
Inflammation and Oxidative Stress Induced by Obesity, Gestational Diabetes, and Preeclampsia in Pregnancy: Role of High-Density Lipoproteins as Vectors for Bioactive Compounds

 , ,
, ,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Methods
3. Bioactive Compounds Contained in Foods Used during Pregnancy
4. Pregnancy–Oxidative Stress–Inflammation Association
5. OS and Inflammation during Pregnancy, Metabolic Changes in Chronic Diseases
5.1. Preeclampsia

{kind=link}
{kind=link}
{kind=link}
| Study Groups | Oxidative and Inflammatory Changes | Effects in Mothers/Neonates | Ref. |
|---|---|---|---|
| Follow-up from 1st to 3rd trimester | |||
| 1st–2nd trim = 22 3rd trim = 11 >35 GW = 9 | ↑ HIF1A mRNA since 1st trimester in PW who later developed PE ↑ MIF, ENG, FLT1, and BACE2 mRNA at week 24–30, and only ENG remains high at 31–34 weeks in PW who developed PE | ND | [81] |
| Group I (risk of HAT without PE) = 82 Group II (risk of HAT developing PE) = 43 | 1st trimester in Group II vs. Group I: ↑ IL-6, TNF-α, IFNγ, TBARS, TAC 2nd trimester in Group II vs. Group I: ↑ hsCRP, TNF-α, IFNγ, TBARS, TAC 3rd trimester in Group II vs. Group I and 3rd vs. 2nd trimester: ↑ IL-6, hsCRP, TNF-α, IFNγ, TBARS, TAC | All parameters correlated with baroreflex sensitivity except BMI and TAC. TBARS had a significant independent contribution to BRS. | [82] |
| Follow up from 1st trimester to 3rd trimester n = 441 | ↑ CRP, TNF-α, and 8-isoprostane early in pregnancy (median 10 weeks) CRP of PE is similar to non-PE at the end of pregnancy ↑ TNF-α in PE across pregnancy and ↑ 8-isoprostane in PE in early gestation. No changes in IL-1, IL-6, IL-10 and 8-OHdG | 50 PW developed PE The strongest associations were observed at approximately 18 weeks of pregnancy. TNF-α and 8-isoprostane were consistently elevated at all four time points in pregnancy. 8-OHdG was found as a protective factor against PE. | [83] |
| Follow up of 96 PW with high risk of PE at 16th week and next at 36th week | ↑ MDA, hsCRP, and IL-6 at 36th week compared to 16th week of pregnancy ↓ Baroreflex sensitivity and NO | Decreased BRS and increased interleukin-6 are associated with reduction in NO | [84] |
| Second trimester | |||
| PE = 49 Ctrl = 50 | ↑ MDA, Cardiac-specific troponin-I, and ↓ CAT, SOD, and GSH in PE vs. Ctrl | Association between SOD and MDA, and CAT with MDA. CAT correlated with hsCRP | [94] |
| Third and delivery | |||
| PE = 20 Ctrl = 18 | ↑ 8-isoprostane in plasma only in post-partum ↓ urinary excretion of 8-iso-prostane in PE | The variation of urinary 8-iso-prostane depends on creatinine clearance and plasmatic endothelin-1 | [97] |
| PE = 10 Ctrl = 11 | ↑ AOPP in PE vs. non-pregnant women but not with Ctrl ↑ CRP of PE vs. Ctrl | ND | [42] |
| PE = 19 Ctrl = 18 Non PW = 20 | ↑ Plasmatic and urinary 8-iso-prostane in PE and Ctrl vs. non-PW but not in PE vs. Ctrl ↓ α-tocopherol in PE and Ctrl vs. Non PW γ-tocopherol decreased in PE vs. Ctrl and vs. non-PW | ND | [98] |
| Mild PE = 47 Severe PE = 36 Ctrl = 50 | ↑ sFlt-1/PlGF, hs-CRP, 8-isoprostane and leptin and lower adiponectin in PE vs. Ctrl. | Significant positive correlations in sFlt-1/PlGF and hs-CRP or leptin. A weak inverse correlation emerged between sFlt-1/PlGF and adiponectin. | [92] |
| Non-pregnant controls = 10 PE = 28 Ctrl = 59 | ↑ Superoxide and ferritin in PE vs. control | During HP, superoxide concentrations correlate significantly with arterial stiffness, while in PE, superoxide is significantly correlated to microvascular endothelial function. | [101] |
| PE = 15 Ctrl = 28 | ↑ AGEs in PE vs. Ctrl | + Correlation between circulating levels of AGEs and gamma-glutamyl transpeptidase, uric acid, glucose, insulin, and HOMA-IR in PE | [102] |
| Non-PW = 20 PW = 20 Ctrl = 20 | ↑ Serum resistin, vaspin, MDA, and IL-8 in PE vs. non-PE and Ctrl | Only resistin and vaspin were reduced in PE after four weeks of postpartum | [85] |
| Mild PE = 45 Severe PE = 40 Ctrl = 48 | ↓ TAC and ↑ TBARS, IL-6, TNF-α and IFN-γ in mild and severe PE groups compared to Ctrl | + Correlation of circulating beta-human chorionic gonadotropin and OS and inflammatory markers | [86] |
| PE = 27 Ctrl = 43 | MPO levels in placenta higher than the levels in normal control subjects | ND | [103] |
| PE = 5 Ctrl = 5 | In preeclamptic women, intense xanthine oxidase immunoreactivity was present within the epidermis. | ND | [104] |
| PE = 67 Ctrl = 70 | ↑ CRP, fetal DNA, MDA and Hsp70 level | In PE serum, Hsp70 levels showed significant correlations with serum CRP, aspartate aminotransferase, and LDH activities MDA | [86] |
| PE = 53 Ctrl = 20 Mild PE = 32 Severe PE = 21 | ↑ hsCRP, IL-6, TNF-α and 8-isoprostane in PE vs. Ctrl and in severe PE vs. mild PE No changes in MDA | Plasma levels of 8-isoprostane were significantly correlated with the plasma levels of hs-CRP, IL-6, and TNF-α in patients with PE | [89] |
| PE = 71 Ctrl = 71 | In amniotic fluid: ↑ sFLT1, sEndoglin, endothelin 1, and leptin in PE. sFLT1, sEndoglin, leptin, and adiponectin ↑ in PE IUGR than those without IUGR. | Leptin has the largest area under the curve (0.753). Amniotic proteins are involved in the inflammatory process of the human placenta. | [93] |
| PE = 44 Ctrl = 44 | ↑ serum CRP, plasma MDAin PE vs. Ctrl | Plasma OPN concentrations are increased in preeclamptic patients with extensive endothelial injury. | [95] |
| PE = 60 Ctrl = 60 | ↑ Hsp70, IL-1Ra, IL2, IL4, IL6, IL8, IL, IL10, IL12p40, IL12p70, IL18, INF&, IP10, MCP-1, ICAM-1, VCAM-1, sFlt-1, P1GF in PE vs. Ctrl | Elevated serum Hsp70 level and sFlt-1/PlGF ratio had a synergistic (joint) effect in the risk of preeclampsia | [86] |
| PE = 46 Ctrl = 42 | ↑ TBARS, TBARS/TAS, IL-6, TNF-a, a1-antitrypsin, CRP in plasma of PE vs. Ctrl ↑ CRP and a1-Antitrypsin in umbilical cord of PE vs. Ctrl ↑ sVCAM, sL-selectin in leucocytes of PE vs. Ctrl | + Correlation between maternal and UCB TAS, IL-6, CRP in Ctrl and PE | [87] |
| PE = 33 Ctrl = 33 | ↑ IL6, MDA, CP, in plasma and placenta of PE vs. Ctrl ↑ TLR-4 and NF-kB in the placenta | ND | [88] |
| PE = 50 (17 mild and 33 severe) Ctrl = 33 | ↑ MDA in PE mild vs. PE severe ↑ IL-6, IL-10, TNF-a in PE vs. Ctrl ↑ IL-6/IL-10 ratio in PE vs. Ctrl | ND | [91] |
| PE = 18 Ctr = 18 | ↑ CAT, Il-6, TNF-α in PE vs. Ctrl Plasma Mg was positively correlated with CAT and GPx activities with concentrations of IL-6 and TNF-α | Plasma magnesium and urinary 8-isoprostane were associated with PE | [99] |
| PE = 100 Ctr = 50 | ↑ SOD, GSH/GSSG ratio, IL-6, and ↓ GSSG in placenta of PE vs. Ctrl | + Associations between placental GSH levels with weight, head, and chest circumference and gestational age at birth | [100] |
5.2. Obesity
5.3. Gestational Diabetes Mellitus
| Study Groups | Oxidative and Inflammatory Markers | Oxidative Stress Changes | Inflammation Changes | Effects in Mothers and Newborn | Ref. |
|---|---|---|---|---|---|
| GDM = 22 Ctrl = 22 | Proteome profiling in plasma | ↓ PON/LAC3 in GDM vs. Ctrl | ↑ CRP and ↓ IGFBP2 in GDM vs. Ctrl | Strong correlations among the inflammation-related proteins and proteins of blood coagulation, lipid homeostasis membrane, and antioxidative enzymes | [128] |
| GDM = 11 Ctrl = 23 | Six F2-isoprostanes isomers | ↑ 8-iso-15(R)-PGF2α levels in GDM vs. Ctrl | ND | Delta-6-desaturase (D6D) activity index, calculated using fatty acid ratios, was 9% lower in pre-existing diabetes than in controls | [146] |
| GDM = 48 Ctrl = 46 | Total oxidative stress (TOS), total antioxidant status (TAS), and oxidative stress index (OSI) in serum | ↑ Serum OGTT, OSI, TOS in GDM vs. Ctrl | ND | ND | [134] |
| GDM = 60 Ctrl = 75 | CRP, NLR and PLR, PCT, and VAP-1 | ND | ↑ CRP, NLR, PLR, PCT and VAP-1 in GDM vs. Ctrl | + Correlation of VAP-1 with glucose, HbA1c, PLR, and CRP | [136] |
| GDM = 114 Ctrl = 114 | ROS (DCFH-DA) and DNA damage (comet assay) in lymphocytesNLR and MVP in blood | ↑ ROS and DNA damage in GDM vs. Ctrl | ↑ NLR and mean platelet volume (MPV) in GDM vs. Ctrl | The elevated parameters are independent determinants of GDM after adjustment for BMI | [135] |
| GDM = 30 | CRP, ferritin, TNF-α, methylglyoxal, glycated albumin | ND | ↑ PAF and TNF-α after diet ↑ MGO in GDM vs. non-pregnant healthy people | + Correlation of MGO levels with HbA1c, pregnancy weight and HOMA-IR at GDM diagnosis and after 12 weeks | [147] |
| GDM = 40 Ctrl = 40 | GDF-15 in serum | ND | ↑ Serum GDF-15 levels in patients with GDM vs. Ctrl | + Correlation of GDF-15 and BMI for both GDM and Ctrl ↓ Apgar scores at 1 min and 5 min in the GDM vs. Ctrl | [141] |
| GDM = 176 Ctrl = 164 | MDA, hsCRP, IL-6, and NO in plasma | ↑ MDA and ↓ NO in GDM vs. Ctrl | ↑ hsCRP and IL-6 in GDM vs. Ctrl | Significant association of HbA1C, MDA, and interleukin-6 with maternal stress and postnatal depression | [140] |
