



Anti-Inflammatory and Antioxidant Properties of β-Sitosterol in Copper Sulfate-Induced Inflammation in Zebrafish (Danio rerio)
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish and Experimental Conditions
2.2. Main Reagents
2.3. Experimental Design
2.3.1. Preparation of Test Samples
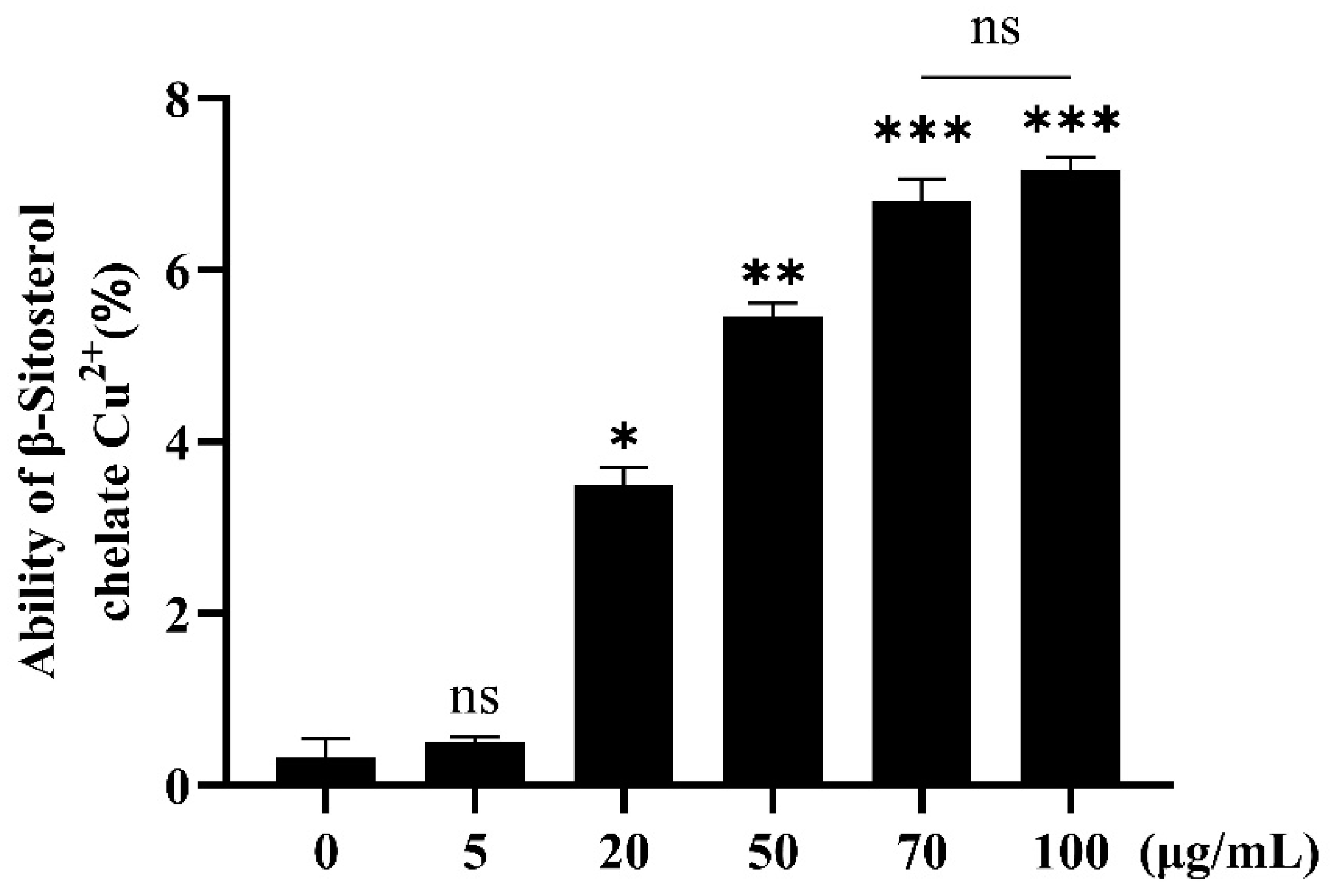
2.3.2. Chelating Ability of β-Sitosterol for Cu2+
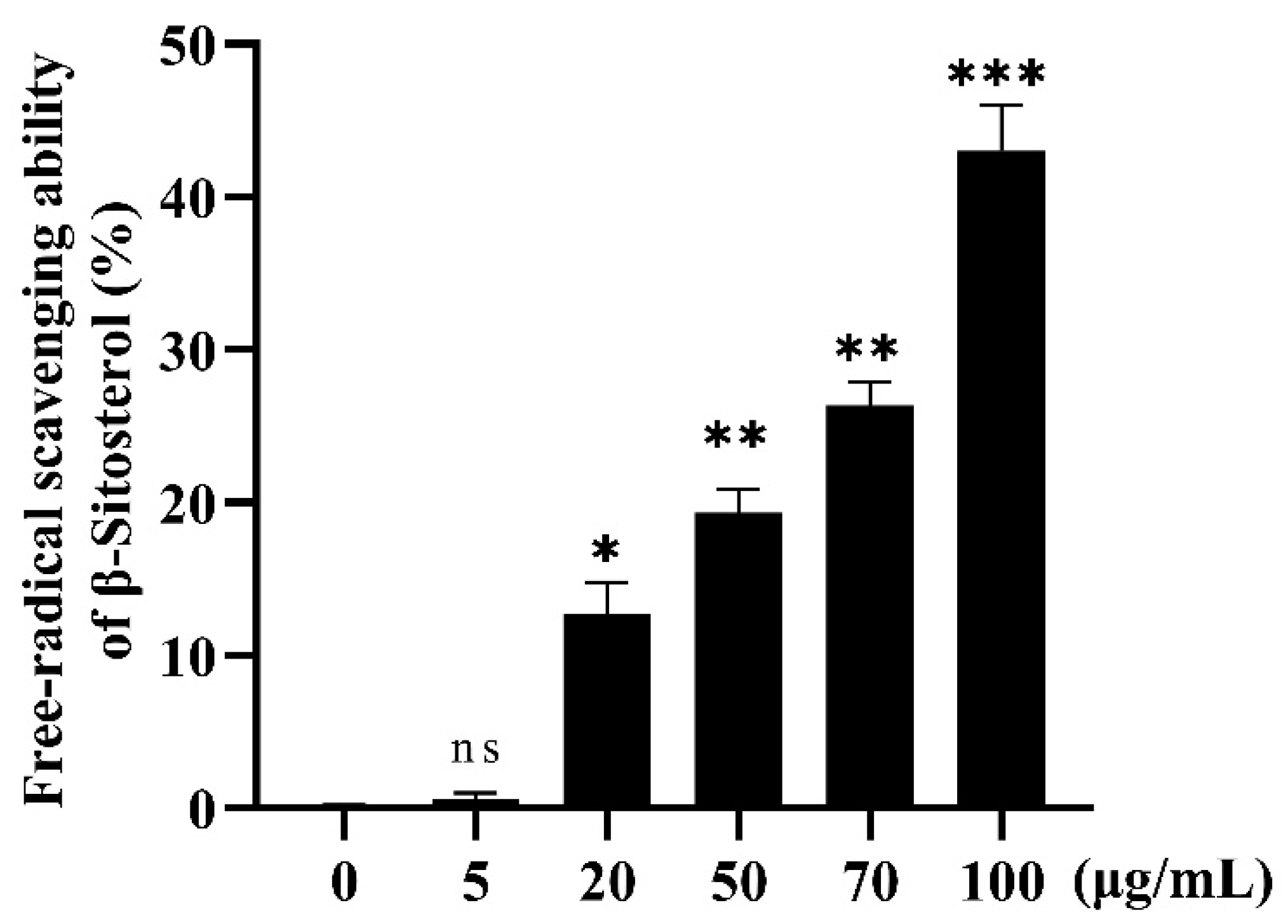
2.3.3. Antioxidant Effect of β-Sitosterol In Vitro
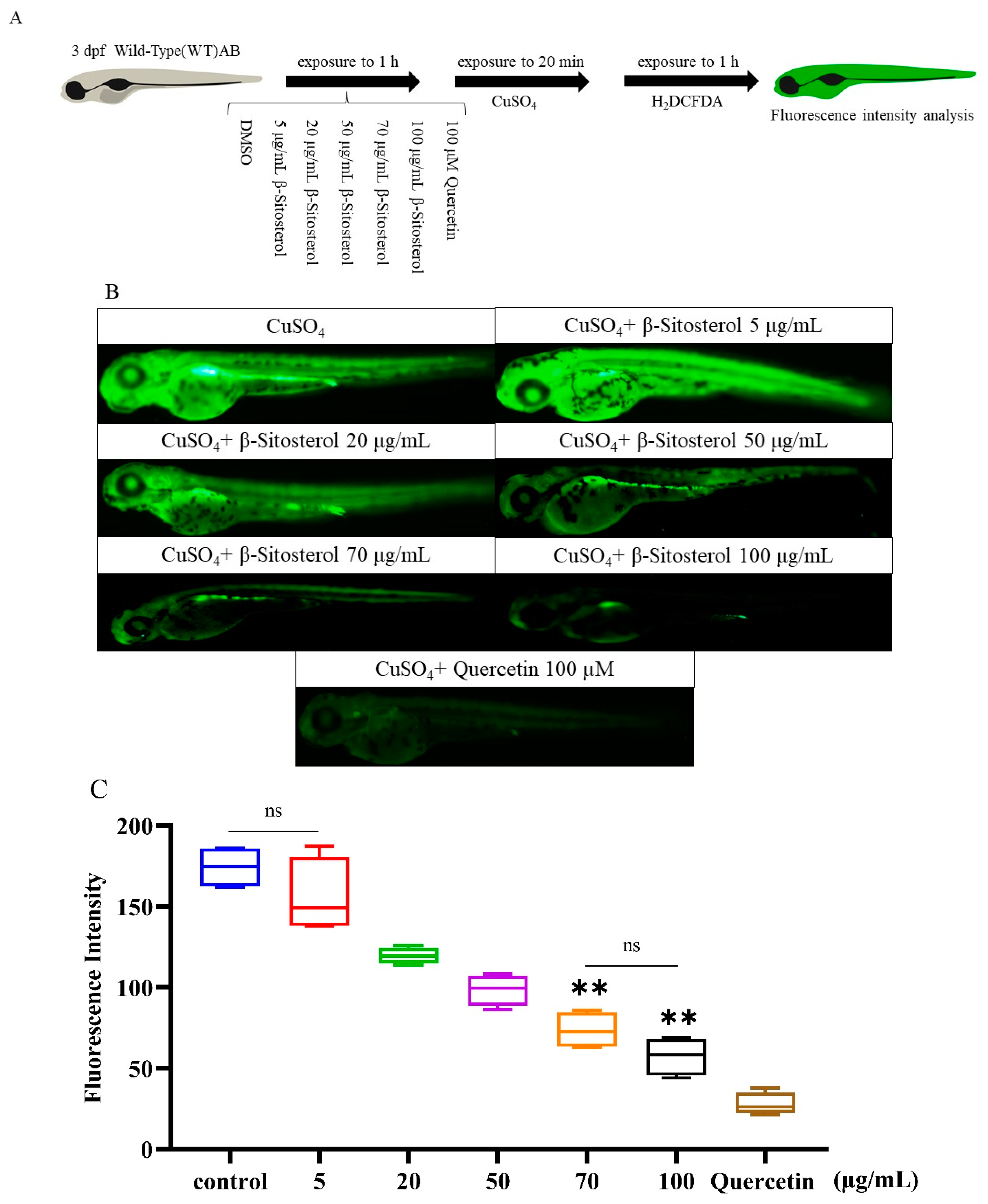
2.3.4. In Vivo Antioxidant Test
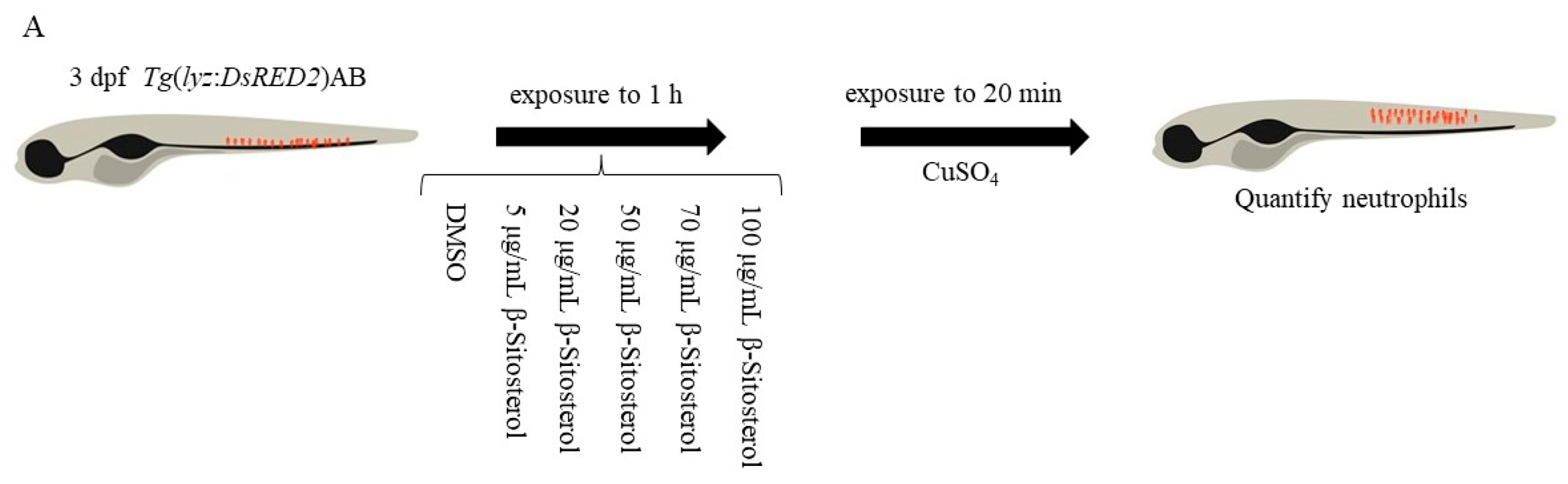
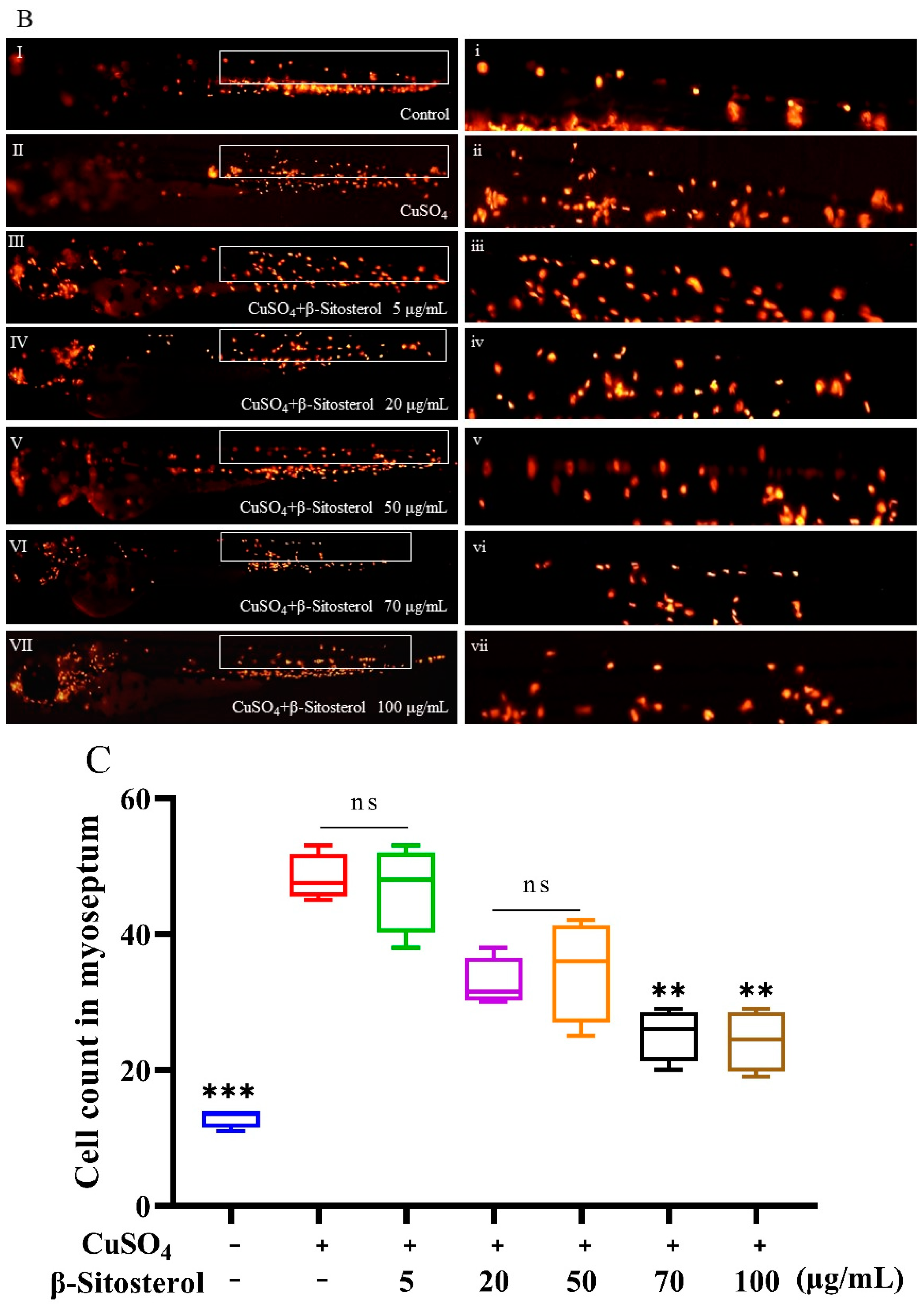
2.3.5. Neutrophil Migration Assay
2.3.6. Fluorescence Assay Quantification Method
2.3.7. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
2.3.8. Data Presentation and Statistical Analyses
3. Results
3.1. Chelating Ability of β-Sitosterol for Cu2+
3.2. Antioxidant Effect of β-Sitosterol
3.2.1. In Vitro Antioxidant Effect of β-Sitosterol
3.2.2. In Vivo Antioxidant Effect of β-Sitosterol
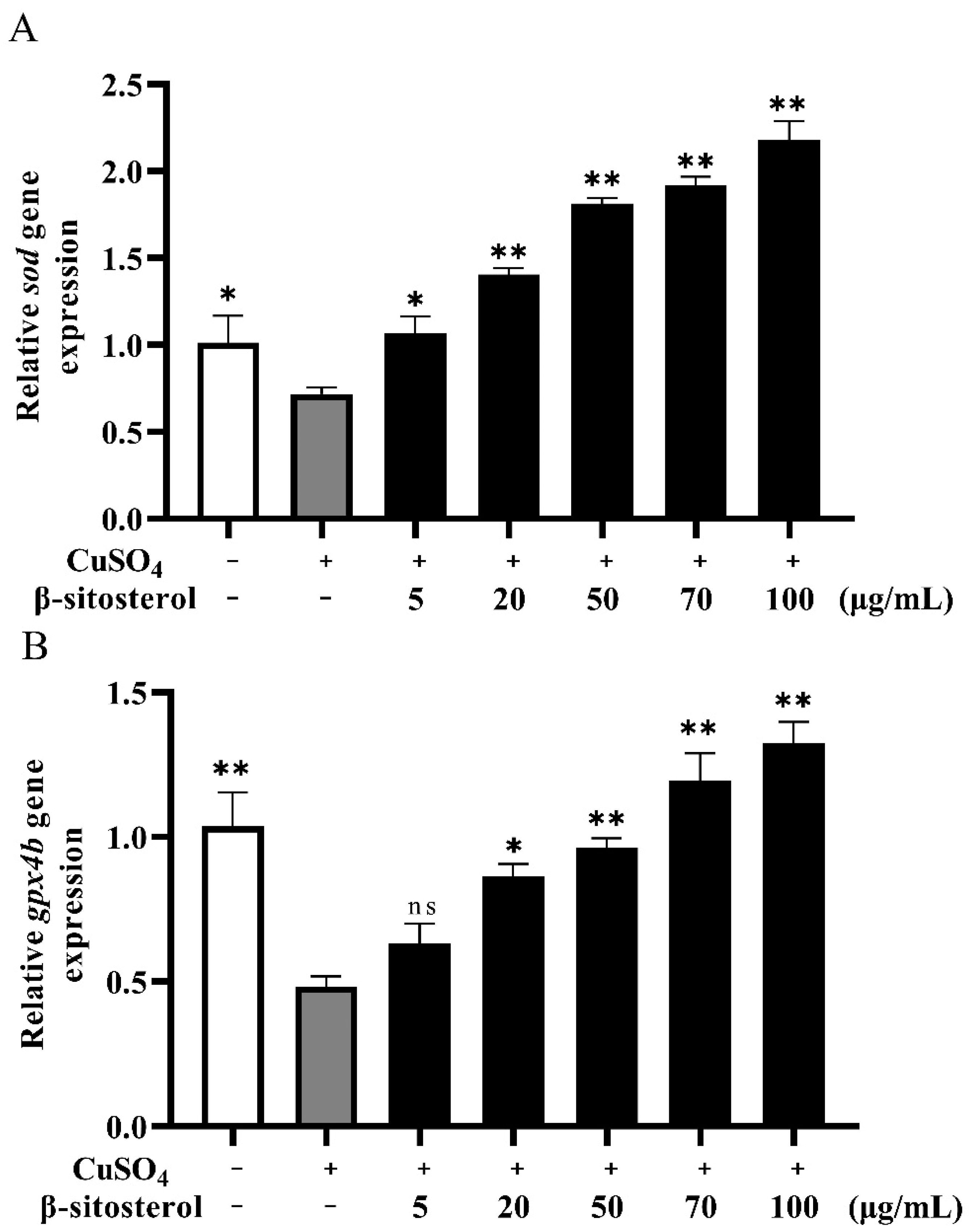
3.2.3. Effect of β-Sitosterol on the Expressions of the Antioxidant Genes sod and gpx4b
3.3. Anti-Inflammatory Effect of β-Sitosterol in Zebrafish
3.3.1. Effect of β-Sitosterol on Neutrophil Migration
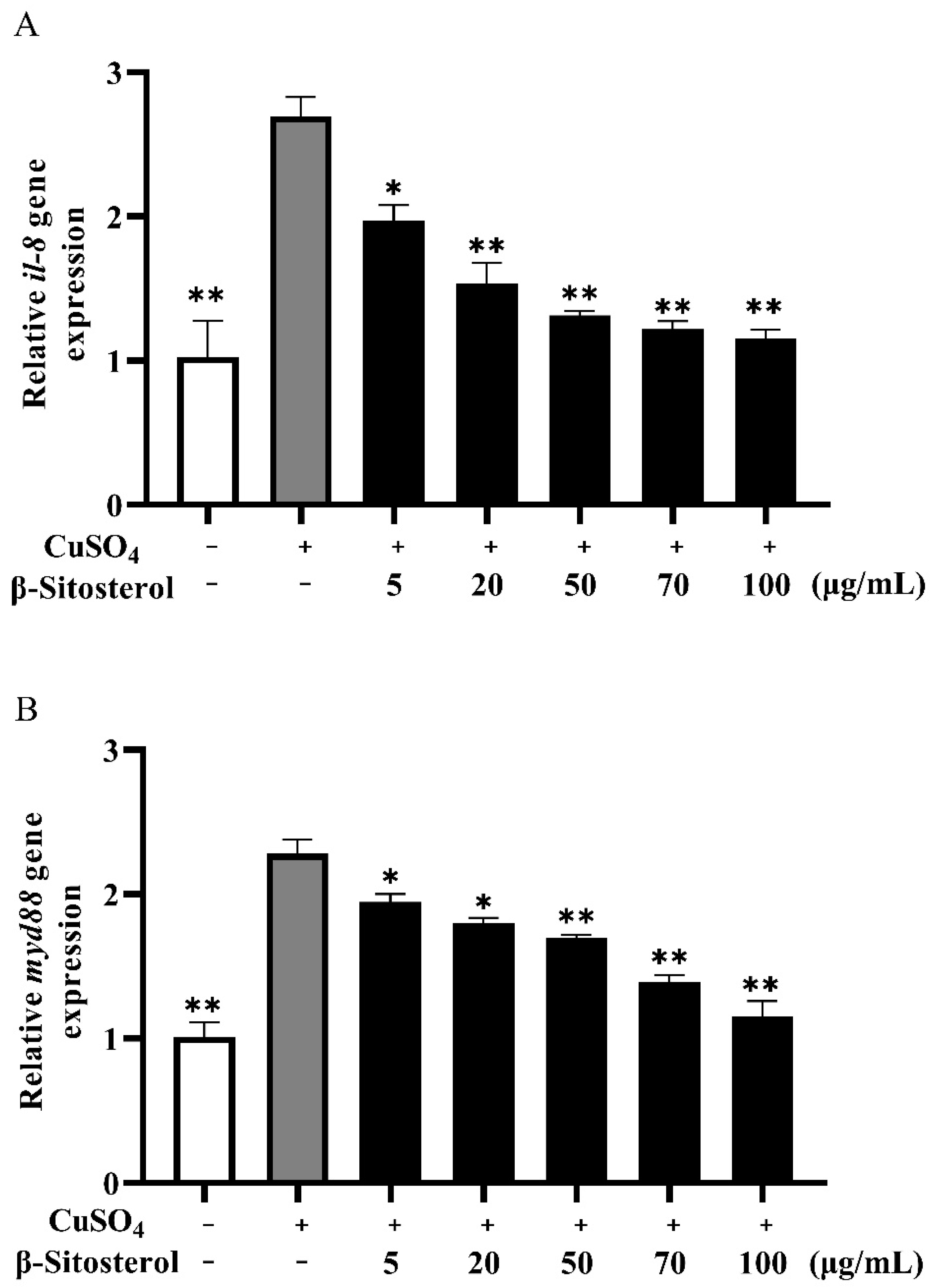
3.3.2. Effects of β-Sitosterol on the Expressions of il-8 and myd88
4. Discussion
4.1. In Vivo Antioxidant Effect of β-Sitosterol
4.2. Anti-Inflammatory Effect of β-Sitosterol in Zebrafish
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hou, C.; Chen, L.; Yang, L.; Ji, X. An insight into anti-inflammatory effects of natural polysaccharides. Int. J. Biol. Macromol. 2020, 153, 248–255. [Google Scholar] [CrossRef]
- Li, F.; Xiaoyu, L.; Weiliang, Z.; Fujiang, G.; Yingchun, W.; Rui, W.; Kaixian, C.; Cheng, H.; Yiming, L. Inhibition of human neutrophil elastase by pentacyclic triterpenes. PLoS ONE 2013, 8, e82794. [Google Scholar] [CrossRef]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxidative Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef]
