
Buspirone Enhances Cell Survival and Preserves Structural Integrity during Oxidative Injury to the Retinal Pigment Epithelium

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cellular Apoptosis Assay
2.3. Animals
2.4. Real Time PCR
2.5. SD-OCT Imaging and Measurement of Total Retinal Thickness
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
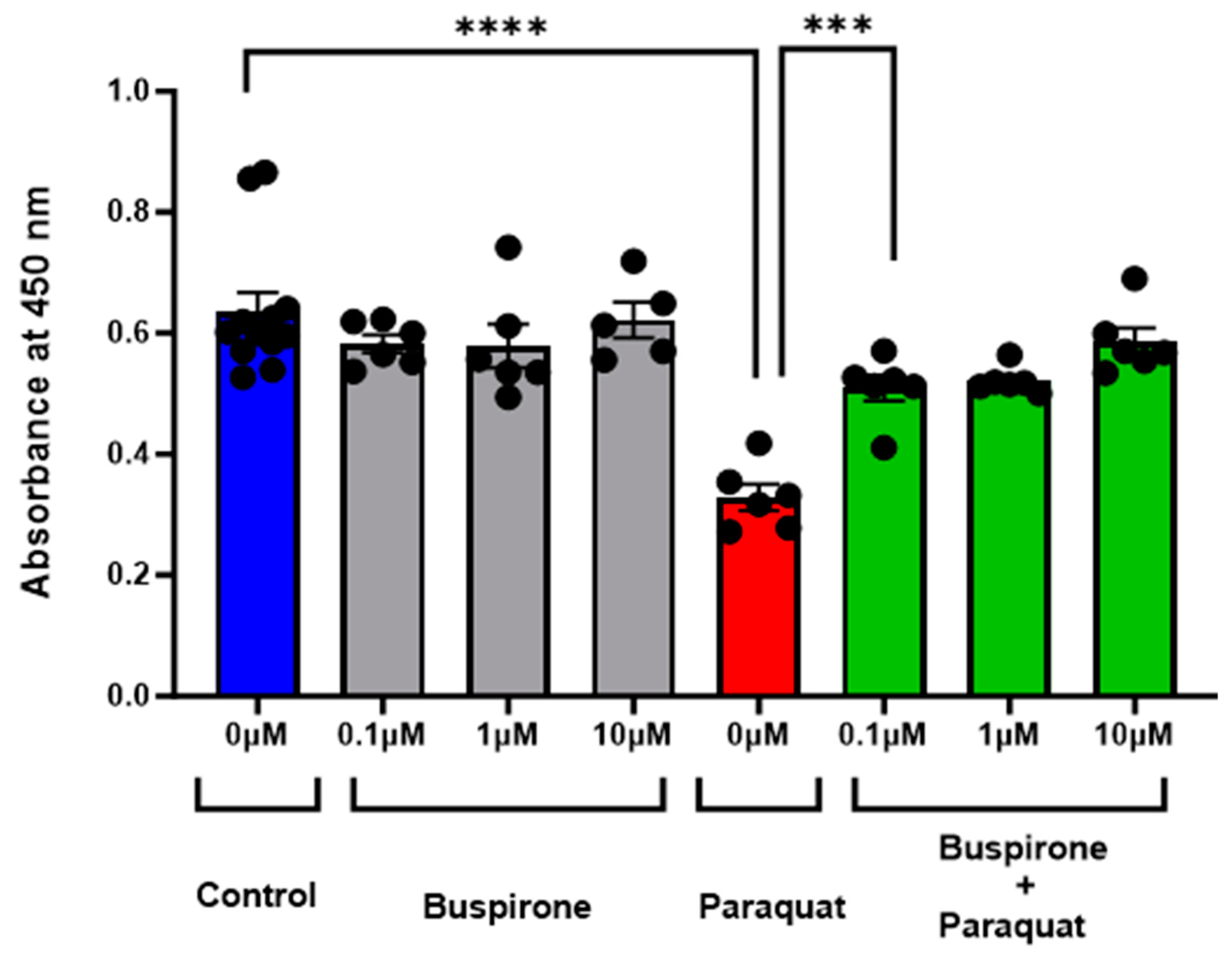
3.1. Buspirone Prevented Oxidative Stress Induced Cell Death
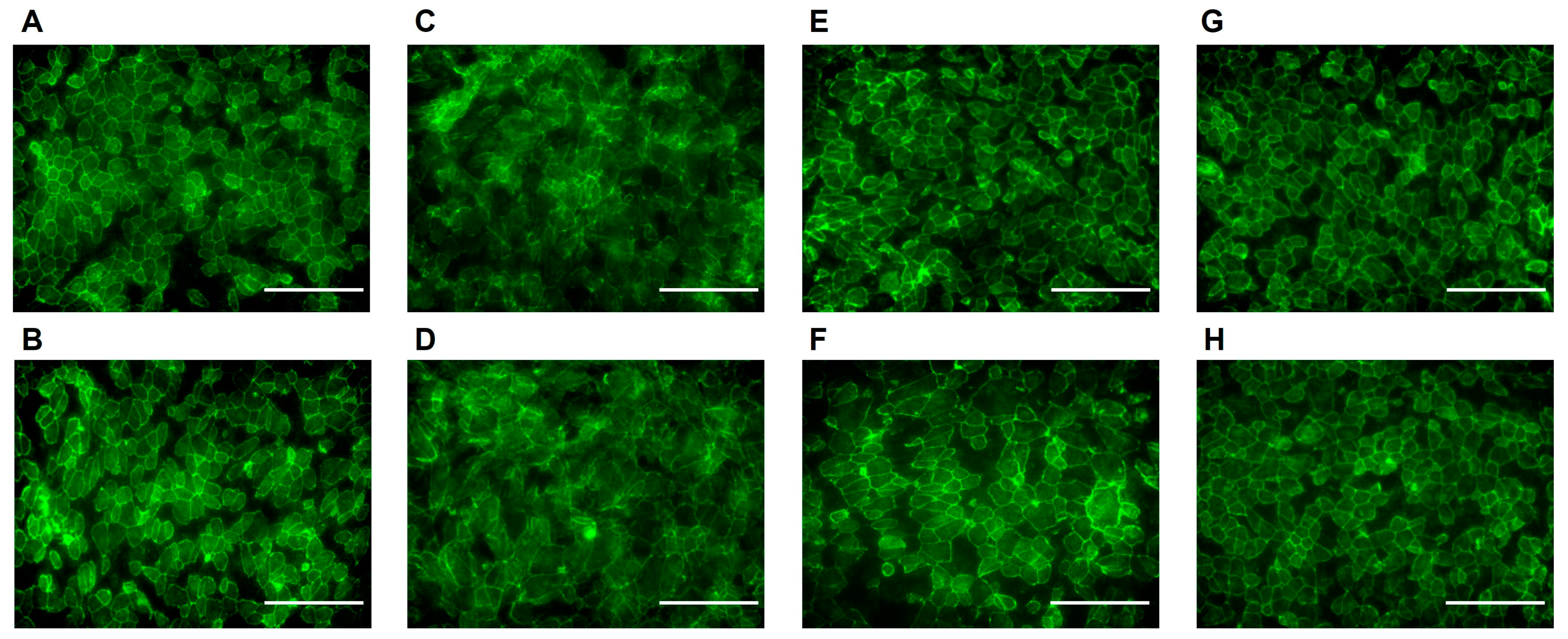
3.2. Buspirone Prevented RPE Structural Disintegration from Oxidative Damage
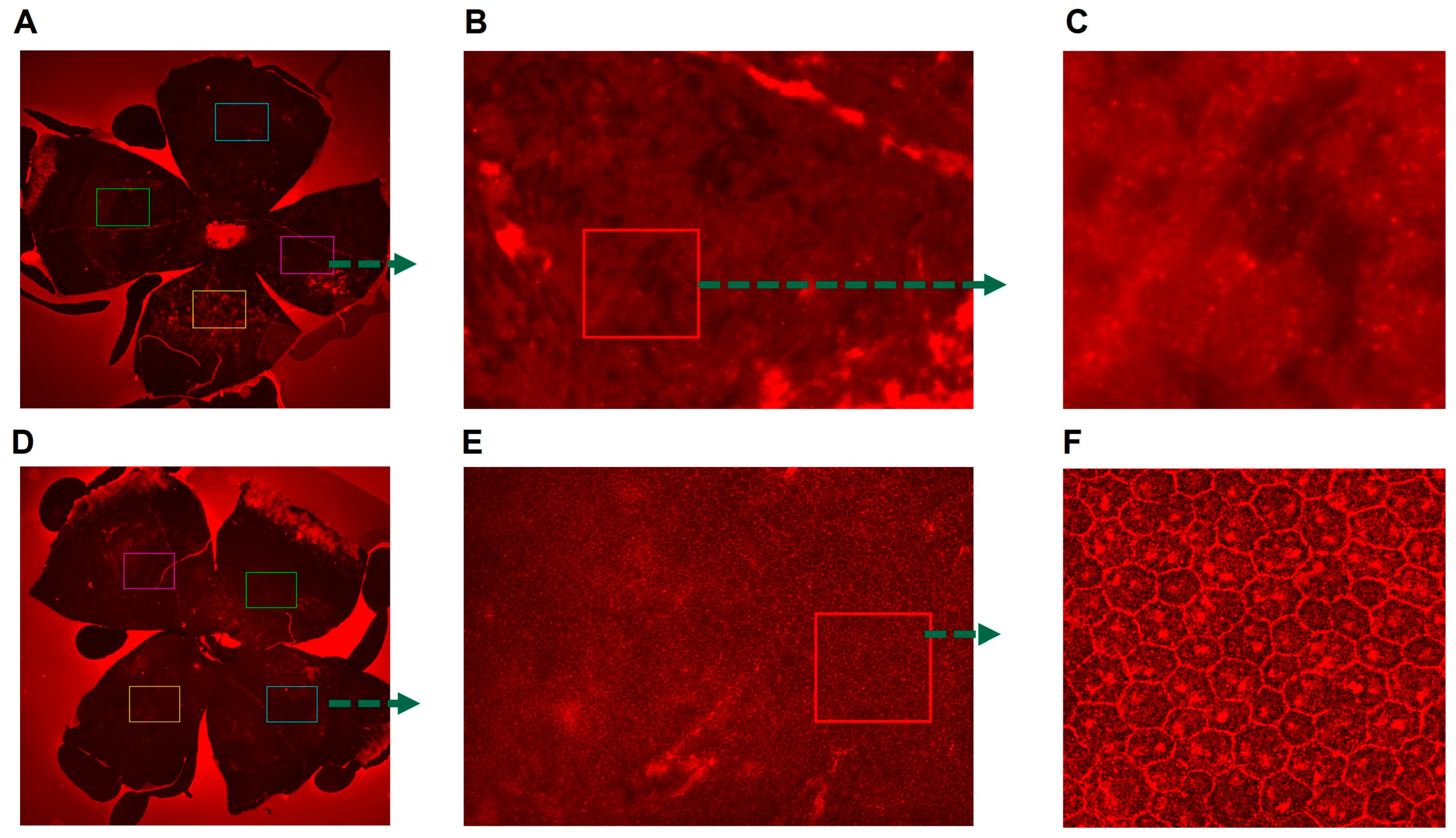
3.3. Presence of Buspirone-Preserved Retinal Structure in an Acute Model of RPE Oxidative Damge
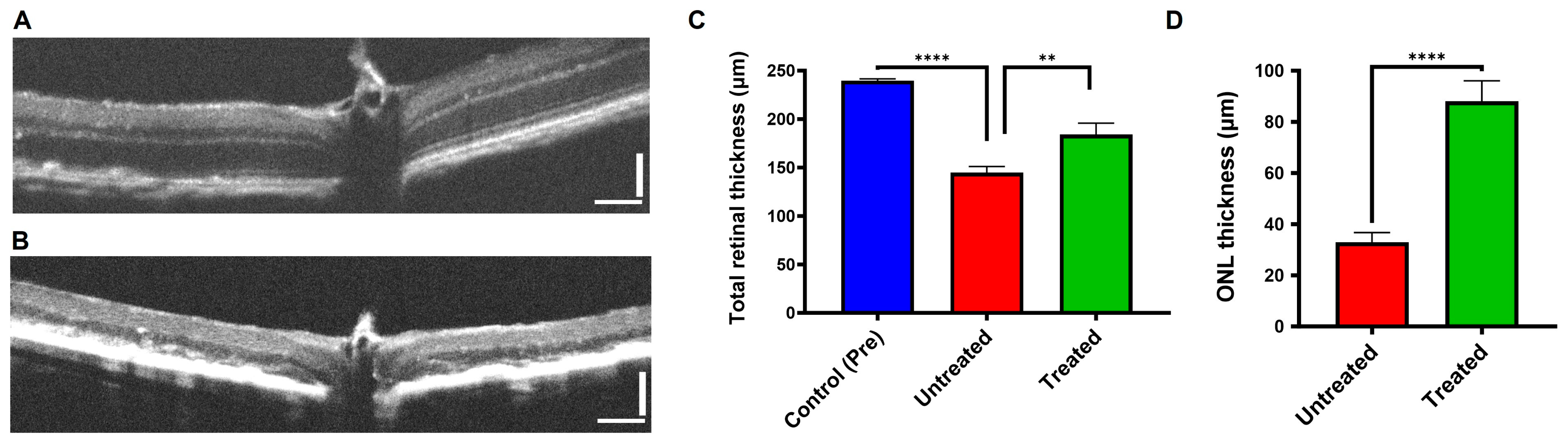
3.4. Buspirone Preserved RPE Structural Integrity In Vivo from Oxidative Damage
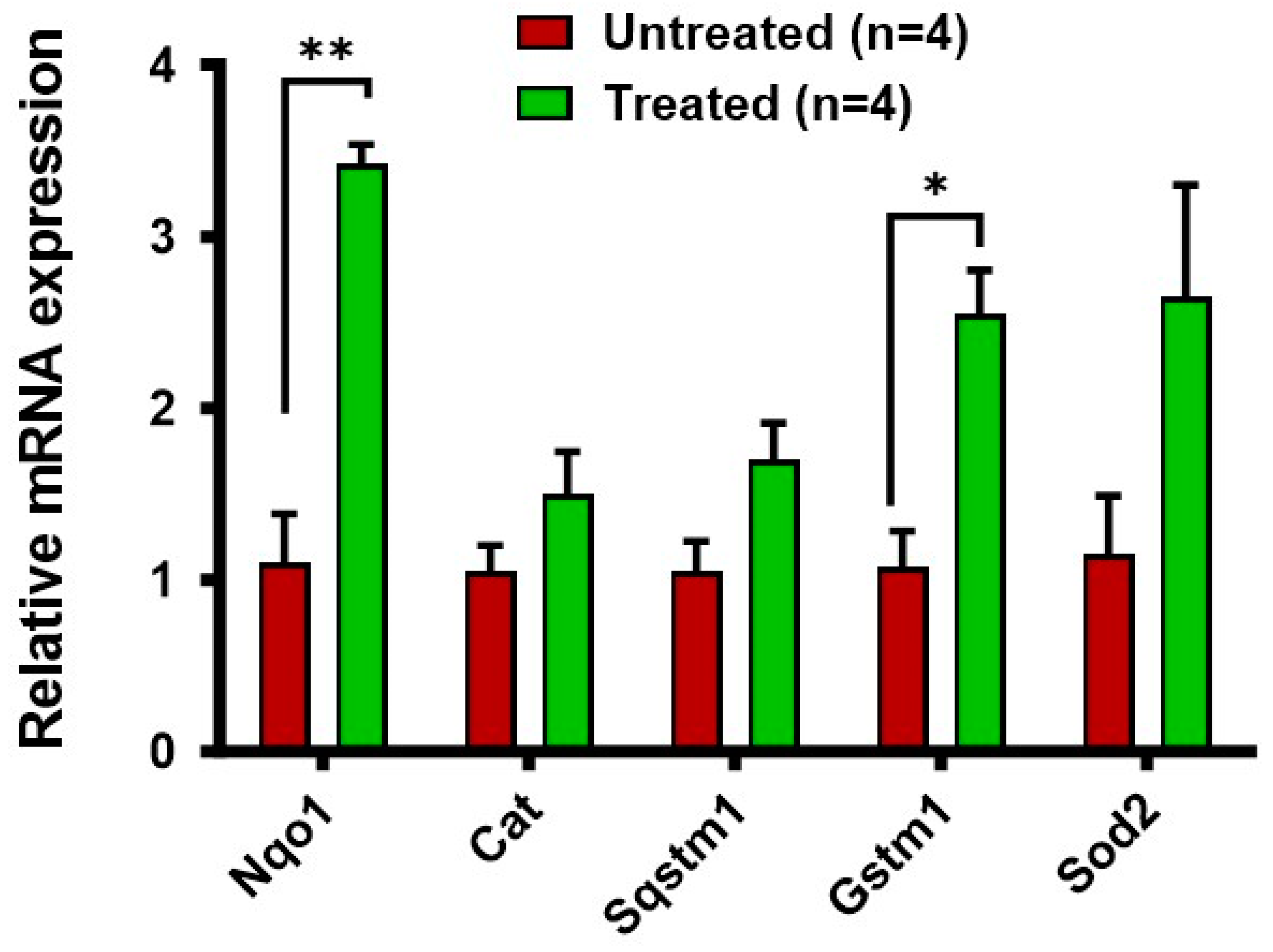
3.5. Increased Expression of Antioxidant Genes in the RPE/Choroid in Response to Buspirone Treatment
4. Discussion
4.1. Buspirone’s Multifaceted Protective Effects in Retinal Health
4.2. Potential Mechanisms Underlying Buspirone’s Protective Effects
4.3. Clinical Implications and Future Directions
4.4. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abokyi, S.; To, C.-H.; Lam, T.T.; Tse, D.Y. Central Role of Oxidative Stress in Age-Related Macular Degeneration: Evidence from a Review of the Molecular Mechanisms and Animal Models. Oxidative Med. Cell. Longev. 2020, 2020, e7901270. [Google Scholar] [CrossRef] [PubMed]
