fruits and Vegetables. Adv. Nutr. 2012, 3, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Guasch-Ferré, M.; Tessier, A.-J.; Petersen, K.S.; Sapp, P.A.; Tapsell, L.C.; Salas-Salvadó, J.; Ros, E.; Kris-Etherton, P.M. Effects of Nut Consumption on Blood Lipids and Lipoproteins: A Comprehensive Literature Update. Nutrients 2023, 15, 596. [Google Scholar] [CrossRef]
- Cena, H.; Calder, P.C. Defining a Healthy Diet: Evidence for the Role of Contemporary Dietary Patterns in Health and Disease. Nutrients 2020, 12, 334. [Google Scholar] [CrossRef] [Green Version]
- Nishi, S.K.; Viguiliouk, E.; Kendall, C.W.C.; Jenkins, D.J.A.; Hu, F.B.; Sievenpiper, J.L.; Atzeni, A.; Misra, A.; Salas-Salvadó, J. Nuts in the Prevention and Management of Type 2 Diabetes. Nutrients 2023, 15, 878. [Google Scholar] [CrossRef]
- Jahanban-Esfahlan, A.; Ostadrahimi, A.; Tabibiazar, M.; Amarowicz, R. A Comparative Review on the Extraction, Antioxidant Content and Antioxidant Potential of Different Parts of Walnut (Juglans regia L.) Fruit and Tree. Molecules 2019, 24, 2133. [Google Scholar] [CrossRef] [Green Version]
- Geng, S.; Ning, D.; Ma, T.; Chen, H.; Zhang, Y.; Sun, X. Comprehensive Analysis of the Components of Walnut Kernel (Juglans regia L.) in China. J. Food Qual. 2021, 2021, 9302181. [Google Scholar] [CrossRef]
- Martínez, M.L.; Labuckas, D.O.; Lamarque, A.L.; Maestri, D.M. Walnut (Juglans regia L.): Genetic Resources, Chemistry, by-Products. J. Sci. Food Agric. 2010, 90, 1959–1967. [Google Scholar] [CrossRef]
- Yang, H.; Xiao, X.; Li, J.; Wang, F.; Mi, J.; Shi, Y.; He, F.; Chen, L.; Zhang, F.; Wan, X. Chemical Compositions of Walnut (Juglans spp.) Oil: Combined Effects of Genetic and Climatic Factors. Forests 2022, 13, 962. [Google Scholar] [CrossRef]
- Santos, A.; Barros, L.; Calhelha, R.C.; Dueñas, M.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Leaves and Decoction of Juglans regia L.: Different Performances Regarding Bioactive Compounds and In Vitro Antioxidant and Antitumor Effects. Ind. Crops Prod. 2013, 51, 430–436. [Google Scholar] [CrossRef]
- Chamorro, F.; Carpena, M.; Lourenço-Lopes, C.; Taofiq, O.; Otero, P.; Cao, H.; Xiao, J.; Simal-Gandara, J.; Prieto, M.A. By-Products of Walnut (Juglans regia) as a Source of Bioactive Compounds for the Formulation of Nutraceuticals and Functional Foods. Biol. Life Sci. Forum 2022, 12, 35. [Google Scholar] [CrossRef]
- Rusu, M.E.; Fizesan, I.; Pop, A.; Mocan, A.; Gheldiu, A.M.; Babota, M.; Vodnar, D.C.; Jurj, A.; Berindan-Neagoe, I.; Vlase, L.; et al. Walnut (Juglans regia L.) Septum: Assessment of Bioactive Molecules and In Vitro Biological Effects. Molecules 2020, 25, 2187. [Google Scholar] [CrossRef] [PubMed]
- Ramishvili, L.; Gordeziani, M.; Tavdishvili, E.; Bedineishvili, N.; Dzidziguri, D.; Kotrikadze, N. The Effect of Extract of Greek Walnut (Juglans regia L.) Septa on Some Function Characteristics of Erytrocytes. Georgian Med. News 2016, 261, 51–57. [Google Scholar]
- Dehghani, F.; Mashhoody, T.; Panjehshahin, M. Effect of Aqueous Extract of Walnut Septum on Blood Glucose and Pancreatic Structure in Streptozotocin-Induced Diabetic Mouse. Iran. J. Pharmacol. Ther. 2012, 11, 10–14. [Google Scholar]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 Explanation and Elaboration: Updated Guidance and Exemplars for Reporting Systematic Reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef] [PubMed]
- Hajikhani, R.; Solati, J. Effects of Walnut Alcoholic Extract (Juglans regia L.) Septum on Serum Glucose, Insulin and Activities of Aminotransferase Enzymes. J. Appl. Chem. Res. 2010, 12, 7–16. [Google Scholar]
- Dzidziguri, D.; Rukhadze, M.; Modebadze, I.; Bakuradze, E.; Kurtanidze, M.; Giqoshvili, V. The Study of the Immune Corrective Properties of Greek Walnut (Juglans regia L.) Septa on the Experimental Model of Leukopenia. Georgian Med. News 2016, 252, 84–89. [Google Scholar]
- Ghiravani, Z.; Hosseini, M.; Hassanzadeh Taheri, M.M.; Reza Abedini, M. Evaluation of Hypoglycemic and Hypolipidemic Effects of Internal Septum of Walnut Fruit in Alloxan-Induced Diabetic Rats. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 94–100. [Google Scholar] [CrossRef]
- Ravanbakhsh, A.; Mahdavi, M.; Jalilzade-Amin, G.; Javadi, S.; Maham, M.; Mohammadnejad, D.; Rashidi, M.R. Acute and Subchronic Toxicity Study of the Median Septum of Juglans regia in Wistar Rats. Adv. Pharm. Bull. 2016, 6, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Bai, H.; Jing, Y.; Li, W.; Yin, S.; Zhou, H. A Pair of Taxifolin-3-O-Arabinofuranoside Isomers from Juglans regia L. Nat. Prod. Res. 2017, 31, 945–950. [Google Scholar] [CrossRef]
- Meng, Q.; Li, Y.; Xiao, T.; Zhang, L.; Xu, D. Antioxidant and Antibacterial Activities of Polysaccharides Isolated and Purified from Diaphragma juglandis Fructus. Int. J. Biol. Macromol. 2017, 105, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Mu, Y.; Dong, H.; Yan, H.; Hao, C.; Wang, X.; Zhang, L. Chemical Constituents of the Ethyl Acetate Extract from Diaphragma juglandis Fructus and Their Inhibitory Activity on Nitric Oxide Production In Vitro. Molecules 2018, 23, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.; Wang, Y.; Chen, F.; Xiao, T.; Zhang, L. Polysaccharides from Diaphragma Juglandis Fructus: Extraction Optimization, Antitumor, and Immune-Enhancement Effects. Int. J. Biol. Macromol. 2018, 115, 835–845. [Google Scholar] [CrossRef]
- Rusu, M.E.; Gheldiu, A.M.; Mocan, A.; Moldovan, C.; Popa, D.S.; Tomuta, I.; Vlase, L. Process Optimization for Improved Phenolic Compounds Recovery from Walnut (Juglans regia L.) Septum: Phytochemical Profile and Biological Activities. Molecules 2018, 23, 2814. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Liu, J.; Li, J.; Liu, H.; Dong, N.; Geng, Y.-Y.; Lu, Y.; Wang, Y. Phenolic Composition and Nutritional Attributes of Diaphragma Juglandis Fructus and Shell of Walnut (Juglans regia L.). Food Sci. Biotechnol. 2020, 29, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, L.; Song, L.; Sun, X.; Yan, S.; Huang, W. Characterisation of Phenolics in Fruit Septum of Juglans regia Linn. by Ultra Performance Liquid Chromatography Coupled with Orbitrap Mass Spectrometer. Food Chem. 2019, 286, 669–677. [Google Scholar] [CrossRef]
- Liu, R.; Zhao, Z.; Dai, S.; Che, X.; Liu, W. Identification and Quantification of Bioactive Compounds in Diaphragma juglandis Fructus by UHPLC-Q-Orbitrap HRMS and UHPLC-MS/MS. J. Agric. Food Chem. 2019, 67, 3811–3825. [Google Scholar] [CrossRef]
- Mehdizadeh, T.; Mohammadipour, N.; Langroodi, A.M.; Raeisi, M. Effect of Walnut Kernel Septum Membranes Hydroalcoholic Extract on the Shelf Life of Traditional Butter. Heliyon 2019, 5, e01296. [Google Scholar] [CrossRef] [Green Version]
