1. Introduction
Chronic kidney disease (CKD) is a prevalent condition that affects approximately 10% of the global population, and its limited treatment options contribute to high mortality rates [
1]. CKD frequently progresses to end-stage renal disease, a life-threatening condition that necessitates renal replacement therapy such as dialysis or kidney transplantation. The development of kidney fibrosis is a key pathological process in CKD. To develop therapies that can prevent or slow the progression of CKD, it is crucial to understand the mechanisms underlying renal fibrosis [
2]. Fibrosis refers to the excessive accumulation of extracellular matrix (ECM) primarily by fibroblasts residing in the kidney tissue [
3,
4]. Normally, the kidneys maintain a minimal amount of ECM to support their structure and function. Upon tissue injury, wound-healing mechanisms are activated to limit inflammation and promote proper tissue regeneration. However, persistent inflammatory responses impede complete regeneration, resulting in fibrotic scar tissue formation. Excessive deposition of ECM during chronic and pathological fibrosis disrupts the normal architecture of the kidney and impairs its function [
5]. Eventually, unresolved kidney fibrosis reaches an irreversible stage and contributes to renal failure.
Extensive research has focused on unraveling the mechanisms involved in the development of kidney fibrosis. Regardless of the causative factors, excessive inflammatory responses are observed in this process. Renal fibrosis is triggered and augmented by the inflammatory response. Multiple cell types, including myeloid-derived inflammatory cells, fibroblasts, pericytes, epithelial cells, and endothelial cells, participate in inflammation [
6]. Recently, an increasing number of studies have focused on the role of tubular epithelial cells (TECs) in renal fibrosis and inflammation [
7,
8,
9]. Once damaged, TECs secrete major cytokines and chemokines that play important roles in fibrosis and inflammation [
7]. TECs directly secrete pro-fibrotic cytokines, including transforming growth factor-β (TGFβ) and connective tissue growth factor, thus activating nearby fibroblasts to produce ECM proteins. Furthermore, TECs secrete major chemokines, including CCL2 and CCL5, that recruit myeloid-derived inflammatory cells [
9]. However, the role of infiltrated myeloid-derived inflammatory cells in fibrosis remains unknown. Interestingly, several recent studies have indicated that bone marrow-derived cells directly contribute to collagen production in kidney fibrosis models [
10].
Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) is a crucial transcription factor involved in the regulation of inflammation. It is present in nearly all types of cells and tissues and responds to various internal and external stimuli. These stimuli include infections, inflammatory cytokines, oxidative stress, and chemical agents [
11]. NF-κB plays a vital role in maintaining normal immune responses against infections; however, when its activation becomes dysregulated, it becomes a significant contributor to inflammatory diseases. NF-κB activation has commonly been observed during kidney injury, whether caused by infection or occurring in a sterile environment [
12,
13]. In models of ischemic acute kidney injury, NF-κB activity has been detected in tubular epithelial cells [
14]. The activation of NF-κB in this context worsens tubular injury and intensifies an inappropriate inflammatory response. Additionally, angiotensin activates NF-κB in tubular epithelial cells, leading to an increased inflammatory response [
15]. Conversely, inhibiting NF-κB activity protects against renal inflammation [
16]. Therefore, targeting NF-κB specifically in tubular epithelial cells represents an interesting approach to the regulation of kidney diseases.
Previous studies have shown that thio-barbiturate-derived compounds have antioxidant effects and prevent LPS-induced inflammation in the liver [
17]. Thio-barbiturate-derived compounds showed significant ROS- and ONOO-scavenging effects in test tubes [
17]. In this study, the authors showed that one of the thio-barbiturate-derived compounds, MHY1025, significantly reduced LPS-induced oxidative stress and NF-κB activation in both macrophage and liver injury models. Moreover, it has been reported by others that barbiturate compounds exhibit antioxidant properties [
18]. However, the effects of these compounds on general aspects of renal fibrosis have not been investigated. Here, we evaluated the antioxidative and anti-inflammatory effects of MHY1025 in renal epithelial cells in vitro. Furthermore, we demonstrated the antifibrotic and anti-inflammatory effects of MHY1025 in mouse models of folic acid-induced renal fibrosis. MHY1025 treatment significantly reduced fibrosis and inflammation in a mouse model of renal fibrosis.