- Piippo, N.; Korhonen, E.; Hytti, M.; Kinnunen, K.; Kaarniranta, K.; Kauppinen, A. Oxidative Stress Is the Principal Contributor to Inflammasome Activation in Retinal Pigment Epithelium Cells with Defunct Proteasomes and Autophagy. CPB 2018, 49, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.P. Buspirone, a New Approach to the Treatment of Anxiety. FASEB J. 1988, 2, 2445–2452. [Google Scholar] [CrossRef] [PubMed]
- Jann, M.W. Buspirone: An Update on a Unique Anxiolytic Agent. Pharmacotherapy 1988, 8, 100–116. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Gupta, M.; Gandhi, R. Buspirone in Autism Spectrum Disorder: A Systematic Review. Cureus 2023, 15, e39304. [Google Scholar] [CrossRef]
- Fernández-Robredo, P.; Sancho, A.; Johnen, S.; Recalde, S.; Gama, N.; Thumann, G.; Groll, J.; García-Layana, A. Current Treatment Limitations in Age-Related Macular Degeneration and Future Approaches Based on Cell Therapy and Tissue Engineering. J. Ophthalmol. 2014, 2014, 510285. [Google Scholar] [CrossRef]
- Gibbs, W.S.; Collier, J.B.; Morris, M.; Beeson, C.C.; Megyesi, J.; Schnellmann, R.G. 5-HT1F Receptor Regulates Mitochondrial Homeostasis and Its Loss Potentiates Acute Kidney Injury and Impairs Renal Recovery. Am. J. Physiol.-Ren. Physiol. 2018, 315, F1119–F1128. [Google Scholar] [CrossRef]
- Eriksen, J.L.; Druse, M.J. Astrocyte-Mediated Trophic Support of Developing Serotonin Neurons: Effects of Ethanol, Buspirone, and S100B. Brain Res. Dev. Brain Res. 2001, 131, 9–15. [Google Scholar] [CrossRef]
- Marazziti, D.; Palego, L.; Giromella, A.; Mazzoni, M.R.; Borsini, F.; Mayer, N.; Naccarato, A.G.; Lucacchini, A.; Cassano, G.B. Region-Dependent Effects of Flibanserin and Buspirone on Adenylyl Cyclase Activity in the Human Brain. Int. J. Neuropsychopharmacol. 2002, 5, 131–140. [Google Scholar] [CrossRef]
