
Portulaca Oleracea L. (Purslane) Extract Protects Endothelial Function by Reducing Endoplasmic Reticulum Stress and Oxidative Stress through AMPK Activation in Diabetic Obese Mice


 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation and Identification of the Purslane Extract
2.3. Animal Experiments
2.4. Functional Assay by Wire Myograph
2.5. Ex vivo Culture of Mouse Aortas
2.6. Primary Culture of Rat Aortic Endothelial Cells (RAECs)
2.7. Western Blotting
2.8. Measurement of NO Release
2.9. ROS Determination by Dihydroethidium (DHE) Staining
2.10. Statistical Analysis
3. Results
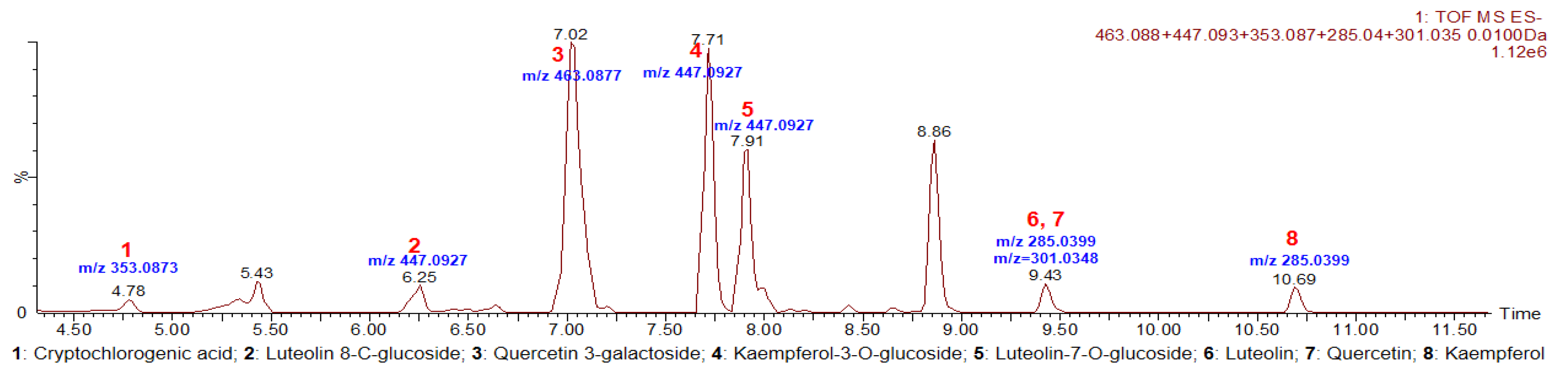
3.1. Identification Results of Purslane Extract
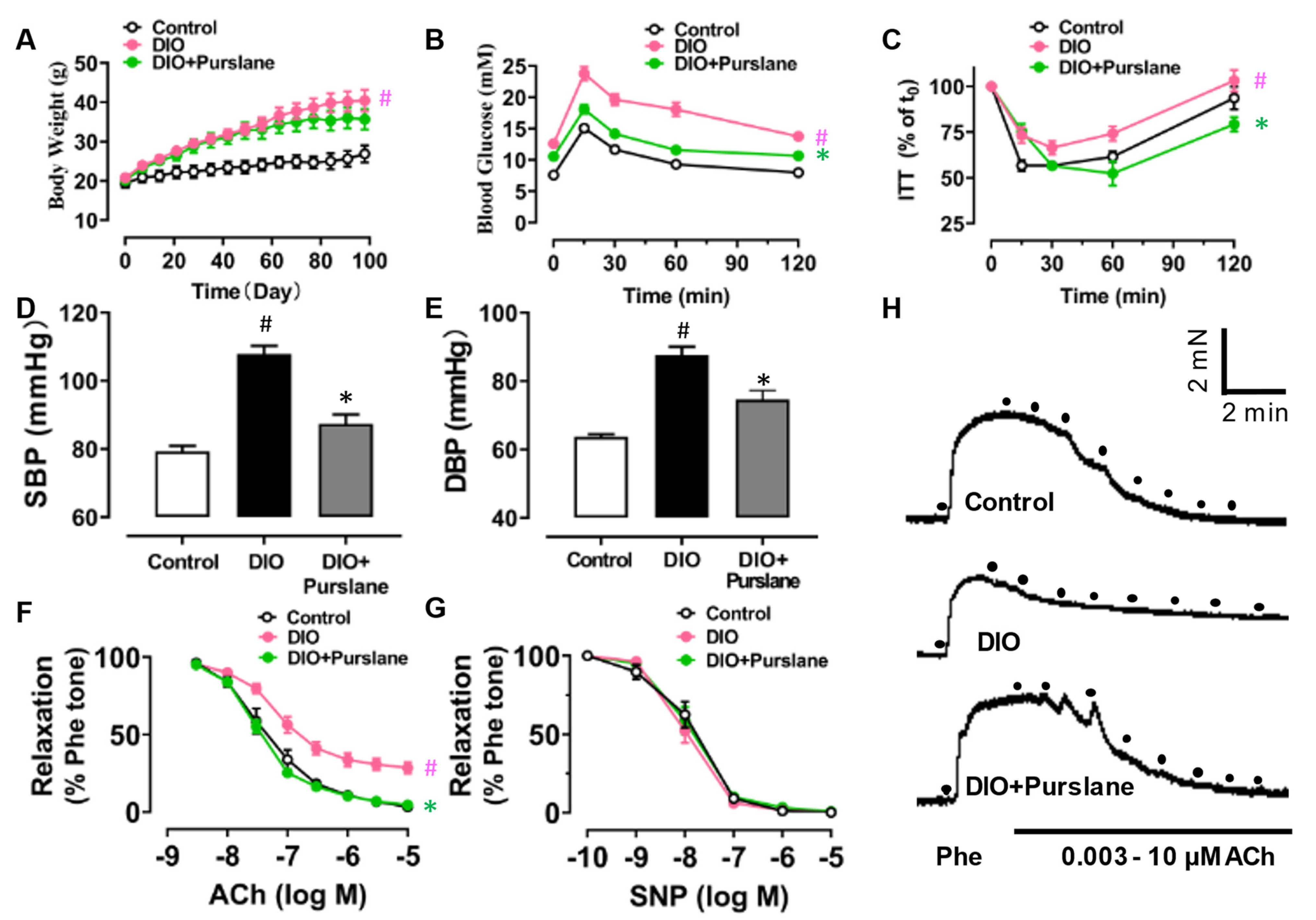
3.2. Purslane Extract Treatment Ameliorates Endothelial Dysfunction and Glucose Metabolism Disorder in DIO Mice
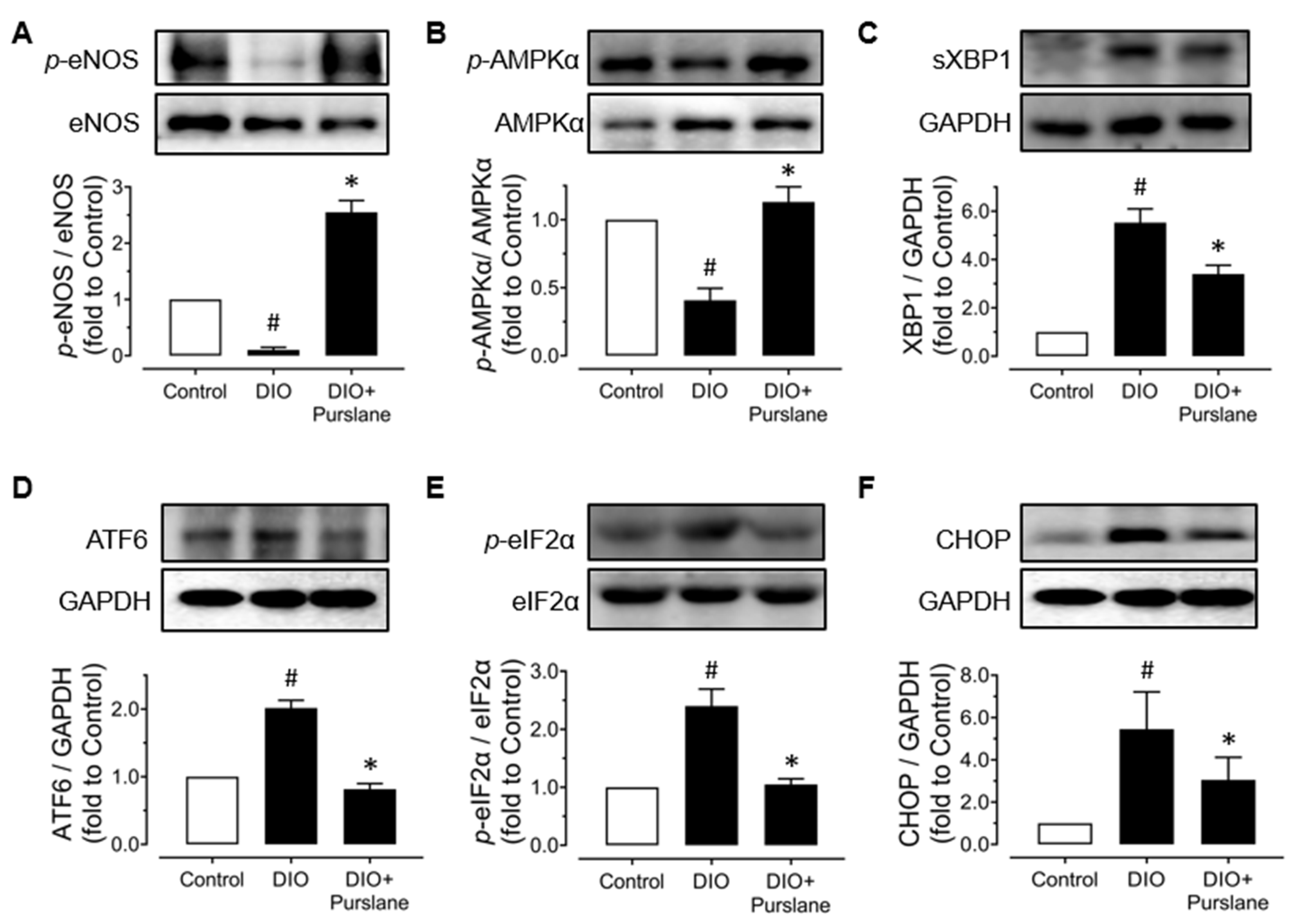
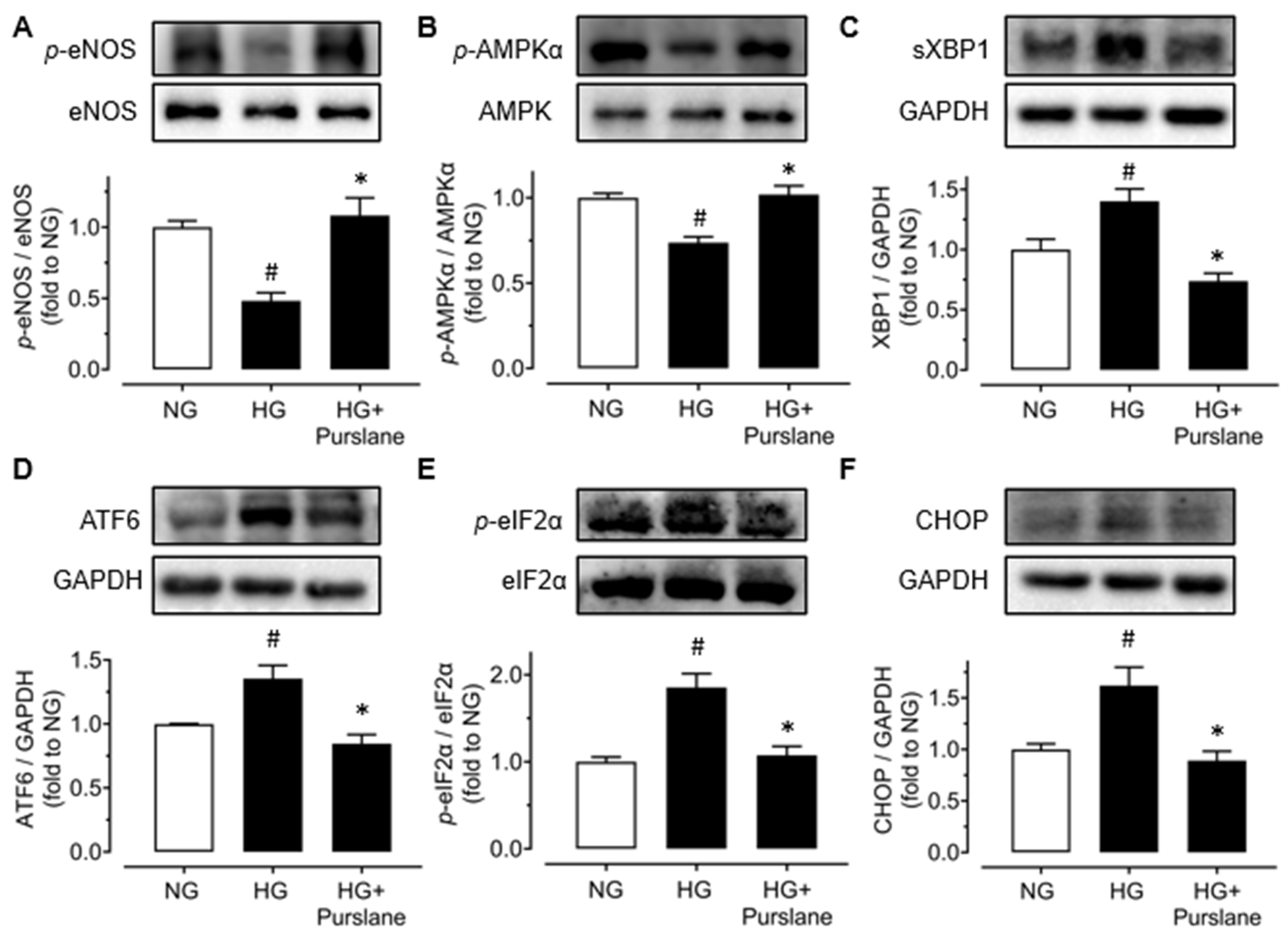
3.3. Purslane Improves Endothelial Function via Activation on AMPK/eNOS Signaling and Suppression on ER Stress
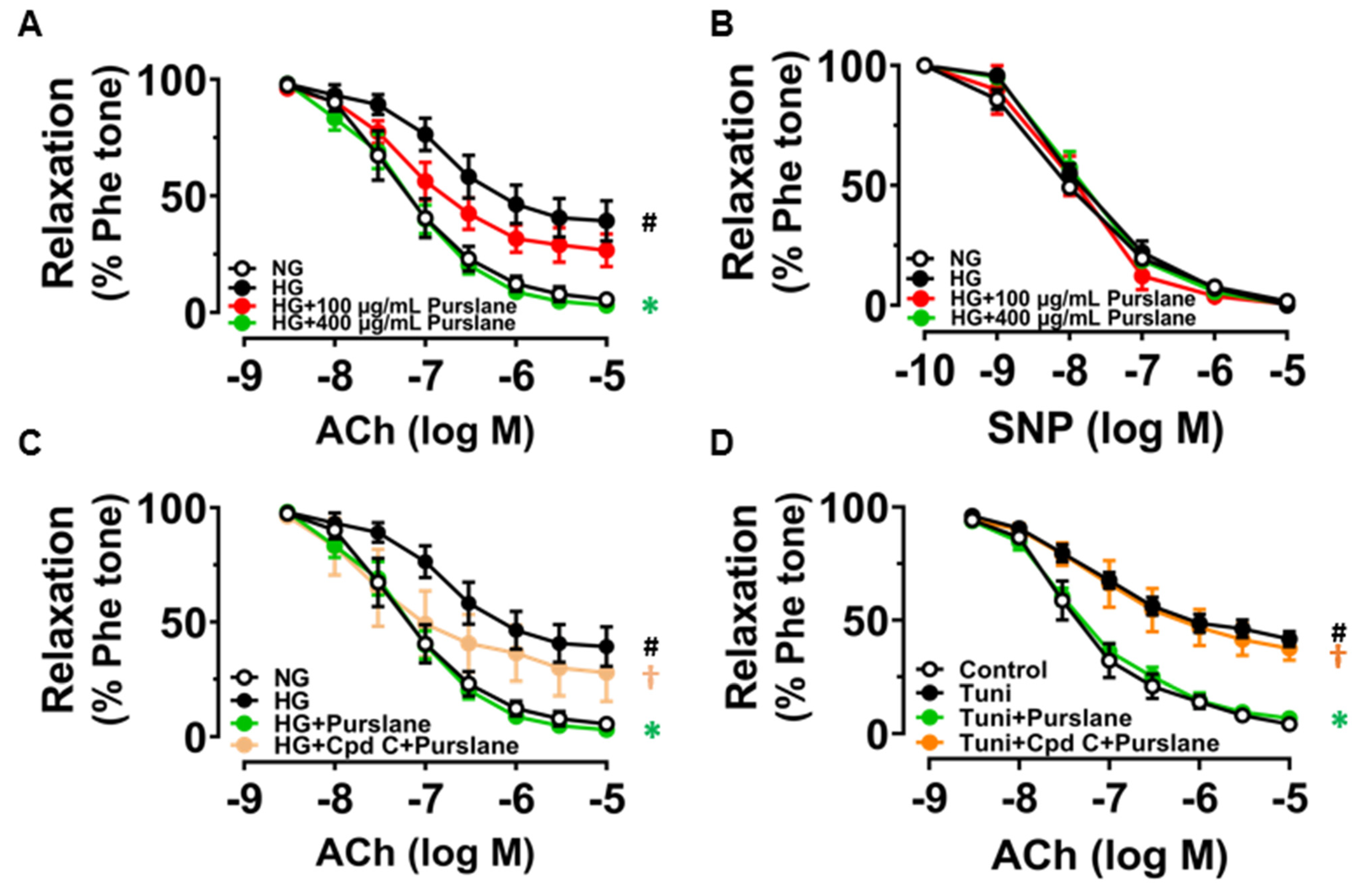
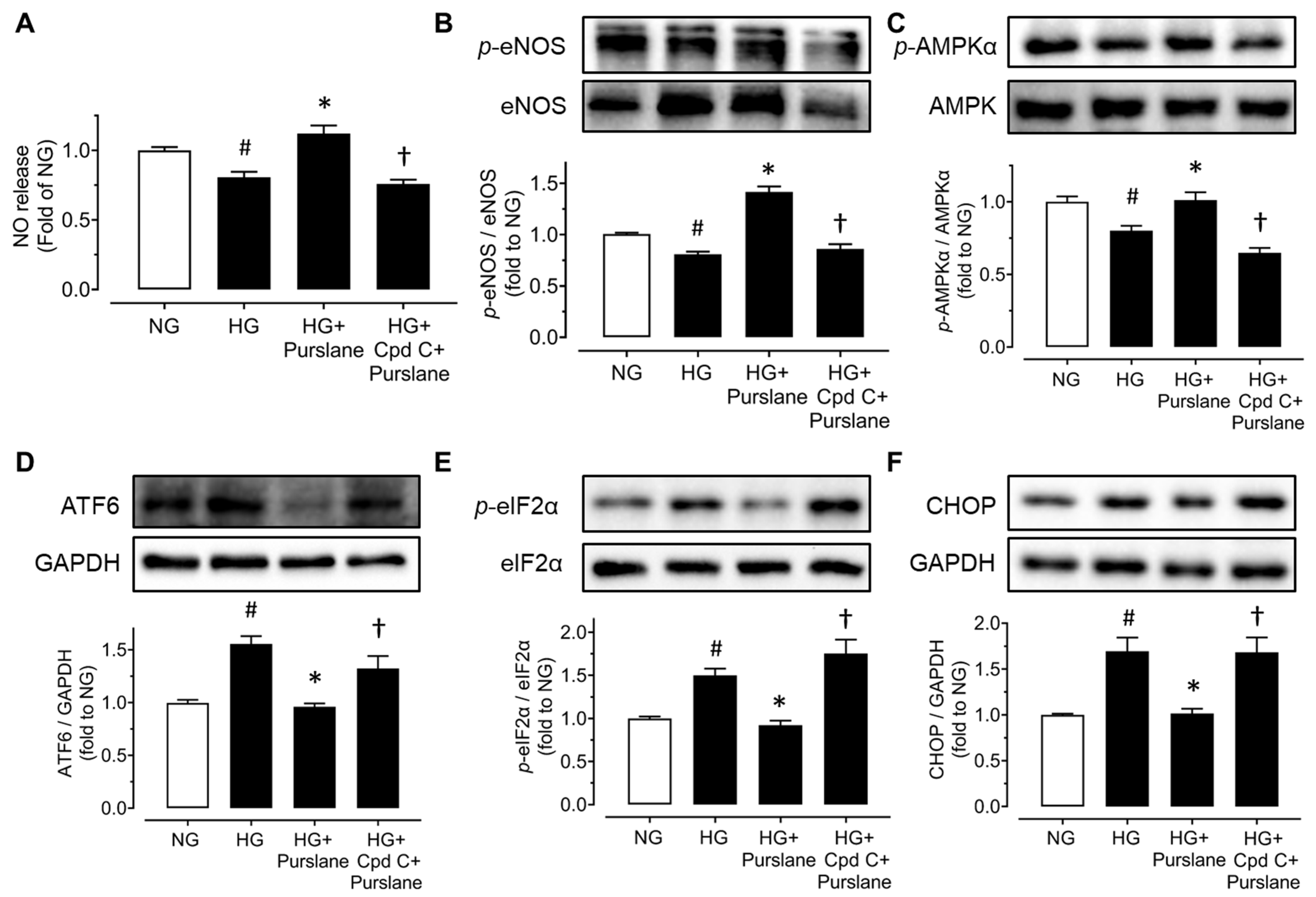
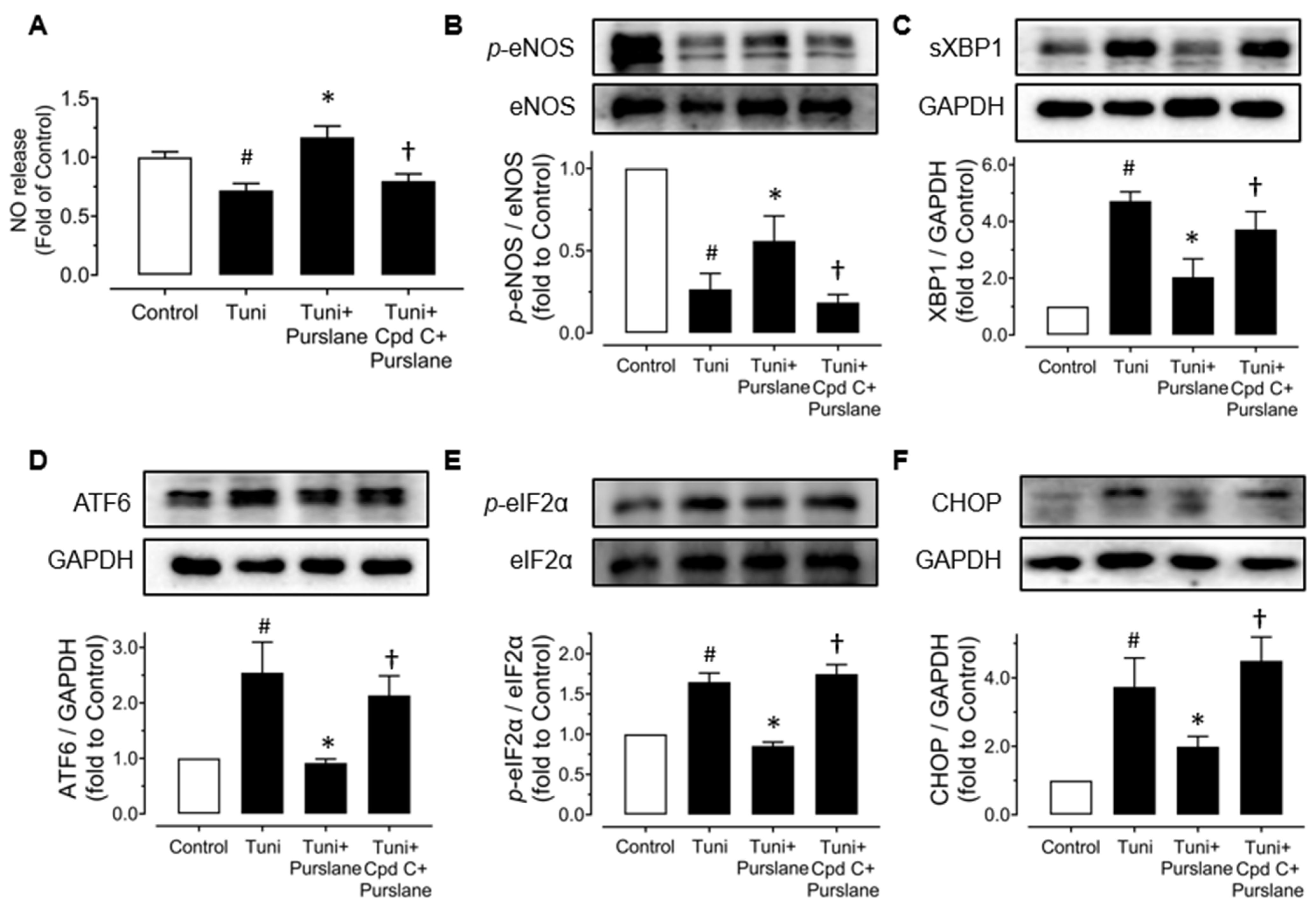
3.4. Purslane Extract Activates AMPK/eNOS Pathway and Alleviates ER Stress in High Glucose/Tunicamycin-Treated Mouse Aortas to Protect Endothelial Function
3.5. Purslane Extract Increases NO Bioavailability via Enhancing AMPK/eNOS Pathway and Reducing ER Stress in RAECs
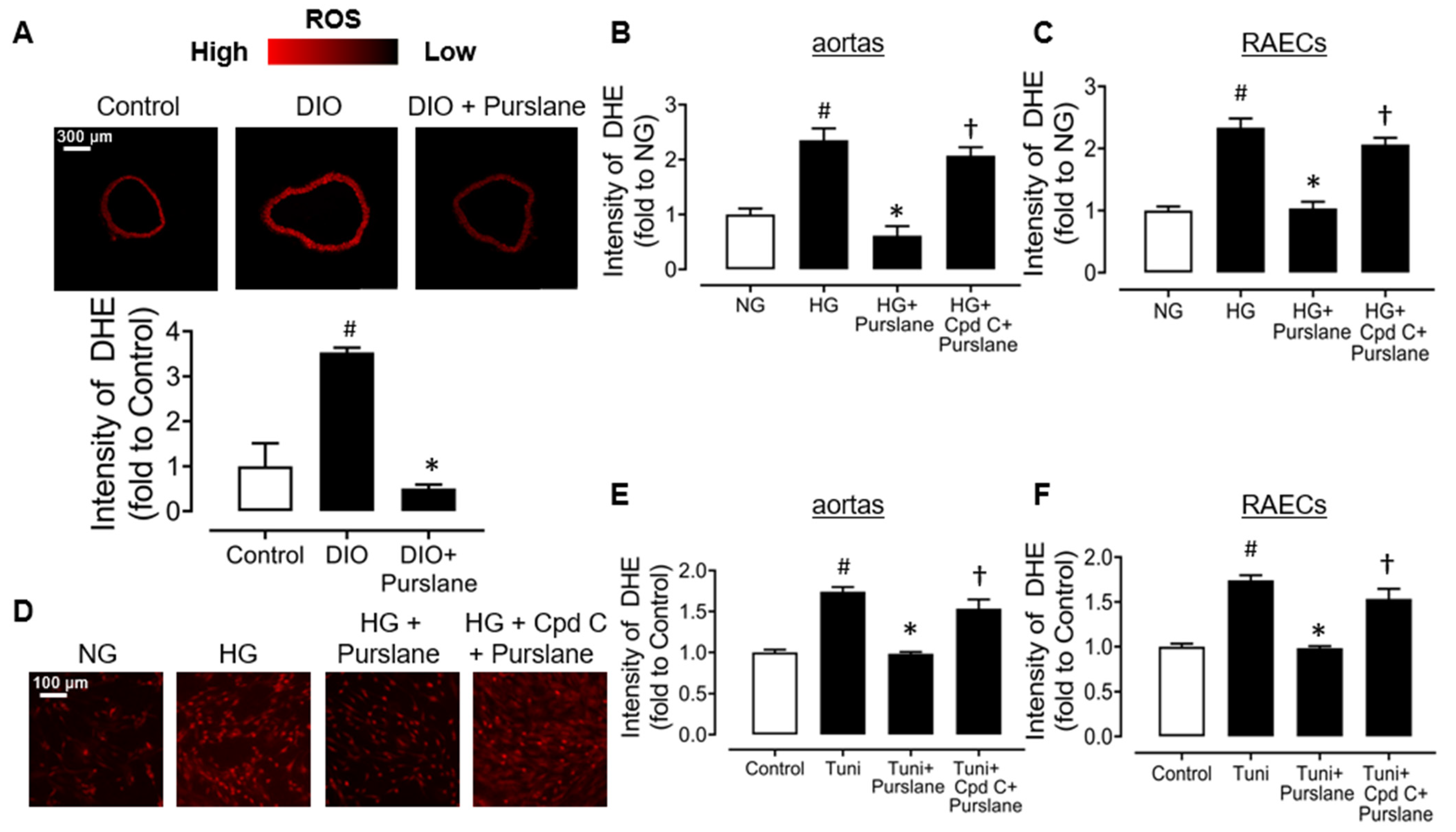
3.6. Purslane Extract Suppresses Oxidative Stress via an AMPK Dependent Mechanism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.; Dellsperger, K.C.; Zhang, C. The link between metabolic abnormalities and endothelial dysfunction in type 2 diabetes: An update. Basic. Res. Cardiol. 2012, 107, 237. [Google Scholar] [CrossRef] [PubMed]