- Meng, Q.; Chen, F.; Xiao, T.; Zhang, L. Inhibitory Effects of Polysaccharide from Diaphragma juglandis Fructus on α-Amylase and α-D-Glucosidase Activity, Streptozotocin-Induced Hyperglycemia Model, Advanced Glycation End-Products Formation, and H2O2-Induced Oxidative Damage. Int. J. Biol. Macromol. 2019, 124, 1080–1089. [Google Scholar] [CrossRef]
- Genovese, C.; Cambria, M.T.; D’Angeli, F.; Addamo, A.P.; Malfa, G.A.; Siracusa, L.; Pulvirenti, L.; Anfuso, C.D.; Lupo, G.; Salmeri, M. The Double Effect of Walnut Septum Extract (Juglans regia L.) Counteracts A172 Glioblastoma Cell Survival and Bacterial Growth. Int. J. Oncol. 2020, 57, 1129–1144. [Google Scholar] [CrossRef]
- Rusu, M.E.; Georgiu, C.; Pop, A.; Mocan, A.; Kiss, B.; Vostinaru, O.; Fizesan, I.; Stefan, M.G.; Gheldiu, A.M.; Mates, L.; et al. Antioxidant Effects of Walnut (Juglans regia L.) Kernel and Walnut Septum Extract in a D-Galactose-Induced Aging Model and in Naturally Aged Rats. Antioxidants 2020, 9, 424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.G.; Kan, H.; Chen, S.X.; Thakur, K.; Wang, S.; Zhang, J.G.; Shang, Y.F.; Wei, Z.J. Comparison of Phenolic Compounds Extracted from Diaphragma Juglandis Fructus, Walnut Pellicle, and Flowers of Juglans regia Using Methanol, Ultrasonic Wave, and Enzyme Assisted-Extraction. Food Chem. 2020, 321, 126672. [Google Scholar] [CrossRef] [PubMed]
- Aghapour, S.K.F.; Sisakhtnezhad, S. Effect of the Internal Septum Extract of the Walnut Kernel on the Mesenchymal Stem Cells Cycle and MSCs-Derived Insulin-Producing Β-Cells Differentiation and Glucose Uptake. Jentashapir J. Cell. Mol. Biol. 2021, 12, e115014. [Google Scholar] [CrossRef]
- Hong, Q.; Geng, S.; Ji, J.; Ye, Y.; Xu, D.; Zhang, Y.; Sun, X. Separation and Identification of Antioxidant Chemical Components in Diaphragma juglandis Fructus and Functional Evaluation in Caenorhabditis elegans. J. Funct. Foods 2021, 80, 104422. [Google Scholar] [CrossRef]
- Hu, G.; Gao, S.; Mou, D. Water and Alcohol Extracts from Diaphragma juglandis on Anti-Fatigue and Antioxidative Effects in Vitro and Vivo. J. Sci. Food Agric. 2021, 101, 3132–3139. [Google Scholar] [CrossRef] [PubMed]
- Fizeșan, I.; Rusu, M.E.; Georgiu, C.; Pop, A.; Ștefan, M.G.; Muntean, D.M.; Mirel, S.; Vostinaru, O.; Kiss, B.; Popa, D.S. Antitussive, Antioxidant, and Anti-Inflammatory Effects of a Walnut (Juglans regia L.) Septum Extract Rich in Bioactive Compounds. Antioxidants 2021, 10, 119. [Google Scholar] [CrossRef] [PubMed]
- Kalogiouri, N.P.; Samanidou, V.F. A Validated Ultrasound-Assisted Extraction Coupled with Spe-Hplc-Dad for the Determination of Flavonoids in by-Products of Plant Origin: An Application Study for the Valorization of the Walnut Septum Membrane. Molecules 2021, 26, 6418. [Google Scholar] [CrossRef]
- Tan, J.; Cheng, Y.; Wang, S.; Li, J.; Ren, H.; Qiao, Y.; Li, Q.; Wang, Y. The Chemical Constituents of Diaphragma Juglandis Fructus and Their Inhibitory Effect on α-Glucosidase Activity. Molecules 2022, 27, 3047. [Google Scholar] [CrossRef]
- Tan, J.Y.; Cheng, Y.G.; Li, J.L.; Ren, H.Q.; Li, H.; Huang, Y.R.; Qiao, Y.B.; Li, Q.S.; Wang, Y.L. New Taraxasterane-Type Triterpenes from Diaphragma juglandis Fructus. Tetrahedron Lett. 2022, 100, 153868. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, L.; Li, Q.; Gao, Y.; Yu, X. Utilization of Diaphragma Juglandis Extract as a Natural Antioxidant for Improving the Oxidative Stability of Soybean Oil during Deep Frying. Food Chem. X 2022, 14, 100359. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Sayed, A.M.; Elmaidomy, A.H. Natural Products’ Extraction and Isolation-between Conventional and Modern Techniques. Front. Nat. Prod. 2022, 1. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Schädel, C.; Blöchl, A.; Richter, A.; Hoch, G. Quantification and Monosaccharide Composition of Hemicelluloses from Different Plant Functional Types. Plant Physiol. Biochem. 2010, 48, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cheung, E.C.; Vousden, K.H. The Role of ROS in Tumour Development and Progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, A.J.P.O.; de Oliveira, J.C.P.L.; da Silva Pontes, L.V.; de Souza Júnior, J.F.; Gonçalves, T.A.F.; Dantas, S.H.; de Almeida Feitosa, M.S.; Silva, A.O.; de Medeiros, I.A. ROS: Basic Concepts, Sources, Cellular Signaling, and Its Implications in Aging Pathways. Oxid. Med. Cell Longev. 2022, 2022, 1225578. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Banc, R.; Rusu, M.E.; Filip, L.; Popa, D.-S. The Impact of Ellagitannins and Their Metabolites through Gut Microbiome on the Gut Health and Brain Wellness within the Gut–Brain Axis. Foods 2023, 12, 270. [Google Scholar] [CrossRef]
- Merly, L.; Smith, S.L. Murine RAW 264.7 Cell Line as an Immune Target: Are We Missing Something? Immunopharmacol. Immunotoxicol. 2017, 39, 55–58. [Google Scholar] [CrossRef]
- Bourais, I.; Elmarrkechy, S.; Taha, D.; Badaoui, B.; Mourabit, Y.; Salhi, N.; Alshahrani, M.M.; al Awadh, A.A.; Bouyahya, A.; Goh, K.W.; et al. Comparative Investigation of Chemical Constituents of Kernels, Leaves, Husk, and Bark of Juglans regia L., Using HPLC-DAD-ESI-MS/MS Analysis and Evaluation of Their Antioxidant, Antidiabetic, and Anti-Inflammatory Activities. Molecules 2022, 27, 8989. [Google Scholar] [CrossRef]
- Fisher, D.; Cahoon, D.; Shukitt-Hale, B. Walnut Oil and Blueberry Treatments Have Beneficial Individual but Not Synergistic Effects on Neuroinflammation In Vitro. Am. Soc. Nutr. 2021, 5, 904. [Google Scholar] [CrossRef]
- Davies, M.J.; Aroda, V.R.; Collins, B.S.; Gabbay, R.A.; Green, J.; Maruthur, N.M.; Rosas, S.E.; del Prato, S.; Mathieu, C.; Mingrone, G.; et al. Management of Hyperglycaemia in Type 2 Diabetes, 2022. A Consensus Report by the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetologia 2022, 65, 1925–1966. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, R.; Aberle, J.; Birkenfeld, A.L.; Gallwitz, B.; Kellerer, M.; Klein, H.; Müller-Wieland, D.; Nauck, M.A.; Reuter, H.M.; Siegel, E. Therapy of Type 2 Diabetes. Exp. Clin. Endocrinol. Diabetes 2019, 127, S73–S92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akpoveso, O.-O.P.; Ubah, E.E.; Obasanmi, G. Antioxidant Phytochemicals as Potential Therapy for Diabetic Complications. Antioxidants 2023, 12, 123. [Google Scholar] [CrossRef]
- Zangeneh, A.; Zangeneh, M.M.; Goodarzi, N.; Najafi, F.; Hagh Nazari, L. Protective Effects of Aqueous Extract of Internal Septum of Walnut Fruit on Diabetic Hepatopathy in Streptozocin-Induced Diabetic Mice. Sci. J. Kurd. Univ. Med. Sci. 2018, 93, 4–14. [Google Scholar]
- Ghiravani, Z.; Hassanzadeh-Taheri, M.; Hassanzadeh-Taheri, M.; Hosseini, M. Internal Septum of Walnut Kernel: A Neglected Functional Food. Res. J. Pharmacogn. (RJP) 2020, 7, 81–92. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Zhao, C.; Sui, H.; Feng Li, C.; Zhong, L.; Zhou, Q.; Bai, Y.; An, S.; Du, X.; et al. Green Walnut Husk Extracts Proliferation and Migration in Gastric Cancer. J. Cancer 2022, 13, 1130–1144. [Google Scholar] [CrossRef]
