
Isolation and Identification of Limosilactobacillus reuteri PSC102 and Evaluation of Its Potential Probiotic, Antioxidant, and Antibacterial Properties


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Conditions, and Media
2.2. Isolation and Selection of L. reuteri PSC102
2.3. Identification of L. reuteri PSC102
2.4. Scanning Electron Microscopy (SEM) Analysis
2.5. Characterization of L. reuteri PSC102
2.6. Extracellular Enzyme Activities
2.7. Acid Tolerance Test
2.8. Bile Tolerance Test
2.9. Autoaggregation and Coaggregation Assay
2.10. Adhesion to Human Colon Carcinoma (Caco-2) Cells
2.11. Antibiotic Sensitivity Test
2.12. Hemolytic and Gelatinase Activities
2.13. Antioxidant Activity
2.13.1. Sample Preparation
2.13.2. 1-Diphenyl-2-Picrylhydrazyl (DPPH) Radical Scavenging Activity
2.13.3. 2-Azinobis-(3-Ethylbenzothiazoline-6-Sulfonic acid) Diammonium Salt (ABTS) Radical Scavenging Activity
2.13.4. Reducing Power Activity
2.14. Determination of Antibacterial Activities
2.14.1. Antibacterial Activity of the L. reuteri PSC102 Culture Supernatant
2.14.2. Time-Kill Assay in Cocultures
2.15. Statistical Analysis
3. Results
3.1. Screening and Isolation of L.reuteri PSC102
3.2. Identification of L. reuteri PSC102
3.3. Biochemical Characteristics of L. reuteri PSC102
3.4. Extracellular Enzyme Activities
3.5. Acid Resistance of L. reuteri PSC102
3.6. Bile Salt Tolerance of L. reuteri PSC102
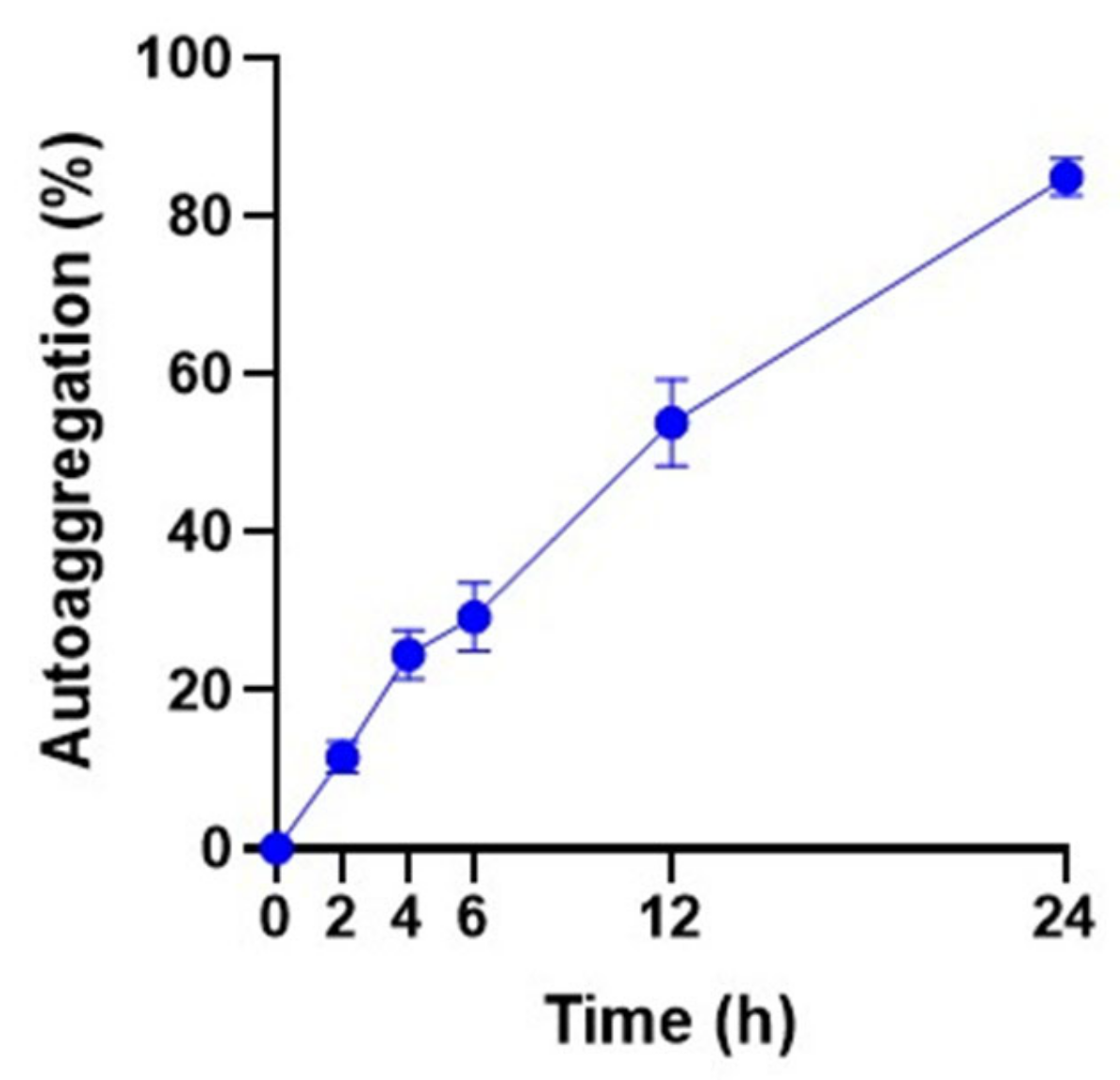
3.7. Autoaggregation Ability
3.8. Coaggregation Ability
3.9. Adhesion to Caco-2 Cells
3.10. Antibiotic Sensitivity Test
3.11. Hemolytic and Gelatinase Activities
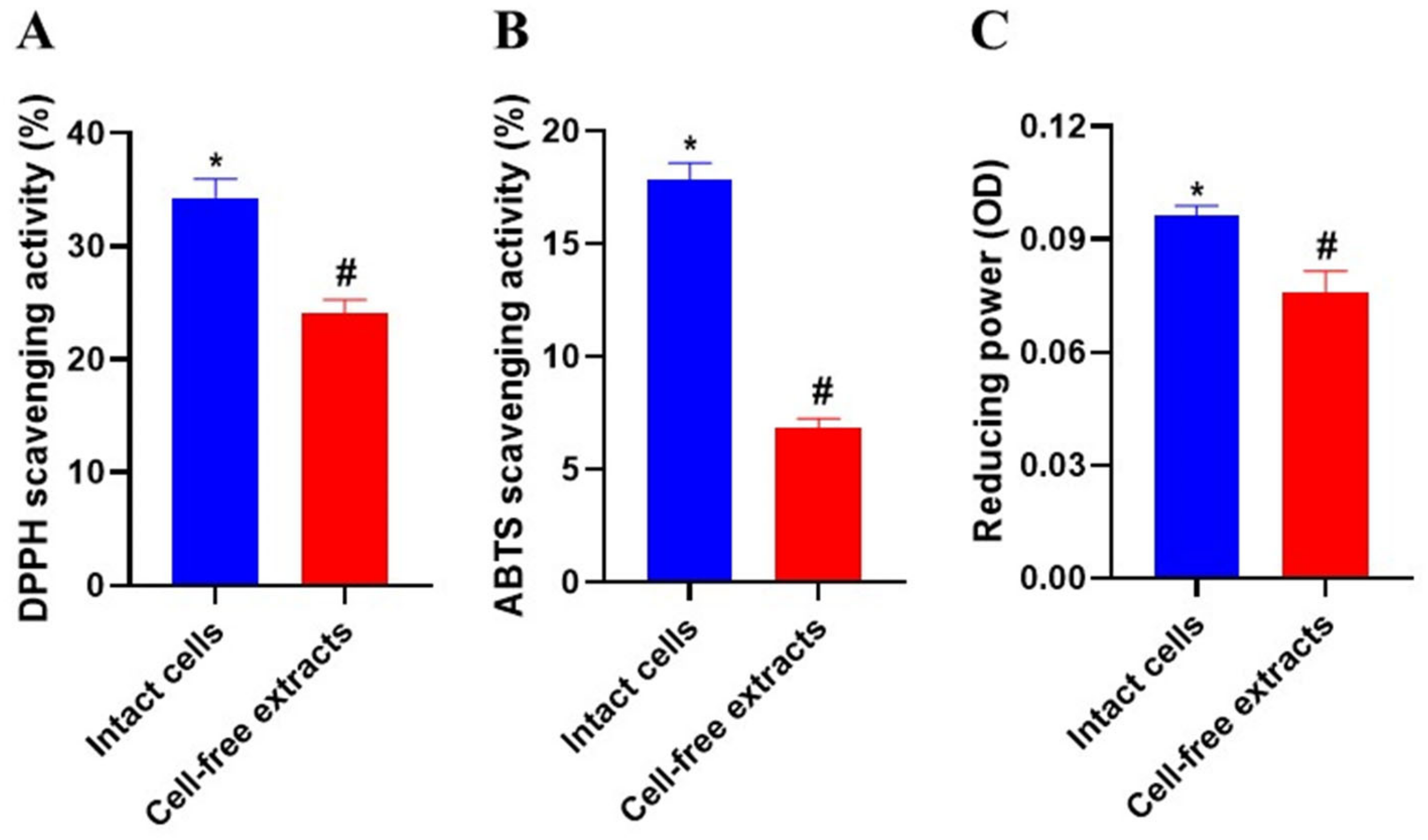
3.12. Antioxidant Activity
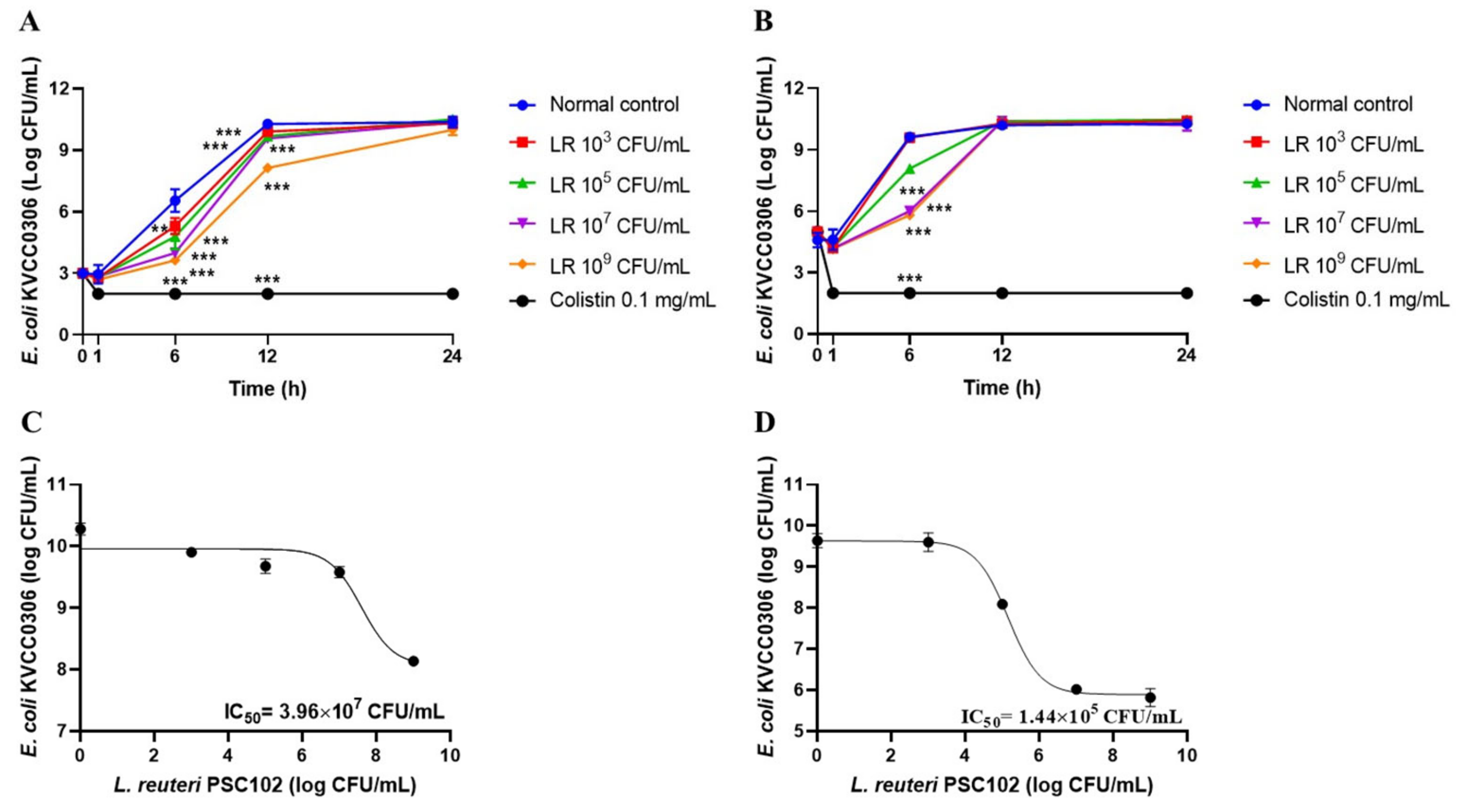
3.13. Antibacterial Activities of L. reuteri PSC102 Culture Supernatant
3.14. Time-Kill Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mingmongkolchai, S.; Panbangred, W. Bacillus Probiotics: An Alternative to Antibiotics for Livestock Production. J. Appl. Microbiol. 2018, 124, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Zhao, P.Y.; Kim, I.H. Probiotics as a Dietary Additive for Pigs: A Review. J. Anim. Vet. Adv. 2011, 10, 2127–2134. [Google Scholar] [CrossRef]
- Guerra, N.P.; Bernárdez, P.F.; Méndez, J.; Cachaldora, P.; Castro, L.P. Production of Four Potentially Probiotic Lactic Acid Bacteria and Their Evaluation as Feed Additives for Weaned Piglets. Anim. Feed Sci. Technol. 2007, 134, 89–107. [Google Scholar] [CrossRef]