- Jafari, A.; Zadehahmad, Z.; Armanmehr, V.; Talebi, M.; Tehrani, H. The evaluation of the role of diabetes health literacy and health locus of control on quality of life among type 2 diabetes using the Path analysis. Sci. Rep. 2023, 13, 5447. [Google Scholar] [CrossRef] [PubMed]
- Jafari, A.; Tehrani, H.; Mansourian, M.; Nejatian, M.; Gholian-Aval, M. Translation and localization the Persian version of diabetes distress scale among type 2 diabetes. Diabetol. Metab. Syndr. 2023, 15, 201. [Google Scholar] [CrossRef] [PubMed]
- Cyr, A.R.; Huckaby, L.V.; Shiva, S.S.; Zuckerbraun, B.S. Nitric Oxide and Endothelial Dysfunction. Crit. Care Clin. 2020, 36, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Clyne, A.M. Endothelial response to glucose: Dysfunction, metabolism, and transport. Biochem. Soc. Trans. 2021, 49, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, J.; Lu, Q.; Ren, D.; Sun, X.; Rousselle, T.; Tan, Y.; Li, J. AMPK: A therapeutic target of heart failure-not only metabolism regulation. Biosci. Rep. 2019, 39, BSR20181767. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Si, L.Y. Protective effects of AMP-activated protein kinase in the cardiovascular system. J. Cell Mol. Med. 2010, 14, 2604–2613. [Google Scholar] [CrossRef] [PubMed]
- Galan, M.; Kassan, M.; Kadowitz, P.J.; Trebak, M.; Belmadani, S.; Matrougui, K. Mechanism of endoplasmic reticulum stress-induced vascular endothelial dysfunction. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2014, 1843, 1063–1075. [Google Scholar] [CrossRef]