| GDM = 17 Ctrl = 23 | CAT, GPX, GSR, and SOD gene expression in placenta and omental and subcutaneous adipose tissue | ↑ CAT and GSR in placenta of GDM vs. Ctrl No changes in GPx or SOD in placenta and no changes in adipose tissue | Treatment with hypoxanthine/xanthine oxidase-stimulated cytokine release (IL1B, MIP1b, and TNF-α) and TNF-α mRNA expression | ND | [137] |
| GDM = 8 T2D = 3 T1D = 12 Ctrl = 13 | IL-8, MCP-1, and CP in plasma and umbilical cord | No changes in CP in mothers with DM vs. Ctrl | No changes in MCP-1 and IL-8 in mothers with DM vs. Ctrl ↑ MCP-1 and CP in umbilical cords from mothers with DM | MCP-1 correlated with ketone bodies and acetoacetate | [138] |
| GDM = 69 PWOb = 44 PWOw = 48 Ctrl = 104 | TBARS in placenta and UCP2, SIRT1, PPARα, TLR4, and GR-a gene expression in placenta tissue | ↑ UCP2 expression in PWOw and PWOb vs. Ctrl | ↑ SIRT1 expression in PWOw and PWOb vs. Ctrl ↑ GR-a expression in GDM vs. Ctrl | ↑ Monocyte count was higher in the cord blood PWOb with normal glucose tolerance ↑ Serum leptin PWOb compared to Ctrl and their offspring | [139] |
5.4. Risk of Preterm Delivery
5.5. Non-Alcoholic Fatty Liver Disease (NAFLD)
6. Lipids and Lipoproteins in Pregnancy
6.1. Lipoproteins
6.2. HDL
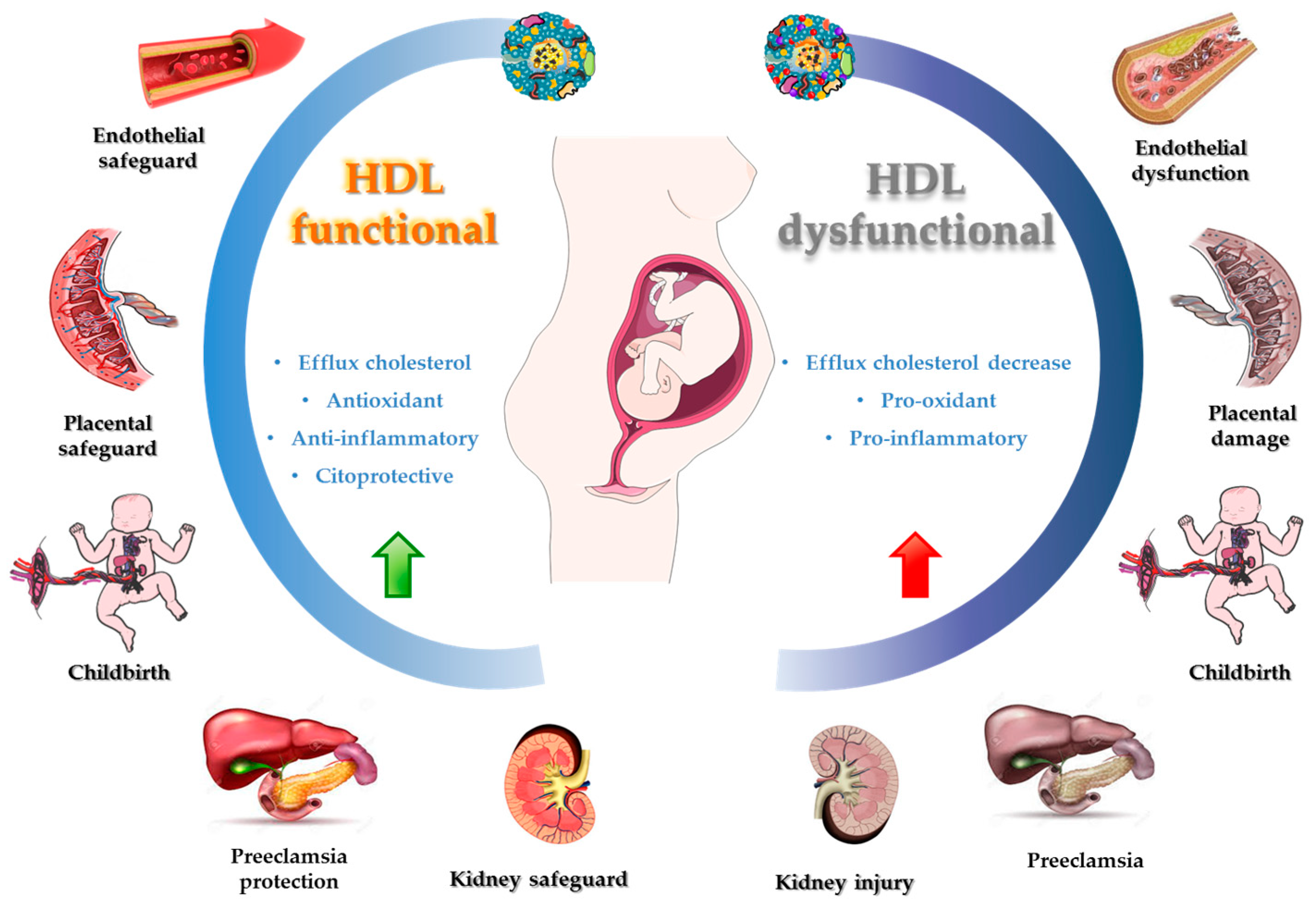
6.3. HDL Role and Upregulation Contribution in Inflammatory Processes and OS
6.4. Bioactive Compounds and HDL Functionality
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kazma, J.M.; van den Anker, J.; Allegaert, K.; Dallmann, A.; Ahmadzia, H.K. Anatomical and Physiological Alterations of Pregnancy. J. Pharmacokinet. Pharmacodyn. 2020, 47, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Kesikburun, S.; Güzelküçük, Ü.; Fidan, U.; Demir, Y.; Ergün, A.; Tan, A.K. Musculoskeletal Pain and Symptoms in Pregnancy: A Descriptive Study. Ther. Adv. Musculoskelet. Dis. 2018, 10, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.K.; Harsha, M.M.; Rathoor, S. Cardiovascular Diseases in Pregnancy—A Brief Overview. Curr. Cardiol. Rev. 2021, 18, 77–88. [Google Scholar] [CrossRef]
- O’Kelly, A.C.; Michos, E.D.; Shufelt, C.L.; Vermunt, J.V.; Minissian, M.B.; Quesada, O.; Smith, G.N.; Rich-Edwards, J.W.; Garovic, V.D.; El Khoudary, S.R.; et al. Pregnancy and Reproductive Risk Factors for Cardiovascular Disease in Women. Circ. Res. 2022, 130, 652–672. [Google Scholar] [CrossRef]
- Estrada-Luna, D.; Ortiz-Rodriguez, M.A.; Medina-Briseño, L.; Carreón-Torres, E.; Izquierdo-Vega, J.A.; Sharma, A.; Cancino-Díaz, J.C.; Pérez-Méndez, O.; Belefant-Miller, H.; Betanzos-Cabrera, G. Current Therapies Focused on High-Density Lipoproteins Associated with Cardiovascular Disease. Molecules 2018, 23, 2730. [Google Scholar] [CrossRef]
- Stadler, J.T.; van Poppel, M.N.M.; Wadsack, C.; Holzer, M.; Pammer, A.; Simmons, D.; Hill, D.; Desoye, G.; Marsche, G. Obesity Affects Maternal and Neonatal HDL Metabolism and Function. Antioxidants 2023, 12, 199. [Google Scholar] [CrossRef] [PubMed]
- Monserrat-Mesquida, M.; Quetglas-Llabrés, M.; Bouzas, C.; Capó, X.; Mateos, D.; Ugarriza, L.; Tur, J.A.; Sureda, A. Peripheral Blood Mononuclear Cells Oxidative Stress and Plasma Inflammatory Biomarkers in Adults with Normal Weight, Overweight and Obesity. Antioxidants 2021, 10, 813. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular Mechanisms Linking Oxidative Stress and Diabetes Mellitus. Oxid. Med. Cell. Longev. 2020, 2020, 8609213. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Mechanistic Insight into Oxidative Stress-Triggered Signaling Pathways and Type 2 Diabetes. Molecules 2022, 27, 950. [Google Scholar] [CrossRef]
- Afrose, D.; Chen, H.; Ranashinghe, A.; Liu, C.C.; Henessy, A.; Hansbro, P.M.; McClements, L. The Diagnostic Potential of Oxidative Stress Biomarkers for Preeclampsia: Systematic Review and Meta-Analysis. Biol. Sex Differ. 2022, 13, 26. [Google Scholar] [CrossRef]
- Sorrell, T.R. Mexican Traditional Medicine: Application of a Traditional and Complementary Medicine System to Improve Opioid Use Treatment in Latinos. J. Holist. Nurs. 2020, 38, 384–399. [Google Scholar] [CrossRef] [PubMed]
- Lucía, C.P.A.; Jacqueline, B.R.; Alberto, B.R.L.; David, B.A.; Beatriz, R.A. Actualized Inventory of Medicinal Plants Used in Traditional Medicine in Oaxaca, Mexico. J. Ethnobiol. Ethnomed. 2021, 17, 7. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.E.; Cooper, K.L.; Relton, C.; Thomas, K.J. Prevalence of Complementary and Alternative Medicine (CAM) Use by the General Population: A Systematic Review and Update. Int. J. Clin. Pract. 2012, 66, 924–939. [Google Scholar] [CrossRef]
- Shewamene, Z.; Dune, T.; Smith, C.A. The Use of Traditional Medicine in Maternity Care among African Women in Africa and the Diaspora: A Systematic Review. BMC Complement. Altern. Med. 2017, 17, 382. [Google Scholar] [CrossRef] [PubMed]
- Okoth, K.; Chandan, J.S.; Marshall, T.; Thangaratinam, S.; Thomas, G.N.; Nirantharakumar, K.; Adderley, N.J. Association between the Reproductive Health of Young Women and Cardiovascular Disease in Later Life: Umbrella Review. BMJ 2020, 371, m3502. [Google Scholar] [CrossRef] [PubMed]
- Cabada-Aguirre, P.; López López, A.M.; Mendoza, K.C.O.; Garay Buenrostro, K.D.; Luna-Vital, D.A.; Mahady, G.B. Mexican Traditional Medicines for Women’s Reproductive Health. Sci. Rep. 2023, 13, 2807. [Google Scholar] [CrossRef]
- Yılmaz, İ.; Enver Dolar, M.; Özpınar, H. Effect of Administering Kefir on the Changes in Fecal Microbiota and Symptoms of Inflammatory Bowel Disease: A Randomized Controlled Trial. Turkish J. Gastroenterol. 2019, 30, 242–253. [Google Scholar] [CrossRef]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsakris, Z.; Rozos, G.; Tsigalou, C.; Bezirtzoglou, E. Interactions between Medical Plant-Derived Bioactive Compounds: Focus on Antimicrobial Combination Effects. Antibiotics 2022, 11, 1014. [Google Scholar] [CrossRef]
- Liang, B.; Zhu, Y.C.; Lu, J.; Gu, N. Effects of Traditional Chinese Medication-Based Bioactive Compounds on Cellular and Molecular Mechanisms of Oxidative Stress. Oxid. Med. Cell. Longev. 2021, 2021, 3617498. [Google Scholar] [CrossRef]
- Chernukha, I.; Kotenkova, E.; Derbeneva, S.; Khvostov, D. Bioactive Compounds of Porcine Hearts and Aortas May Improve Cardiovascular Disorders in Humans. Int. J. Environ. Res. Public Health 2021, 18, 7330. [Google Scholar] [CrossRef]
- Li, H.Y.; Gan, R.Y.; Shang, A.; Mao, Q.Q.; Sun, Q.C.; Wu, D.T.; Geng, F.; He, X.Q.; Li, H. Bin Plant-Based Foods and Their Bioactive Compounds on Fatty Liver Disease: Effects, Mechanisms, and Clinical Application. Oxid. Med. Cell. Longev. 2021, 2021, 6621644. [Google Scholar] [CrossRef] [PubMed]
- Bocanegra, A.; Macho-González, A.; Alba Garcimartín, J.B.; Sánchez-Muniz, F.J. Whole Alga, Algal Extracts, and Compounds as Ingredients of Functional Foods: Composition and Action Mechanism Relationships in the Prevention and Treatment of Type-2 Diabetes Mellitus. Int. J. Mol. Sci. 2021, 22, 3816. [Google Scholar] [CrossRef] [PubMed]
- Mohd Sairazi, N.S.; Sirajudeen, K.N.S. Natural Products and Their Bioactive Compounds: Neuroprotective Potentials against Neurodegenerative Diseases. Evid.-Based Complement. Altern. Med. 2020, 2020, 6565396. [Google Scholar] [CrossRef] [PubMed]
- Ghosh; Rajarshi; Bryant; Daniel, L.; Farone, A.L. Polysaccharides as Immunomodulators : Current Research Status and Future Directions. Molecules 2020, 25, 5854. [Google Scholar] [CrossRef] [PubMed]
- Ge, P.Y.; Qi, Y.Y.; Qu, S.Y.; Zhao, X.; Ni, S.J.; Yao, Z.Y.; Guo, R.; Yang, N.Y.; Zhang, Q.C.; Zhu, H.X. Potential Mechanism of s. Baicalensis on Lipid Metabolism Explored via Network Pharmacology and Untargeted Lipidomics. Drug Des. Devel. Ther. 2021, 15, 1915–1930. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.W.; Crew, J.; Planinic, P.; Alexander, J.M.; Basu, A. Associations of Dietary Bioactive Compounds with Maternal Adiposity and Inflammation in Gestational Diabetes: An Update on Observational and Clinical Studies. Int. J. Environ. Res. Public Health 2020, 17, 7528. [Google Scholar] [CrossRef]
- Ajanaku, C.O.; Ademosun, O.T.; Atohengbe, P.O.; Ajayi, S.O.; Obafemi, Y.D.; Owolabi, O.A.; Akinduti, P.A.; Ajanaku, K.O. Functional Bioactive Compounds in Ginger, Turmeric, and Garlic. Front. Nutr. 2022, 9, 1012023. [Google Scholar] [CrossRef]
- Nacka-Aleksić, M.; Pirković, A.; Vilotić, A.; Bojić-Trbojević, Ž.; Jovanović Krivokuća, M.; Giampieri, F.; Battino, M.; Dekanski, D. The Role of Dietary Polyphenols in Pregnancy and Pregnancy-Related Disorders. Nutrients 2022, 14, 5246. [Google Scholar] [CrossRef]
- Fogacci, S.; Fogacci, F.; Cicero, A.F.G. Nutraceuticals and Hypertensive Disorders in Pregnancy: The Available Clinical Evidence. Nutrients 2020, 12, 378. [Google Scholar] [CrossRef]
- Zhang, H.; Su, S.; Yu, X.; Li, Y. Dietary Epigallocatechin 3-Gallate Supplement Improves Maternal and Neonatal Treatment Outcome of Gestational Diabetes Mellitus: A Double-Blind Randomised Controlled Trial. J. Hum. Nutr. Diet. 2017, 30, 753–758. [Google Scholar] [CrossRef]