- Lim, Y.H.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdullah, N. Comparative Studies of Versatile Extracellular Proteolytic Activities of Lactic Acid Bacteria and Their Potential for Extracellular Amino Acid Productions as Feed Supplements. J. Anim. Sci. Biotechnol. 2019, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Hou, C.; Zeng, X.; Yang, F.; Liu, H.; Qiao, S. Study and Use of the Probiotic Lactobacillus reuteri in Pigs: A Review. J. Anim. Sci. Biotechnol. 2015, 6, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markowiak, P.; Śliżewska, K. The Role of Probiotics, Prebiotics and Synbiotics in Animal Nutrition. Gut Pathog. 2018, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Lavilla-Lerma, L.; Pérez-Pulido, R.; Martínez-Bueno, M.; Maqueda, M.; Valdivia, E. Characterization of Functional, Safety, and Gut Survival Related Characteristics of Lactobacillus Strains Isolated from Farmhouse Goat’s Milk Cheeses. Int. J. Food Microbiol. 2013, 163, 136–145. [Google Scholar] [CrossRef]
- Cizeikiene, D.; Jagelaviciute, J. Investigation of Antibacterial Activity and Probiotic Properties of Strains Belonging to Lactobacillus and Bifidobacterium Genera for Their Potential Application in Functional Food and Feed Products. Probiotics Antimicrob. Proteins 2021, 13, 1387–1403. [Google Scholar] [CrossRef]
- Wang, J.; Ji, H.F.; Wang, S.X.; Zhang, D.Y.; Liu, H.; Shan, D.C.; Wang, Y.M. Lactobacillus plantarum ZLP001: In Vitro Assessment of Antioxidant Capacity and Effect on Growth Performance and Antioxidant Status in Weaning Piglets. Asian Australas. J. Anim. Sci. 2012, 25, 1153. [Google Scholar] [CrossRef] [Green Version]
- Firuzi, O.; Miri, R.; Tavakkoli, M.; Saso, L. Antioxidant Therapy: Current Status and Future Prospects. Curr. Med. Chem. 2011, 18, 3871–3888. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The Role of Antioxidants in the Chemistry of Oxidative Stress: A Review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Valeriano, V.D.V.; Balolong, M.P.; Kang, D. Probiotic Roles of Lactobacillus Sp. in Swine: Insights from Gut Microbiota. J. Appl. Microbiol. 2017, 122, 554–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohland, C.L.; MacNaughton, W.K. Probiotic Bacteria and Intestinal Epithelial Barrier Function. Am. J. Physiol. Liver Physiol. 2010, 298, G807–G819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietrangelo, L.; Magnifico, I.; Petronio Petronio, G.; Cutuli, M.A.; Venditti, N.; Nicolosi, D.; Perna, A.; Guerra, G.; Di Marco, R. A Potential “Vitaminic Strategy” against Caries and Halitosis. Appl. Sci. 2022, 12, 2457. [Google Scholar] [CrossRef]