- Kim, I.; Xu, W.; Reed, J.C. Cell death and endoplasmic reticulum stress: Disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2008, 7, 1013–1030. [Google Scholar] [CrossRef]
- Battson, M.L.; Lee, D.M.; Gentile, C.L. Endoplasmic reticulum stress and the development of endothelial dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H355–H367. [Google Scholar] [CrossRef]
- Zhou, Y.; Murugan, D.D.; Khan, H.; Huang, Y.; Cheang, W.S. Roles and Therapeutic Implications of Endoplasmic Reticulum Stress and Oxidative Stress in Cardiovascular Diseases. Antioxidants 2021, 10, 1167. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.X.; Xin, H.L.; Rahman, K.; Wang, S.J.; Peng, C.; Zhang, H. Portulaca oleracea L.: A review of phytochemistry and pharmacological effects. Biomed. Res. Int. 2015, 2015, 925631. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Mo, F.; Ling, C.; Peng, H.; Gu, W.; Li, M.; Chen, Z. Portulaca oleracea L. alleviates liver injury in streptozotocin-induced diabetic mice. Drug Des. Devel Ther. 2018, 12, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Hwang, S.B.; Park, H.J.; Jin, G.R.; Lee, B.H. Antiobesity and Antidiabetic Effects of Portulaca oleracea Powder Intake in High-Fat Diet-Induced Obese C57BL/6 Mice. Evid. Based Complement. Altern. Med. 2021, 2021, 5587848. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zang, X.; Ma, J.; Xu, G. Anti-Diabetic Effect of Portulaca oleracea L. Polysaccharideandits Mechanism in Diabetic Rats. Int. J. Mol. Sci. 2016, 17, 1201. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Hou, K.; Xin, C.; Zeng, D.; Cheng, C.; Zhao, H.; Wang, Z.; Wang, L. Portulaca oleracea L. Extract Alleviated Type 2 Diabetes Via Modulating the Gut Microbiota and Serum Branched-Chain Amino Acid Metabolism. Mol. Nutr. Food Res. 2022, 66, e2101030. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, J.E.; Han, J.S. Portulaca oleracea L. extract reduces hyperglycemia via PI3k/Akt and AMPK pathways in the skeletal muscles of C57BL/Ksj-db/db mice. J. Ethnopharmacol. 2020, 260, 112973. [Google Scholar] [CrossRef] [PubMed]
- Baradaran Rahimi, V.; Rakhshandeh, H.; Raucci, F.; Buono, B.; Shirazinia, R.; Samzadeh Kermani, A.; Maione, F.; Mascolo, N.; Askari, V.R. Anti-Inflammatory and Anti-Oxidant Activity of Portulaca oleracea Extract on LPS-Induced Rat Lung Injury. Molecules 2019, 24, 139. [Google Scholar] [CrossRef]
- Miao, L.; Tao, H.; Peng, Y.; Wang, S.; Zhong, Z.; El-Seedi, H.; Dragan, S.; Zengin, G.; Cheang, W.S.; Wang, Y.; et al. The anti-inflammatory potential of Portulaca oleracea L. (purslane) extract by partial suppression on NF-κB and MAPK activation. Food Chem. 2019, 290, 239–245. [Google Scholar] [CrossRef]
- Lee, A.S.; Lee, Y.J.; Lee, S.M.; Yoon, J.J.; Kim, J.S.; Kang, D.G.; Lee, H.S. Portulaca oleracea Ameliorates Diabetic Vascular Inflammation and Endothelial Dysfunction in db/db Mice. Evid.-Based Complement. Altern. Med. 2012, 2012, 741824. [Google Scholar] [CrossRef]
- Zhou, C.; Tan, Y.; Xu, B.; Wang, Y.; Cheang, W.-S. 3,4′,5-Trimethoxy-trans-stilbene Alleviates Endothelial Dysfunction in Diabetic and Obese Mice via Activation of the AMPK/SIRT1/eNOS Pathway. Antioxidants 2022, 11, 1286. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Borji, A.; Farkhondeh, T. Attenuation of Oxidative Stress and Inflammation by Portulaca oleracea in Streptozotocin-Induced Diabetic Rats. J. Evid.-Based Integr. Med. 2017, 22, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, B.K.; Schaalan, M.F.; Tolba, A.M. Hypoglycemic and pancreatic protective effects of Portulaca oleracea extract in alloxan induced diabetic rats. BMC Complement. Altern. Med. 2017, 17, 37. [Google Scholar] [CrossRef] [PubMed]