- Yuan, L.J.; Qin, Y.; Wang, L.; Zeng, Y.; Chang, H.; Wang, J.; Wang, B.; Wan, J.; Chen, S.H.; Zhang, Q.Y.; et al. Capsaicin-Containing Chili Improved Postprandial Hyperglycemia, Hyperinsulinemia, and Fasting Lipid Disorders in Women with Gestational Diabetes Mellitus and Lowered the Incidence of Large-for-Gestational-Age Newborns. Clin. Nutr. 2016, 35, 388–393. [Google Scholar] [CrossRef]
- Jaworsky, K.; DeVillez, P.; Basu, A. The Role of Phytochemicals and Plant-Based Diets in Gestational Diabetes: Evidence from Clinical Trials. Int. J. Environ. Res. Public Health 2023, 20, 4188. [Google Scholar] [CrossRef]
- Majed, B.H.; Khalil, R.A. Molecular Mechanisms Regulating the Vascular Prostacyclin Pathways and Their Adaptation during Pregnancy and in the Newborn. Pharmacol. Rev. 2012, 64, 540–582. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Roca, B.; Rubió-Piqué, L.; Montull-López, A. Polyphenol Intake in Pregnant Women on Gestational Diabetes Risk and Neurodevelopmental Disorders in Offspring: A Systematic Review. Nutrients 2022, 14, 3753. [Google Scholar] [CrossRef]
- Parham, S.; Kharazi, A.Z.; Bakhsheshi-Rad, H.R.; Nur, H.; Ismail, A.F.; Sharif, S.; Ramakrishna, S.; Berto, F. Antioxidant, Antimicrobial and Antiviral Properties of Herbal Materials. Antioxidants 2020, 9, 1309. [Google Scholar] [CrossRef] [PubMed]
- Ozkur, M.; Benlier, N.; Takan, I.; Vasileiou, C.; Georgakilas, A.G.; Pavlopoulou, A.; Cetin, Z.; Saygili, E.I. Ginger for Healthy Ageing: A Systematic Review on Current Evidence of Its Antioxidant, Anti-Inflammatory, and Anticancer Properties. Oxid. Med. Cell. Longev. 2022, 2022, 4748447. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.Y.; Ryu, J.H.; Shin, J.H.; Kang, M.J.; Kang, J.R.; Han, J.; Kang, D. Comparison of Anti-Oxidant and Anti-Inflammatory Effects between Fresh and Aged Black Garlic Extracts. Molecules 2016, 21, 430. [Google Scholar] [CrossRef]
- Sah, A.; Naseef, P.P.; Kuruniyan, M.S.; Jain, G.K.; Zakir, F.; Aggarwal, G. A Comprehensive Study of Therapeutic Applications of Chamomile. Pharmaceuticals 2022, 15, 1284. [Google Scholar] [CrossRef]
- Tafrihi, M.; Imran, M.; Tufail, T.; Gondal, T.A.; Caruso, G.; Sharma, S.; Sharma, R.; Atanassova, M.; Atanassov, L.; Fokou, P.V.T.; et al. The Wonderful Activities of the Genus Mentha: Not Only Antioxidant Properties. Molecules 2021, 26, 1118. [Google Scholar] [CrossRef]
- Sánchez, M.; González-Burgos, E.; Iglesias, I.; Lozano, R.; Gómez-Serranillos, M.P. The Pharmacological Activity of Camellia sinensis (L.) Kuntze on Metabolic and Endocrine Disorders: A Systematic Review. Biomolecules 2020, 10, 603. [Google Scholar] [CrossRef]
- Arikan, S.; Konukog, D.; Tülay, A.; Davas, I. Lipid Peroxidation and Antioxidant Status in Maternal and Cord Blood. Gynecol. Obs. Investig. 2001, 51, 145–149. [Google Scholar] [CrossRef]
- Fialová, L.; Malbohan, I.; Kalousová, M.; Soukupová, J.; Krofta, L.; Štípek, S.; Zima, T. Oxidative Stress and Inflammation in Pregnancy. Scand. J. Clin. Lab. Investig. 2006, 66, 121–128. [Google Scholar] [CrossRef]
- Guerby, P.; Tasta, O.; Swiader, A.; Pont, F.; Bujold, E.; Parant, O.; Vayssiere, C.; Salvayre, R.; Negre-Salvayre, A. Role of Oxidative Stress in the Dysfunction of the Placental Endothelial Nitric Oxide Synthase in Preeclampsia. Redox Biol. 2021, 40, 101861. [Google Scholar] [CrossRef] [PubMed]
- Sultana, Z.; Maiti, K.; Aitken, J.; Morris, J.; Dedman, L.; Smith, R. Oxidative Stress, Placental Ageing-Related Pathologies and Adverse Pregnancy Outcomes. Am. J. Reprod. Immunol. 2017, 77, e12653. [Google Scholar] [CrossRef]
- Myatt, L. Review: Reactive Oxygen and Nitrogen Species and Functional Adaptation of the Placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Murtaza, G.; Metwally, E.; Kalhoro, D.H.; Kalhoro, M.S.; Rahu, B.A.; Sahito, R.G.A.; Yin, Y.; Yang, H.; Chughtai, M.I.; et al. The Role of Oxidative Stress and Antioxidant Balance in Pregnancy. Mediat. Inflamm. 2021, 2021, 9962860. [Google Scholar] [CrossRef]
- Aouache, R.; Biquard, L.; Vaiman, D.; Miralles, F. Oxidative Stress in Preeclampsia and Placental Diseases. Int. J. Mol. Sci. 2018, 19, 1496. [Google Scholar] [CrossRef]
- Parrettini, S.; Caroli, A.; Torlone, E. Nutrition and Metabolic Adaptations in Physiological and Complicated Pregnancy: Focus on Obesity and Gestational Diabetes. Front. Endocrinol. 2020, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. A Novel and Compact Review on the Role of Oxidative Stress in Female Reproduction. Reprod. Biol. Endocrinol. 2018, 16, 80. [Google Scholar] [CrossRef]
- Mor, G.; Aldo, P.; Alvero, A.B. The Unique Immunological and Microbial Aspects of Pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482. [Google Scholar] [CrossRef]
- Denizli, M.; Capitano, M.L.; Kua, K.L. Maternal Obesity and the Impact of Associated Early-Life Inflammation on Long-Term Health of Offspring. Front. Cell. Infect. Microbiol. 2022, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Puttabyatappa, M.; Banker, M.; Zeng, L.; Goodrich, J.M.; Domino, S.E.; Dolinoy, D.C.; Meeker, J.D.; Pennathur, S.; Song, P.X.K.; Padmanabhan, V. Maternal Exposure to Environmental Disruptors and Sexually Dimorphic Changes in Maternal and Neonatal Oxidative Stress. J. Clin. Endocrinol. Metab. 2020, 105, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Gümüş, P.; Emingil, G.; Öztürk, V.Ö.; Belibasakis, G.N.; Bostanci, N. Oxidative Stress Markers in Saliva and Periodontal Disease Status: Modulation during Pregnancy and Postpartum. BMC Infect. Dis. 2015, 15, 261. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla-García, C.V.; Uribarri, J.; Fajardo-Araujo, M.E.; Barrientos-Romero, J.J.; Romero-Gutiérrez, G.; Reynaga-Ornelas, M.G.; Garay-Sevilla, M.E. Changes in Circulating Levels of Carboxymethyllysine, Soluble Receptor for Advanced Glycation End Products (SRAGE), and Inflammation Markers in Women during Normal Pregnancy. J. Matern. Neonatal Med. 2019, 32, 4102–4107. [Google Scholar] [CrossRef]
- Lean, S.C.; Jones, R.L.; Roberts, S.A.; Heazell, A.E.P. A Prospective Cohort Study Providing Insights for Markers of Adverse Pregnancy Outcome in Older Mothers. BMC Pregnancy Childbirth 2021, 21, 706. [Google Scholar] [CrossRef]
- Cave, C.; Hanson, C.; Schumacher, M.; Lyden, E.; Furtado, J.; Obaro, S.; Delair, S.; Kocmich, N.; Rezac, A.; Izevbigie, N.I.; et al. A Comparison of Vitamin E Status and Associated Pregnancy Outcomes in Maternal–Infant Dyads between a Nigerian and a United States Population. Nutrients 2018, 10, 1300. [Google Scholar] [CrossRef]
- Merlo, A.C.; Rosa, G.M.; Porto, I. Pregnancy-Related Acute Myocardial Infarction: A Review of the Recent Literature. Clin. Res. Cardiol. 2022, 111, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Hu, W.; Xia, B.; Liu, L.; Han, X.; Liu, Q. Association between Gestational Diabetes Mellitus and the Risks of Type-Specific Cardiovascular Diseases. Front. Public Health 2022, 10, 1–10. [Google Scholar] [CrossRef]
- Grønberg, M.; Flatmark, T. Inhibition of the H+-ATPase in Bovine Adrenal Chromaffin Granule Ghosts by Diethylstilbestrol Evidence for a Tight Coupling between ATP Hydrolysis and Proton Translocation. FEBS Lett. 1988, 229, 40–44. [Google Scholar] [CrossRef]
- Zvolsky, P.; Dvorakova, M.; Grof, P. Genealogic Study of Patients with Admission Diagnosis of Functional Psychosis. Arch. Psychiatr. Nervenkr. 1982, 232, 157–166. [Google Scholar] [CrossRef]
- Kalagiri, R.R.; Carder, T.; Choudhury, S.; Vora, N.; Ballard, A.R.; Govande, V.; Drever, N.; Beeram, M.R.; Uddin, M.N. Inflammation in Complicated Pregnancy and Its Outcome. Am. J. Perinatol. 2016, 33, 1337–1356. [Google Scholar] [CrossRef] [PubMed]
- Catalano, P.M.; Shankar, K. Obesity and Pregnancy: Mechanisms of Short Term and Long Term Adverse Consequences for Mother and Child. BMJ 2017, 356, j1. [Google Scholar] [CrossRef]
- Pantham, P.; Aye, I.L.M.H.; Powella, T.L. Inflammation in Maternal Obesity and Gestational Diabetes Mellitus. Placenta 2015, 176, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Humberg, A.; Fortmann, I.; Siller, B.; Kopp, M.V.; Herting, E.; Göpel, W.; Härtel, C.; Neonatal, G. Preterm Birth and Sustained Inflammation: Consequences for the Neonate Network, German Center for Lung Research and Priming Immunity at the Beginning of Life (PRIMAL) Consortium. Semin. Immunopathol. 2020, 42, 451–468. [Google Scholar] [CrossRef] [PubMed]
- Musa, E.; Salazar-Petres, E.; Arowolo, A.; Levitt, N.; Matjila, M.; Sferruzzi-Perri, A.N. Obesity and Gestational Diabetes Independently and Collectively Induce Specific Effects on Placental Structure, Inflammation and Endocrine Function in a Cohort of South African Women. J. Physiol. 2023, 7, 1287–1306. [Google Scholar] [CrossRef] [PubMed]
- Green, E.S.; Arck, P.C. Pathogenesis of Preterm Birth: Bidirectional Inflammation in Mother and Fetus. Semin. Immunopathol. 2020, 42, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Alston, M.C.; Redman, L.M.; Sones, J.L. An Overview of Obesity, Cholesterol, and Systemic Inflammation in Preeclampsia. Nutrients 2022, 14, 2087. [Google Scholar] [CrossRef]