- Yong, C.-C.; Khoo, B.-Y.; Sasidharan, S.; Piyawattanametha, W.; Kim, S.-H.; Khemthongcharoen, N.; Chuah, L.-O.; Ang, M.-Y.; Liong, M.-T. Activity of Crude and Fractionated Extracts by Lactic Acid Bacteria (LAB) Isolated from Local Dairy, Meat, and Fermented Products against Staphylococcus aureus. Ann. Microbiol. 2015, 65, 1037–1047. [Google Scholar] [CrossRef]
- Marianelli, C.; Cifani, N.; Pasquali, P. Evaluation of Antimicrobial Activity of Probiotic Bacteria against Salmonella enterica subsp. enterica Serovar Typhimurium 1344 in a Common Medium under Different Environmental Conditions. Res. Microbiol. 2010, 161, 673–680. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic Activities of Lactobacilli and Bifidobacteria against Microbial Pathogens. FEMS Microbiol. Rev. 2004, 28, 405–440. [Google Scholar] [CrossRef] [Green Version]
- Hyronimus, B.; Le Marrec, C.; Hadj Sassi, A.; Deschamps, A. Acid and Bile Tolerance of Spore-Forming Lactic Acid Bacteria. Int. J. Food Microbiol. 2000, 61, 193–197. [Google Scholar] [CrossRef]
- Abbas, M.A.; Lee, E.-B.; Boby, N.; Biruhanu, B.T.; Park, S.-C. A Pharmacodynamic Investigation to Assess the Synergism of Orbifloxacin and Propyl Gallate against Escherichia coli. Front. Pharmacol. 2022, 13, 989395. [Google Scholar] [CrossRef]
- Kim, E.-Y.; Kim, Y.-H.; Rhee, M.-H.; Song, J.-C.; Lee, K.-W.; Kim, K.-S.; Lee, S.-P.; Lee, I.-S.; Park, S.-C. Selection of Lactobacillus sp. PSC101 That Produces Active Dietary Enzymes Such as Amylase, Lipase, Phytase and Protease in Pigs. J. Gen. Appl. Microbiol. 2007, 53, 111–117. [Google Scholar] [CrossRef]
- Lee, J.-S.; Damte, D.; Lee, S.-J.; Hossain, M.-A.; Belew, S.; Kim, J.-Y.; Rhee, M.-H.; Kim, J.-C.; Park, S.-C. Evaluation and Characterization of a Novel Probiotic Lactobacillus pentosus PL11 Isolated from Japanese Eel (Anguilla japonica) for Its Use in Aquaculture. Aquac. Nutr. 2015, 21, 444–456. [Google Scholar] [CrossRef]