- Jun, L.; Yixin, W.; John, H.W.; Yu, C.; Zhaoting, L.; Quanyin, H. Mucoadhesive probiotic backpacks with ROS nanoscavengers enhance the bacteriotherapy for inflammatory bowel diseases. Sci. Adv. 2022, 8, eabp8798. [Google Scholar] [CrossRef]
- Alessandro, D.; Jessica, K.; Mona, S.; Federica, M.; Bruno, C.; Fowler, C.J.; Valentina, O. Exploring the fatty acid amide hydrolase and cyclooxygenase inhibitory properties of novel amide derivatives of ibuprofen. J. Enzym. Inhib. Med. Chem. 2020, 35, 815–823. [Google Scholar] [CrossRef]
- Christian, C.; Emmanuel, M. Non-steroidal anti-inflammatory drugs in the pharmacological management of osteoarthritis in the very old: Prescribe or proscribe? Ther. Adv. Musculoskelet. Dis. 2021, 13, 1759720X211022149. [Google Scholar] [CrossRef]
- Liang, M.; Tzu-Wen, L.; Wallig, M.A.; Dobrucki, I.T.; Dobrucki, L.W.; Nelson, E.R.; Swanson, K.S.; Smith, A.M. Efficient Targeting of Adipose Tissue Macrophages in Obesity with Polysaccharide Nanocarriers. ACS Nano 2016, 10, 6952–6962. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Kim, H.-S.; Kim, S.-Y.; Lee, S.-H.; Lee, W.W.; Jeon, Y.-J. Identification of sterols from the soft coral Dendronephthya gigantea and their anti-inflammatory potential. Environ. Toxicol. Pharmacol. 2017, 55, 37–43. [Google Scholar] [CrossRef]
- Hussain, A.; AlAjmi, M.F.; Rehman, M.T.; Amir, S.; Husain, F.M.; Alsalme, A.; Siddiqui, M.A.; AlKhedhairy, A.A.; Khan, R.A. Copper(II) complexes as potential anticancer and Nonsteroidal anti-inflammatory agents: In vitro and in vivo studies. Sci. Rep. 2019, 9, 5237. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.J.; Medici, V.; Heffern, M.C. Correction: A caged imidazopyrazinone for selective bioluminescence detection of labile extracellular copper(ii). Chem. Sci. 2022, 13, 5774. [Google Scholar] [CrossRef]