- Thomas Broome, S.; Castorina, A. The Anxiolytic Drug Buspirone Prevents Rotenone-Induced Toxicity in a Mouse Model of Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 1845. [Google Scholar] [CrossRef]
- Kumar, A.; Kaur, G.; Rinwa, P. Buspirone along with Melatonin Attenuates Oxidative Damage and Anxiety-like Behavior in a Mouse Model of Immobilization Stress. Chin. J. Nat. Med. 2014, 12, 582–589. [Google Scholar] [CrossRef]
- Kushwah, N.; Bora, K.; Maurya, M.; Pavlovich, M.C.; Chen, J. Oxidative Stress and Antioxidants in Age-Related Macular Degeneration. Antioxidants 2023, 12, 1379. [Google Scholar] [CrossRef] [PubMed]
- Beatty, S.; Koh, H.; Phil, M.; Henson, D.; Boulton, M. The Role of Oxidative Stress in the Pathogenesis of Age-Related Macular Degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.; Siegel, D. Functions of NQO1 in Cellular Protection and CoQ10 Metabolism and Its Potential Role as a Redox Sensitive Molecular Switch. Front. Physiol. 2017, 8, 595. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Du, W.; He, X.; Zhang, D.; He, W. Tribulus Terrestris Ameliorates Oxidative Stress-Induced ARPE-19 Cell Injury through the PI3K/Akt-Nrf2 Signaling Pathway. Oxidative Med. Cell. Longev. 2020, 2020, 7962393. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Barrio, C.; Del Olmo-Aguado, S.; García-Pérez, E.; de la Fuente, M.; Muruzabal, F.; Anitua, E.; Baamonde-Arbaiza, B.; Fernández-Vega-Cueto, L.; Fernández-Vega, L.; Merayo-Lloves, J. Antioxidant Role of PRGF on RPE Cells after Blue Light Insult as a Therapy for Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 1021. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Parsons, K.K.; Chi, L.; Malakauskas, S.M.; Le, T.H. Glutathione S-Transferase-Μ1 Regulates Vascular Smooth Muscle Cell Proliferation, Migration, and Oxidative Stress. Hypertension 2009, 54, 1360–1368. [Google Scholar] [CrossRef]