- Ali, M.S.; Lee, E.-B.; Quah, Y.; Birhanu, B.T.; Suk, K.; Lim, S.-K.; Park, S.-C. Heat-Killed Limosilactobacillus reuteri PSC102 Ameliorates Impaired Immunity in Cyclophosphamide-Induced Immunosuppressed Mice. Front. Microbiol. 2022, 13, 820838. [Google Scholar] [CrossRef] [PubMed]
- Kondrotiene, K.; Lauciene, L.; Andruleviciute, V.; Kasetiene, N.; Serniene, L.; Sekmokiene, D.; Malakauskas, M. Safety Assessment and Preliminary in Vitro Evaluation of Probiotic Potential of Lactococcus Lactis Strains Naturally Present in Raw and Fermented Milk. Curr. Microbiol. 2020, 77, 3013–3023. [Google Scholar] [CrossRef]
- Jena, P.K.; Trivedi, D.; Thakore, K.; Chaudhary, H.; Giri, S.S.; Seshadri, S. Isolation and Characterization of Probiotic Properties of Lactobacilli Isolated from Rat Fecal Microbiota. Microbiol. Immunol. 2013, 57, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Barik, A.; Patel, G.D.; Sen, S.K.; Rajhans, G.; Nayak, C.; Raut, S. Probiotic Characterization of Indigenous Kocuria flava Y4 Strain Isolated from Dioscorea villosa Leaves. Probiotics Antimicrob. Proteins 2021, 1–16. [Google Scholar] [CrossRef]
- Hossain, M.A.; Park, H.-C.; Park, S.-W.; Park, S.-C.; Seo, M.-G.; Her, M.; Kang, J. Synergism of the Combination of Traditional Antibiotics and Novel Phenolic Compounds against Escherichia coli. Pathogens 2020, 9, 811. [Google Scholar] [CrossRef]
- Perin, L.M.; Miranda, R.O.; Todorov, S.D.; de Melo Franco, B.D.G.; Nero, L.A. Virulence, Antibiotic Resistance and Biogenic Amines of Bacteriocinogenic Lactococci and Enterococci Isolated from Goat Milk. Int. J. Food Microbiol. 2014, 185, 121–126. [Google Scholar] [CrossRef]
- Tian, L.; Liu, R.; Zhou, Z.; Xu, X.; Feng, S.; Kushmaro, A.; Marks, R.S.; Wang, D.; Sun, Q. Probiotic Characteristics of Lactiplantibacillus Plantarum N-1 and Its Cholesterol-Lowering Effect in Hypercholesterolemic Rats. Probiotics Antimicrob. Proteins 2022, 14, 337–348. [Google Scholar] [CrossRef]
- Son, S.-H.; Yang, S.-J.; Jeon, H.-L.; Yu, H.-S.; Lee, N.-K.; Park, Y.-S.; Paik, H.-D. Antioxidant and Immunostimulatory Effect of Potential Probiotic Lactobacillus paraplantarum SC61 Isolated from Korean Traditional Fermented Food, jangajji. Microb. Pathog. 2018, 125, 486–492. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. Clin. Lab Stand. Inst. 2016, 35, 16–38. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. 2019. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 27 November 2022).