2. Materials and Methods
2.1. Animal Studies
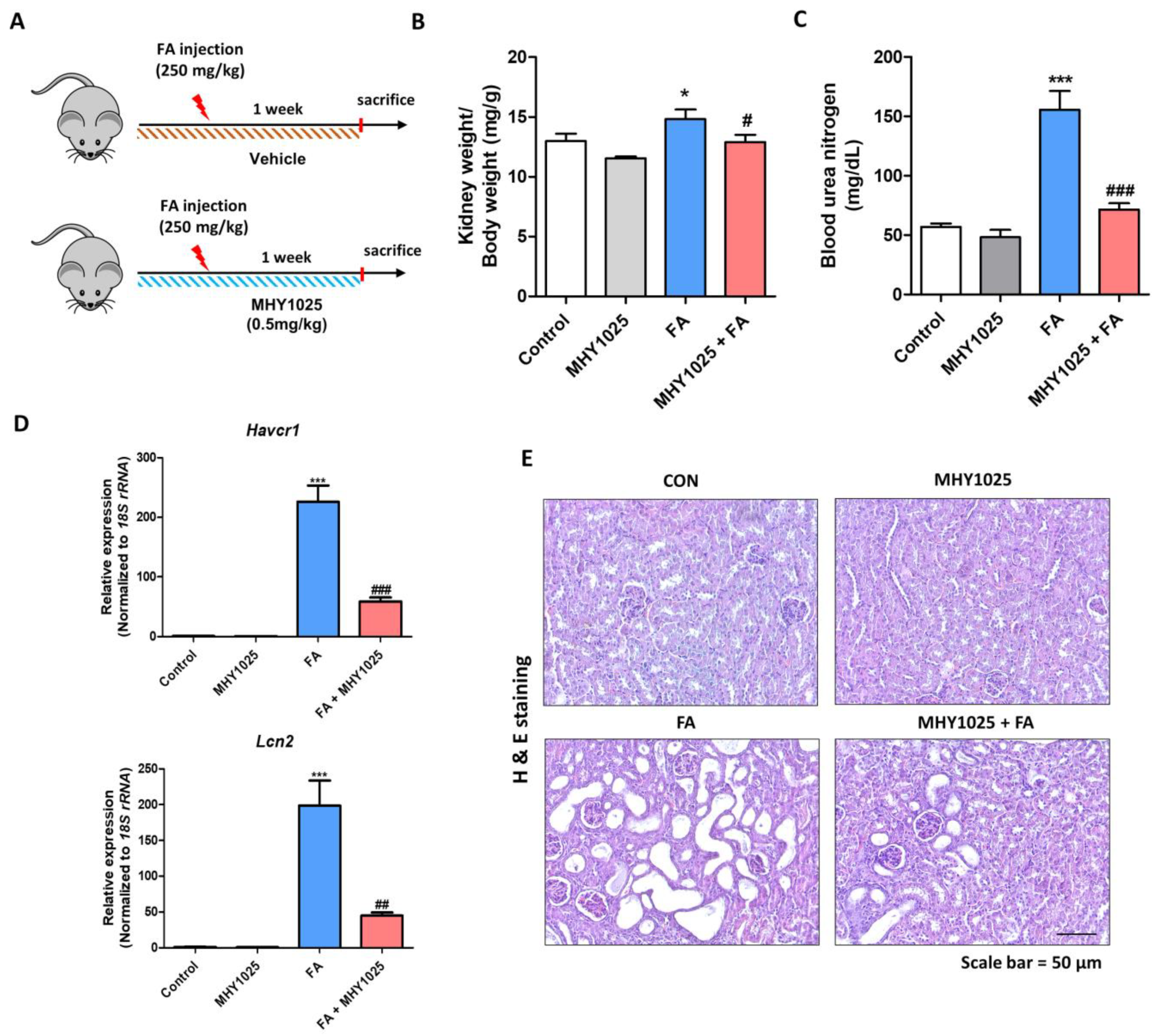
All animal experiments were performed in accordance with the guidelines for animal experimentation issued by Pusan National University (PNU) and were approved by the Institutional Animal Care Committee of PNU (IACUC approval No. PNU-2022-3180). The C57BL/6J mice were purchased from Hyochang Science (Daegu, Korea). All mice were maintained at 23 ± 2 °C with a relative humidity of 60 ± 5% and 12 h light/dark cycles and were provided with free access to water and food. The animals were randomly sorted into four different groups (n = 7~8). To examine the therapeutic effect of MHY1025 in a renal fibrosis model, male mice were intraperitoneally administered 250 mg/kg of folic acid dissolved in 0.3 M NaHCO3 or a vehicle. Throughout the experimental period, a daily oral gavage was administered for one week, delivering either 0.5 mg/kg MHY1025 dissolved in 1% DMSO in water or a solution of 1% DMSO alone. Following the sacrifice of the mice, the kidneys and sera were collected. The kidneys were promptly preserved at −80 °C or fixed in 10% neutral formalin for histochemical analysis. Serum samples were collected by centrifugation at 3000 rpm for 20 min at 4 °C. Blood urea nitrogen (BUN) levels were measured using a commercial assay kit from Shinyang Diagnostics (SICDIA L-BUN, 1120171, Seoul, Korea) according to the manufacturer’s instructions.