- Sandbach, J.M.; Coscun, P.E.; Grossniklaus, H.E.; Kokoszka, J.E.; Newman, N.J.; Wallace, D.C. Ocular Pathology in Mitochondrial Superoxide Dismutase (Sod2)-Deficient Mice. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2173–2178. [Google Scholar]
- Kasahara, E.; Lin, L.-R.; Ho, Y.-S.; Reddy, V.N. SOD2 Protects against Oxidation-Induced Apoptosis in Mouse Retinal Pigment Epithelium: Implications for Age-Related Macular Degeneration. Investig. Ophthalmol. Vis. Sci. 2005, 46, 3426–3434. [Google Scholar] [CrossRef]
- Collier, R.J.; Patel, Y.; Martin, E.A.; Dembinska, O.; Hellberg, M.; Krueger, D.S.; Kapin, M.A.; Romano, C. Agonists at the Serotonin Receptor (5-HT(1A)) Protect the Retina from Severe Photo-Oxidative Stress. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2118–2126. [Google Scholar] [CrossRef]
- Biswal, M.R.; Ahmed, C.M.; Ildefonso, C.J.; Han, P.; Li, H.; Jivanji, H.; Mao, H.; Lewin, A.S. Systemic Treatment with a 5HT1a Agonist Induces Anti-Oxidant Protection and Preserves the Retina from Mitochondrial Oxidative Stress. Exp. Eye Res. 2015, 140, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, C.M.; Biswal, M.R.; Li, H.; Han, P.; Ildefonso, C.J.; Lewin, A.S. Repurposing an Orally Available Drug for the Treatment of Geographic Atrophy. Mol. Vis. 2016, 22, 294–310. [Google Scholar] [PubMed]
- Coyner, A.S.; Ryals, R.C.; Ku, C.A.; Fischer, C.M.; Patel, R.C.; Datta, S.; Yang, P.; Wen, Y.; Hen, R.; Pennesi, M.E. Retinal Neuroprotective Effects of Flibanserin, an FDA-Approved Dual Serotonin Receptor Agonist-Antagonist. PLoS ONE 2016, 11, e0159776. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Wang, C.; Zhang, Y.; Su, G. Morphologic and Histopathologic Change of Sodium Iodate-Induced Retinal Degeneration in Adult Rats. Int. J. Clin. Exp. Pathol. 2019, 12, 443–454. [Google Scholar] [PubMed]