- Ghanbari, M.; Rezaei, M.; Jami, M.; Nazari, R.M. Isolation and Characterization of Lactobacillus Species from Intestinal Contents of Beluga (Huso huso) and Persian Sturgeon (Acipenser persicus). Iran. J. Vet. Res. 2009, 10, 152–157. [Google Scholar]
- Aragón, F.; Carino, S.; Perdigón, G.; de Moreno de LeBlanc, A. Inhibition of Growth and Metastasis of Breast Cancer in Mice by Milk Fermented with Lactobacillus casei CRL 431. J. Immunother. 2015, 38, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Kwon, B.; Ku, S.; Ji, G.E. The Efficacy of Bifidobacterium longum BORI and Lactobacillus acidophilus AD031 Probiotic Treatment in Infants with Rotavirus Infection. Nutrients 2017, 9, 887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, S.S.Y.; Wan, M.L.Y.; El-Nezami, H. Probiotics-Mediated Suppression of Cancer. Curr. Opin. Oncol. 2017, 29, 62–72. [Google Scholar] [CrossRef]
- Galdeano, C.M.; Cazorla, S.I.; Dumit, J.M.L.; Vélez, E.; Perdigón, G. Beneficial Effects of Probiotic Consumption on the Immune System. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar]
- Venditti, N.; Vergalito, F.; Magnifico, I.; Cutuli, M.A.; Pietrangelo, L.; Cozzolino, A.; Angiolillo, A.; Succi, M.; Petronio, G.P.; Di Marco, R. The Lepidoptera Galleria Mellonella “In Vivo” Model: A Preliminary Pilot Study on Oral Administration of Lactobacillus plantarum (Now Lactiplantibacillus plantarum). New Microbiol. 2021, 44, 42–50. [Google Scholar]
- Lee, Y.; Choi, Y.; Yoon, Y. Lactic Acid Bacteria in Kimchi Might Be a Cause for Carcinogen Production in Intestine. Food Control 2021, 126, 108045. [Google Scholar] [CrossRef]
- Delgado, S.; O’sullivan, E.; Fitzgerald, G.; Mayo, B. In Vitro Evaluation of the Probiotic Properties of Human Intestinal Bifidobacterium Species and Selection of New Probiotic Candidates. J. Appl. Microbiol. 2008, 104, 1119–1127. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.-A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of Potential Probiotic Lactic Acid Bacteria from Fermented Olives by in Vitro Tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Sánchez, B.; Champomier-Vergès, M.-C.; Collado, M.D.C.; Anglade, P.; Baraige, F.; Sanz, Y.; de los Reyes-Gavilán, C.G.; Margolles, A.; Zagorec, M. Low-PH Adaptation and the Acid Tolerance Response of Bifidobacterium longum Biotype Longum. Appl. Environ. Microbiol. 2007, 73, 6450–6459. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Ohishi, H.; Benno, Y. H+-ATPase Activity in Bifidobacterium with Special Reference to Acid Tolerance. Int. J. Food Microbiol. 2004, 93, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zhang, Y.; Zhang, Y.; Liu, Y.; Wang, S.; Dong, X.; Wang, Y.; Zhang, H. Screening of Potential Probiotic Properties of Lactobacillus fermentum Isolated from Traditional Dairy Products. Food Control 2010, 21, 695–701. [Google Scholar] [CrossRef]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile Resistance Mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzolino, A.; Vergalito, F.; Tremonte, P.; Iorizzo, M.; Lombardi, S.J.; Sorrentino, E.; Luongo, D.; Coppola, R.; Di Marco, R.; Succi, M. Preliminary Evaluation of the Safety and Probiotic Potential of Akkermansia muciniphila DSM 22959 in Comparison with Lactobacillus rhamnosus GG. Microorganisms 2020, 8, 189. [Google Scholar] [CrossRef] [PubMed]
- Grześkowiak, Ł.; Collado, M.C.; Salminen, S. Evaluation of Aggregation Abilities between Commensal Fish Bacteria and Pathogens. Aquaculture 2012, 356, 412–414. [Google Scholar] [CrossRef]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and Aggregation Ability of Probiotic Strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Rubayet Ul Alam, A.S.M.; Jahid, I.K. Characterization and Evaluation of Lactic Acid Bacteria from Indigenous Raw Milk for Potential Probiotic Properties. J. Dairy Sci. 2020, 103, 1223–1237. [Google Scholar] [CrossRef]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Measurement of Aggregation Properties between Probiotics and Pathogens: In Vitro Evaluation of Different Methods. J. Microbiol. Methods 2007, 71, 71–74. [Google Scholar] [CrossRef]