2.2. Cell Culture Experiments
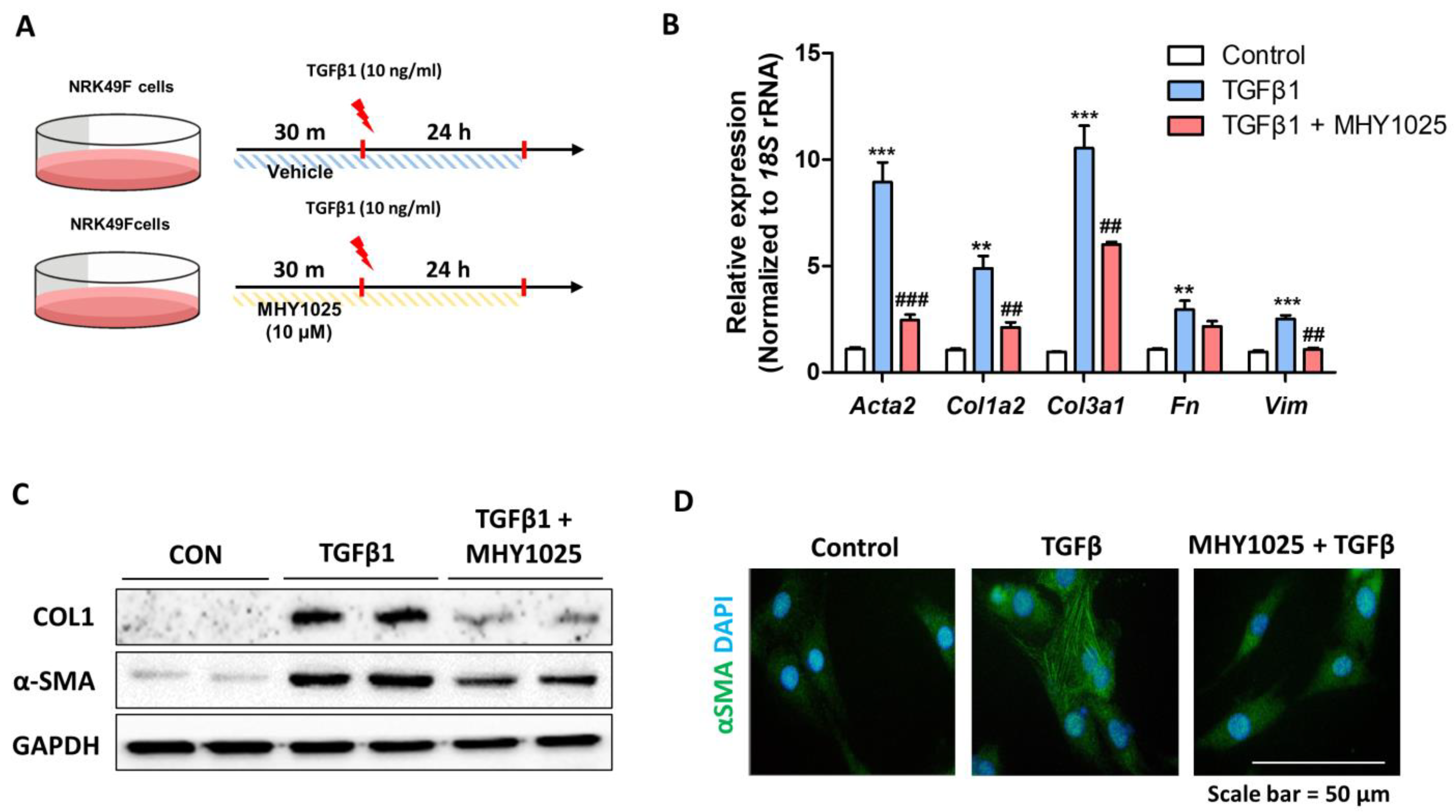
The rat tubular epithelial cell line (NRK52E) and rat kidney fibroblasts (NRK49F) were purchased from the ATCC. All cell lines were cultured in an incubator at 37 °C in humidified 95% air and 5% CO2 in Dulbecco’s modified Eagle’s medium (DMEM) containing 5% fetal bovine serum (FBS) and 1% penicillin. A stock solution of MHY1025 was prepared at a concentration of 10 mM in DMSO and stored at −20 °C until use. To determine the antioxidant and anti-inflammatory effects of MHY1025, LPS (L2630, Sigma, St. Louis, MO, USA) treatment was used in NRK52E cells. To evaluate the inhibitory effect of MHY1025 on fibrotic cell activation, NRK49F cells were pretreated with MHY1025 and then treated with 10 ng/mL of TGF-β1 (100-21, PeproTech, Cranbury, NJ, USA) for 24 h. The concentration range of MHY1025 administered to the cells ranged from 1 to 20 μM, as determined by the experimental conditions. All the cell culture experiments were repeated at least 3 times.
2.3. Cell Viability Assay
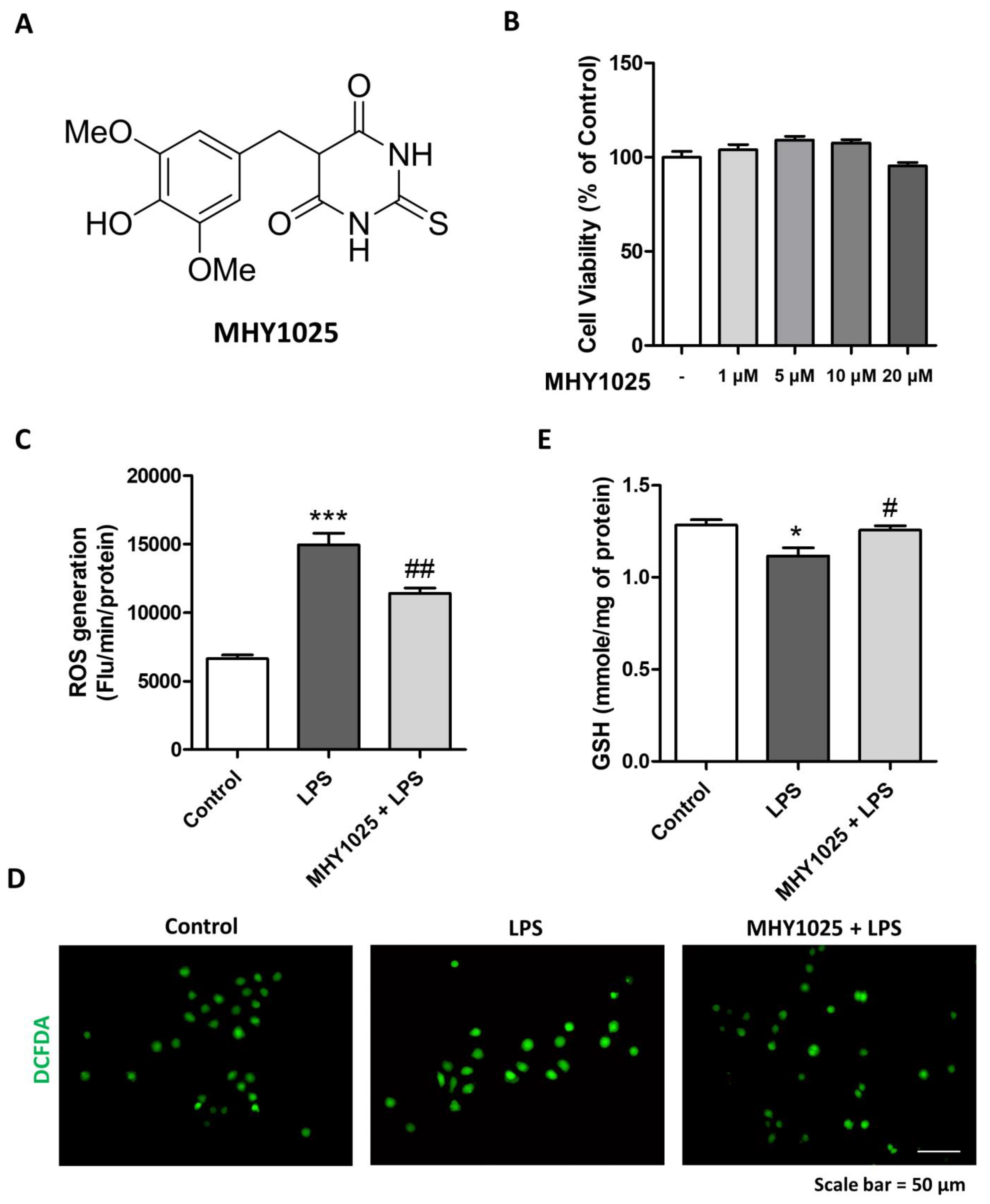
Cell viability was measured to determine the cytotoxicity of MHY1025 in NRK52E cells. The EZ-cytox Cell Viability Assay Kit (Dogen, Seoul, South Korea) was used according to the manufacturer’s instructions. Briefly, NRK52E cells were cultured to 70–80% confluence in serum-free media and treated with MHY1025 at the indicated concentrations for 24 h. The cells were further cultured for 30 min by adding the cell viability solution in the incubator, and the absorbance was measured at 450 nm using a spectrophotometer. The percentage of viable cells was calculated according to the following formula: [(chemically treated group/control group) × 100].
2.4. Determination of Antioxidant Effect
Cellular oxidative stress was quantified to determine the antioxidant effects of MHY1025. ROS generation was measured using a 2′,7′-dichlorodihydrofluorescein diacetate (DCFDA) fluorescent dye. Briefly, the cells were scarpered and centrifuged and the supernatants were removed. Cells were incubated with 50 μM DCFDA dissolved in PBS. Changes in fluorescence were detected