- Boulfia, M.; Lamchouri, F.; Toufik, H. Mineral Analysis, in Vitro Evaluation of Alpha-Amylase, Alpha-Glucosidase, and Beta-Galactosidase Inhibition, and Antibacterial Activities of Juglans regia L. Bark Extracts. BioMed Res. Int. 2021, 2021, 1585692. [Google Scholar] [CrossRef]
- Medic, A.; Jakopic, J.; Solar, A.; Hudina, M.; Veberic, R. Walnut (J. regia) Agro-Residues as a Rich Source of Phenolic Compounds. Biology 2021, 10, 535. [Google Scholar] [CrossRef]
- Sun, Y.; Li, S.; Zeng, F.; Qi, J.; Qin, W.; Tan, C.; Luo, Q.; Wu, D.; Zhang, Q.; Lin, D.; et al. Functional Components, Antioxidant Activity and Hypoglycemic Ability Following Simulated Gastro-Intestinal Digestion of Pigments from Walnut Brown Shell and Green Husk. Antioxidants 2019, 8, 573. [Google Scholar] [CrossRef] [Green Version]
- Jelodar, G.; Mohammadi, M.; Akbari, A.; Nazifi, S. Cyclohexane Extract of Walnut Leaves Improves Indices of Oxidative Stress, Total Homocysteine and Lipids Profiles in Streptozotocin-Induced Diabetic Rats. Physiol. Rep. 2020, 8, e14348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabiei, K.; Ebrahimzadeh, M.A.; Saeedi, M.; Bahar, A.; Akha, O.; Kashi, Z. Effects of a Hydroalcoholic Extract of Juglans regia (Walnut) Leaves on Blood Glucose and Major Cardiovascular Risk Factors in Type 2 Diabetic Patients: A Double-Blind, Placebo-Controlled Clinical Trial. BMC Complement. Altern. Med. 2018, 18, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Vedeanu, N.; Voica, C.; Magdas, D.A.; Kiss, B.; Stefan, M.G.; Simedrea, R.; Georgiu, C.; Berce, C.; Vostinaru, O.; Boros, R.; et al. Subacute Co-Exposure to Low Doses of Ruthenium(III) Changes the Distribution, Excretion and Biological Effects of Silver Ions in Rats. Environ. Chem. 2020, 17, 163–172. [Google Scholar] [CrossRef]
- Rusu, M.E.; Simedrea, R.; Gheldiu, A.M.; Mocan, A.; Vlase, L.; Popa, D.S.; Ferreira, I.C.F.R. Benefits of Tree Nut Consumption on Aging and Age-Related Diseases: Mechanisms of Actions. Trends Food Sci. Technol. 2019, 88, 104–120. [Google Scholar] [CrossRef]
- Vieira, V.; Pereira, C.; Abreu, R.M.V.; Calhelha, R.C.; Alves, M.J.; Coutinho, J.A.P.; Ferreira, O.; Barros, L.; Ferreira, I.C.F.R. Hydroethanolic Extract of Juglans regia L. Green Husks: A Source of Bioactive Phytochemicals. Food Chem. Toxicol. 2020, 137, 111189. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadi, S.; Moghadam, H.N.; Mahdavi-Ourtakand, M. Evaluating the Anti-Biofilm and Antibacterial Effects of Juglans regia L. Extracts against Clinical Isolates of Pseudomonas Aeruginosa. Microb. Pathog. 2018, 118, 285–289. [Google Scholar] [CrossRef]
- Acquaviva, R.; D’Angeli, F.; Malfa, G.A.; Ronsisvalle, S.; Garozzo, A.; Stivala, A.; Ragusa, S.; Nicolosi, D.; Salmeri, M.; Genovese, C. Antibacterial and Anti-Biofilm Activities of Walnut Pellicle Extract (Juglans regia L.) against Coagulase-Negative Staphylococci. Nat. Prod. Res. 2021, 35, 2076–2081. [Google Scholar] [CrossRef]
- Barekat, S.; Nasirpour, A.; Keramat, J.; Dinari, M.; Meziane-Kaci, M.; Paris, C.; Desobry, S. Phytochemical Composition, Antimicrobial, Anticancer Properties, and Antioxidant Potential of Green Husk from Several Walnut Varieties (Juglans regia L.). Antioxidants 2022, 12, 52. [Google Scholar] [CrossRef]
- Soto-Madrid, D.; Gutiérrez-Cutiño, M.; Pozo-Martínez, J.; Zúñiga-López, M.C.; Olea-Azar, C.; Matiacevich, S. Dependence of the Ripeness Stage on the Antioxidant and Antimicrobial Properties of Walnut (Juglans regia L.) Green Husk Extracts from Industrial by-Products. Molecules 2021, 26, 2878. [Google Scholar] [CrossRef]
- Delibaş, E.A.Ö.; Kiray, E. Investigation of Antioxidant and Antimicrobial Activities of Walnut (Juglans regia L.) Kernel Septum. Eur. Res. J. 2022, 9, 87–96. [Google Scholar] [CrossRef]
- Kovacs, D.; Cardinali, G.; Picardo, M.; Bastonini, E. Shining Light on Autophagy in Skin Pigmentation and Pigmentary Disorders. Cells 2022, 11, 2999. [Google Scholar] [CrossRef] [PubMed]
- Besrour, N.; Oludemi, T.; Mandim, F.; Pereira, C.; Dias, M.I.; Soković, M.; Stojković, D.; Ferreira, O.; Ferreira, I.C.F.R.; Barros, L. Article Valorization of Juglans regia Leaves as Cosmeceutical Ingredients: Bioactivity Evaluation and Final Formulation Development. Antioxidants 2022, 11, 677. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Keogh, J.; Clifton, P.M. Nuts and Cardio-Metabolic Disease: A Review of Meta-Analyses. Nutrients 2018, 10, 1935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojdyło, A.; Turkiewicz, I.P.; Tkacz, K.; Nowicka, P.; Bobak, Ł. Nuts as Functional Foods: Variation of Nutritional and Phytochemical Profiles and Their In Vitro Bioactive Properties. Food Chem. X 2022, 15, 100418. [Google Scholar] [CrossRef]
- Tuzimski, T.; Petruczynik, A. Determination of Anti-Alzheimer’s Disease Activity of Selected Plant Ingredients. Molecules 2022, 27, 3222. [Google Scholar] [CrossRef] [PubMed]
- Amat-Ur-rasool, H.; Symes, F.; Tooth, D.; Schaffert, L.N.; Elmorsy, E.; Ahmed, M.; Hasnain, S.; Carter, W.G. Potential Nutraceutical Properties of Leaves from Several Commonly Cultivated Plants. Biomolecules 2020, 10, 1556. [Google Scholar] [CrossRef]
- Sharma, P.; Verma, P.K.; Sood, S.; Pankaj, N.K.; Agarwal, S.; Raina, R. Neuroprotective Potential of Hydroethanolic Hull Extract of Juglans regia L. on Isoprenaline Induced Oxidative Damage in Brain of Wistar Rats. Toxicol. Rep. 2021, 8, 223–229. [Google Scholar] [CrossRef]
- Li, Y.; Schoufour, J.; Wang, D.D.; Dhana, K.; Pan, A.; Liu, X.; Song, M.; Liu, G.; Shin, H.J.; Sun, Q.; et al. Healthy Lifestyle and Life Expectancy Free of Cancer, Cardiovascular Disease, and Type 2 Diabetes: Prospective Cohort Study. BMJ 2020, 368, l6669. [Google Scholar] [CrossRef] [Green Version]
- Rusu, M.E.; Fizeșan, I.; Vlase, L.; Popa, D.S. Antioxidants in Age-Related Diseases and Anti-Aging Strategies. Antioxidants 2022, 11, 1868. [Google Scholar] [CrossRef]
- Ros, E.; Singh, A.; O’keefe, J.H. Nuts: Natural Pleiotropic Nutraceuticals. Nutrients 2021, 13, 3269. [Google Scholar] [CrossRef]
- Rusu, M.E.; Mocan, A.; Ferreira, I.C.F.R.; Popa, D.S. Health Benefits of Nut Consumption in Middle-Aged and Elderly Population. Antioxidants 2019, 8, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardman, W.E.; Primerano, D.A.; Legenza, M.T.; Morgan, J.; Fan, J.; Denvir, J. Dietary Walnut Altered Gene Expressions Related to Tumor Growth, Survival, and Metastasis in Breast Cancer Patients: A Pilot Clinical Trial. Nutr. Res. 2019, 66, 82–94. [Google Scholar] [CrossRef]
- Tang, Y.T.; Li, Y.; Chu, P.; Ma, D.X.; Tang, Z.Y.; Sun, L.Z. Molecular Biological Mechanism of Action in Cancer Therapies: Juglone and Its Derivates, the Future of Development. Biomed. Pharmacother. 2022, 148, 112785. [Google Scholar] [CrossRef] [PubMed]