- Brandão, P.T.C.; Martha, C.M.; Reis, B.M. Copper toxicology, oxidative stress and inflammation using zebrafish as experimental model. J. Appl. Toxicol. 2016, 36, 876–885. [Google Scholar] [CrossRef]
- Di Paola, D.; Capparucci, F.; Lanteri, G.; Crupi, R.; Marino, Y.; Franco, G.A.; Cuzzocrea, S.; Spano, N.; Gugliandolo, E.; Peritore, A.F. Environmental Toxicity Assessment of Sodium Fluoride and Platinum-Derived Drugs Co-Exposure on Aquatic Organisms. Toxics 2022, 10, 272. [Google Scholar] [CrossRef]
- Moreau, R.A.; Nyström, L.; Whitaker, B.D.; Winkler-Moser, J.K.; Baer, D.J.; Gebauer, S.K.; Hicks, K.B. Phytosterols and their derivatives: Structural diversity, distribution, metabolism, analysis, and health-promoting uses. Prog. Lipid Res. 2018, 70, 35–61. [Google Scholar] [CrossRef]
- Valerio, M.; Awad, A.B. β-Sitosterol down-regulates some pro-inflammatory signal transduction pathways by increasing the activity of tyrosine phosphatase SHP-1 in J774A.1 murine macrophages. Int. Immunopharmacol. 2011, 11, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.; Tan, Y.Y.; Ding, Y.; Fang, Y.; Yang, X.; Fang, J.; Xu, D.C.; Zhang, H.; Lu, W.; Li, M.; et al. β-Sitosterol improves experimental colitis in mice with a target against pathogenic bacteria. J. Cell. Biochem. 2019, 120, 5687–5694. [Google Scholar] [CrossRef] [PubMed]
- Paniagua-Pérez, R.; Madrigal-Bujaida, E.; Reyes-Cadena, S.; Álvarez-González, I.; Sánchez-Chapul, L.; Pérez-Gallaga, L.; Hernández, N.; Flores-Mondragón, G.; Velasco, O. Cell protection induced by beta-sitosterol: Inhibition of genotoxic damage, stimulation of lymphocyte production, and determination of its antioxidant capacity. Arch. Toxicol. 2008, 82, 615–622. [Google Scholar] [CrossRef]
- Di Paola, D.; Abbate, J.M.; Iaria, C.; Cordaro, M.; Crupi, R.; Siracusa, R.; D’Amico, R.; Fusco, R.; Impellizzeri, D.; Cuzzocrea, S.; et al. Environmental Risk Assessment of Dexamethasone Sodium Phosphate and Tocilizumab Mixture in Zebrafish Early Life Stage (Danio rerio). Toxics 2022, 10, 279. [Google Scholar] [CrossRef]