- Upadhyay, M.; Bonilha, V.L. Regulated Cell Death Pathways in the Sodium Iodate Model: Insights and Implications for AMD. Exp. Eye Res. 2023, 238, 109728. [Google Scholar] [CrossRef] [PubMed]
- Hazim, R.A.; Volland, S.; Yen, A.; Burgess, B.L.; Williams, D.S. Rapid Differentiation of the Human RPE Cell Line, ARPE-19, Induced by Nicotinamide. Exp. Eye Res. 2019, 179, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Dunn, K.C.; Aotaki-Keen, A.E.; Putkey, F.R.; Hjelmeland, L.M. ARPE-19, a Human Retinal Pigment Epithelial Cell Line with Differentiated Properties. Exp. Eye Res. 1996, 62, 155–169. [Google Scholar] [CrossRef]
- Chen, W.; Lin, B.; Xie, S.; Yang, W.; Lin, J.; Li, Z.; Zhan, Y.; Gui, S.; Lin, B. Naringenin Protects RPE Cells from NaIO3-Induced Oxidative Damage in Vivo and in Vitro through up-Regulation of SIRT1. Phytomedicine 2021, 80, 153375. [Google Scholar] [CrossRef]
- Juel, H.B.; Faber, C.; Svendsen, S.G.; Vallejo, A.N.; Nissen, M.H. Inflammatory Cytokines Protect Retinal Pigment Epithelial Cells from Oxidative Stress-Induced Death. PLoS ONE 2013, 8, e64619. [Google Scholar] [CrossRef]
- Xie, R.; Wang, B.; Zuo, S.; Du, M.; Wang, X.; Yu, Y.; Yan, H. Protective Effects of CRTH2 Suppression in Dry Age-Related Macular Degeneration. Biochem. Biophys. Res. Commun. 2022, 624, 8–15. [Google Scholar] [CrossRef]