- Wainstein, J.; Landau, Z.; Bar Dayan, Y.; Jakubowicz, D.; Grothe, T.; Perrinjaquet-Moccetti, T.; Boaz, M. Purslane Extract and Glucose Homeostasis in Adults with Type 2 Diabetes: A Double-Blind, Placebo-Controlled Clinical Trial of Efficacy and Safety. J. Med. Food 2016, 19, 133–140. [Google Scholar] [CrossRef]
- Lee, A.S.; Lee, Y.J.; Lee, S.M.; Yoon, J.J.; Kim, J.S.; Kang, D.G.; Lee, H.S. An Aqueous Extract of Portulaca oleracea Ameliorates Diabetic Nephropathy Through Suppression of Renal Fibrosis and Inflammation in Diabetic db/db Mice. Am. J. Chin. Med. 2012, 40, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, S.; Mashatan, N.; Farmani, E.; Khodashenas, M.; Musazadeh, V.; Ahrabi, S.S.; Moridpour, A.H.; Faghfouri, A.H. The effects of purslane (Portulaca oleracea) on glycemic indices: A GRADE-assessed systematic review and meta-analysis of randomized controlled trials. Phytother. Res. 2023, 37, 5529–5540. [Google Scholar] [CrossRef] [PubMed]
- Zanella, M.T.; Kohlmann, O., Jr.; Ribeiro, A.B. Treatment of obesity hypertension and diabetes syndrome. Hypertension 2001, 38, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lee, T.S.; Kolb, E.M.; Sun, K.; Lu, X.; Sladek, F.M.; Kassab, G.S.; Garland, T., Jr.; Shyy, J.Y. AMP-activated protein kinase is involved in endothelial NO synthase activation in response to shear stress. Arter. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1281–1287. [Google Scholar] [CrossRef]