- Crocetto, F.; di Zazzo, E.; Buonerba, C.; Aveta, A.; Pandolfo, S.D.; Barone, B.; Trama, F.; Caputo, V.F.; Scafuri, L.; Ferro, M.; et al. Kaempferol, Myricetin and Fisetin in Prostate and Bladder Cancer: A Systematic Review of the Literature. Nutrients 2021, 13, 3750. [Google Scholar] [CrossRef] [PubMed]
- Asgharian, P.; Tazekand, A.P.; Hosseini, K.; Forouhandeh, H.; Ghasemnejad, T.; Ranjbar, M.; Hasan, M.; Kumar, M.; Beirami, S.M.; Tarhriz, V.; et al. Potential Mechanisms of Quercetin in Cancer Prevention: Focus on Cellular and Molecular Targets. Cancer Cell Int. 2022, 22, 257. [Google Scholar] [CrossRef] [PubMed]
- Vrânceanu, M.; Galimberti, D.; Banc, R.; Dragoş, O.; Cozma-Petruţ, A.; Hegheş, S.C.; Voştinaru, O.; Cuciureanu, M.; Stroia, C.M.; Miere, D.; et al. The Anticancer Potential of Plant-Derived Nutraceuticals via the Modulation of Gene Expression. Plants 2022, 11, 2524. [Google Scholar] [CrossRef] [PubMed]
- D’Angeli, F.; Malfa, G.A.; Garozzo, A.; Volti, G.L.; Genovese, C.; Stivala, A.; Nicolosi, D.; Attanasio, F.; Bellia, F.; Ronsisvalle, S.; et al. Antimicrobial, Antioxidant, and Cytotoxic Activities of Juglans regia L. Pellicle Extract. Antibiotics 2021, 10, 159. [Google Scholar] [CrossRef]
- Hu, J.R.; Jung, C.J.; Ku, S.M.; Jung, D.H.; Bashir, K.M.I.; Ku, S.K.; Choi, J.S. Anti-Inflammatory, Expectorant, and Antitussive Properties of Kyeongok-Go in ICR Mice. Pharm. Biol. 2021, 59, 321–334. [Google Scholar] [CrossRef]
- Bottoni, M.; Milani, F.; Colombo, L.; Nallio, K.; Colombo, P.S.; Giuliani, C.; Bruschi, P.; Fico, G. Using Medicinal Plants in Valmalenco (Italian Alps): From Tradition to Scientific Approaches. Molecules 2020, 25, 4144. [Google Scholar] [CrossRef]
- Papakyriakopoulou, P.; Velidakis, N.; Khattab, E.; Valsami, G.; Korakianitis, I.; Kadoglou, N.P.E. Potential Pharmaceutical Applications of Quercetin in Cardiovascular Diseases. Pharmaceuticals 2022, 15, 1019. [Google Scholar] [CrossRef]
- Maurya, P.K. Health Benefits of Quercetin in Age-Related Diseases. Molecules 2022, 27, 2498. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Wang, R.; Wang, M.; Meng, Z.; Wang, X.; Han, M.; Guo, Y.; Wang, X. Preparation of Naringenin Nanosuspension and Its Antitussive and Expectorant Effects. Molecules 2022, 27, 741. [Google Scholar] [CrossRef]
- Najafian, Y.; Hamedi, S.S.; Kaboli Farshchi, M.; Feyzabadi, Z. Plantago Major in Traditional Persian Medicine and Modern Phytotherapy: A Narrative Review. Electron Phys. 2018, 10, 6390–6399. [Google Scholar] [CrossRef] [Green Version]
- Steven, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Kuntic, M.; Jimenez, M.T.B.; Vujacic-Mirski, K.; Helmstädter, J.; Kröller-Schön, S.; Münzel, T.; et al. Vascular Inflammation and Oxidative Stress: Major Triggers for Cardiovascular Disease. Oxid. Med. Cell Longev. 2019, 2019, 7092151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, V.D.; Anderson, R.M. Nutrition, Longevity and Disease: From Molecular Mechanisms to Interventions. Cell 2022, 185, 1455–1470. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ref. | Country | Study Type | Study Purpose | Materials/Type of Extracts | Analysis Metods/Biological Systems/Animal Models | Study Outcomes/Biological Activities |
|---|---|---|---|---|---|---|
| [16] | Iran | In vivo | Hypoglicemiant effect of WSE | WSE—90% ethanol | Streptozotocin-induced diabetic rats (60 mg/kg bw) | WSE (gavage: 100, 200, 500, 1000 mg/kg bw, 2 weeks) significantly decreased the blood glucose level. |
| [14] | Iran | In vivo | Hypoglicemiant effect of WSE; Histopathological pancreatic structure | WSE—aqueous | Streptozotocin-induced diabetic mice (220 mg/kg bw) | WSE (oral; 200, 400, 600, 800 mg/kg bw, 4 weeks) reduced blood glucose level. No effect on pancreatic structure. |
| [17] | Georgia | In vivo | Immune corrective properties of WSE on leukopenia induced experimental model | WSE—aqueous (2:1, water:WS) | Cyclophosphamide—induced leukopenia mice (350 mg/kg bw) | WSE stimulates myelopoiesis in mice with leukopenia induced by chemotherapy. |
| [18] | Iran | In vivo | The antidiabetic and hypolipidemic effects of WSE on diabetic rats | WSE—1:10 w/v in 80% ethanol | Alloxan-induced diabetic rats (150 mg/kg bw) | WSE (oral: 200, 400 mg/kg bw, 4 weeks) significantly decreased blood glucose, ALAT, ASAT, TG; WSE (200 mg/kg bw) attenuated TC and LDL-C. |
| [13] | Georgia | In vivo | The effect of WSE on the functional characteristics of erythrocytes during the administration of cyclophosphamide | WSE—2:1 (water:WS) | Cyclophosphamide—cytotoxicity induced mice (350 mg/kg bw) | WSE had a preventive effect on the process of hemolysis; WSE increased sorption capacity and resistance of erythrocytes to lysis. |
| [19] | Iran | In vivo | Acute and subacute toxicity of WSE; Effect of WSE on oxidative stress and enzymes; Histopathology | WSE—95% methanol | Female Wistar rats treated with WSE (oral: 10, 100, 1000, 1600, 2900, and 5000 mg/kg bw and gavage: 1000 mg/kg bw, 4 weeks) | WSE showed no acute or subacute adverse effects with dose of 1000 mg/kg bw (oral by gavage). WSE may improve kidney structure and function. |
| [20] | China | In vitro | Isolation of taxifolin-3-O-arabinofuranoside isomers from WSE | WSE—70% ethanol | Compounds identification: silica gel column chromatography (CC), Sephadex LH-20 CC, ODS CC, and preparative HPLC | (2S,3S)-taxifolin-3-O-α-d-arabinofuranoside and its isomer (2S,3S)-taxifolin-3-O-α-l-arabinofuranoside were isolated from WSE. |
| [21] | China | In vitro | Antioxidant and antibacterial activities of polysaccharides from WS (WSP) | WSP—water-soluble, purified | Composition: HPIC; Antioxidant activity: DPPH, ABTS, -OH, FRAP. Antibacterial activity: on E. coli, P. aeruginosa, S. aureus, E. faecalis strains | WSP mainly contained glucose, galactose, arabinose, xylose, and traces of mannose. WSP showed antioxidant and antibacterial activities in vitro. |
| [22] | China | In vitro | Chemical constituents and anti-inflammatory effects of WSE | WSE—ethyl acetate extract; | Isolation: pre-HPLC, HSCCC; identification: NRM, ESI-MS; Lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages model (in vitro) | Fourteen compounds isolated; Dihydrophaseic acid, blumenol B and (4S)-4-hydroxy-1-tetralone for the first time in WS; Gallic acid, ethyl gallate. and (+)-dehydrovomifoliol—anti-inflammatory activity. |
| [23] | China | In vitro | Antitumor, and immune-enhancement effects of polysaccharides from WS (WSP) | WSP—water-soluble, purified (Water/raw material ratio: 20 mL/g) | Cytotoxicity (MTT assay) on Murine macrophage cell line RAW 264.7, HepG2, and BGC-823 cells. Immune-enhancement activity: cytokines production, RT and RT-PCR on RAW 264.7 cells | WSP could effectively suppress the proliferation of tumor cells. WSP could significantly enhance phagocytosis, stimulate the production of NO, TNF-α, and interleukins (IL-6 and IL-1β), and promote the mRNA expression levels of iNOS, TNF-α, IL-6, and IL-1β in a dose-dependent manner. |