- Schneider, D.; Hernández, C.; Farías, M.; Uauy, R.; Krause, B.J.; Casanello, P. Oxidative Stress as Common Trait of Endothelial Dysfunction in Chorionic Arteries from Fetuses with IUGR and LGA. Placenta 2015, 36, 552–558. [Google Scholar] [CrossRef]
- Kamath, U.; Rao, G.; Raghothama, C.; Rai, L.; Rao, P. Erythrocyte Indicators of Oxidative Stress in Gestational Diabetes. Acta Paediatr. Int. J. Paediatr. 1998, 87, 676–679. [Google Scholar] [CrossRef]
- Jarmuzek, P.; Wielgos, M.; Bomba-Opon, D.A. Placental Pathologic Changes in Gestational Diabetes Mellitus. Neuroendocrinol. Lett. 2015, 36, 101–105. [Google Scholar]
- van den Bosch, W.J.H.M. The Practice Guideline “Problematic Alcohol Consumption” (Second Revision) from the Dutch College of General Practitioners. Ned. Tijdschr. Geneeskd. 2006, 150, 2523–2524. [Google Scholar]
- Alexander, B.T.; Dasinger, J.H.; Intapad, S. Fetal Programming and Cardiovascular Pathology. Compr. Physiol. 2017, 176, 139–148. [Google Scholar]
- Phipps, E.A.; Thadhani, R.; Benzing, T.; Karumanchi, S.A. Pre-Eclampsia: Pathogenesis, Novel Diagnostics and Therapies. Nat. Rev. Nephrol. 2019, 15, 275–289. [Google Scholar] [CrossRef]
- MacDonald, T.M.; Walker, S.P.; Hannan, N.J.; Tong, S.; Kaitu’u-Lino, T.J. Clinical Tools and Biomarkers to Predict Preeclampsia. eBioMedicine 2022, 75, 103780. [Google Scholar] [CrossRef]
- Hutcheon, J.A.; Lisonkova, S.; Joseph, K.S. Epidemiology of Pre-Eclampsia and the Other Hypertensive Disorders of Pregnancy. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Offer, E.R.E.Z.; Romero, R.; Eunjung, J.U.N.G.; Chaemsaithong, P.; Bosco, M.; Suksai, M.; Gotsch, F. Preeclampsia/Eclampsia: The Conceptual Evolution of a Syndrome. Am. J. Obs. Gynecol. 2022, 226, 1–42. [Google Scholar] [CrossRef]
- Bartsch, E.; Medcalf, K.E.; Park, A.L.; Ray, J.G.; Al-Rubaie, Z.T.A.; Askie, L.M.; Berger, H.; Blake, J.; Graves, L.; Kingdom, J.C.; et al. Clinical Risk Factors for Pre-Eclampsia Determined in Early Pregnancy: Systematic Review and Meta-Analysis of Large Cohort Studies. BMJ 2016, 353, i1753. [Google Scholar] [CrossRef] [PubMed]
- Southcombe, J.H.; Redman, C.W.G.; Sargent, I.L.; Granne, I. Interleukin-1 Family Cytokines and Their Regulatory Proteins in Normal Pregnancy and Pre-Eclampsia. Clin. Exp. Immunol. 2015, 181, 480–490. [Google Scholar] [CrossRef]
- Wang, Y.; Li, B.; Zhao, Y. Inflammation in Preeclampsia: Genetic Biomarkers, Mechanisms, and Therapeutic Strategies. Front. Immunol. 2022, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Gil-Acevedo, L.; Ceballos, G.; Torres-Ramos, Y. Foetal Lipoprotein Oxidation and Preeclampsia. Lipids Health Dis. 2022, 21, 51. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, S.; Inversetti, A.; Causarano, V.; Stenirri, S.; Soriani, N.; Ambrosi, A.; Valsecchi, L.; Candiani, M.; Cremonesi, L.; Ferrari, M.; et al. HIF1A and MIF as Potential Predictive MRNA Biomarkers of Pre-Eclampsia: A Longitudinal Prospective Study in High Risk Population. Clin. Chem. Lab. Med. 2015, 53, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Subha, M.; Pal, P.; Pal, G.K.; Habeebullah, S.; Adithan, C.; Sridhar, M.G. Decreased Baroreflex Sensitivity Is Linked to Sympathovagal Imbalance, Low-Grade Inflammation, and Oxidative Stress in Pregnancy-Induced Hypertension. Clin. Exp. Hypertens. 2016, 38, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, K.K.; Meeker, J.D.; McElrath, T.F.; Mukherjee, B.; Cantonwine, D.E. Repeated Measures of Inflammation and Oxidative Stress Biomarkers in Preeclamptic and Normotensive Pregnancies. Am. J. Obstet. Gynecol. 2017, 216, 527.e1–527.e9. [Google Scholar] [CrossRef]
- Karthiga, K.; Pal, G.K.; Dasari, P.; Nanda, N.; Velkumary, S.; Chinnakali, P. Attenuation of Baroreflex Sensitivity and Heart Rate Variability Is Linked to Reduced Levels of Nitric Oxide in Pregnant Women Having Risks of Developing Gestational Hypertension. Clin. Exp. Hypertens. 2021, 43, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Talab, A.Y.; Hamza, H.A.; Mostafa, T.M. Antepartum and Postpartum Changes in Adipokines, Endothelial Dysfunction, Inflammatory Markers and Other Biochemical Parameters in Preeclamptic Women: A Prospective Observational Cohort Study. J. Appl. Biomed. 2021, 19, 62–72. [Google Scholar] [CrossRef]
- Molvarec, A.; Szarka, A.; Walentin, S.; Beko, G.; Karádi, I.; Prohászka, Z.; Rigó, J. Serum Heat Shock Protein 70 Levels in Relation to Circulating Cytokines, Chemokines, Adhesion Molecules and Angiogenic Factors in Women with Preeclampsia. Clin. Chim. Acta 2011, 412, 1957–1962. [Google Scholar] [CrossRef]
- Catarino, C.; Santos-Silva, A.; Belo, L.; Rocha-Pereira, P.; Rocha, S.; Patrício, B.; Quintanilha, A.; Rebelo, I. Inflammatory Disturbances in Preeclampsia: Relationship between Maternal and Umbilical Cord Blood. J. Pregnancy 2012, 2012, 684384. [Google Scholar] [CrossRef]
- Bernardi, F.C.B.; Felisberto, F.; Vuolo, F.; Petronilho, F.; Souza, D.R.; Luciano, T.F.; De Souza, C.T.; Ritter, C.; Dal-Pizzol, F. Oxidative Damage, Inflammation, and Toll-like Receptor 4 Pathway Are Increased in Preeclamptic Patients: A Case-Control Study. Oxid. Med. Cell. Longev. 2012, 2012, 636419. [Google Scholar] [CrossRef]
- Ouyang, Y.Q.; Li, S.J.; Zhang, Q.; Cai, H.B.; Chen, H.P. Interactions between Inflammatory and Oxidative Stress in Preeclampsia. Hypertens. Pregnancy 2009, 28, 56–62. [Google Scholar] [CrossRef]
- Molvarec, A.; Rigó, J.; Lázár, L.; Balogh, K.; Makó, V.; Cervenak, L.; Mézes, M.; Prohászka, Z. Increased Serum Heat-Shock Protein 70 Levels Reflect Systemic Inflammation, Oxidative Stress and Hepatocellular Injury in Preeclampsia. Cell Stress Chaperones 2009, 14, 151–159. [Google Scholar] [CrossRef]
- Silva, D.M.C.E.; Marreiro, D.D.N.; Moita Neto, J.M.; Brito, J.A.; Neta, E.A.D.S.; Matias, J.P.; Sampaio, F.A.; Nogueira, N.D.N. Oxidative Stress and Immunological Alteration in Women with Preeclampsia. Hypertens. Pregnancy 2013, 32, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.Q.; Li, S.J.; Zhang, Q.; Xiang, W.P.; Shen, H.L.; Chen, H.P.; Chen, H.; Chen, H.Z. Plasma SFlt-1-to-PlGF Ratio Is Correlated with Inflammatory but Not with Oxidative Stress in Chinese Preeclamptic Women. Arch. Gynecol. Obstet. 2009, 280, 91–97. [Google Scholar] [CrossRef]
- Wang, C.N.; Chang, S.D.; Peng, H.H.; Lee, Y.S.; Chang, Y.L.; Cheng, P.J.; Chao, A.S.; Wang, T.H.; Wang, H.S. Change in Amniotic Fluid Levels of Multiple Anti-Angiogenic Proteins before Development of Preeclampsia and Intrauterine Growth Restriction. J. Clin. Endocrinol. Metab. 2010, 95, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Ekun, O.A.; Ogidi, N.O.; Lawal, R.A.; Ogunmuyiwa, O.A.; Umewune, M.C.; Adefolaju, F.O.; Oshundun, M.F.; Oremosu, A.I. Interrelationship between Markers of Oxidative Stress, Inflammation and Hematological Parameters Among Preeclamptic Nigerian Women. Med. Sci. Monit. Basic Res. 2018, 24, 225–231. [Google Scholar] [CrossRef]
- Stenczer, B.; Rigó, J.; Prohászka, Z.; Derzsy, Z.; Lázár, L.; Makó, V.; Cervenak, L.; Balogh, K.; Mézes, M.; Karádi, I.; et al. Plasma Osteopontin Concentrations in Preeclampsia—Is There an Association with Endothelial Injury? Clin. Chem. Lab. Med. 2010, 48, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, L.; Wang, X.; Liu, Y. Association between Serum Beta-Human Chorionic Gonadotropin and Inflammation, Oxidative Stress in Pregnancy-Induced Hypertension. Microvasc. Res. 2021, 135, 104130. [Google Scholar] [CrossRef]
- Barden, A.; Beilin, L.J.; Ritchie, J.; Croft, K.D.; Walters, B.N.; Michael, C.A. Plasma and Urinary 8-Iso-Prostane as an Indicator of Lipid Peroxidation in Pre-Eclampsia and Normal Pregnancy. Clin. Sci. 1996, 91, 711–718. [Google Scholar] [CrossRef]
- Ishihara, O.; Hayashi, M.; Osawa, H.; Kobayashi, K.; Takeda, S.; Vessby, B.; Basu, S. Isoprostanes, Prostaglandins and Tocopherols in Pre-Eclamsia, Normal Pregnancy and Non-Pregnancy. Free Radic. Res. 2004, 38, 913–918. [Google Scholar] [CrossRef]
- de Sousa Rocha, V.; Della Rosa, F.B.; Ruano, R.; Zugaib, M.; Colli, C. Association between Magnesium Status, Oxidative Stress and Inflammation in Preeclampsia: A Case-Control Study. Clin. Nutr. 2015, 34, 1166–1171. [Google Scholar] [CrossRef]
- Ferreira, R.C.; Fragoso, M.B.T.; dos Santos Tenório, M.C.; da Paz Martins, A.S.; Borbely, A.U.; Moura, F.A.; Goulart, M.O.F.; de Oliveira, A.C.M. Biomarkers of Placental Redox Imbalance in Pregnancies with Preeclampsia and Consequent Perinatal Outcomes. Arch. Biochem. Biophys. 2020, 691, 108464. [Google Scholar] [CrossRef]
- Mannaerts, D.; Faes, E.; Cos, P.; Briedé, J.J.; Gyselaers, W.; Cornette, J.; Gorbanev, Y.; Bogaerts, A.; Spaanderman, M.; Van Craenenbroeck, E.; et al. Oxidative Stress in Healthy Pregnancy and Preeclampsia Is Linked to Chronic Inflammation, Iron Status and Vascular Function. PLoS ONE 2018, 13, e0202919. [Google Scholar] [CrossRef]