- Santos, J.S.; Alvarenga Brizola, V.R.; Granato, D. High-throughput assay comparison and standardization for metal chelating capacity screening: A proposal and application. Food Chem. 2017, 214, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Yuan, H.H.; Bao, X.L.; Lan, M.B. In vitro antioxidant and cytotoxic properties of ethanol extract of Alpinia oxyphylla fruits. Pharm. Biol. 2013, 51, 1419–1425. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Le, H.D.; Kim, T.N.T.; The, H.P.; Nguyen, T.M.; Cornet, V.; Lambert, J.; Kestemont, P. Anti–Inflammatory and Antioxidant Properties of the Ethanol Extract of Clerodendrum Cyrtophyllum Turcz in Copper Sulfate-Induced Inflammation in Zebrafish. Antioxidants 2020, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Li, C.; Jia, X.; Wang, K.; Tu, Y.; Wang, R.; Liu, K.; Lu, T.; He, C. In Vitro and In Vivo Anti-inflammatory Effects of Polyphyllin Vii Through Downregulating Mapk and Nf-kappa B Pathways. Chem. Chem. 2019, 24, 875. [Google Scholar] [CrossRef]
- Marc Sultan, V.A.; Risch, T.; Schuette, M.; Dökel, S.; Ralser, M.; Balzereit, D.; Lehrach, H.; Yaspo, M.-L. Influence of RNA extraction methods and library selection schemes on RNA-seq data. BMC Genom. 2014, 15, 675. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Leite, C.E.; Maboni, L.d.O.; Cruz, F.F.; Rosemberg, D.B.; Zimmermann, F.F.; Pereira, T.C.B.; Bogo, M.R.; Bonan, C.D.; Campos, M.M.; Morrone, F.B.; et al. Involvement of purinergic system in inflammation and toxicity induced by copper in zebrafish larvae. Toxicol. Appl. Pharmacol. 2013, 272, 681–689. [Google Scholar] [CrossRef]
- Hernandez, P.P.; Undurraga, C.; Gallardo, V.E.; Mackenzie, N.; Allende, M.L.; Reyes, A.E. Sublethal concentrations of waterborne copper induce cellular stress and cell death in zebrafish embryos and larvae. Biol. Res. 2011, 44, 7–15. [Google Scholar] [CrossRef]