using a GENios plate reader (Tecan Instruments, Salzburg, Austria) at excitation and emission wavelengths of 485 and 530 nm, respectively. Changes in the ROS levels were detected using a fluorescence microscope. The cellular glutathione (GSH) levels were quantified. Briefly, 25% of the meta-phosphoric acid-added cell pellets were centrifuged at 12,000 rpm for 10 min and the supernatant was collected for the assay. For GSH, 1 mM EDTA-50 mM phosphate buffer was added to the supernatant, followed by the addition of o-phthaladehyde. After 20 min at room temperature, the fluorescence was measured at excitation and emission wavelengths of 360 and 460 nm, respectively.
2.5. Determination of the Transcriptional Activity
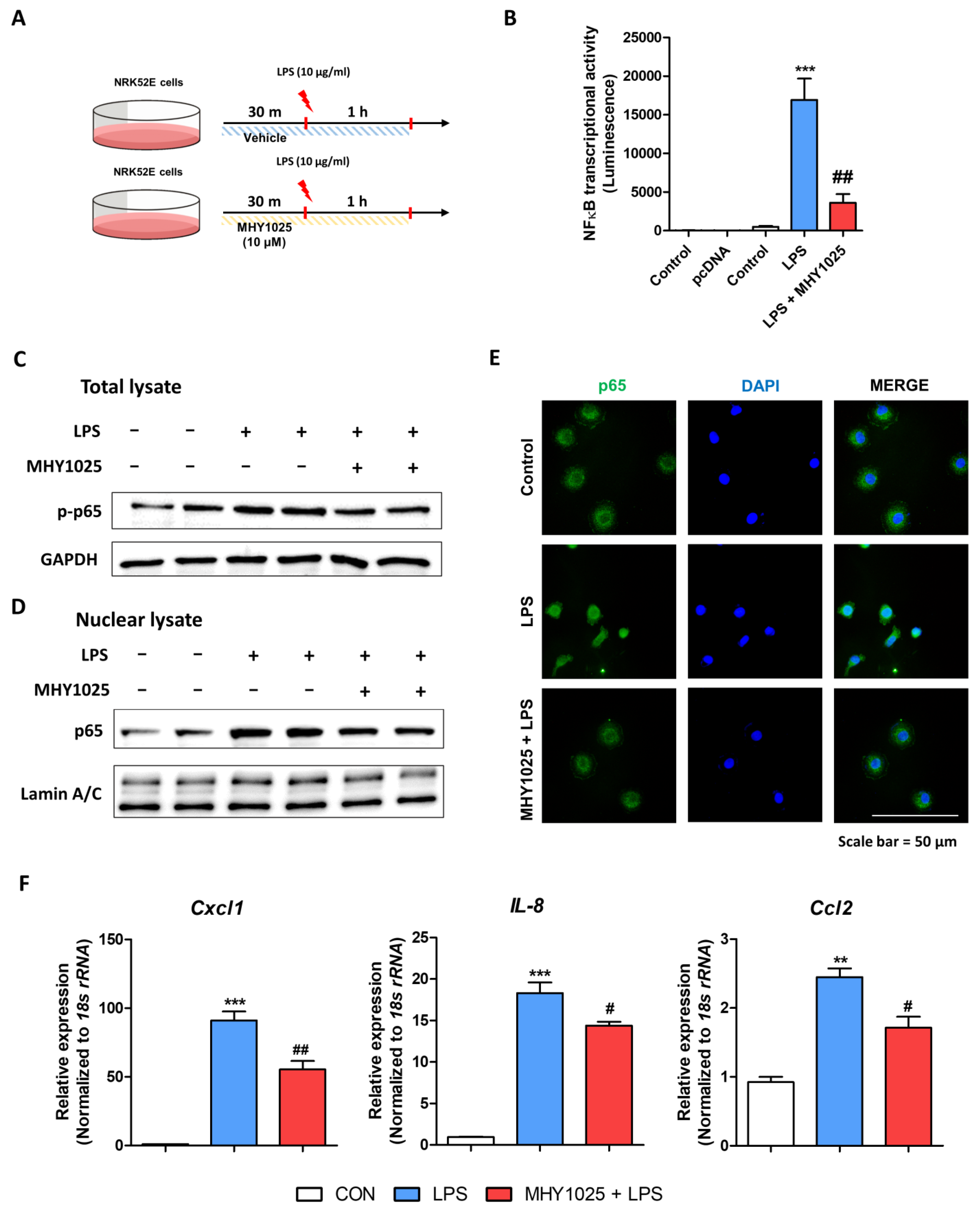
A luciferase assay was performed to detect the transcriptional activity of NF-κB in the NRK52E cells. Lipofectamine 3000 transfection reagent (Invitrogen, Carlsbad, CA, USA) and Opti-MEM (Gibco, Grand Island, NY, USA) were used according to the manufacturer’s instructions for the plasmid transfection. Cells were cultured to 70–80% confluence in serum-free media and transfected with 0.05 ng of NF-κB promoter-LUC plasmids. After 24 h of NF-κB transfection, the cells were treated with 10 μM MHY1025 for 2 h. Luciferase activity was measured using the One-Glo Luciferase Assay System (Promega, Madison, WI, USA) and detected using a luminescence plate reader (Berthold Technologies GmbH & Co., Bad Wildbad, Germany).
2.6. Immunofluorescence
Immunofluorescence staining (IF) was used to detect the location of proteins in the cells. Briefly, cells were fixed in 4% formaldehyde for 10 min, washed three times with ice-cold phosphate-buffered saline (PBS), and exposed to 0.25% Triton-X 100 in PBS for 10 min for permeabilization. After blocking with 1% BSA/0.1% Tween 20 in PBS at room temperature for 30 min, the cells were incubated overnight with primary antibody diluted in blocking buffer at 4 °C. The cells were washed with PBS, incubated with the secondary antibody for 1 h in the dark, and counterstained with Hoechst 33,258 in PBS for 1 min. The images were acquired using a fluorescence microscope (LS30; Leam Solution, Seoul, Korea).