- García-Gómez, E.; Bobadilla-Bravo, M.; Díaz-Díaz, E.; Vázquez-Martínez, E.R.; Nava-Salazar, S.; Torres-Ramos, Y.; García-Romero, C.S.; Camacho-Arroyo, I.; Cerbón, M. High Plasmatic Levels of Advanced Glycation End Products Are Associated with Metabolic Alterations and Insulin Resistance in Preeclamptic Women. Curr. Mol. Med. 2020, 20, 751–759. [Google Scholar] [CrossRef]
- Gandley, R.E.; Ph, D.; Rohland, J.; Zhou, Y.; Shibata, E.; Gail, F.; Rajakumar, A.; Ph, D.; Kagan, V.E.; Ph, D.; et al. Increased Myeloperoxidase in the Placenta and circulation of women with preeclampsia. Hypertension 2008, 52, 387–393. [Google Scholar] [CrossRef]
- Bainbridge, S.A.; Deng, J.S.; Roberts, J.M. Increased Xanthine Oxidase in the Skin of Preeclamptic Women. Reprod. Sci. 2009, 16, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Rees, A.; Richards, O.; Chambers, M.; Jenkins, B.J.; Cronin, J.G.; Thornton, C.A. Immunometabolic Adaptation and Immune Plasticity in Pregnancy and the Bi-Directional Effects of Obesity. Clin. Exp. Immunol. 2022, 208, 132–146. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.J.; Rigel, J.; Kishton, J.R. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Loftus, R.M.; Finlay, D.K. Immunometabolism: Cellular Metabolism Turns Immune Regulator. J. Biol. Chem. 2016, 291, 1–10. [Google Scholar] [CrossRef]
- Joo, E.H.; Kim, Y.R.; Kim, N.; Jung, J.E.; Han, S.H.; Cho, H.Y. Effect of Endogenic and Exogenic Oxidative Stress Triggers on Adverse Pregnancy Outcomes: Preeclampsia, Fetal Growth Restriction, Gestational Diabetes Mellitus and Preterm Birth. Int. J. Mol. Sci. 2021, 22, 10122. [Google Scholar] [CrossRef]
- Kasture, V.; Sahay, A.; Joshi, S. Cell Death Mechanisms and Their Roles in Pregnancy Related Disorders, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2021; Volume 126, ISBN 9780323853170. [Google Scholar]
- Langley-Evans, S.C.; Pearce, J.; Ellis, S. Overweight, Obesity and Excessive Weight Gain in Pregnancy as Risk Factors for Adverse Pregnancy Outcomes: A Narrative Review. J. Hum. Nutr. Diet. 2022, 35, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Wawer, A.A.; Hodyl, N.A.; Fairweather-Tait, S.; Froessler, B. Are Pregnant Women Who Are Living with Overweight or Obesity at Greater Risk of Developing Iron Deficiency/Anaemia? Nutrients 2021, 13, 1572. [Google Scholar] [CrossRef]
- Álvarez, D.; Muñoz, Y.; Ortiz, M.; Maliqueo, M.; Chouinard-Watkins, R.; Valenzuela, R. Impact of Maternal Obesity on the Metabolism and Bioavailability of Polyunsaturated Fatty Acids during Pregnancy and Breastfeeding. Nutrients 2021, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Scifres, C.M.; Catov, J.M. The Impact of Maternal Obesity and Gestational Weight Gain on Early and Mid-Pregnancy Lipid Profiles. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Maguire, R.L.; House, J.S.; Lloyd, D.T.; Skinner, H.G.; Allen, T.K.; Raffi, A.M.; Skaar, D.A.; Park, S.S.; McCullough, L.E.; Kollins, S.H.; et al. Associations between Maternal Obesity, Gestational Cytokine Levels, and Child Obesity in the NEST Cohort. Pediatr. Obes. 2021, 176, 139–148. [Google Scholar] [CrossRef] [PubMed]
- León-Reyes, G.; Guzmán-Grenfell, A.M.; Medina-Navarro, R.; Montoya-Estrada, A.; Moreno-Eutimio, M.A.; Fuentes-García, S.; Perichart Perera, O.; Muñoz-Manrique, C.; Espino y Sosa, S.; Hicks, G.J.J.; et al. Is Gestational Diabetes Mellitus in Obese Women Predicted by Oxidative Damage in Red Blood Cells? Gynecol. Endocrinol. 2018, 34, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Mostafavi, E.; Nargesi, A.A.; Asbagh, F.A.; Ghazizadeh, Z.; Heidari, B.; Mirmiranpoor, H.; Esteghamati, A.; Vigneron, C.; Nakhjavani, M. Abdominal Obesity and Gestational Diabetes: The Interactive Role of Magnesium. Magnes. Res. 2015, 28, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Solis-Paredes, M.; Estrada-Gutierrez, G.; Perichart-Perera, O.; Montoya-Estrada, A.; Guzmán-Huerta, M.; Borboa-Olivares, H.; Bravo-Flores, E.; Cardona-Pérez, A.; Zaga-Clavellina, V.; Garcia-Latorre, E.; et al. Key Clinical Factors Predicting Adipokine and Oxidative Stress Marker Concentrations among Normal, Overweight and Obese Pregnant Women Using Artificial Neural Networks. Int. J. Mol. Sci. 2018, 19, 86. [Google Scholar] [CrossRef]
- Malti, N.; Merzouk, H.; Merzouk, S.A.; Loukidi, B.; Karaouzene, N.; Malti, A.; Narce, M. Oxidative Stress and Maternal Obesity: Feto-Placental Unit Interaction. Placenta 2014, 35, 411–416. [Google Scholar] [CrossRef]
- Hernández-Trejo, M.; Montoya-Estrada, A.; Torres-Ramos, Y.; Espejel-Núñez, A.; Guzmán-Grenfell, A.; Morales-Hernández, R.; Tolentino-Dolores, M.; Laresgoiti-Servitje, E. Oxidative Stress Biomarkers and Their Relationship with Cytokine Concentrations in Overweight/Obese Pregnant Women and Their Neonates. BMC Immunol. 2017, 18, 3. [Google Scholar] [CrossRef]
- Dao, M.C.; Sen, S.; Iyer, C.; Klebenov, D.; Meydani, S.N. Obesity during Pregnancy and Fetal Iron Status: Is Hepcidin the Link? J. Perinatol. 2013, 33, 177–181. [Google Scholar] [CrossRef]
- Zambon, M.; Mandò, C.; Lissoni, A.; Anelli, G.M.; Novielli, C.; Cardellicchio, M.; Leone, R.; Monari, M.N.; Massari, M.; Cetin, I.; et al. Inflammatory and Oxidative Responses in Pregnancies with Obesity and Periodontal Disease. Reprod. Sci. 2018, 25, 1474–1484. [Google Scholar] [CrossRef]
- Santos-Rosendo, C.; Bugatto, F.; González-Domínguez, A.; Lechuga-Sancho, A.M.; Mateos, R.M.; Visiedo, F. Placental Adaptive Changes to Protect Function and Decrease Oxidative Damage in Metabolically Healthy Maternal Obesity. Antioxidants 2020, 9, 794. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Iyer, C.; Meydani, S.N. Obesity during Pregnancy Alters Maternal Oxidant Balance and Micronutrient Status. J. Perinatol. 2014, 34, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Solis Paredes, J.M.; Perichart Perera, O.; Montoya Estrada, A.; Reyes Muñoz, E.; Espino, Y.; Sosa, S.; Ortega Castillo, V.; Medina Bastidas, D.; Tolentino Dolores, M.; Sanchez Martinez, M.; et al. Gestational Weight Gain Influences the Adipokine-Oxidative Stress Association during Pregnancy. Obes. Facts 2021, 14, 604–612. [Google Scholar] [CrossRef]
- Xie, W.; Wang, Y.; Xiao, S.; Qiu, L.; Yu, Y.; Zhang, Z. Association of Gestational Diabetes Mellitus with Overall and Type Specific Cardiovascular and Cerebrovascular Diseases: Systematic Review and Meta-Analysis. BMJ 2022, 378, e070244. [Google Scholar] [CrossRef]
- Sweeting, A.; Wong, J.; Murphy, H.R.; Ross, G.P. A Clinical Update on Gestational Diabetes Mellitus. Endocr. Rev. 2022, 43, 763–793. [Google Scholar] [CrossRef] [PubMed]
- Gaumnitz, T.; Jain, A.; Pertot, Y.; Huppert, M.; Jordan, I.; Ardana-Lamas, F.; Wörner, H.J. Streaking of 43-Attosecond Soft-X-Ray Pulses Generated by a Passively CEP-Stable Mid-Infrared Driver. Opt. Express 2017, 25, 27506. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, J.; Wen, X.; Duan, J.; Xue, D.; Pan, Y.; Sun, J.; Zhang, W.; Cheng, X.; Wang, C. Proteome Profiling of Gestational Diabetes Mellitus at 16–18 Weeks Revealed by LC-MS/MS. J. Clin. Lab. Anal. 2020, 34, e23424. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E.; Herrero, L.; Naaz, A. Obesity, Inflammation, and Insulin Resistance. Gastroenterology 2007, 132, 2169–2180. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, X.; Wu, Y.; Duan, R.; Zhang, J.; Du, F.; Zhang, Q.; Li, Y.; Li, N. Effects of Vaspin on Pancreatic β Cell Secretion via PI3K/Akt and NF-ΚB Signaling Pathways. PLoS ONE 2017, 12, e0189722. [Google Scholar] [CrossRef]
- Li, C.; Cheng, H.; Adhikari, B.K.; Wang, S.; Yang, N.; Liu, W.; Sun, J.; Wang, Y. The Role of Apelin–APJ System in Diabetes and Obesity. Front. Endocrinol. 2022, 13, 1–11. [Google Scholar] [CrossRef]
- Sperling, M.; Grzelak, T.; Pelczyńska, M.; Bogdański, P.; Formanowicz, D.; Czyżewska, K. Association of Serum Omentin-1 Concentration with the Content of Adipose Tissue and Glucose Tolerance in Subjects with Central Obesity. Biomedicines 2023, 11, 331. [Google Scholar] [CrossRef]
- Briana, D.D.; Malamitsi-Puchner, A. Reviews: Adipocytokines in Normal and Complicated Pregnancies. Reprod. Sci. 2009, 16, 921–937. [Google Scholar] [CrossRef]
- Usluoğullari, B.; Usluogullari, C.A.; Balkan, F.; Orkmez, M. Role of Serum Levels of Irisin and Oxidative Stress Markers in Pregnant Women with and without Gestational Diabetes. Gynecol. Endocrinol. 2017, 33, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Basu, J.; Datta, C.; Chowdhury, S.; Mandal, D.; Mondal, N.K.; Ghosh, A. Gestational Diabetes Mellitus in a Tertiary Care Hospital of Kolkata, India: Prevalence, Pathogenesis and Potential Disease Biomarkers. Exp. Clin. Endocrinol. Diabetes 2020, 128, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Dincgez Cakmak, B.; Dundar, B.; Ketenci Gencer, F.; Yildiz, D.E.; Bayram, F.; Ozgen, G.; Aydin Boyama, B. Assessment of Relationship between Serum Vascular Adhesion Protein-1 (VAP-1) and Gestational Diabetes Mellitus. Biomarkers 2019, 24, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Lappas, M.; Mittion, A.; Permezel, M. In Response to Oxidative Stress, the Expression of Inflammatory Cytokines and Antioxidant Enzymes Are Impaired in Placenta, but Not Adipose Tissue, of Women with Gestational Diabetes. J. Endocrinol. 2010, 204, 75–84. [Google Scholar] [CrossRef]