- Olivari, F.A.; Hernández, P.P.; Allende, M.L. Acute copper exposure induces oxidative stress and cell death in lateral line hair cells of zebrafish larvae. Brain Res. 2008, 1244, 1–12. [Google Scholar] [CrossRef]
- Francesco, S.; Gianluca, P.; Domenico, C.; Domenico, C.; Tonino, C.; Ermanno, V. Monitoring antioxidants by coulometry: Quantitative assessment of the strikingly high antioxidant capacity of bergamot (Citrus bergamia R.) by-products. Talanta 2023, 251, 123765. [Google Scholar] [CrossRef]
- Jesús, R.M. Anti-Inflammatory and Antioxidant Properties of Plant Extracts. Antioxidants 2021, 10, 921. [Google Scholar] [CrossRef]
- Liu, H.; Wang, L.; Wang, M.-H. Antioxidant and nitric oxide release inhibition activities of methanolic extract from Clerodendrum cyrtophyllum Turcz. Hortic. Environ. Biotechnol. 2011, 52, 309–314. [Google Scholar] [CrossRef]
- Wang, Y.T.; Chen, G.C. Regulation of oxidative stress-induced autophagy by ATG9A ubiquitination. Autophagy 2022, 18, 2008–2010. [Google Scholar] [CrossRef] [PubMed]
- Lirong, H.; Kun, J.; Haibin, X.; Guiyou, T.; Jiaxin, X.; Wei, Y.; Chen, L.; Xiaoping, X.; Huiqiang, L. Oxyfluorfen exposure can cause acute kidney injury by promoting ROS-induced oxidative stress and inflammation in zebrafish. J. Hazard. Mater. 2022, 440, 129823. [Google Scholar] [CrossRef]
- Ying, W.; Junhua, L.; Huiming, Q.; Chunjing, C.; Hui, L.; Jie, C. β-Carotene extracted from Blakeslea trispora attenuates oxidative stress, inflammatory, hepatic injury and immune damage induced by copper sulfate in zebrafish (Danio rerio). Comp. Biochem. Physiology. Toxicol. Pharmacol. 2022, 258, 109366. [Google Scholar] [CrossRef]
- Lee, S.-H.; Ko, C.-I.; Jee, Y.; Jeong, Y.; Kim, M.; Kim, J.-S.; Jeon, Y.-J. Anti-inflammatory effect of fucoidan extracted from Ecklonia cava in zebrafish model. Carbohydr. Polym. 2013, 92, 84–89. [Google Scholar] [CrossRef]
- Shibo, S.; Yici, Z.; Weiping, X.; Yue, Z.; Rui, Y.; Jianli, G.; Shui, G.; Qiang, M.; Kun, M.; Jianqiang, X. Chlorophyllin Inhibits Mammalian Thioredoxin Reductase 1 and Triggers Cancer Cell Death. Antioxidants 2021, 10, 1733. [Google Scholar] [CrossRef]