- Ma, H.; Yang, F.; Ding, X.-Q. Inhibition of Thyroid Hormone Signaling Protects Retinal Pigment Epithelium and Photoreceptors from Cell Death in a Mouse Model of Age-Related Macular Degeneration. Cell Death Dis. 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Bardak, H.; Uğuz, A.C.; Bardak, Y.; Rocha-Pimienta, J.; Delgado-Adámez, J.; Espino, J. Selenium Protects ARPE-19 and ACBRI 181 Cells against High Glucose-Induced Oxidative Stress. Molecules 2023, 28, 5961. [Google Scholar] [CrossRef] [PubMed]
- Hanneken, A.; Lin, F.-F.; Johnson, J.; Maher, P. Flavonoids Protect Human Retinal Pigment Epithelial Cells from Oxidative-Stress–Induced Death. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3164–3177. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Kuriyan, A.E.; Su, C.-W.; Mahabole, M.; Zhang, Y.; Zhu, Y.-T.; Flynn, H.W.; Parel, J.-M.; Tseng, S.C.G. Inhibition of Proliferation and Epithelial Mesenchymal Transition in Retinal Pigment Epithelial Cells by Heavy Chain-Hyaluronan/Pentraxin 3. Sci. Rep. 2017, 7, srep43736. [Google Scholar] [CrossRef] [PubMed]
- Haranahalli Shivarudrappa, A.; Gopal, S.S.; Ponesakki, G. An in Vitro Protocol to Study the Effect of Hyperglycemia on Intracellular Redox Signaling in Human Retinal Pigment Epithelial (ARPE-19) Cells. Mol. Biol. Rep. 2019, 46, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Biswal, M.R.; Justis, B.D.; Han, P.; Li, H.; Gierhart, D.; Dorey, C.K.; Lewin, A.S. Daily Zeaxanthin Supplementation Prevents Atrophy of the Retinal Pigment Epithelium (RPE) in a Mouse Model of Mitochondrial Oxidative Stress. PLoS ONE 2018, 13, e0203816. [Google Scholar] [CrossRef]
- Wenzel, A.A.; O’Hare, M.N.; Shadmand, M.; Corson, T.W. Optical Coherence Tomography Enables Imaging of Tumor Initiation in the TAg-RB Mouse Model of Retinoblastoma. Mol. Vis. 2015, 21, 515–522. [Google Scholar]