2.7. Protein Extraction and Western Blot Analysis
ProEXTM CETi (Translab, Daejeon, Korea) containing a protease inhibitor cocktail and phosphate inhibitor was used to extract total protein lysates from the tissues. Total protein was extracted from cells using RIPA buffer (#9806, Cell Signaling Technology, Danvers, MA, USA) containing a protease inhibitor cocktail. Throughout the extraction process, all tubes, solutions, and centrifuges were maintained at a temperature of 4 °C. To determine protein concentrations, BCA reagent from Thermo Scientific (Waltham, MA, USA) was employed. Extracted proteins (10–20 μg) underwent a 5 min boiling step with 4× sample buffer (Cat#1610747, Bio-Rad, CA, USA) in a volume ratio of 3:1. Following protein extraction, sodium dodecyl sulfate-polyacrylamide gel electrophoresis was used to separate proteins from each sample on 8–15% acrylamide gels. Subsequently, a Bio-Rad Western blot system was used to transfer proteins to polyvinylidene difluoride (PVDF) membranes (Millipore, Burlington, MA, USA). The membranes were promptly placed in blocking buffer (5% non-fat milk) containing 10 mM Tris (pH 7.5), 100 mM NaCl, and 0.1% Tween 20. The membranes were subjected to a 30 min wash with Tris-buffered saline (TBS; 50 mM Tris and 150 mM NaCl [pH 7.6]) and Tween buffer (10 mM Tris [pH 7.5], 100 mM NaCl, and 0.1% Tween 20). The membranes were incubated with specific primary antibodies (diluted at 1:500 to 1:2000;
Table S1) at a temperature of 4 °C overnight. After the primary antibody incubation, the membranes were washed three times with TBS-Tween buffer for 10 min each, followed by incubation with a horseradish peroxidase-conjugated anti-mouse, anti-rabbit, or anti-goat secondary antibody (1:10,000 dilution) at 25 °C for 1 h. Immunoblots were visualized using Western Bright Peroxide solution from Advansta (San Jose, CA, USA) and a ChemiDoc imaging system from Bio-Rad, following the manufacturer’s instructions. Western blot analysis was performed on the kidneys of every animal tested in the experiments, and the figures display representative results.