- Devi, R.R.; Arumughan, C. Antiradical efficacy of phytochemical extracts from defatted rice bran. Food Chem. Toxicol. 2007, 45, 2014–2021. [Google Scholar] [CrossRef]
- Mohanan, P.V.; Devi, K.S. Toxicological evaluation of sobatum. Cancer Lett. 1998, 127, 135–140. [Google Scholar] [CrossRef]
- Ambavade, S.D.; Misar, A.V.; Ambavade, P.D. Pharmacological, nutritional, and analytical aspects of β-sitosterol: A review. Orient. Pharm. Exp. Med. 2014, 14, 193–211. [Google Scholar] [CrossRef]
- Maharajan, K.; Muthulakshmi, S.; Nataraj, B.; Ramesh, M.; Kadirvelu, K. Toxicity assessment of pyriproxyfen in vertebrate model zebrafish embryos (Danio rerio): A multi biomarker study. Aquat. Toxicol. 2018, 196, 132–145. [Google Scholar] [CrossRef]
- Wang, C.; Harwood, J.D.; Zhang, Q. Oxidative stress and DNA damage in common carp (Cyprinus carpio) exposed to the herbicide mesotrione. Chemosphere 2018, 193, 1080–1086. [Google Scholar] [CrossRef]
- Nair, P.M.G.; Park, S.Y.; Choi, J. Expression of catalase and glutathione S-transferase genes in Chironomus riparius on exposure to cadmium and nonylphenol. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 154, 399–408. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, H.; Huang, H.; Ma, Y.; Wang, R.; Hu, Y.; Zheng, X.; Chen, C.; Tang, H. Squid Ink Polysaccharides Protect Human Fibroblast Against Oxidative Stress by Regulating NADPH Oxidase and Connexin43. Front. Pharmacol. 2019, 10, 1574. [Google Scholar] [CrossRef]
- Chaudhary, V.; Katyal, P.; Panwar, H.; Kaur, J.; Aluko, R.E.; Puniya, A.K.; Poonia, A.K. Antioxidative, anti-inflammatory, and anticancer properties of the red biopigment extract from Monascus purpureus (MTCC 369). J. Food Biochem. 2022, 46, e14249. [Google Scholar] [CrossRef]