| [24] | Romania | In vitro | Process optimization for improved phenolic compounds recovery from WS Phytochemical profile and biological activities | WSE—70% ethanol | Phytochemical profile: UTE, LC-MS/MS; Quantitative det.: TPC, TFC, CTC. Antioxidant activity: ABTS, DPPH, FRAP assays; Tyrosinase inhibitory effect | The content in phenolic compounds, tannins, and phytosterols was correlated with the evaluated antioxidant and tyrosinase inhibitory activities. |
| [25] | China | In vitro | Phenolic composition and nutritional attributes of WS | WS—dried under 35 °C in oven, grinded | Identification: minerals (ICP-MS), fatty acids (GC/MS), amino acids, monosaccharides, phenolic compounds (HPLC); Phenolic compounds extract: HPLC–DAD/ESI–MS | Phenolic acids, hydroxybenzoicacid, isoflavone, and flavone were identified in the free phenolic fractions (FPFs) of WS. |
| [26] | China | In vitro | Characterization of phenolics in WSE | WSE—70% methanol | Analysis of phenolic compounds: HPLC-UV; Individual phenolic compounds: UPLC-MS; TPC—Folin–Ciocalteau method | A total of 75 phenolic compounds were identified: flavonols, flavanols, procyanidins, ellagitannins, gallotannins, phenolic acids and related compounds |
| [27] | China | In vitro | Identification and quantification of bioactive compounds in WSE | WSE—acetone/cyclohexane (2:1, v/v) | Identification: hydrolyzable tannins, flavonoids, phenolic acids, quinones: UHPLC-Q-Orbitrap HRMS; Polyphenols quantification: UHPLC-MS/MS | Two-hundred compounds, including hydrolyzable tannins, flavonoids, phenolic acids, and quinones, were identified. Total contents of polyphenols: 2.88−6.18 mg/g. |
| [28] | Iran | In vitro | Antioxidant and antimicrobial effect of (WSE) in traditional butter | WSE—hydroalcoholic extract | Antioxidant effect: DPPH, reducing power, total phenol assays; Physicochemical analysis; Microbiological analysis; Sensory evaluation | The utilization of WSE is safe in the 0.05%, 0.1% and 0.5% conc. and has benefits for oxidative stability and inhibited the microbial growth of butter stored under suitable conditions. |
| [29] | China | In vitro; In vivo | The hemolysis inhibitory effects of DJP-2; Antioxidant effect of WSP (in vitro); Hypoglycemic activity (in vivo); Antiglycation activities. | WSP- water-soluble, purified | H2O2-induced hemolysis of RBCs from Kunming mice; Hepatic L02 cells model; Type 2 diabetes mellitus mice model (male ICR mice, ip. STZ 1%) Hypoglycemic assay (α-amylase and α-D-glucosidase activity) | WSP showed remarkable hemolysis inhibitory activity. WSP markedly weakened the oxidative damage induced by H2O2 in hepatic L02 cells via strengthening the cell viability. WSP showed hypoglycemic activities in vivo and in vitro. WSP (3 mg/mL) exerted more significant antiglycation activities than aminoguanidine. |
| [30] | Italy | In vitro | The cytostatic potential and antibacterial activity of WSE | WSE—96% ethanol | Human glioblastoma cells (A172). Clinical isolates: Gram-positive and Gram-negative strains | WSE (70 μg/mL) caused a significant reduction of healthy A172 cells and an increment of apoptotic and necrotic cells. WSE (275 μg/mL) affected the growth of G-pos and G-neg bacterial strains, resistant to Ciprofloxacin. |
| [31] | Romania | In vivo | Antioxidant effect of WSE in aged induced and old rats | WSE—water: acetone (50:50, v/v) | D-Galactose-induced aging model (Wistar female rats, D-gal 1200 mg/week), old rats; Cellular antioxidant status: ABTS, DPPH, FRAP; Oxidative stress biomarkers: OS, AGEs, NO, MDA, GSH; AChE activity; Histopathological and immunohistochemical analyses | A significant improvement in cellular antioxidant activity and decrease of ROS, advanced glycation end products, NO, MDA, or increase of glutathione after WSE intake. |
| [12] | Romania | In vitro | Assessment of bioactive molecules and in vitro biological effects of WS | WSE WS mixed with water/acetone (50:50, v/v) at a ratio of 1:10 (w/v) | Tocopherol content: LC-MS/MS; Biological activities: enzyme, cholinesterase, α-glucosidase, and lipase inhibitory activity; Antibacterial and antifungal activity: G. pos, G. neg. bacteria, and fungi strains; Antimutagenic assay: Salmonella typhimurium TA 98 and TA 100 strains; Cytocompatibility: cancerous A549 and T47D-KBluc cells; Antioxidant potential: ROS in A549, MCF-7 and HGF cell types; Anti-inflammatory potential in HGF cells | WSE has inhibitory effect on acetylcholinesterase, α-glucosidase, and lipase activity. WSE—strong antimicrobial potential against Staphylococcus aureus, Pseudomonas aeruginosa, and Salmonella enteritidis. Strong antimutagenic inhibitory effects against TA 98 and TA 100 strains. WSE—cytotoxic, antioxidant, and anti-inflammatory activity (IL-6, IL-8, IL-1β) in the selected cell lines. |
| [32] | China | In vitro | Comparison of phenolic compounds extracted from WS, walnut pellicle (WP), and flowers of Juglans regia (WFl) | ME—methanolic condensation reflux extraction UE—ultrasonic waves extraction EE—enzymes assisted-extraction | TPC—Folin-Ciocalteu method, FL, CT; Antioxidant activity: OH radical scavenging, DPPH, ABTS, Fe2+ chelating | A total of 50 phenolic compounds were identified by LC-MS/MS. The antioxidant capacity of WP was stronger than that of WFl and WS. |
| [33] | Iran | In vitro | The effect of the DISWK on the survival, TAC, and cell cycle of rat bone marrow. The expression of genes involved in pancreatic insulin-producing β-cell (IPCs) commitment and glucose uptake in MSCs-derived IPCs. | DISWK—aqueous | Survival, TAC, and cell cycle on rat bone marrow-derived mesenchymal stem cells (rBM-MSCs). The expression of genes involved in pancreatic insulin-producing β-cell (IPCs) commitment and glucose uptake in MSCs-derived IPCs | DISWK significantly reduced the viability of rBM-MSCs and decreased their TAC in long-term cultures. DISWK suppressed the rBM-MSCs cell cycle at S and G2 phases. DISWK significantly upregulated Ins1/2, Insr, and Glut1 genes and downregulated Pdx1 gene in the rBM-MSCs-derived IPCs. |
| [34] | China | In vivo | Separation and identification of antioxidant chemical components in WS and functional evaluation in Caenorhabditis elegans | WSE—ethanol and water | C. elegans—to evaluate antioxidant capacity and examined the changes in metabolome by GC-TOF/MS | Four monomers were isolated, three for the first time in WS CDG could improve oxidative stress and prolong lifespan in C. elegans by improving the activities of antioxidant enzymes and reducing ROS and MDA content. |
| [35] | China | In vitro; In vivo | Anti-fatigue and antioxidative effects of water and alcohol extracts from WS | WSE—water (1:20, w/v); WSE—70% ethanol | H2O2-induced oxidative stress in HepG2 cells; Adult male Kunming mice—behavioral tests and biochemical assay | Both extracts—HepG2 protective capacities; capable of scavenging DPPH and ABTS as VitC |