- Zou, X.-L.; Wang, G.-F.; Li, D.-D.; Chen, J.-X.; Zhang, C.-L.; Yu, Y.-Z.; Zhou, W.-J.; Zou, Y.-P.; Rao, B.-Q. Protection of Tight Junction between RPE Cells with Tissue Factor Targeting Peptide. Int. J. Ophthalmol. 2018, 11, 1594–1599. [Google Scholar] [CrossRef]
- Obert, E.; Strauss, R.; Brandon, C.; Grek, C.; Ghatnekar, G.; Gourdie, R.; Rohrer, B. Targeting The Tight Junction Protein, Zonula Occludens-1, With The Connexin 43 Mimetic Peptide, αCT1, Reduces VEGF-Dependent RPE Pathophysiology. J. Mol. Med. 2017, 95, 535–552. [Google Scholar] [CrossRef]
- Trakkides, T.-O.; Schäfer, N.; Reichenthaler, M.; Kühn, K.; Brandwijk, R.J.M.G.E.; Toonen, E.J.M.; Urban, F.; Wegener, J.; Enzmann, V.; Pauly, D. Oxidative Stress Increases Endogenous Complement-Dependent Inflammatory and Angiogenic Responses in Retinal Pigment Epithelial Cells Independently of Exogenous Complement Sources. Antioxidants 2019, 8, 548. [Google Scholar] [CrossRef]
- Frede, K.; Ebert, F.; Kipp, A.P.; Schwerdtle, T.; Baldermann, S. Lutein Activates the Transcription Factor Nrf2 in Human Retinal Pigment Epithelial Cells. J. Agric. Food Chem. 2017, 65, 5944–5952. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.A.; Ryals, R.C.; Jiang, D.; Coyner, A.S.; Weller, K.K.; Sinha, W.; Robb, B.M.; Yang, P.; Pennesi, M.E. The Role of ERK1/2 Activation in Sarpogrelate-Mediated Neuroprotection. Investig. Ophthalmol. Vis. Sci. 2018, 59, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Tullis, B.E.; Ryals, R.C.; Coyner, A.S.; Gale, M.J.; Nicholson, A.; Ku, C.; Regis, D.; Sinha, W.; Datta, S.; Wen, Y.; et al. Sarpogrelate, a 5-HT2A Receptor Antagonist, Protects the Retina From Light-Induced Retinopathy. Investig. Ophthalmol. Vis. Sci. 2015, 56, 4560–4569. [Google Scholar] [CrossRef] [PubMed]