- Li, H.; Forstermann, U. Nitric oxide in the pathogenesis of vascular disease. J. Pathol. 2000, 190, 244–254. [Google Scholar] [CrossRef]
- Deng, G.; Long, Y.; Yu, Y.R.; Li, M.R. Adiponectin directly improves endothelial dysfunction in obese rats through the AMPK-eNOS Pathway. Int. J. Obes 2010, 34, 165–171. [Google Scholar] [CrossRef]
- Han, F.; Zhang, S.; Hou, N.; Wang, D.; Sun, X. Irisin improves endothelial function in obese mice through the AMPK-eNOS pathway. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1501–H1508. [Google Scholar] [CrossRef] [PubMed]
- Minamino, T.; Komuro, I.; Kitakaze, M. Endoplasmic reticulum stress as a therapeutic target in cardiovascular disease. Circ. Res. 2010, 107, 1071–1082. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Hare, J.M.; Stamler, J.S. NO/redox disequilibrium in the failing heart and cardiovascular system. J. Clin. Invest. 2005, 115, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Ceolotto, G.; Gallo, A.; Papparella, I.; Franco, L.; Murphy, E.; Iori, E.; Pagnin, E.; Fadini, G.P.; Albiero, M.; Semplicini, A.; et al. Rosiglitazone reduces glucose-induced oxidative stress mediated by NAD(P)H oxidase via AMPK-dependent mechanism. Arter. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2627–2633. [Google Scholar] [CrossRef]
- Jalali, J.; Ghasemzadeh Rahbardar, M. Ameliorative effects of Portulaca oleracea L. (purslane) on the metabolic syndrome: A review. J. Ethnopharmacol. 2022, 299, 115672. [Google Scholar] [CrossRef] [PubMed]