| [36] | Romania | In vivo | Antitussive, antioxidant, and anti-inflammatory effects of a WSE | WSE—water: acetone (50:50, v/v) | Citric acid aerosol-induced cough experimental model in rodents (Wistar male rats) | A significant antitussive effect of WSE, superior to codeine; The antioxidant and anti-inflammatory effects of WSE were confirmed by biochemical assays and histopathological analysis. |
| [37] | Greece | In vitro | Determination of flavonoids in by-products of plant origin: An application for the valorization of the WS | WS | A UAE-SPE-HPLC-DAD analytical method was developed and validated | Seven flavonoids were determined: catechin, rutin, myricetin, luteolin, quercetin, apigenin, and kaempferol. |
| [38] | China | In vitro | Chemical constituents of WS; The hypoglycemic effect of WSE | WSE—70% ethanol | Identification of constituents: LC–MS, HPLC Hypoglycemic assay—α-glucosidase activity (in vitro) | 37 constituents identified; the most abundant: rugosin F isomer, gallic acid, phlorizin, p-coumaric acid, vanillic acid, and quercetin. Flavonoids: the main bioactive substances contributing to the α-glucosidase inhibitory activity of WSE. |
| [39] | China | In vitro | Phytochemical investigation on WSE | WSE—70% ethanol | Identification of constituents: HRESIMS, NMR and X-ray single-crystal diffraction analysis; Molecular docking: α-glycosidase inhibitory activities | The isolation of four new taraxasterane-type triterpenes. The structure-activity relationship of the bioactive taraxasterane-type triterpenes against the α-glucosidase molecular docking was established. |
| [40] | China | In vitro | The effect of WSE on the oxidative stability of soybean oil during deep frying. | WSE—80% ethanol | TPC—Folin-Ciocalteu method; Quantification of bioactive compounds: Q Exactive Orbitrap mass spectrometer coupled to a Vanquish ultraperformance liquid chromatography system; Determination of oxidized and polymeric TAGs: HPSEC method; Quantification of oxidation products: 1H NMR | 31 polyphenols determined; the main components: catechin, quercitrin, taxifolin, quercetin 3-β-d-glucoside, epicatechin, gallic acid, and 3,4-dihydroxybenzoic acid. The antioxidants delayed the degradation of triglycerides and inhibited the increase in the contents of panisidine, oxidized triglyceride monomers, triglyceride dimers, and TG oligomers. WSE: better inhibitory effect on the formation of (E)-2-alkenals, (E,E)-2,4- alkadienals, 4-oxo-alkanals, primary alcohols, and secondary alcohols detected by 1H NMR TBHQ and TP |
| Ref. | Extract | Analytical Method | Compounds | Amount |
|---|---|---|---|---|
| [20] | 70% ethanolic extract | LC-ESI-MS; HPLC-UV | Flavonoids: (2S,3S)-Taxifolin-3-O-α-d-arabinofuranoside, (2S,3S)-Taxifolin-3-O-α-l-arabinofuranoside | NQ |
| [22] | Ethanolic extract | LC-ESI-MS; HPLC-UV | Phenolic acids: Gallic acid, p-Hydroxybenzoic acid, Protocatechuic acid, Vanillic acid | NA |
| Flavonoids: Quercitrin, Taxifolin (dihydroquercetin), Taxifolin-3-O-α-l-arabinofuranoside | NA | |||
| Esters: Ethyl gallate | NA | |||
| [24] | Extract in water:acetone (50:50, v/v) | LC-ESI-MS; HPLC-UV | Phenolic acids: p-Coumaric acid, Ferulic acid, Gentisic acid | <LOQ |
| Flavonoids: Hyperoside | 67.32 µg/g dw | |||
| Isoquercitrin | 103.60 µg/g dw | |||
| Quercitrin | 1073.04 µg/g dw | |||
| Quercetin | <LOQ | |||
| LC-ESI-MS | Phenolic acids: Gallic acid | 79.58 µg/g dw | ||
| Protocatechuic acid | 9.94 µg/g dw | |||
| Syringic acid | 5.20 µg/g dw | |||
| Vanillic acid | 5.57 µg/g dw | |||
| Flavonoids: Catechin | 597.65 µg/g dw | |||
| Epicatechin | 12.54 µg/g dw | |||
| [25] | Extracts in acetone and methanol | LC-ESI-MS; HPLC-UV | Phenolic acids: Gallic acid, Phtalic acid, Vanillic acid | NQ |
| Flavonoids: Catechin, Kaempferol, Taxifolin, Taxifolin-3-O-α-l-arabinofuranoside, Quercitrin, Quercetin-3-O-(4-O-acetyl)-α-l-rhamnopyranoside, | NQ | |||
| Other polyphenols: Vanillin | NQ | |||
| Esters: Ethyl gallate, Propyl gallate | NQ | |||
| [27] | Extracts in acetone/cyclohexane (for identification) and 50% methanol (for quantification) obtained by UE | UHPLC-Q-Orbitrap HRMS | Phenolic acids: Ellagic acid | 518.38–1733.64 μg/g dw a |
| Neochlorogenic acid | 32.49–126.57 μg/g dw | |||
| Gallic acid | 89.87–219.09 μg/g dw | |||
| Protocatechuic acid | 44.28–154.04 μg/g dw | |||
| Syringic acid | 11.44–26.76 μg/g dw | |||
| Vanillic acid | 18.68–44.13 μg/g dw | |||
| Flavonoids: (+)-Catechin | 289.19–693.32 μg/g dw | |||
| (−)-Epicatechin | 8.82–36.44 μg/g dw | |||
| (−)-Epigallocatechin gallate | 0.43–2.21 μg/g dw | |||
| (−)-Epicatechin gallate | 63.47–194.79 μg/g dw | |||
| Taxifolin | 33.99–153.31 μg/g dw | |||
| Myricitrin | 11.22–28.13 μg/g dw | |||
| Hyperoside | 4.20–60.27 μg/g dw | |||
| Isoquercitrin | 20.89–116.68 μg/g dw | |||
| Astilbin | 9.19–31.62 μg/g dw | |||
| Taxifolin-3-O-arabinofuranoside | 519.60–2181.84 μg/g dw | |||
| Quercitrin | 145.42–983.58 μg/g dw | |||
| Phlorizin | 3.18–34.57 μg/g dw | |||
| Quercetin | 4.76–17.43 μg/g dw | |||
| Esters: Methyl gallate | 34.32–355.66 μg/g dw | |||
| Hydroxibenzaldehydes: Protocatechualdehyde | 9.79–34.60 μg/g dw | |||
| [26] | 70% methanolic extracts obtained by UE | HPLC-UV | Phenolic acids: Chlorogenic acid, Coumaroylquinic acid isomers, Ellagic acid, Gallic acid, Neochlorogenic acid, p-Coumaroylquinic acid | NQ |
| Flavonoids: (−)-Epicatechin, (+)-Catechin, Kaempferol-rhamnoside, Hyperoside, Quercetin-rhamnose-pentoside Quercetin-3-o-glucoside, Quercetin-rhamnoside, Quercetin-rhamnoside isomers, Quercetin-rhamnose-hexoside, Quercetin-pentoside isomers, Quercetin-galloylhexoside isomers B-type procyanidin dimer, trimer, tetramer, pentamer, hexamer, and isomers | NQ | |||
| Tannins: Ellagic acid hexoside, Ellagic acid pentoside, Ellagic acid pentoside isomers, Digalloyl-glucose isomer, HHDP-glucose isomer, bis-HHDP-glucose, Galloyl-HHDP-glucose, Monogalloyl-glucose isomers, Digalloyl-glucose isomers, Trigalloyl-glucose isomers, Euprostin A isomer | NQ | |||
| [32] | 70% methanolic extracts obtained by ME, UE, and EE | LC-MS/MS | Phenolic acids: Ellagic acid | 239.65–351.92 µg/g dw b |
| Quinic acid | 28.09–44.62 µg/g dw | |||
| Coumaroylquinic acid | 7.64–10.65 µg/g dw | |||
| P-coumaric acid | 19.82–28.74 µg/g dw | |||
| Hydroxymandelic acid | 3.58–5.01 μg/g dw | |||
| Ferulic acid isomer | 55.77–87.96 μg/g dw | |||
| Protocatehuic acid | 1.15–1.24 μg/g dw | |||
| Caffeic acid | 5.54–5.88 μg/g dw | |||
| P-Hydroxybenzoic | 7.06–8.54 μg/g dw | |||
| Flavonoids: (+)-Catechin | 2.29–3.65 μg/g dw | |||
| Isoquercitrin | 3.0–4.27 μg/g dw | |||
| Luteolin | 20.41–28.39 μg/g dw | |||
| Myricitrin | 21.34–31.05 μg/g dw | |||
| Quercetin | 38.01–39.84 μg/g dw | |||
| Quercetin galloylhexoside isomer | 8.30–13.80 μg/g dw | |||
| Quercitrin | 29.59–33.83 μg/g dw | |||
| Taxifolin | 11.75–15.68 μg/g dw | |||