2.8. RNA Extraction and qPCR
Total RNA was extracted using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s guidelines. Briefly, kidney tissues or cells were homogenized in the TRIzol reagent. Subsequently, 0.2 mL of chloroform per 1 mL of homogenate was added, and the samples were vigorously shaken for 15 min. The resulting aqueous phases were transferred to fresh tubes and an equal volume of isopropanol was added. Following a 15 min incubation at 4 °C, the samples were centrifuged at 12,000×
g for 15 min at 4 °C. The supernatants were carefully decanted and the RNA pellets were washed with 75% ethanol by vortexing. Afterward, centrifugation at 7500×
g for 8 min at 4 °C was performed. The resulting pellets were air-dried for 10–15 min and dissolved in diethyl pyrocarbonate-treated water. For reverse transcription, 1.0 μg of isolated RNA was used with a cDNA synthesis kit from GenDEPOT (Katy, TX, USA). Quantitative real-time polymerase chain reaction (qPCR) was performed using SYBR Green Master Mix (BIOLINE, Taunton, MA, USA) and the CFX Connect System (Bio-Rad). The primers were designed using Primer3Plus (
Table S2). To analyze qPCR data, the 2-ΔΔCT method was employed as a relative quantification approach.
2.9. Histological Analysis
To examine histological alterations in the kidneys, kidney specimens were fixed in a 10% neutral formalin solution. Paraffin-embedded sections were prepared and stained with hematoxylin and eosin (H&E) staining. To assess the extent of renal fibrosis and injury, Sirius Red (SR) staining was performed using a commercially available kit (VB-3017; Rockville, MD, USA). Immunohistochemical analysis was performed to evaluate protein expression in the kidneys. Paraffin-embedded sections were incubated with specific primary antibodies as indicated. Visualization was achieved using diaminobenzidine substrates and counterstaining was performed using hematoxylin. Images were captured using a microscope (LS30).