- Schellenberg, J.; Smoragiewicz, W.; Karska-Wysocki, B. A Rapid Method Combining Immunofluorescence and Flow Cytometry for Improved Understanding of Competitive Interactions between Lactic Acid Bacteria (LAB) and Methicillin-Resistant S. aureus (MRSA) in Mixed Culture. J. Microbiol. Methods 2006, 65, 1–9. [Google Scholar] [CrossRef]
- Han, S.; Lu, Y.; Xie, J.; Fei, Y.; Zheng, G.; Wang, Z.; Liu, J.; Lv, L.; Ling, Z.; Berglund, B. Probiotic Gastrointestinal Transit and Colonization after Oral Administration: A Long Journey. Front. Cell. Infect. Microbiol. 2021, 11, 609722. [Google Scholar] [CrossRef]
- Śliżewska, K.; Chlebicz-Wójcik, A.; Nowak, A. Probiotic Properties of New Lactobacillus Strains Intended to Be Used as Feed Additives for Monogastric Animals. Probiotics Antimicrob. Proteins 2021, 13, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Klare, I.; Konstabel, C.; Werner, G.; Huys, G.; Vankerckhoven, V.; Kahlmeter, G.; Hildebrandt, B.; Müller-Bertling, S.; Witte, W.; Goossens, H. Antimicrobial Susceptibilities of Lactobacillus, Pediococcus and Lactococcus Human Isolates and Cultures Intended for Probiotic or Nutritional Use. J. Antimicrob. Chemother. 2007, 59, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Kumherová, M.; Veselá, K.; Kosová, M.; Mašata, J.; Horáčková, Š.; Šmidrkal, J. Novel Potential Probiotic Lactobacilli for Prevention and Treatment of Vulvovaginal Infections. Probiotics Antimicrob. Proteins 2021, 13, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Kodner, C.; Kudrimoti, A. Diagnosis and Management of Acute Interstitial Nephritis. Am. Fam. Physician 2003, 67, 2527–2534. [Google Scholar] [PubMed]
- Barbosa, J.; Gibbs, P.A.; Teixeira, P. Virulence Factors among Enterococci Isolated from Traditional Fermented Meat Products Produced in the North of Portugal. Food Control 2010, 21, 651–656. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Miyazaki, T. Beneficial Effects of Antioxidative Lactic Acid Bacteria. AIMS Microbiol. 2017, 3, 1–7. [Google Scholar] [CrossRef]
- Wang, A.N.; Yi, X.W.; Yu, H.F.; Dong, B.; Qiao, S.Y. Free Radical Scavenging Activity of Lactobacillus fermentum in Vitro and Its Antioxidative Effect on Growing–Finishing Pigs. J. Appl. Microbiol. 2009, 107, 1140–1148. [Google Scholar] [CrossRef]
- Halder, D.; Mandal, M.; Chatterjee, S.S.; Pal, N.K.; Mandal, S. Indigenous Probiotic Lactobacillus Isolates Presenting Antibiotic like Activity against Human Pathogenic Bacteria. Biomedicines 2017, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Suzuki, Y.; Guan, L.L. Dissect the Mode of Action of Probiotics in Affecting Host-Microbial Interactions and Immunity in Food Producing Animals. Vet. Immunol. Immunopathol. 2018, 205, 35–48. [Google Scholar] [CrossRef]
- Langa, S.; Martín-Cabrejas, I.; Montiel, R.; Peirotén, Á.; Arqués, J.L.; Medina, M. Protective Effect of Reuterin-Producing Lactobacillus Reuteri against Listeria Monocytogenes and Escherichia coli O157:H7 in Semi-Hard Cheese. Food Control 2018, 84, 284–289. [Google Scholar] [CrossRef]
- Bennett, S.; Ben Said, L.; Lacasse, P.; Malouin, F.; Fliss, I. Susceptibility to Nisin, Bactofencin, Pediocin and Reuterin of Multidrug Resistant Staphylococcus aureus, Streptococcus dysgalactiae and Streptococcus uberis Causing Bovine Mastitis. Antibiotics 2021, 10, 1418. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, L.; Auchtung, T.A.; Hermans, K.E.; Whitehead, D.; Borhan, B.; Britton, R.A. The Antimicrobial Compound Reuterin (3-Hydroxypropionaldehyde) Induces Oxidative Stress via Interaction with Thiol Groups. Microbiology 2010, 156, 1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šušković, J.; Kos, B.; Beganović, J.; Leboš Pavunc, A.; Habjanič, K.; Matošić, S. Antimicrobial Activity–the Most Important Property of Probiotic and Starter Lactic Acid Bacteria. Food Technol. Biotechnol. 2010, 48, 296–307. [Google Scholar]