- Sun, Y.; Gao, L.; Hou, W.; Wu, J. β-Sitosterol Alleviates Inflammatory Response via Inhibiting the Activation of ERK/p38 and NF-κB Pathways in LPS-Exposed BV2 Cells. Biomed. Res. Int. 2020, 2020, 7532306. [Google Scholar] [CrossRef]
- Christine, W.; Markus, R.; Shah, A.H.; Ralf, M.; Urban, L.; Clemens, G. Facilitating drug discovery: An automated high-content inflammation assay in zebrafish. J. Vis. Exp. 2012, 16, e4203. [Google Scholar] [CrossRef]
- d’Alençon, C.A.; Oscar, P.; Christine, W.; Viviana, G.; Rebecca, J.; Felix, L.; Urban, L.; Clemens, G.; Miguel, A. A high-throughput chemically induced inflammation assay in zebrafish. BMC Biol. 2010, 8, 151. [Google Scholar] [CrossRef]
- Mara, C.; Keat, L.W.; Adina, G.; Nandula, S.V.; Manisha, B.; Qiong, S.; Francesco, B.; Maurilio, P.; Marta, S.; Andrea, C.; et al. Mutations of multiple genes cause deregulation of NF-kappaB in diffuse large B-cell lymphoma. Nature 2009, 459, 717–721. [Google Scholar] [CrossRef]
- Jacques, D.; Barton, G.M. MyD88: A central player in innate immune signaling. F1000Prime Rep. 2014, 6, 97. [Google Scholar] [CrossRef]
- Delitto, D.; Delitto, A.E.; DiVita, B.E.; Pham, K.; Han, S.; Hartlage, E.R.; Newby, B.N.; Gerber, M.H.; Behrns, K.E.; Moldawer, L.L.; et al. Human Pancreatic Cancer Cells Induce a MyD88-Dependent Stromal Response to Promote a Tumor-Tolerant Immune Microenvironment. Cancer Res. 2017, 77, 672–683. [Google Scholar] [CrossRef]
- Zhu, X.; Burfeind, K.G.; Michaelis, K.A.; Braun, T.P.; Olson, B.; Pelz, K.R.; Morgan, T.K.; Marks, D.L. MyD88 signalling is critical in the development of pancreatic cancer cachexia. J. Cachexia Sarcopenia Muscle 2019, 10, 378–390. [Google Scholar] [CrossRef]
- Ruslan, M. The spectrum of inflammatory responses. Science 2021, 734, 1070–1075. [Google Scholar] [CrossRef]
- Holmes, W.; Lee, J.; Kuang, W.; Rice, G.; Wood, W. Structure and functional expression of a human interleukin-8 receptor. Science 1991, 253, 1278–1280. [Google Scholar] [CrossRef]