- Kurepa, D.; Pramanik, A.K.; Kakkilaya, V.; Caldito, G.; Groome, L.J.; Bocchini, J.A.; Jain, S.K. Elevated Acetoacetate and Monocyte Chemotactic Protein-1 Levels in Cord Blood of Infants of Diabetic Mothers. Neonatology 2012, 102, 163–168. [Google Scholar] [CrossRef]
- Martino, J.; Sebert, S.; Segura, M.T.; Garcia-Valdés, L.; Florido, J.; Padilla, M.C.; Marcos, A.; Rueda, R.; McArdle, H.J.; Budge, H.; et al. Maternal Body Weight and Gestational Diabetes Differentially Influence Placental and Pregnancy Outcomes. J. Clin. Endocrinol. Metab. 2016, 101, 59–68. [Google Scholar] [CrossRef]
- Renugasundari, M.; Pal, G.K.; Chaturvedula, L.; Nanda, N.; Harichandrakumar, K.T.; Durgadevi, T. Inflammation and Decreased Cardiovagal Modulation Are Linked to Stress and Depression at 36th Week of Pregnancy in Gestational Diabetes Mellitus. Sci. Rep. 2023, 13, 10348. [Google Scholar] [CrossRef]
- Yakut, K.; Öcal, D.F.; Öztürk, F.H.; Öztürk, M.; Oğuz, Y.; Sınacı, S.; Çağlar, T. Is GDF-15 Level Associated with Gestational Diabetes Mellitus and Adverse Perinatal Outcomes? Taiwan J. Obstet. Gynecol. 2021, 60, 221–224. [Google Scholar] [CrossRef]
- Mandò, C.; Anelli, G.M.; Novielli, C.; Panina-Bordignon, P.; Massari, M.; Mazzocco, M.I.; Cetin, I. Impact of Obesity and Hyperglycemia on Placental Mitochondria. Oxid. Med. Cell. Longev. 2018, 2018, 2378189. [Google Scholar] [CrossRef]
- Clausen, T.D.; Mathiesen, E.R.; Hansen, T.; Pedersen, O.; Jensen, D.M.; Lauenborg, J.; Schmidt, L.; Damm, P. Overweight and the Metabolic Syndrome in Adult Offspring of Women with Diet-Treated Gestational Diabetes Mellitus or Type 1 Diabetes. J. Clin. Endocrinol. Metab. 2009, 94, 2464–2470. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhang, S.; Li, W.; Leng, J.; Wang, L.; Liu, H.; Li, W.; Zhang, C.; Qi, L.; Tuomilehto, J.; et al. Maternal Gestational Diabetes Is Associated with Offspring’s Hypertension. Am. J. Hypertens. 2019, 32, 335–342. [Google Scholar] [CrossRef]
- Bayman, M.G.; Inal, Z.O.; Hayiroglu, F.; Ozturk, E.N.Y.; Gezginc, K. Foetal Umbilical Cord Brain-Derived Neurotrophic Factor (BDNF) Levels in Pregnancy with Gestational Diabetes Mellitus. J. Obstet. Gynaecol. 2022, 42, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Taschereau-Charron, A.; Bilodeau, J.F.; Larose, J.; Greffard, K.; Berthiaume, L.; Audibert, F.; Fraser, W.D.; Julien, P.; Rudkowska, I. F 2-Isoprostanes and Fatty Acids Profile in Early Pregnancy Complicated by Pre-Existing Diabetes. Prostaglandins Leukot. Essent. Fat. Acids 2018, 135, 115–120. [Google Scholar] [CrossRef]
- Piuri, G.; Basello, K.; Rossi, G.; Soldavini, C.M.; Duiella, S.; Privitera, G.; Spadafranca, A.; Costanzi, A.; Tognon, E.; Cappelletti, M.; et al. Methylglyoxal, Glycated Albumin, PAF, and TNF-α: Possible Inflammatory and Metabolic Biomarkers for Management of Gestational Diabetes. Nutrients 2020, 12, 479. [Google Scholar] [CrossRef]
- Phoswa, W.N.; Khaliq, O.P. The Role of Oxidative Stress in Hypertensive Disorders of Pregnancy (Preeclampsia, Gestational Hypertension) and Metabolic Disorder of Pregnancy (Gestational Diabetes Mellitus). Oxid. Med. Cell. Longev. 2021, 2021, 5581570. [Google Scholar] [CrossRef]
- Zych, B.; Górka, A.; Myszka, A.; Błoniarz, D.; Siekierzyńska, A.; Błaż, W. Status of Oxidative Stress during Low-Risk Labour: Preliminary Data. Int. J. Environ. Res. Public Health 2023, 20, 157. [Google Scholar] [CrossRef]
- Li, J.; Xu, Y.; Sun, Z.; Cai, Y.; Wang, B.; Zhang, M.; Ban, Y.; Hou, X.; Hao, Y.; Ouyang, Q.; et al. Differential Lipids in Pregnant Women with Subclinical Hypothyroidism and Their Correlation to the Pregnancy Outcomes. Sci. Rep. 2021, 11, 19689. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Peng, H.; Xu, H.; Li, J.; Golovko, M.; Cheng, H.; Lynch, E.C.; Liu, L.; McCauley, N.; Kennedy, L.; et al. Maternal Diet Intervention before Pregnancy Primes Offspring Lipid Metabolism in Liver. Lab. Investig. 2020, 100, 553–569. [Google Scholar] [CrossRef]
- Galvan-Martinez, D.H.; Bosquez-Mendoza, V.M.; Ruiz-Noa, Y.; del Rocio Ibarra-Reynoso, L.; Barbosa-Sabanero, G.; Lazo-De-La-Vega-Monroy, M.L. Nutritional, Pharmacological, and Environmental Programming of NAFLD in Early Life. Am. J. Physiol.—Gastrointest. Liver Physiol. 2023, 324, G99–G114. [Google Scholar] [CrossRef]
- Hagström, H.; Simon, T.G.; Roelstraete, B.; Stephansson, O.; Söderling, J.; Ludvigsson, J.F. Maternal Obesity Increases the Risk and Severity of NAFLD in Offspring. J. Hepatol. 2021, 75, 1042–1048. [Google Scholar] [CrossRef]
- Mikolajczak, A.; Sallam, N.A.; Singh, R.D.; Scheidl, T.B.; Walsh, E.J.; Larion, S.; Huang, C.; Thompson, J.A. Accelerated Developmental Adipogenesis Programs Adipose Tissue Dysfunction and Cardiometabolic Risk in Offspring Born to Dams with Metabolic Dysfunction. Am. J. Physiol.—Endocrinol. Metab. 2021, 321, E581–E591. [Google Scholar] [CrossRef] [PubMed]
- Scheidl, T.B.; Wager, J.L.; Baker, L.G.; Brightwell, A.L.; Melan, K.M.; Larion, S.; Sarr, O.; Regnault, T.; Urbanski, S.J.; Thompson, J. High Maternal Adiposity During Pregnancy Programs an Imbalance in the Lipidome and Predisposes to Diet-Induced Hepatosteatosis in the Offspring. Biosci. Rep. 2023, 43, BSR20231060. [Google Scholar] [CrossRef] [PubMed]
- Reyes, R.B.; Carrocera, L.A.F. Programación Metabólica Fetal. Perinatol. Y Reprod. Humana 2015, 29, 99–105. [Google Scholar] [CrossRef]
- Charlton, F.; Tooher, J.; Rye, K.A.; Hennessy, A. Cardiovascular Risk, Lipids and Pregnancy: Preeclampsia and the Risk of Later Life Cardiovascular Disease. Hear. Lung Circ. 2014, 23, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Adank, M.C.; Johansen, A.K.; Benschop, L.; Van Streun, S.P.; Smak Gregoor, A.M.; Øyri, L.K.L.; Mulder, M.T.; Steegers, E.A.P.; Holven, K.B.; Roeters van Lennep, J.E. Maternal Lipid Levels in Early Pregnancy as a Predictor of Childhood Lipid Levels: A Prospective Cohort Study. BMC Pregnancy Childbirth 2022, 22, 588. [Google Scholar] [CrossRef] [PubMed]
- Bausserman, L.L.; Herbert, P.N.; McAdam, K.P. Heterogeneity of Human Serum Amyloid A Proteins. J. Exp. Med. 1980, 52, 641–656. [Google Scholar] [CrossRef]
- Davidson, W.S.; Shah, A.S.; Sexmith, H.; Gordon, S.M. The HDL Proteome Watch: Compilation of Studies Leads to New Insights on HDL Function. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2022, 1867, 159072. [Google Scholar] [CrossRef]
- Shah, A.S.; Tan, L.; Long, J.L.; Davidson, W.S. Proteomic Diversity of High Density Lipoproteins: Our Emerging Understanding of Its Importance in Lipid Transport and Beyond. J. Lipid Res. 2013, 54, 2575–2585. [Google Scholar] [CrossRef]
- Ouimet, M.; Barrett, T.J.; Fisher, E.A. HDL and Reverse Cholesterol Transport: Basic Mechanisms and Their Roles in Vascular Health and Disease. Circ. Res. 2019, 124, 1505–1518. [Google Scholar] [CrossRef]
- Cuchel, M.; Rader, D.J. Macrophage Reverse Cholesterol Transport: Key to the Regression of Atherosclerosis? Circulation 2006, 113, 2548–2555. [Google Scholar] [CrossRef]
- Chen, C.; Khismatullin, D.B. Oxidized Low-Density Lipoprotein Contributes to Atherogenesis via Co-Activation of Macrophages and Mast Cells. PLoS ONE 2015, 10, e0123088. [Google Scholar] [CrossRef]
- Rysz, J.; Gluba-Brzózka, A.; Rysz-Górzyńska, M.; Franczyk, B. The Role and Function of HDL in Patients with Chronic Kidney Disease and the Risk of Cardiovascular Disease. Int. J. Mol. Sci. 2020, 21, 601. [Google Scholar] [CrossRef] [PubMed]
- Buendía-Abad, M.; García-Palencia, P.; de Pablos, L.M.; Alunda, J.M.; Osuna, A.; Martín-Hernández, R.; Higes, M. First Description of Lotmaria Passim and Crithidia Mellificae Haptomonad Stages in the Honeybee Hindgut. Int. J. Parasitol. 2022, 52, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Stadler, J.T.; Scharnagl, H.; Wadsack, C.; Marsche, G. Preeclampsia Affects Lipid Metabolism and HDL Function in Mothers and Their Offspring. Antioxidants 2023, 12, 795. [Google Scholar] [CrossRef] [PubMed]
- Eren, E. High Density Lipoprotein and It’s Dysfunction. Open Biochem. J. 2012, 6, 78–93. [Google Scholar] [CrossRef]
- Gordon, T.; Castelli, W.P.; Hjortland, M.C.; Kannel, W.B.; Dawber, T.R. High Density Lipoprotein as a Protective Factor against Coronary Heart Disease. The Framingham Study. Am. J. Med. 1977, 62, 707–714. [Google Scholar] [CrossRef]
- Patanapirunhakit, P.; Karlsson, H.; Mulder, M.; Ljunggren, S.; Graham, D.; Freeman, D. Sphingolipids in HDL—Potential Markers for Adaptation to Pregnancy? Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2021, 1866, 158955. [Google Scholar] [CrossRef]
- Luna-Luna, M.; Niesor, E.; Pérez-Méndez, Ó. HDL as Bidirectional Lipid Vectors: Time for New Paradigms. Biomedicines 2022, 10, 1180. [Google Scholar] [CrossRef]