| Taxifolin-3-O-arabinofuranoside | 7.17–13.84 μg/g dw | |||
| Tannins: HHDP-glucose isomer | 3.84–6.75 μg/g dw | |||
| Galloyl-HHDP-glucose | 4.21–5.77 μg/g dw | |||
| Monogalloyl-glucose | 3.20–4.87 μg/g dw | |||
| Trigalloyl-glucose | 2.64–4.90 μg/g dw | |||
| Esters: Methyl gallate | 3.54–10.60 μg/g dw | |||
| [34] | Ethanolic extract | GC-TOF/MS | Phenolic acids: p-Coumaric acid | NQ |
| Phenylpropanoids: (1′-methyl-2′-hydroxy) propane-O-α-D-glucopyranoside, (4′–hydroxyphenyl) methylene-O-β-D-glucopyranosyl-(4 → 1)-α-L-arabinopyranoside, 2-carboxy-5,7-dihydroxy-3-naphthyl-β-D-glucopyranoside | NQ | |||
| [37] | Methanolic extracts (Chandler, Vina, and Franquette varieties) obtained by UE | HPLC-DAD | Flavonoids: Apigenin | 1–10 µg/g dw c |
| Catechin | 25–53 µg/g dw | |||
| Kaempferol | 2–9 µg/g dw | |||
| Luteolin | 1–5 µg/g dw c | |||
| Myricetin | 1–11 µg/g d | |||
| Quercetin | 4–16 µg/g dw | |||
| Rutin | 1–6 µg/g dw | |||
| [38] | 70% ethanolic extracts | HPLC | Flavonoids: (+)-Catechin, Catechin lactone A, (2R)-eriodictyol-5-O-β-D-glucoside,3,5,7-trihydroxylchromone-3-O-α-L-arabinofuranoside, Luteolin, Naringenin 7-O-β-D—glucopyranoside, Quercetin, 3-O-methylquercetin, Avicularin, Quercetin-3-O-α-D-arabinofuranoside, Quercetin 3-O-β-D-xylopyranoside, Quercetin-3-O-(6″-O-galloyl)-β-D-galactopyranoside, Quercetin-3-O-β-D-glucopyranoside, Sakuranetin 5-O-β-D xylopyranoside, Taxifolin, Taxifolin-3-O-α-L-arabinofuranoside, Taxifolin-3-β-D-xylopyranoside | NQ |
| Phenylpropanoids: 1,6-di-O-(E)-Coumaroyl-β-D-glucopyranoside,3,5,7-Trihydroxylchromone-3-O-α-L-arabinofuranoside, 1-O-(Z)-Coumaroyl,6-O-(E)-coumaroyl-β-D-glucopyranoside, Erythro-(7S,8R)-guaiacyl-glycerol-β-O-4′-dihydroconiferyl ether, 1-(4′-Hydroxy-3′-methoxyphenyl)-2-[4″-(3-hydroxypropyl)-2″,6″-dimethoxyphenoxy]propane-1,3-diol, Rosalaevin B | NQ | |||
| Lignans: (5-methoxy-(+)-Isolariciresinol, Erythro-guaiacyl-glycerol-β-O-4′-(5′)-methoxylariciresinol, Rhoiptelol B, Dihydrodehydodiconiferyl alcohol | NQ | |||
| [40] | 80% ethanolic extracts obtained by UE | UPLC-Q-MS | Phenolic acids: Gallic acid | 272.52 ± 0.52 μg/g dw d |
| 3,4-Dihydroxybenzoic acid | 259.06 ± 0.09 μg/g dw | |||
| Vanillic acid | 66.06 ± 1.30 μg/g dw | |||
| Syringic acid | 37.94 ± 0.22 μg/g dw | |||
| Benzoic acid | 18.55 ± 0.07 μg/g dw | |||
| 4-Hydroxybenzoic acid | 17.14 ± 0.10 μg/g dw | |||
| Caffeic acid | 9.81 ± 0.07 μg/g dw | |||
| Salicylic acid | 6.50 ± 0.28 μg/g dw | |||
| Trans-ferulic acid | 4.56 ± 0.19 μg/g dw | |||
| p-Hydroxycinnamic acid | 10.17 ± 0.58 μg/g dw | |||
| Trans-cinnamic acid | 0.74 ± 0.11 μg/g dw | |||
| 4-Hydroxy-3,5-dimethoxycinnamic acid | 0.27 ± 0.06 μg/g dw | |||
| Flavonoids: Catechin | 9989.16 ± 0.73 μg/g dw | |||
| Quercitrin | 6816.18 ± 1.61 μg/g dw | |||
| Taxifolin | 569.39 ± 0.88 μg/g dw | |||
| Quercetin 3-β-D-glucoside | 399.00 ± 1.07 μg/g dw | |||
| Epicatechin | 362.10 ± 0.50 μg/g dw | |||
| Quercetin | 54.10 ± 0.06 μg/g dw | |||
| Naringenin | 31.84 ± 0.27 μg/g dw | |||
| Rutin | 17.32 ± 0.44 μg/g dw | |||
| Vanillin | 15.53 ± 0.45 μg/g dw | |||
| Dihydrokaempferol | 14.57 ± 0.17 μg/g dw | |||
| Luteolin | 7.60 ± 0.45 μg/g dw | |||
| Dihydromyricetin | 5.75 ± 0.12 μg/g dw | |||
| Naringenin chalcone | 3.34 ± 0.10 μg/g dw | |||
| Vitexin | 1.90 ± 0.19 μg/g dw | |||
| Isorhamnetin | 0.99 ± 0.14 μg/g dw | |||
| Kaempferol | 0.96 ± 0.03 μg/g dw | |||
| Apigenin | 0.36 ± 0.05 μg/g dw | |||
| Benzaldehydes: Protocatechualdehyde | 85.46 ± 0.63 μg/g dw | |||
| Syringaldehyde | 4.90 ± 0.03 μg/g dw |
| Ref. | Extracts | Analytical Method | Compounds | Amount |
|---|---|---|---|---|
| [24] | Acetone and ethanol extracts obtained by UTE | LC-MS | Phytosterols: | |
| β-Sitosterol | 31,018.16 µg/g dw | |||
| Stigmasterol | <LOQ | |||
| Campesterol | 292.07 µg/g dw | |||
| [25] | Methanolic extract | GC/MS | Fatty acids: | |
| Octanoic acid | 1.37 ± 0.05 mg/kg dw * | |||
| Decylic acid | 1.86 ± 0.02 mg/kg dw | |||
| Lauric acid | 5.33 ± 0.10 mg/kg dw | |||
| Myristic acid | 11.62 ± 0.20 mg/kg dw | |||
| Pentadecanoic acid | 22.24 ± 0.26 mg/kg dw | |||
| Palmitic acid | 700.55 ± 8.80 mg/kg dw | |||
| Margaric acid | 28.95 ± 0.16 mg/kg dw | |||
| Stearic acid | 143.96 ± 4.02 mg/kg dw | |||
| Arachidic acid | 18.98 ± 0.27 mg/kg dw | |||
| Heneicosanoic acid | 18.66 ± 0.06 mg/kg dw | |||
| Behenic acid | 50.79 ± 0.31 mg/kg dw | |||
| Tricosanoic acid | 44.91 ±0.28 mg/kg dw | |||
| Lignoceric acid | 49.89 ± 0.24 mg/kg dw | |||
| Palmitoleic acid | 17.45 ± 0.24 mg/kg dw | |||
| cis-10-Heptadecenoic acid | 7.51 ± 0.14 mg/kg dw | |||
| Oleic acid | 549.92 ± 18.98 mg/kg dw | |||
| Linoleic acid | 1314.06 ± 10.71 mg/kg dw | |||
| c-Linolenic acid | 5.39 ± 0.09 mg/kg dw | |||
| cis-11-Eicosenoic acid | 5.47 ± 0.16 mg/kg dw | |||
| cis-11,14-Eicosadienoic acid | 3.90 ± 0.15 mg/kg dw | |||
| cis-8,11,14-Poxyeicosatrienoic acid | 0.77 ± 0.05 mg/kg dw | |||
| Arachidonic acid | 1.06 ± 0.05 mg/kg dw | |||
| cis-5,8,11,14,17-Timnodonic acid | 2.20 ± 0.08 mg/kg dw | |||
| Erucic acid | 1.82 ± 0.06 mg/kg dw | |||
| cis-13,16-Docosadienoic acid | 1.36 ± 0.09 mg/kg dw | |||
| Nervonic acid | 1.36 ± 0.11 mg/kg dw | |||
| [12] | Water/acetone (50:50, v/v) extract | LC-MS | Tocopherols: | |
| α-Tocopherol | 3.35 ± 0.04 mg/100 g dw * | |||
| β/γ-Tocopherols | 1.73 ± 0.01 mg/100 g dw | |||
| δ-Tocopherol | 1.47 ± 0.02 mg/100 g dw |
| Ref. | Extracts | Analytical Method | Compounds | Amount |
|---|---|---|---|---|
| [21] | Petroleum ether, acetone, ethanol extracts | HPGFC-RID | Monosaccharides: Arabinose, Galactose, Glucose, Xylose, Mannose | NQ |
| [25] | WS extract [44] | HPLC-MS | Monosaccharides, disaccharides: | |
| Mannose | 11.45 ± 0.52 mg/g dw * | |||
| Rhamnose | 8.99 ± 0.41 mg/g dw * | |||
| Ribose | 2.99 ± 0.27 mg/g dw * | |||
| Glucuronic acid | 3.09 ± 0.09 mg/g dw * | |||
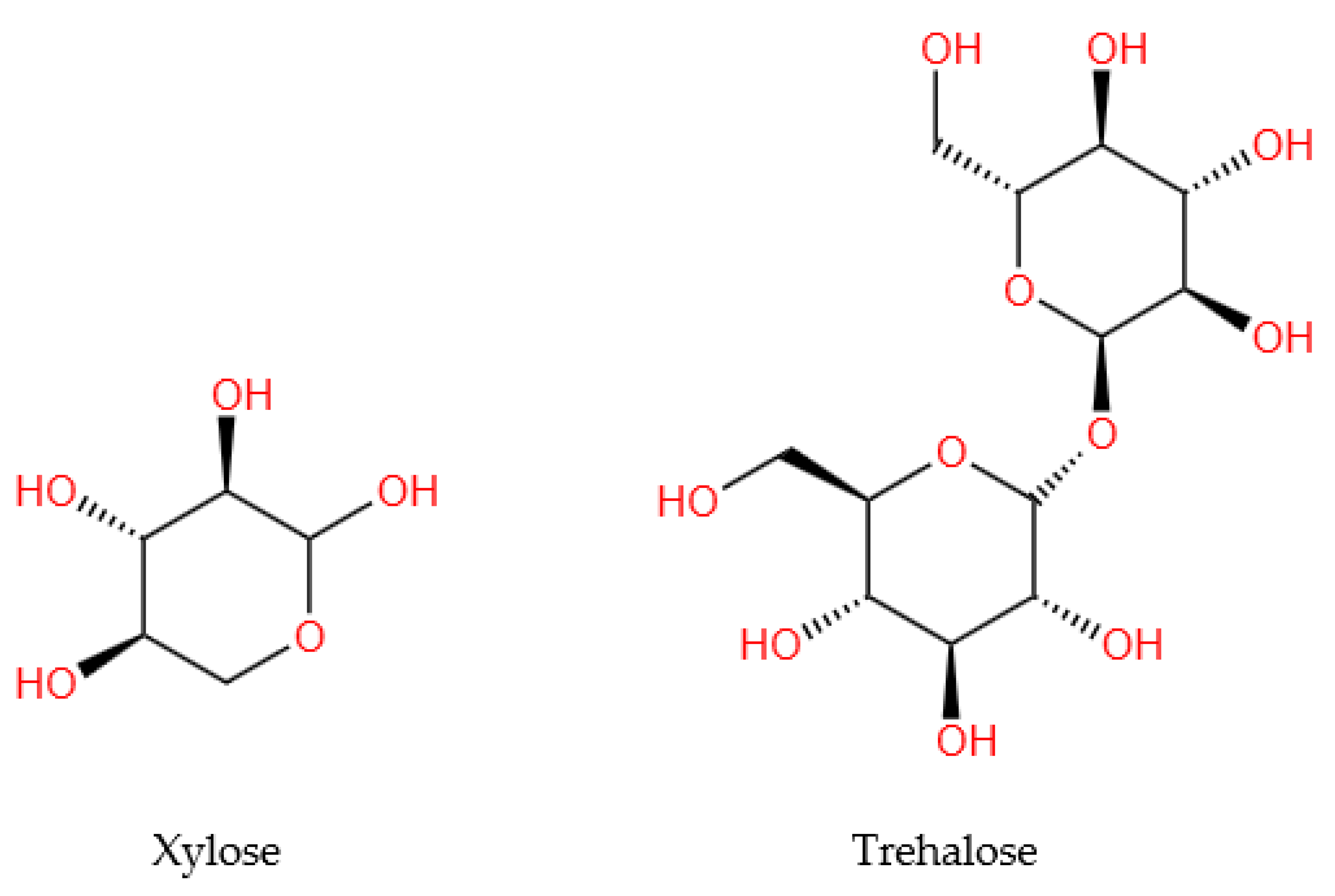
| Trehalose | 223.76 ± 10.01 mg/g dw * | |||
| Galacturonic acid | 8.18 ± 0.05 mg/g dw * | |||
| Xylose | 44.79 ± 2.08 mg/g dw * | |||
| Galactose | 2.77 ± 0.02 mg/g dw * | |||
| Arabinose | 8.11 ± 1.57 mg/g dw * |
| Classification | Compounds | Amount | Ref. |
|---|---|---|---|
| Sesquiterpenoids | Megastigmanes: Blumenol B, Dehydrovomifoliol, (6R,9R)-9-Hydroxymegastigman-4-en-3-one (blumenol C glucoside), (6R,9S)-9- Hydroxymegastigman-4-en-3-one (blumenol C glucoside) | NQ | [22] |
| Carotenoids: Dihydrophaseic acid | NQ | ||
| Megastigmanes: Blumenol B | NQ | [25] | |
| Megastigmanes: Diamegastigmane A, Diamegastigmane B, Diamegastigmane C, Blumenol A (vomifoliol), Blumenol B, Aglycone of euodionoside G, Bridelionol C, Myrsinionoside A, Byzantionoside B, (6R, 9S)-60-(400-hydroxybenzoyl)-roseoside | NQ | [38] | |
| Taraxasteranes: Juglansin A, Juglansin B, Juglansin C, Juglansin D | NQ | [39] | |
| Quinones | (4s)-4-hydroxy-1- tetralone | NA | [22] |
| Coumarins | 7-hydroxy-methylcoumarin | 240.45–332.18 µg/g dw * | [32] |