- Iranshahy, M.; Javadi, B.; Iranshahi, M.; Jahanbakhsh, S.P.; Mahyari, S.; Hassani, F.V.; Karimi, G. A review of traditional uses, phytochemistry and pharmacology of Portulaca oleracea L. J. Ethnopharmacol. 2017, 205, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Feng, J.; Zhang, J.; Kang, X.; Qian, D. Quercetin modulates AMPK/SIRT1/NF-κB signaling to inhibit inflammatory/oxidative stress responses in diabetic high fat diet-induced atherosclerosis in the rat carotid artery. Exp. Ther. Med. 2020, 20, 280. [Google Scholar] [CrossRef]
- Miao, L.; Cheong, M.S.; Zhou, C.; Farag, M.; Cheang, W.S.; Xiao, J. Apigenin alleviates diabetic endothelial dysfunction through activating AMPK/PI3K/Akt/eNOS and Nrf2/HO-1 signaling pathways. Food Front. 2023, 4, 420–431. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Retention Time, min | Observed Mass, m/z | Theoretical Mass, m/z | Predicted Chemical Formula | Error, ppm | Observed MS/MS Fragment Ions | Method of Identification | Chemical Compound Name/CAS No. |
|---|---|---|---|---|---|---|---|---|
| 1 | 4.78 | 353.0877 | 353.0873 | C16H18O9 | 1.1 | 135.0446, 191.0556, 173.0450, 179.0345 | Standard comparison | Cryptochlorogenic acid 905-99-7 |
| 2 | 6.25 | 447.0933 | 447.0927 | C21H20O11 | 1.3 | 357.0615, 327.0507, 297.0341 | Standard comparison | Luteolin 8-C-glucoside 28608-75-5 |
| 3 | 7.02 | 463.0890 | 463.0877 | C21H20O12 | 2.8 | 300.0281 | Standard comparison | Quercetin 3-galactoside 482-36-0 |
| 4 | 7.71 | 447.0932 | 447.0927 | C21H20O11 | 1.1 | 284.0331, 255.0297, 227.0344 | PubChem database | Kaempferol-3-O-glucoside 480-10-4 |
| 5 | 7.91 | 447.0930 | 447.0927 | C21H20O11 | 0.6 | 285.0399, 284.0324, 151.0033, 257.0457 | PubChem database | Luteolin-7-O-glucoside 5373-11-5 |
| 6 | 9.43 | 285.0391 | 285.0399 | C15H10O6 | −2.8 | 285.0401, 133.0295, 151.0116, 175.0441, 199.0446 | Standard comparison | Luteolin 491-70-3 |