2.10. In Situ Hybridization
In situ hybridization (ISH) was performed on tissue samples that had been fixed in formalin and embedded in paraffin. For visualization of RNA expression within the tissue, either the RNAscope 2.5 HD Assay (322300, Biotechne, Minneapolis, MN, USA) or the RNAscope 2.5 HD Duplex Detection Kit (322436, bio-techne, Minneapolis, MN, USA) was used, following the manufacturer’s instructions. The RNAscope assay was conducted using the following probes: Mm-Ccl2 cat# 311791, Mm-Emr1 cat# 317969-C2, and Mm-Col1a1 cat# 319379. A microscope (LS30) was used to capture images.
2.11. Quantification and Statistical Analysis
Student’s t-test was used to analyze differences between two groups, and analysis of variance was used to analyze intergroup differences. Statistical significance was set at p < 0.05. Statistical analyses were performed using GraphPad Prism version 5 (GraphPad Software Inc., San Diego, CA, USA). Image calculations were performed using ImageJ software (Version 1.54d, National Institutes of Health, Bethesda, MD, USA).
4. Discussion
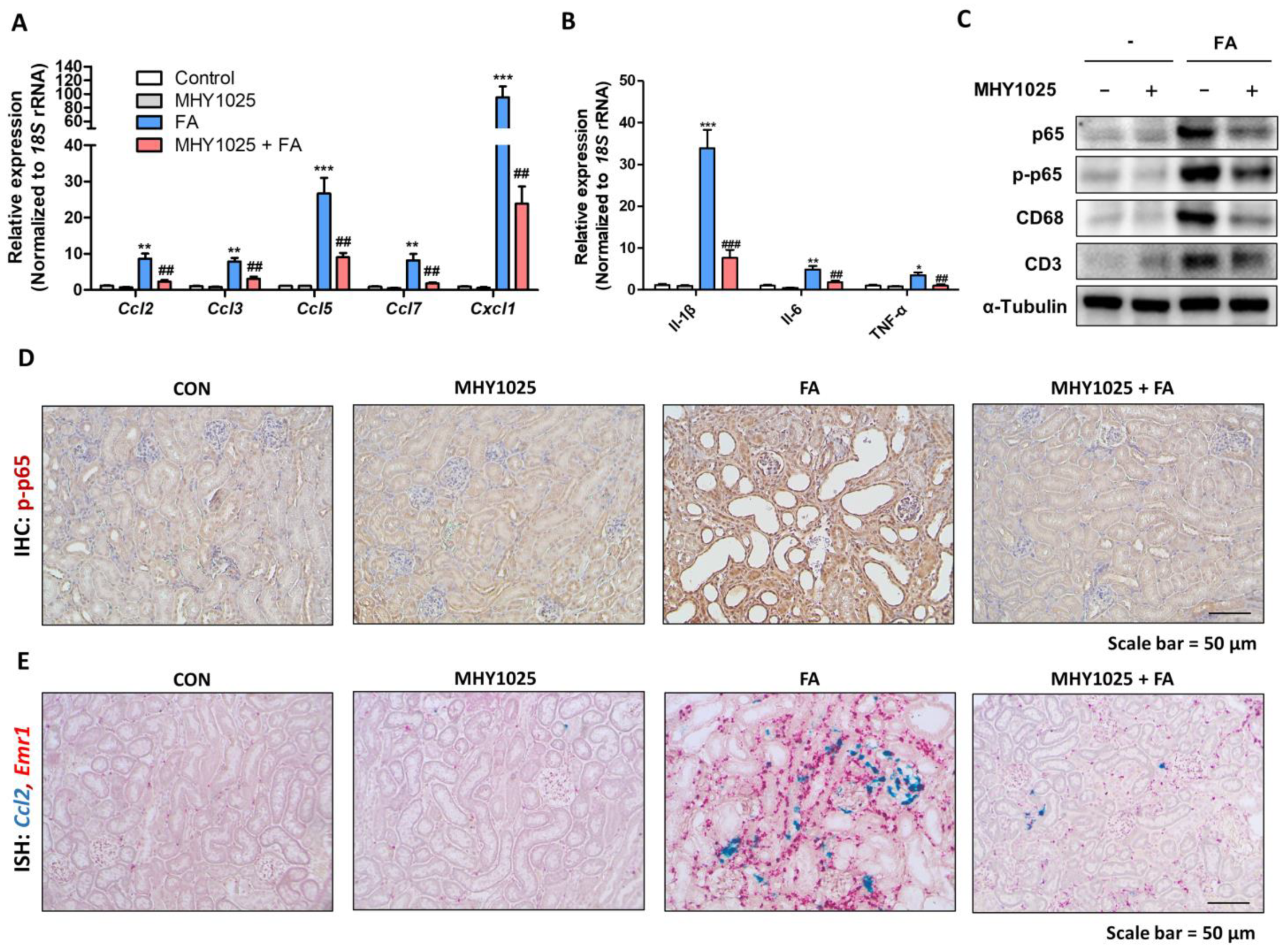
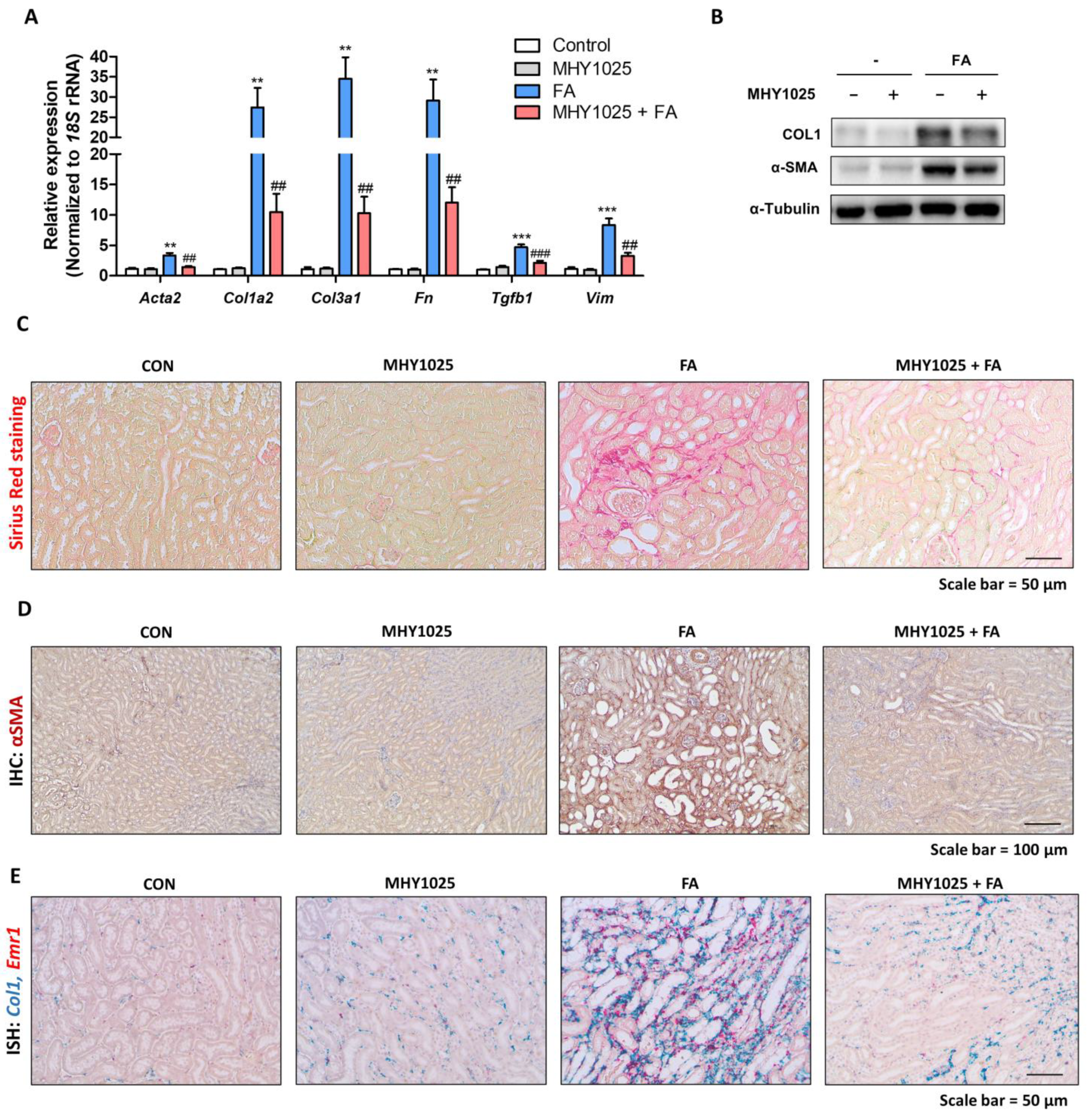
The results of our study demonstrated the multifaceted beneficial effects of MHY1025 in kidney disease. Our investigation focused on elucidating the protective mechanisms of MHY1025 against various aspects of kidney injury ranging from oxidative stress and inflammation to fibrotic progression. The antioxidative properties of MHY1025 reduced the inflammatory response in the kidney epithelial cells. Furthermore, MHY1025 directly suppressed TGFβ-induced fibroblast activation and reduced fibrosis markers. Based on the in vitro observations, we further demonstrated the in vivo efficacy of MHY1025 in a folic acid-induced kidney fibrosis model. MHY1025 significantly reduced kidney inflammation and fibrosis Collectively, these data highlight the potential therapeutic value of MHY1025 in mitigating renal damage and dysfunction.