- Chen, R.; Chen, Q.; Zheng, J.; Zeng, Z.; Chen, M.; Li, L.; Zhang, S. Serum Amyloid Protein A in Inflammatory Bowel Disease: From Bench to Bedside. Cell Death Discov. 2023, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Streese, L.; Habisch, H.; Deiseroth, A.; Carrard, J.; Infanger, D.; Schmidt-Trucksäss, A.; Madl, T.; Hanssen, H. Lipoprotein Subclasses Independently Contribute to Subclinical Variance of Microvascular and Macrovascular Health. Molecules 2022, 27, 4760. [Google Scholar] [CrossRef]
- Woudberg, N.J.; Goedecke, J.H.; Blackhurst, D.; Frias, M.; James, R.; Opie, L.H.; Lecour, S. Association between Ethnicity and Obesity with High-Density Lipoprotein (HDL) Function and Subclass Distribution. Lipids Health Dis. 2016, 15, 92. [Google Scholar] [CrossRef]
- Muñoz-Vega, M.; Massó, F.; Páez, A.; Vargas-Alarcón, G.; Coral-Vázquez, R.; Mas-Oliva, J.; Carreón-Torres, E.; Pérez-Méndez, Ó. Hdl-Mediated Lipid Influx to Endothelial Cells Contributes to Regulating Intercellular Adhesion Molecule (Icam)-1 Expression and Enos Phosphorylation. Int. J. Mol. Sci. 2018, 19, 3394. [Google Scholar] [CrossRef] [PubMed]
- Brites, F.; Martin, M.; Guillas, I.; Kontush, A. Antioxidative Activity of High-Density Lipoprotein (HDL): Mechanistic Insights into Potential Clinical Benefit. BBA Clin. 2017, 8, 66–77. [Google Scholar] [CrossRef]
- Jayaraman, S.; Haupt, C.; Gursky, O. Thermal Transitions in Serum Amyloid A in Solution and on the Lipid: Implications for Structure and Stability of Acute-Phase HDL. J. Lipid Res. 2015, 56, 1531–1542. [Google Scholar] [CrossRef]
- Göçmen, A.Y.; Şahin, E.; Semiz, E.; Gümüslü, S. Is Elevated Serum Ceruloplasmin Level Associated with Increased Risk of Coronary Artery Disease? Can. J. Cardiol. 2008, 24, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Khovidhunkit, W.; Kim, M.S.; Memon, R.A.; Shigenaga, J.K.; Moser, A.H.; Feingold, K.R.; Grunfeld, C. Effects of Infection and Inflammation on Lipid and Lipoprotein Metabolism: Mechanisms and Consequences to the Host. J. Lipid Res. 2004, 45, 1169–1196. [Google Scholar] [CrossRef]
- Rosenson, R.S.; Brewer, H.B., Jr.; Ansell, B.J.; Barter, P.; Chapman, M.J.; Heinecke, J.W.; Kontush, A.; Tall, A.R.; Webb, N.R. Dysfunctional HDL and Atherosclerotic Cardiovascular Disease. Nat. Rev. Cardiol. 2016, 13, 48–60. [Google Scholar] [CrossRef]
- Zafaranieh, S.; Stadler, J.T.; Pammer, A.; Marsche, G.; van Poppel, M.N.M.; Desoye, G. The Association of Physical Activity and Sedentary Behavior with Maternal and Cord Blood Anti-Oxidative Capacity and HDL Functionality: Findings of DALI Study. Antioxidants 2023, 12, 827. [Google Scholar] [CrossRef]
- Schliefsteiner, C.; Hirschmugl, B.; Kopp, S.; Curcic, S.; Bernhart, E.M.; Marsche, G.; Lang, U.; Desoye, G.; Wadsack, C. Maternal Gestational Diabetes Mellitus Increases Placental and Foetal Lipoprotein-Associated Phospholipase A2 Which Might Exert Protective Functions against Oxidative Stress. Sci. Rep. 2017, 7, 12628. [Google Scholar] [CrossRef]
- León-Reyes, G.; Espino, Y.; Sosa, S.; Medina-Navarro, R.; Guzmán-Grenfell, A.M.; Medina-Urrutia, A.X.; Fuentes-García, S.; Hicks, G.J.J.; Torres-Ramos, Y.D. Oxidative Modifications of Foetal LDL-c and HDL-c Lipoproteins in Preeclampsia. Lipids Health Dis. 2018, 17, 110. [Google Scholar] [CrossRef] [PubMed]
- Jayalekshmi, V.S.; Jaikumar, V.S.; Mehra, P.; Thulaseedharan, T.; Vinod, V.M.; Ramachandran, S. Differential Expression of Lipid Metabolic Genes in Hypercholesterolemic Rabbit Placenta Predisposes the Offspring to Develop Atherosclerosis in Early Adulthood. Life Sci. 2023, 327, 121823. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, H.; Simental-Mendía, L.E. The Triglycerides and Glucose Index Is Highly Associated with Non-Alcoholic Fatty Liver Disease in Overweight and Obese Women. Ir. J. Med. Sci. 2023, 2023, 11845. [Google Scholar] [CrossRef] [PubMed]
- Mocciaro, G.; Allison, M.; Jenkins, B.; Azzu, V.; Huang-Doran, I.; Herrera-Marcos, L.V.; Hall, Z.; Murgia, A.; Susan, D.; Frontini, M.; et al. Non-Alcoholic Fatty Liver Disease Is Characterised by a Reduced Polyunsaturated Fatty Acid Transport via Free Fatty Acids and High-Density Lipoproteins (HDL). Mol. Metab. 2023, 73, 101728. [Google Scholar] [CrossRef]
- Pelczyńska, M.; Moszak, M.; Wesołek, A.; Bogdański, P. The Preventive Mechanisms of Bioactive Food Compounds against Obesity-Induced Inflammation. Antioxidants 2023, 12, 1232. [Google Scholar] [CrossRef]
- Pathikkal, A.; Puthusseri, B.; Divya, P.; Rudrappa, S.; Chauhan, V.S. Folate Derivatives, 5-Methyltetrahydrofolate and 10-Formyltetrahydrofolate, Protect BEAS-2B Cells from High Glucose–Induced Oxidative Stress and Inflammation. Vitr. Cell. Dev. Biol.—Anim. 2022, 58, 419–428. [Google Scholar] [CrossRef]
- Zhou, K.; Raffoul, J.J. Potential Anticancer Properties of Grape Antioxidants. J. Oncol. 2012, 2012, 803294. [Google Scholar] [CrossRef]
- Delmastro-Greenwood, M.; Freeman, B.A.; Wendell, S.G. Redox-Dependent Anti-Inflammatory Signaling Actions of Unsaturated Fatty Acids. Annu. Rev. Physiol. 2014, 76, 79–105. [Google Scholar] [CrossRef] [PubMed]
- Gong, S.; Ji, X.; Su, J.; Wang, Y.; Yan, X.; Wang, G.; Xiao, B.; Dong, H.; Xiang, X.; Liu, S. Yeast Fermentate Prebiotic Ameliorates Allergic Asthma, Associating with Inhibiting Inflammation and Reducing Oxidative Stress Level through Suppressing Autophagy. Mediat. Inflamm. 2021, 2021, 4080935. [Google Scholar] [CrossRef]
- Dai, Y.; Quan, J.; Xiong, L.; Luo, Y.; Yi, B. Probiotics Improve Renal Function, Glucose, Lipids, Inflammation and Oxidative Stress in Diabetic Kidney Disease: A Systematic Review and Meta-Analysis. Ren. Fail. 2022, 44, 862–880. [Google Scholar] [CrossRef] [PubMed]
- Kelesidis, T.; Jackson, N.; Mccomsey, G.A.; Wang, X.; Elashoff, D.; Dube, M.P.; Brown, T.T.; Yang, O.O.; Stein, J.H.; Currier, J.S. Oxidized Lipoproteins Are Associated with Markers of Inflammation and Immune Activation in HIV-1 Infection. AIDS 2016, 30, 2625–2633. [Google Scholar] [CrossRef]
- Vergès, B. Dyslipidemia in Type 1 Diabetes: A Masked Danger. Trends Endocrinol. Metab. 2020, 31, 422–434. [Google Scholar] [CrossRef]
- Ganjali, S.; Dallinga-Thie, G.M.; Simental-Mendía, L.E.; Banach, M.; Pirro, M.; Sahebkar, A. HDL Functionality in Type 1 Diabetes. Atherosclerosis 2017, 267, 99–109. [Google Scholar] [CrossRef]
- Stadler, J.T.; Marsche, G. Dietary Strategies to Improve Cardiovascular Health: Focus on Increasing High-Density Lipoprotein Functionality. Front. Nutr. 2021, 8, 1–16. [Google Scholar] [CrossRef]
- Cardner, M.; Yalcinkaya, M.; Goetze, S.; Luca, E.; Balaz, M.; Hunjadi, M.; Hartung, J.; Shemet, A.; Kränkel, N.; Radosavljevic, S.; et al. Structure-Function Relationships of HDL in Diabetes and Coronary Heart Disease. JCI Insight 2020, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Luna-Castillo, K.P.; Lin, S.; Muñoz-Valle, J.F.; Vizmanos, B.; López-Quintero, A.; Márquez-Sandoval, F. Functional Food and Bioactive Compounds on the Modulation of the Functionality of Hdl-c: A Narrative Review. Nutrients 2021, 13, 1165. [Google Scholar] [CrossRef]
- Vaisar, T.; Kanter, J.E.; Wimberger, J.; Irwin, A.D.; Gauthier, J.; Wolfson, E.; Bahnam, V.; Wu, I.H.; Shah, H.; Keenan, H.A.; et al. High Concentration of Medium-Sized HDL Particles and Enrichment in HDL Paraoxonase 1 Associate with Protection from Vascular Complications in People with Long-Standing Type 1 Diabetes. Diabetes Care 2020, 43, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Huang, Y.; Levison, B.S.; Gerstenecker, G.; DiDonato, A.J.; Hazen, L.B.; Lee, J.; Gogonea, V.; DiDonato, J.A.; Hazen, S.L. Identification of Critical Paraoxonase 1 Residues Involved in High Density Lipoprotein Interaction. J. Biol. Chem. 2016, 291, 1890–1904. [Google Scholar] [CrossRef] [PubMed]
- Dorantes-Morales, A.; Estrada-Luna, D.; Bautista-Pérez, R.; Betanzos-Cabrera, G.; Luna-Luna, M.; Flores-Castillo, C.; Vargas-Alarcón, G.; Fragoso, J.M.; Pérez-Méndez, Ó.; Carreón-Torres, E.; et al. Microencapsulated Pomegranate Modifies the Composition and Function of High-Density. Molecules 2020, 25, 3297. [Google Scholar] [CrossRef]
- Hernáez, Á.; Castañer, O.; Elosua, R.; Pintó, X.; Estruch, R.; Salas-Salvadó, J.; Corella, D.; Arós, F.; Serra-Majem, L.; Fiol, M.; et al. Mediterranean Diet Improves High-Density Lipoprotein Function in High-Cardiovascular-Risk Individuals. Circulation 2017, 135, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Hernáez, Á.; Fernández-Castillejo, S.; Farràs, M.; Catalán, Ú.; Subirana, I.; Montes, R.; Solà, R.; Muñoz-Aguayo, D.; Gelabert-Gorgues, A.; Díaz-Gil, Ó.; et al. Olive Oil Polyphenols Enhance High-Density Lipoprotein Function in Humans: A Randomized Controlled Trial. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2115–2119. [Google Scholar] [CrossRef] [PubMed]
- Sola, R.; Valls, R.M.; Puzo, J.; Calabuig, J.R.; Brea, A.; Pedret, A.; Morinã, D.; Villar, J.; Millán, J.; Anguera, A. Effects of Poly-Bioactive Compounds on Lipid Profile and Body Weight in a Moderately Hypercholesterolemic Population with Low Cardiovascular Disease Risk: A Multicenter Randomized Trial. PLoS ONE 2014, 9, e101978. [Google Scholar] [CrossRef]