- de Oliveira, S.; Reyes-Aldasoro, C.C.; Candel, S.; Renshaw, S.A.; Mulero, V.; Calado, Â. Cxcl8 (IL-8) mediates neutrophil recruitment and behavior in the zebrafish inflammatory response. J. Immunol. 2013, 190, 4349–4359. [Google Scholar] [CrossRef]
- Jiang, J.; Shi, D.; Zhou, X.Q.; Yin, L.; Feng, L.; Jiang, W.D.; Liu, Y.; Tang, L.; Wu, P.; Zhao, Y. Vitamin D inhibits lipopolysaccharide-induced inflammatory response potentially through the Toll-like receptor 4 signalling pathway in the intestine and enterocytes of juvenile Jian carp (Cyprinus carpio var. Jian). Br. J. Nutr. 2015, 114, 1560–1568. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank Accession No. | Forward and Reverse Primer Sequences (5′-3′) |
|---|---|---|
| β-actin [20] | AF057040 | F: CCCCATTGAGCACGGTATTG R: ATACATGGCAGGGGTGTTGA |
| il-8 | XM_009306855.3 | F: GAGAGGTCTGGCTGTAGATC R: AGTTGTCATCAAGGTGGCAAT |
| myd88 | NM_212814.2 | F: GTGATGCCTGTGATTTTCAGACTAA R: CGGCCTCTTCATGGATTTGT |
| sod [20] | NM_131294.1 | F: ATGGTGAACAAGGCCGTTTG R: AAAGCATGGACGTGGAAACC |
| gpx4b [20] | BC095133.1 | F: TGAGAAGGGTTTACGCATCCTG R: TGTTGTTCCCCAGTGTTCCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Liu, N.; Xue, M.; Zhang, M.; Liu, W.; Xu, C.; Fan, Y.; Meng, Y.; Zhang, Q.; Zhou, Y. Anti-Inflammatory and Antioxidant Properties of β-Sitosterol in Copper Sulfate-Induced Inflammation in Zebrafish (Danio rerio). Antioxidants 2023, 12, 391. https://doi.org/10.3390/antiox12020391
Zhang P, Liu N, Xue M, Zhang M, Liu W, Xu C, Fan Y, Meng Y, Zhang Q, Zhou Y. Anti-Inflammatory and Antioxidant Properties of β-Sitosterol in Copper Sulfate-Induced Inflammation in Zebrafish (Danio rerio). Antioxidants. 2023; 12(2):391. https://doi.org/10.3390/antiox12020391
Chicago/Turabian StyleZhang, Peng, Naicheng Liu, Mingyang Xue, Mengjie Zhang, Wei Liu, Chen Xu, Yuding Fan, Yan Meng, Qinghua Zhang, and Yong Zhou. 2023. "Anti-Inflammatory and Antioxidant Properties of β-Sitosterol in Copper Sulfate-Induced Inflammation in Zebrafish (Danio rerio)" Antioxidants 12, no. 2: 391. https://doi.org/10.3390/antiox12020391