Oxidative stress is a fundamental pathological process implicated in various chronic diseases, including kidney disorders [
19,
20]. The kidneys are highly metabolically active organs, and their physiological functions, such as filtration, reabsorption, and excretion, involve complex cellular processes that generate ROS as byproducts [
21]. Oxidative stress plays a pivotal role in the initiation and propagation of renal injury and contributes to inflammation, cellular damage, and fibrosis [
22]. Kidneys are highly susceptible to various sources of oxidative stress, including ischemia/reperfusion injury, inflammation, metabolic disorders, and hypertension [
23]. Oxidative stress triggers a cascade of detrimental effects on the kidney function and structure. ROS directly induce cellular damage by influencing cellular components such as lipids, proteins, and DNA, leading to cell dysfunction and death. ROS also contribute to the activation of pro-inflammatory pathways, promote fibroblast activation, and directly cause vascular endothelial dysfunction. Understanding the role of oxidative stress in kidney disease has prompted research on antioxidant therapies as potential interventions.
Inflammation, which is often associated with oxidative stress, is a hallmark of kidney injury [
24]. During the initial phase, inflammation is activated as a protective response to eliminate the cause of the injury and facilitate tissue repair. However, during prolonged inflammation, fibroblasts can become overactive, leading to the excessive production of collagen and other extracellular matrix components. During extended inflammation, leukocytes originating from the bone marrow, particularly neutrophils and macrophages, play a central role in kidney inflammation [
25]. Accumulation of these cells is a prominent hallmark of pro-inflammatory kidney disease. In addition, research has revealed a significant contribution of kidney cells activated within the kidney itself, including TECs, mesangial cells, podocytes, and endothelial cells. TECs play a pivotal role in initiating inflammatory responses during the progression of interstitial fibrosis [
26]. Under injured or damaged conditions, TECs actively engage in pro-inflammatory reactions by producing chemokines. Based on these observations, the regulation of epithelial inflammation is an interesting target for modulating kidney inflammation and fibrosis. MHY1025 reduced oxidative stress and LPS-induced inflammatory responses in the renal tubular epithelial cells. MHY1025 significantly reduced NF-κB activation, translocation, and chemokine production in the epithelial cells. Since ROS play a significant role in NF-κB activation, it is reasonable to assume that the antioxidant properties of MHY1025 may have contributed to the control of epithelial inflammation.
In addition to its role in inflammation, oxidative stress has also been implicated in the development of kidney fibrosis. TGF-β is recognized as a pivotal facilitator of renal fibrosis, as it activates fibroblasts within the kidney. Therefore, it is an intriguing candidate for fibrosis treatment. While it is widely accepted that the Smad pathway plays a central role in TGF-β-induced fibrogenesis, recent findings suggest that oxidative stress exerts regulatory influence over TGF-β signaling through various pathways, including the Smad pathway [
27]. Moreover, the interplay between TGF-β and oxidative stress creates a detrimental feedback loop. Consequently, pursuing interventions that address TGF-β-induced and ROS-driven cellular signaling emerges as an innovative strategy for tackling fibrotic conditions. We also found that MHY1025 exerts direct antifibrotic effects in TGF-β-induced fibroblast activation in NRK49F cells. MHY1025 effectively blocked TGF-β-induced collagen production and αSMA expression in fibroblasts.
Based on the in vitro observations, we further evaluated the MHY1025’s efficacy using an in vivo animal model of kidney fibrosis. The widely used folic acid-induced kidney fibrosis model is characterized by an excessive inflammatory response in epithelial cells. MHY1025 treatment significantly reduced renal damage, inflammation, and fibrosis. Nonetheless, our study was subject to several limitations. Our primary evaluation of the role of MHY1025 largely relied on cell lines, and it will be essential to utilize other in vitro models, including primary kidney cells, to validate its efficacy. Additionally, our investigation solely employed a single animal model of kidney fibrosis. To further substantiate the effectiveness of MHY1025, it is advisable to incorporate additional models such as the ischemia–reperfusion kidney injury model. Collectively, our results suggest that the administration of antioxidant compounds effectively prevented renal fibrosis in both in vitro and in vivo models of kidney fibrosis, indicating the therapeutic potential of antioxidants against CKD.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