| 7 | 9.43 | 301.0356 | 301.0348 | C15H10O7 | 2.7 | 151.0084, 107.0188, 121.0332, 179.0057 | Standard comparison | Quercetin 117-39-5 |
| 8 | 10.69 | 285.0404 | 285.0399 | C15H10O6 | 1.8 | 285.0432, 239.0361, 185.0600 | Standard comparison | Kaempferol 520-18-3 |
| Plasma Levels of | Control | DIO | DIO + Purslane |
|---|---|---|---|
| Total cholesterol (mg/dL) | 74.33 ± 8.16 | 138.4 ± 2.65 # | 123.4 ± 2.56 * |
| Triglycerides (mg/dL) | 101.7 ± 4.43 | 223.7 ± 7.05 # | 188.9 ± 9.95 |
| HDL (mg/dL) | 129.93 ± 8.89 | 97.83 ± 20.11 | 141.92 ± 11.60 |
| LDL (mg/dL) | 11.21 ± 1.55 | 93.19 ± 5.41 # | 26.68 ± 3.09 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, L.; Zhou, C.; Zhang, H.; Cheong, M.S.; Tan, Y.; Wang, Y.; Zhang, X.; Yu, H.; Cheang, W.S. Portulaca Oleracea L. (Purslane) Extract Protects Endothelial Function by Reducing Endoplasmic Reticulum Stress and Oxidative Stress through AMPK Activation in Diabetic Obese Mice. Antioxidants 2023, 12, 2132. https://doi.org/10.3390/antiox12122132
Miao L, Zhou C, Zhang H, Cheong MS, Tan Y, Wang Y, Zhang X, Yu H, Cheang WS. Portulaca Oleracea L. (Purslane) Extract Protects Endothelial Function by Reducing Endoplasmic Reticulum Stress and Oxidative Stress through AMPK Activation in Diabetic Obese Mice. Antioxidants. 2023; 12(12):2132. https://doi.org/10.3390/antiox12122132
Chicago/Turabian StyleMiao, Lingchao, Chunxiu Zhou, Haolin Zhang, Meng Sam Cheong, Yi Tan, Yuehan Wang, Xutao Zhang, Hua Yu, and Wai San Cheang. 2023. "Portulaca Oleracea L. (Purslane) Extract Protects Endothelial Function by Reducing Endoplasmic Reticulum Stress and Oxidative Stress through AMPK Activation in Diabetic Obese Mice" Antioxidants 12, no. 12: 2132. https://doi.org/10.3390/antiox12122132