- Alakomi, H.-L.; Skytta, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic Acid Permeabilizes Gram-Negative Bacteria by Disrupting the Outer Membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Gram Staining | Cell Morphology | Catalase | Acid Production | Lactic Acid Production |
|---|---|---|---|---|---|
| L0002 | + | Rod | − | + | D, L |
| L0006 | + | Rod | − | + | D, L |
| L0010 | + | Rod | − | + | D, L |
| L0013 | + | Rod | − | + | D, L |
| L0014 | + | Rod | − | + | D, L |
| L0017 | + | Rod | − | + | D, L |
| L0018 | + | Rod | − | + | D, L |
| L0102 | + | Rod | − | + | D, L |
| Isolates | Protease | Lipase | Amylase | Phytase |
|---|---|---|---|---|
| L0002 | ++ | + | + | + |
| L0006 | +++ | + | + | ++ |
| L0010 | ++ | − | ++ | − |
| L0013 | ++ | − | + | + |
| L0014 | +++ | − | + | + |
| L0017 | +++ | + | +++ | ++ |
| L0018 | ++ | + | +++ | + |
| L0102 | +++ | ++ | +++ | +++ |
| Active Ingredient | Result | Active Ingredient | Result | Active Ingredient | Result |
|---|---|---|---|---|---|
| Control | − | Inositol | − | Inulin | + |
| Glycerol | + | Mannitol | + | Melezitose | − |
| Erythritol | − | Sorbitol | + | Raffinose | + |
| D-arabinose | − | α-Methyl-D-mannoside | + | Starch | + |
| L-arabinose | − | β-Methyl-xyloside | − | Glycogen | + |
| D-ribose | + | α-Methyl-D-glucoside | − | Xylitol | − |
| D-xylose | + | N-Acetyl-glucosamine | − | Gentiobiose | − |
| L-xylose | + | Amygdalin | + | D-Turanose | + |
| D-adonitol | − | Arbutine | + | D-Lyxose | − |
| Methyl-β-D-xylopyranoside | − | Esculine | + | D-Tagatose | − |
| D-galactose | + | Salicine | + | D-fucose | + |
| D-glucose | + | Cellobiose | + | L-fucose | − |
| D-fructose | + | Maltose | + | D-arabitol | − |
| D-mannose | + | Lactose | + | L-arabitol | − |
| L-sorbose | + | Melibiose | + | Gluconate | − |
| Rhamnose | − | Sucrose | + | 2-keto-Gluconate | + |
| Dulcitol | − | Trehalose | + |
| No. | Enzymes | Limosilactobacillus reuteri PSC102 | Limosilactobacillus reuteri KCTC 3594 | Lactobacillus acidophilus KCTC 3146 |
|---|---|---|---|---|
| 1 | Control | 0 | 0 | 0 |
| 2 | Alkaline phosphatase | 0 | 0 | 0 |
| 3 | Esterase | 3 | 2 | 2 |
| 4 | Esterase lipase | 1 | 1 | 1 |
| 5 | Lipase | 0 | 0 | 0 |
| 6 | Leucine arylamides | 5 | 5 | 4 |
| 7 | Valine arylamides | 1 | 3 | 1 |
| 8 | Cystine arylamides | 1 | 0 | 0 |
| 9 | Trypsin | 0 | 0 | 0 |
| 10 | α-Chymotrypsin | 0 | 0 | 0 |
| 11 | Acid phosphatase | 4 | 2 | 2 |
| 12 | Naphthol-AS-BI-phosphohydrolase | 4 | 2 | 2 |
| 13 | α-Galactosidase | 4 | 4 | 1 |
| 14 | β-Galactosidase | 1 | 5 | 4 |
| 15 | β-Glucuronidase | 0 | 0 | 0 |
| 16 | α-Glucosidase | 5 | 2 | 1 |
| 17 | β-Glucosidase | 0 | 0 | 0 |
| 18 | N-acyl-glucosaminidase | 0 | 0 | 0 |
| 19 | α-Mannosidase | 0 | 0 | 0 |
| 20 | α-Fructosidase | 0 | 0 | 0 |
| Bacteria | pH 2 | pH 3 | pH 5 | pH 7 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 1 h | 6 h | 12 h | 0 h | 1 h | 6 h | 12 h | 0 h | 1 h | 6 h | 12 h | 0 h | 1 h | 6 h | 12 h | |
| Log CFU/mL | Log CFU/mL | Log CFU/mL | Log CFU/mL | |||||||||||||
| L. reuteri PSC102 | 5.55 ± 0.08 | 3.75 ± 0.16 * | 3.52 ± 0.09 * | 2.00 ± 0.00 | 5.55 ± 0.08 | 4.81 ± 0.12 | 3.74 ± 0.08 * | 2.00 ± 0.00 | 5.55 ± 0.08 | 4.95 ± 0.05 | 4.52 ± 0.08 * | 7.65 ± 0.14 * | 5.55 ± 0.08 | 5.97 ± 0.06 | 6.52 ± 0.23 | 10.75 ± 0.15 * |
| L. reuteri KCCM 40,717 | 5.29 ± 0.16 | 2.00 ± 0.00 # | 2.00 ± 0.00 # | 2.00 ± 0.00 | 5.29 ± 0.16 | 4.35 ± 0.24 | 2.00 ± 0.00 # | 2.00 ± 0.00 | 5.29 ± 0.16 | 4.94 ± 0.05 | 5.13 ± 0.11 # | 5.16 ± 0.07 # | 5.29 ± 0.16 | 5.99 ± 0.15 | 6.51 ± 0.05 | 9.17 ± 0.07 # |