- Bub, A.; Malpuech-Brugère, C.; Orfila, C.; Amat, J.; Arianna, A.; Blot, A.; Di Nunzio, M.; Holmes, M.; Kertész, Z.; Marshall, L.; et al. A Dietary Intervention of Bioactive Enriched Foods Aimed at Adults at Risk of Metabolic Syndrome: Protocol and Results from PATHWAY-27 Pilot Study. Nutrients 2019, 11, 1814. [Google Scholar] [CrossRef]
- Muralidharan, J.; Papandreou, C.; Soria-Florido, M.T.; Sala-Vila, A.; Blanchart, G.; Estruch, R.; Martínez-González, M.A.; Corella, D.; Ros, E.; Ruiz-Canela, M.; et al. Cross-Sectional Associations between HDL Structure or Function, Cell Membrane Fatty Acid Composition, and Inflammation in Elderly Adults. J. Nutr. 2022, 152, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Luna, D.; Carreón-Torres, E.; Bautista-Perez, R.; Betanzos-Cabrera, G.; Dorantes-Morales, A.; Luna-Luna, M.; Vargas-Barrón, J.; Mejía, A.M.; Fragoso, J.M.; Carvajal-Aguilera, K.; et al. Microencapsulated Pomegranate Reverts High-Density Lipoprotein (HDL)-Induced Endothelial Dysfunction and Reduces Postprandial Triglyceridemia in Women with Acute Coronary Syndrome. Nutrients 2019, 11, 1710. [Google Scholar] [CrossRef] [PubMed]



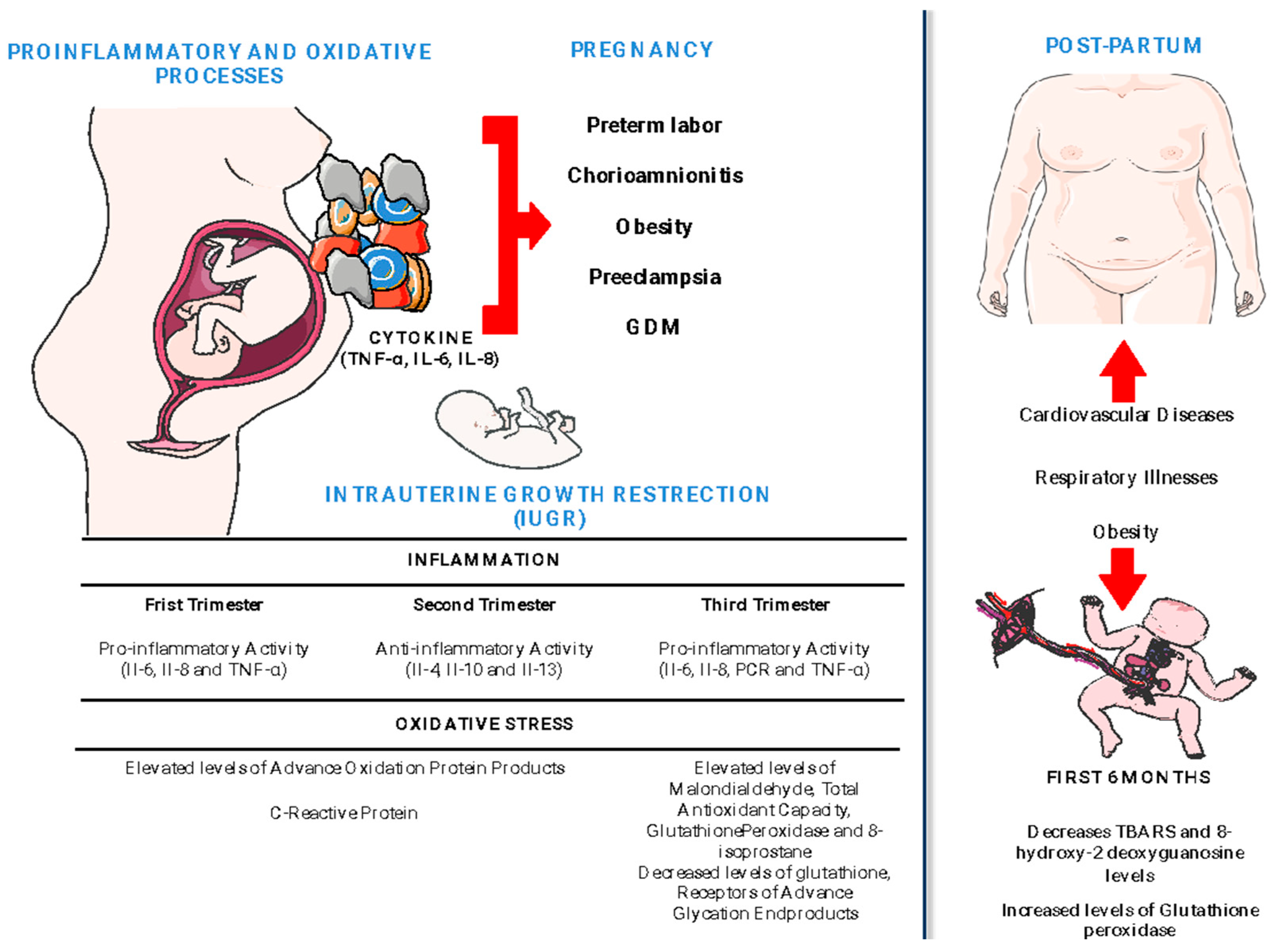
| Trimester | Cytokines | Clinical Outcome in the Mother | Clinical Outcome in the Fetus/Infant | Ref. |
|---|---|---|---|---|
| First Trimester (Proinflammatory) | IL-6 | Infection and chorioamnionitis increased prevalence | Fetal inflammatory response disturbances Reactive airway disease Ectopic lipid accumulation in liver | [61,62,63] |
| IL-8 | Preeclampsia development | Insulin resistance Bronchopulmonary dysplasia | [51,61,63,64] | |
| TNF-α | Increases obesity and GDM risk | Adipose tissue alterations | [61,65] | |
| MCP-1 | Increased risk of miscarriage and GDM risk | Undetermined | [61] | |
| RANTES G-SCF | Increased risk of miscarriage | Undetermined | [61] | |
| Second Trimester | IL-4 | Decreases the risk of preterm birth, preeclampsia Increased risk of systemic lupus erythematosus (SLE) | Undetermined | [61] |
| IL-10 | Decreases the risk of preterm birth and preeclampsia Increases obesity and GDM risk | Decreased expression of the inflammatory cytokine gene in the uterus and placenta | [61,66] | |
| IL-13 | Development of fetal inflammatory response syndrome | Undetermined | [61] | |
| Third Trimester (Proinflammatory) | IL-6 | Increased risk of miscarriage, obesity, GDM, and preeclampsia risk Development of reactive airway disease (RAD) | Development of reactive airway disease (RAD) | [51,61,62,63,65,66,67] |
| IL-8 | Increased prevalence of infection, preeclampsia, and chorioamnionitis Development of fetal inflammatory response syndrome | Undetermined | [61,66] | |
| MCP-1 | Increased risk of miscarriage, autoimmune diseases, and GDM | Undetermined | [61] | |
| RANTES-G-SCF | Increased risk of miscarriage | Undetermined | [61] | |
| TNF-α | Preeclampsia | Wheezing and lower respiratory tract infections. Intrauterine growth restriction | [61,63,67] |
| Study Groups | Oxidative and Inflammatory Markers | OS Changes | Inflammation Changes | Effects in Mothers and Newborns | Ref. |
|---|---|---|---|---|---|
| PW Ctrl = 28 PWOw = 26 PWOb = 26 | MDA, LPH in plasma CP in RBCs Plasmatic IL-6 and TNF-α | ↑ MDA, LOOH and CP in PWOb and PWOw vs. Ctrl in the 3rd trimester vs. the 2nd trimester | ↑ TNF-α in PWOb vs. PW at 12–13 months of gestation ↑ TNF-α and IL-6 in women who developed GDM | + Correlation of LOOH, MDA and CP with gestational age in all groups and higher levels of these markers in PW who developed GDM vs. PWOb in 2nd and 3rd trimesters | [115] |
| Ctrl = 15 PWOb = 15 | Serum iron and transferrin saturation (Tsat), CRP, IL-6, GSH, and GSSG | Ratio of serum GSSG/GSH was higher in the PWOb compared to Ctrl | ↑ TNF-α in PWOb vs. Ctrl. No changes in IL-6 and iron | + Correlation between maternal BMI and CRP. - Correlation between BMI and iron status in cord blood. Serum iron and Tsat were found to be significantly lower in cord blood from PWOb compared to Ctrl | [120] |
| Ctrl = 40 PWOb = 40 | Serum MDA and hsCRP at 28 weeks of gestation | ↑ MDA and glucose after 3 h OGTT were higher in PWOb No changes at baseline. | Higher levels of hsCRP at baseline and after 3 h of OGTT in PWOb vs. Ctrls | After the OGTT at 28 weeks of gestation, 16 patients in the obese group and 1 in the control group had GDM. | [116] |
| Weight gain Insufficient, IWG = 28 Normal, NWG = 20 Excessive, EWG = 26 | Serum LOOH, MDA, CP, 8-oxodG, adiponectin, leptin, and resistin | No significant differences in LOOH, MDA, and CP concentrations between groups. A trend to lower 8-oxodG in PWOb | ↓ Adiponectin in EWG + Correlation between Adiponectin and 8-oxodG concentration in NWG. In EWG, leptin was associated with LOOH, MDA, and CP. | ND | [124] |
| Ctrl = 25 PWOw = 21 PWOb = 22 | Adiponectin, leptin, and resistin in serum. MDA, CP, and 8-oxodG in plasma | ↑ MDA and CP in PWOb vs. Ctrl | ↓ Adiponectin and resistin in PWOb | BMI estimating adiponectin and CP concentrations (37%). Gestational age predicts resistin and MDA (34%). | [117] |
| Ctrl = 15 PWOb = 15 | CRP, GSSH, GSH, TNF-α and IL-6 levels in serum and cord | ↓ GSSG/GSH ratio in PWOb vs. Ctrl | ↑ CRP and IL-6 in PWOb vs. Ctrl. ↑ CRP in cord of PWOb vs. Ctrl | Maternal BMI correlated with CRP, GSSG/GSH ratio, and IL-6. Infants born from PWOb mothers did not have higher measures of OS. | [120] |
| Ctrl = 50 PWOb = 40 | Nitric oxide (NO), CAT, SOD, GSH, CP and MDA in plasma and placenta | ↓ SOD and CAT and GSSG levels in PWOb and their newborns vs. Ctrl ↑ MDA, CP, Nitrite and Red-NTB in plasma PWOb and their newborns vs. Ctrl ↑ CAT, SOD, GSH, MDA and CP in placenta of PWOb vs. Ctrl | + Correlation between maternal and fetal levels of TG and plasma and placental MDA. Maternal nitrite and reduced NBT levels were significantly correlated with those of newborns of PWOb | [118] | |
| Ctrl = 27 PWOb = 18 PW with obesity and GDM (ObGDM) = 17 | TAC and CRP in saliva CRP in plasma | ↑ TAC in saliva in ObGDM vs. PW. + Correlation between TAC and CRP in saliva or plasma | ↑ CRP in saliva and plasma in ObGDM vs. PW. ↑ CRP in plasma in PWOb vs. PW | TAC and CRP correlates positively with BMI and glycemia. A significant interaction between maternal BMI and periodontitis for s-TAC levels. | [121] |
| HDL Function | Proteins and Lipids Associated with HDL Function | Ref. | |
|---|---|---|---|
| Reverse cholesterol transport | HDL promotes cholesterol efflux from various cell types. | ABCAI, ABCG1, SR-BI, cubilin, ApoE receptor | [168,169] |
| Removing excess cholesterol from lipid-laden macrophages is a crucial process in HDL-mediated vascular protection. | |||
| Oxidant | HDL has antioxidant properties whereby it can remove and inactivate lipid peroxides from LDL and cells. | PON 1, Apo AI, PAF-AH, LCAT, Apo M, S1P, phospholipids | [159] |
| Inflammation | Controlling the activation of monocytes, preventing macrophage migration, and inhibiting the oxidation of LDL by blocking the 12-lipoxygenase that produces lipid hydroperoxides and leads to the oxidation of the LDL | VCAM, ICAM, TNF-α, SAA, ceramides | [169] |
| Vascular function | Modulation of endothelial nitric oxide synthase (eNOS) expression, leading to increased nitric oxide (NO) production and vasodilation | ABCA1, SR-BI, S1PR, S1P, Apo M | [170,171,172] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Osorio, A.S.; Carreón-Torres, E.; Correa-Solís, E.; Ángel-García, J.; Arias-Rico, J.; Jiménez-Garza, O.; Morales-Castillejos, L.; Díaz-Zuleta, H.A.; Baltazar-Tellez, R.M.; Sánchez-Padilla, M.L.; et al. Inflammation and Oxidative Stress Induced by Obesity, Gestational Diabetes, and Preeclampsia in Pregnancy: Role of High-Density Lipoproteins as Vectors for Bioactive Compounds. Antioxidants 2023, 12, 1894. https://doi.org/10.3390/antiox12101894
Jiménez-Osorio AS, Carreón-Torres E, Correa-Solís E, Ángel-García J, Arias-Rico J, Jiménez-Garza O, Morales-Castillejos L, Díaz-Zuleta HA, Baltazar-Tellez RM, Sánchez-Padilla ML, et al. Inflammation and Oxidative Stress Induced by Obesity, Gestational Diabetes, and Preeclampsia in Pregnancy: Role of High-Density Lipoproteins as Vectors for Bioactive Compounds. Antioxidants. 2023; 12(10):1894. https://doi.org/10.3390/antiox12101894
Chicago/Turabian StyleJiménez-Osorio, Angélica Saraí, Elizabeth Carreón-Torres, Emmanuel Correa-Solís, Julieta Ángel-García, José Arias-Rico, Octavio Jiménez-Garza, Lizbeth Morales-Castillejos, Hugo Alexander Díaz-Zuleta, Rosa María Baltazar-Tellez, María Luisa Sánchez-Padilla, and et al. 2023. "Inflammation and Oxidative Stress Induced by Obesity, Gestational Diabetes, and Preeclampsia in Pregnancy: Role of High-Density Lipoproteins as Vectors for Bioactive Compounds" Antioxidants 12, no. 10: 1894. https://doi.org/10.3390/antiox12101894