- Urbina, M.; Schmeer, C.; Lima, L. 5HT1A Receptor Agonist Differentially Increases Cyclic AMP Concentration in Intact and Lesioned Goldfish Retina. In Vitro Inhibition of Outgrowth by Forskolin. Neurochem. Int. 1996, 29, 453–460. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Ref. Seq | Forward Primer | Reverse Primer |
|---|---|---|---|
| Beta-Actin | NM_007393.3 | CGAGCACAGCTTCTTTGCAG | TTCCCACCATCACACCCTGG |
| Nqo1 | NM_008706.5 | CGACAACGGTCCTTTCCAGA | CCAGACGGTTTCCAGACGTT |
| Cat | NM_009804.2 | CGCAATCCTACACCATGTCG | AGTATCCAAAAGCACCTGCTCC |
| Sqstm1 | NM_011018.2 | GGAAGCTGCCCTATACCCAC | GCTTGGCCACAGCACTATCA |
| Gstm1 | NM_010358.5 | GGGATACTGGAACGTCCGC | GCTCTGGGTGATCTTGTGTGA |
| Sod2 | NM_013671.3 | CAGGATGCCGCTCCGTTAT | TGAGGTTTACACGACCGCTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biswal, M.R.; Paulson, R.J.; Vichare, R.; Lewin, A.S. Buspirone Enhances Cell Survival and Preserves Structural Integrity during Oxidative Injury to the Retinal Pigment Epithelium. Antioxidants 2023, 12, 2129. https://doi.org/10.3390/antiox12122129
Biswal MR, Paulson RJ, Vichare R, Lewin AS. Buspirone Enhances Cell Survival and Preserves Structural Integrity during Oxidative Injury to the Retinal Pigment Epithelium. Antioxidants. 2023; 12(12):2129. https://doi.org/10.3390/antiox12122129
Chicago/Turabian StyleBiswal, Manas R., Ryan J. Paulson, Riddhi Vichare, and Alfred S. Lewin. 2023. "Buspirone Enhances Cell Survival and Preserves Structural Integrity during Oxidative Injury to the Retinal Pigment Epithelium" Antioxidants 12, no. 12: 2129. https://doi.org/10.3390/antiox12122129