| Bacteria | Bile Salt (0.1%) | Bile Salt (0.3%) | Bile Salt (1%) | Bile Salt (0%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 1 h | 6 h | 12 h | 0 h | 1 h | 6 h | 12 h | 0 h | 1 h | 6 h | 12 h | 0 h | 1 h | 6 h | 12 h | |
| Log CFU/mL | Log CFU/mL | Log CFU/mL | Log CFU/mL | |||||||||||||
| L. reuteri PSC102 | 5.23 ± 0.04 | 5.27 ± 0.09 | 5.41 ± 0.11 | 7.28 ± 0.09 | 5.23 ± 0.09 | 5.26 ± 0.02 | 5.38 ± 0.01 | 6.83 ± 0.08 * | 5.23 ± 0.04 | 5.20 ± 0.06 | 5.32 ± 0.06 | 6.53 ± 0.09 * | 5.23 ± 0.04 | 5.36 ± 0.11 | 6.65 ± 0.08 | 9.19 ± 0.09 |
| L. reuteri KCCM 40,717 | 5.19 ± 0.10 | 5.22 ± 0.08 | 5.53 ± 0.10 | 7.27 ± 0.11 | 5.19 ± 0.05 | 5.25 ± 0.06 | 5.37 ± 0.04 | 6.28 ± 0.06 # | 5.19 ± 0.10 | 5.17 ± 0.06 | 5.28 ± 0.04 | 5.18 ± 0.09 # | 5.19 ± 0.10 | 5.75 ± 0.06 | 6.53 ± 0.06 | 9.09 ± 0.07 |
| Pathogenic Bacteria | Coaggregation with L. reuteri PSC102 (%) | |
|---|---|---|
| 2 h | 24 h | |
| E. coli ATCC 35,218 | 6.52 ± 0.75 | 77.16 ± 1.59 |
| E. coli KVCC0306 | 9.02 ± 0.91 | 81.13 ± 0.87 |
| S. aureus ATCC 29,213 | 3.35 ± 0.51 | 72.41 ± 0.69 |
| Initial Concentration (L. reuteri PSC102, CFU/mL) | Final Concentration (L. reuteri PSC102, CFU/mL) | Adhesion (%) |
|---|---|---|
| 1.08 × 108 | 4 × 106 | 3.70 |
| 1.26 × 108 | 6 × 106 | 4.76 |
| 1.10 × 108 | 4 × 106 | 3.63 |
| Average | ||
| 1.14 × 108 | 4.6 × 106 | 4.03 |
| Antibiotics | Limosilactobacillus reuteri PSC102 | Limosilactobacillus reuteri KCCM 40,717 | Staphylococcus aureus ATCC 25,922 | Enterotoxigenic E. coli KVCC0306 | ||||
|---|---|---|---|---|---|---|---|---|
| MIC (μg/mL) | MBC (μg/mL) | MIC (μg/mL) | MBC (μg/mL) | MIC (μg/mL) | MBC (μg /mL) | MIC (μg/mL) | MBC (μg/mL) | |
| Cephalexin | >256 | >256 | >256 | >256 | 2 | 4 | 32 | 64 |
| Colistin sulfate | >64 | >128 | >64 | >128 | >64 | >128 | 0.05 | 1 |
| Enrofloxacin | 8 | 16 | 4 | 8 | 0.05 | 1 | 0.025 | 1 |
| Cefalonium | 32 | 64 | 64 | 32 | 2 | 4 | 16 | 32 |
| Amoxicillin trihydrate | 4 | 2 | 1 | 1 | 0.5 | 2 | 1 | 2 |
| Penicillin G procaine | 32 | 64 | 16 | 64 | 0.05 | 1 | 32 | 64 |
| Norfloxacin | >256 | >256 | >256 | >256 | 0.5 | 4 | 0.5 | 1 |
| Spectinomycin | 64 | 128 | 8 | 128 | 8 | 64 | 2 | 4 |
| Tylosin base | 4 | 8 | 2 | 4 | 1 | 8 | 32 | 64 |
| Cefuroxime sodium | 2 | 32 | 1 | 8 | 0.05 | 2 | 1 | 2 |
| Florfenicol | 2 | 16 | 2 | 16 | 2 | 8 | 4 | 16 |
| Penicillin G benzathine | 4 | 8 | 2 | 4 | 0.05 | 2 | 16 | 32 |
| Gentamicin sulfate | 64 | 128 | 32 | 128 | 4 | 16 | 1 | 2 |
| Streptomycin sulfate | 64 | 128 | 32 | 128 | 4 | 16 | 1 | 2 |
| Pathogens | Zone of Inhibition (mm) | |||
|---|---|---|---|---|
| Concentrated Supernatant of L. reuteri PSC102 | ||||
| ×10 | ×5 | ×1 | Positive Control (Ampicillin, 10 µg) | |
| E. coli KVCC0306 | 14.10 ± 0.08 | 8.96 ± 0.03 | - | - |
| E. coli KVCC0543 | 14.20 ± 0.06 | 8.94 ± 0.04 | - | 20.35 ± 0.35 |
| E. coli KVCC1423 | 16.17 ± 0.51 | 9.60 ± 0.41 | - | 21.47 ± 0.65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.S.; Lee, E.-B.; Lim, S.-K.; Suk, K.; Park, S.-C. Isolation and Identification of Limosilactobacillus reuteri PSC102 and Evaluation of Its Potential Probiotic, Antioxidant, and Antibacterial Properties. Antioxidants 2023, 12, 238. https://doi.org/10.3390/antiox12020238
Ali MS, Lee E-B, Lim S-K, Suk K, Park S-C. Isolation and Identification of Limosilactobacillus reuteri PSC102 and Evaluation of Its Potential Probiotic, Antioxidant, and Antibacterial Properties. Antioxidants. 2023; 12(2):238. https://doi.org/10.3390/antiox12020238
Chicago/Turabian StyleAli, Md. Sekendar, Eon-Bee Lee, Suk-Kyung Lim, Kyoungho Suk, and Seung-Chun Park. 2023. "Isolation and Identification of Limosilactobacillus reuteri PSC102 and Evaluation of Its Potential Probiotic, Antioxidant, and Antibacterial Properties" Antioxidants 12, no. 2: 238. https://doi.org/10.3390/antiox12020238