| Amino acids | Lysine (Lys) | 2.41 ± 0.02 mg/g dw | [25] |
| Phenylalanine (Phe) | 1.23 ± 0.02 mg/g dw | ||
| Threonine (Thr) | 1.04 ± 0.03 mg/g dw | ||
| Isoleucine (Ile) | 1.25 ± 0.04 mg/g dw | ||
| Leucine (Leu) | 2.09 ± 0.07 mg/g dw | ||
| Valine (Val) | 1.66 ± 0.04 mg/g dw | ||
| Aspartic acid (Asp) | 0.59 ± 0.03 mg/g dw | ||
| Serine (Ser) | 1.61 ± 0.03 mg/g dw | ||
| Glutamate (Glu) | 3.86 ± 0.02 mg/g dw | ||
| Glycine (Gly) | 2.10 ± 0.05 mg/g dw | ||
| Alanine (Ala) | 1.56 ± 0.02 mg/g dw | ||
| Cystine (Cys) | 2.15 ± 0.04 mg/g dw | ||
| Tyrosine (Tyr) | 0.88 ± 0.04 mg/g dw | ||
| Proline (Pro) | 2.40 ± 0.02 mg/g dw | ||
| Arginine (Arg) | 1.13 ± 0.03 mg/g dw | ||
| Histidine (His) | 2.61 ± 0.04 mg/g dw | ||
| Methionine (Met) | 0.98 ± 0.04 mg/g dw | ||
| Minerals | K | 8362.87 ± 21.90 mg/kg dw | [25] |
| Na | 479.17 ± 6.29 mg/kg dw | ||
| Ca | 3526.37 ± 15.71 mg/kg dw | ||
| Mg | 636.80 ± 12.94 mg/kg dw | ||
| Fe | 36.93 ± 0.90 mg/kg dw | ||
| Cu | 3.78 ± 0.16 mg/kg dw | ||
| Zn | 4.93 ± 0.11 mg/kg dw | ||
| Mn | 14.70 ± 0.10 mg/kg dw | ||
| Se | 0.049 ± 0.002 mg/kg dw |
| Ref. | Animals | Extract/Substance Tested | Groups and Doses | Parameters/Biomarkers Analyzed | Effects/Biological Activities |
| [16] |
|
|
| Biochemical parameters: serum glucose, ALAT, ASAT, ALP, insulin levels |
|
| [14] |
|
|
| Histopathological analyses of the pancreatic tissues and blood glucose levels |
|
| [18] |
|
|
| Antidiabetic and hypolipidemic effects:
| Both doses of WSE:
|
| [17] |
| WSE preparation: in water (water:walnut septa, 2:1) and boiled on a water bath until the evaporation of ½ of the water volume |
| Leucocytes in peripheral blood Peripheral blood cells in blood and bone marrow on the 8th day after the cyclophosphamide administration |
|
| [13] |
|
|
| Sorption capacity of erythrocytes Resistance to hemolysis of erythrocytes |
|
| [13] |
|
|
| Acute toxicity assay: lethality, signs of toxicity, and weight daily monitored |
|
| [19] |
|
|
| Subchronic toxicity assay Biochemical parameters (Cr, ALAT, ASAT, and urea) Antioxidant status: enzyme activities (GPx, SOD, PON-1, AO, XDH) and MDA level in liver and kidney tissues Histological analysis of the liver, kidney, heart, brain, and eye tissue samples Quantitative assessment of kidney tissue changes: epithelium thickness and diameter of renal glomerulus | JRSME:
|
| [29] |
|
|
| Blood glucose levels | DJP-2:
|
| [31] |
|
|
WK dose: calculated for 9% of the daily diet given to the animals (quantity/rat); this dose is equivalent to 43 g or 1.5 oz of nuts/2000 kcal/day; administered daily added to SF WSE dose: the equivalent amount of TPC as the WK dose (9%) determined by FC assay; administered daily added to SF | Hematological and biochemical analyses (fasting glycemia, blood lipid profile, ASAT, ALAT, creatinine, urea, CRP) Antioxidant (antiradical) activity (AA) of the brain and/or liver homogenates (by TEAC, DPPH, and FRAP assays) Oxidative stress biomarkers in the brain and liver homogenates: ROS (by DCFH-DA assay), AGEs (by fluorimetry), total NO (by Griess assay), total MDA by UPLC-PDA), total GSH level (by Ellman’s assay) AChE activity (by Ellman’s assay) Histopathological and immunohistochemical analyses of the brain and liver tissues | Both WK and WSE intake normalized some hematological parameters significantly lowered by D-gal (vs. D-gal):
Both WK and WSE intake protected hepatocytes to be affected by D-gal (that induced an atrophy with attenuation of the nuclei and dilatation and hyperemia of sinusoids in the direction of the centrilobular vein (HE-staining) WK intake, but more evidence for WSE intake, prevented the liver glycogen accumulation induced by D-gal exposure (complete absence of the hepatic glycogen stores in D-gal + WSE group) |
| [31] |
|
|
WSE dose: the equivalent amount of TPC as the WK dose (9%) determined by FC assay; administered daily added to SF | Hematological and biochemical analyses (fasting glycemia, blood lipid profile, ASAT, ALAT, creatinine, urea, CRP) AA of the brain and/or liver homogenates (by TEAC, DPPH, and FRAP assays) Oxidative stress biomarkers in the brain and liver homogenates: ROS (by DCFH-DA assay), AGEs (by fluorimetry), total NO (by Griess assay), total MDA by UPLC-PDA), total GSH level (by Ellman’s assay) AChE activity (by Ellman’s assay) Histopathological and immunohistochemical analyses of the brain and liver tissues | WK or WSE intake significantly improved some biochemical parameters affected by natural aging (vs. CO):
WK intake reduced glycogen deposits in the hepatic parenchyma; this effect was more highlight for WSE intake |
| [34] | Caenorhabditis elegans in vivo model | 2-carboxy-5,7-dihydroxy-3-naphthyl-β-D-glucopyranoside (CDG) was isolated from DJF |
| Lifespan assay and lipofuscin accumulation Stress resistance assays: the thermotolerance assay and the resistance to paraquat ROS level, antioxidant enzyme activities, and MDA level |
CDG (200 μg/mL) increased lifespan of nematodes exposed to paraquat by 40.83% relative to the control group CDG (50, 100, and 200 μg/mL) showed antioxidant activity in nematodes:
|
| [35] |
|
|
| The climbing bar time and the force swimming test at 30 min after final treatment Serum LDH activity SUN level Liver and muscle glycogen levels TAC, SOD, GSH, CAT, and MDA in serum, liver, and skeletal muscle |
|
| [36] |
|
|
WSE dose: 10 mL WSE (containing 134 mg GAE)/kg bw/day, administered orally by gavage, 1x/day, 3 consecutive days WSE 1:2 dose: 5 mL WSE (containing 134 mg GAE)/kg bw/day, administered orally by gavage, 1x/day, 3 consecutive days [mg GAE were calculated analyzing TPC by FC assay] | The number and frequency of coughs caused by citric acid aerosols (17.5 %; 4 min-period of exposure, 1 h after the last oral treatment; 8 min-period of observation), and cough latency after exposure. Oxidative Stress Biomarkers (6 h after the citric acid aerosol exposure): TAC in serum (by TEAC assay); NO and ROS levels in lung homogenates Inflammatory biomarkers (6 h after the citric acid aerosol exposure): IL-6, CXC-R1, and CXC-R2 Histopathological analysis of lung tissue (6 h after the citric acid aerosol exposure) | Significant antitussive effect of WSE, comparable with codeine:
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateș, L.; Rusu, M.E.; Popa, D.-S. Phytochemicals and Biological Activities of Walnut Septum: A Systematic Review. Antioxidants 2023, 12, 604. https://doi.org/10.3390/antiox12030604
Mateș L, Rusu ME, Popa D-S. Phytochemicals and Biological Activities of Walnut Septum: A Systematic Review. Antioxidants. 2023; 12(3):604. https://doi.org/10.3390/antiox12030604
Chicago/Turabian StyleMateș, Letiția, Marius Emil Rusu, and Daniela-Saveta Popa. 2023. "Phytochemicals and Biological Activities of Walnut Septum: A Systematic Review" Antioxidants 12, no. 3: 604. https://doi.org/10.3390/antiox12030604