
The Inhibition of Neuropathic Pain Incited by Nerve Injury and Accompanying Mood Disorders by New Heme Oxygenase-1 Inducers: Mechanisms Implicated
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Neuropathic Pain
2.3. Nociceptive Behavioral Responses
2.4. Emotional Behavioral Responses
2.5. Western Blot
2.6. Drugs
2.7. Experimental Design
2.8. Statistical Analyses
3. Results
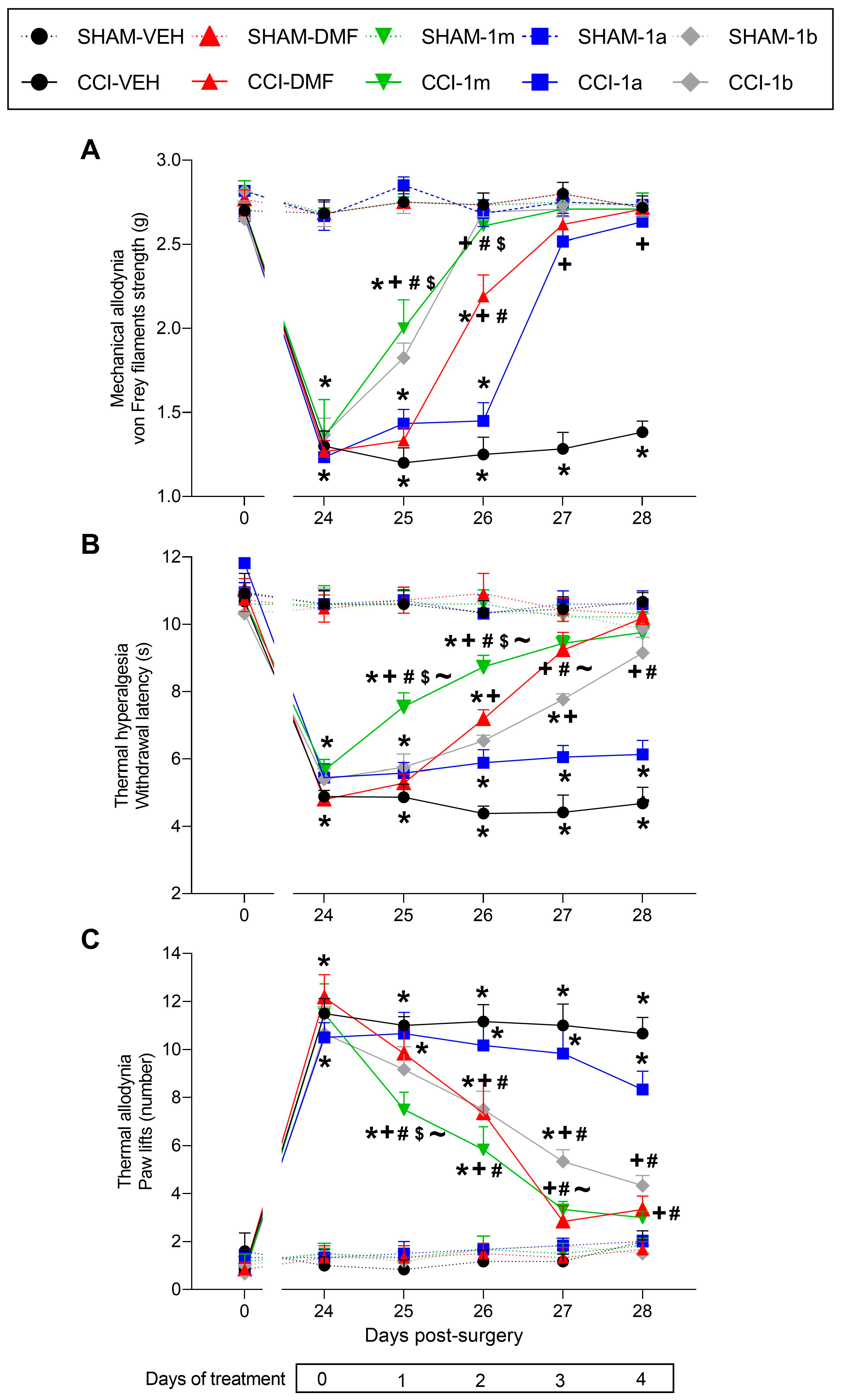
3.1. Effects of 1m, 1b, 1a, and DMF Given Orally on the Hyperalgesia and Allodynia Provoked by Sciatic Nerve Injury
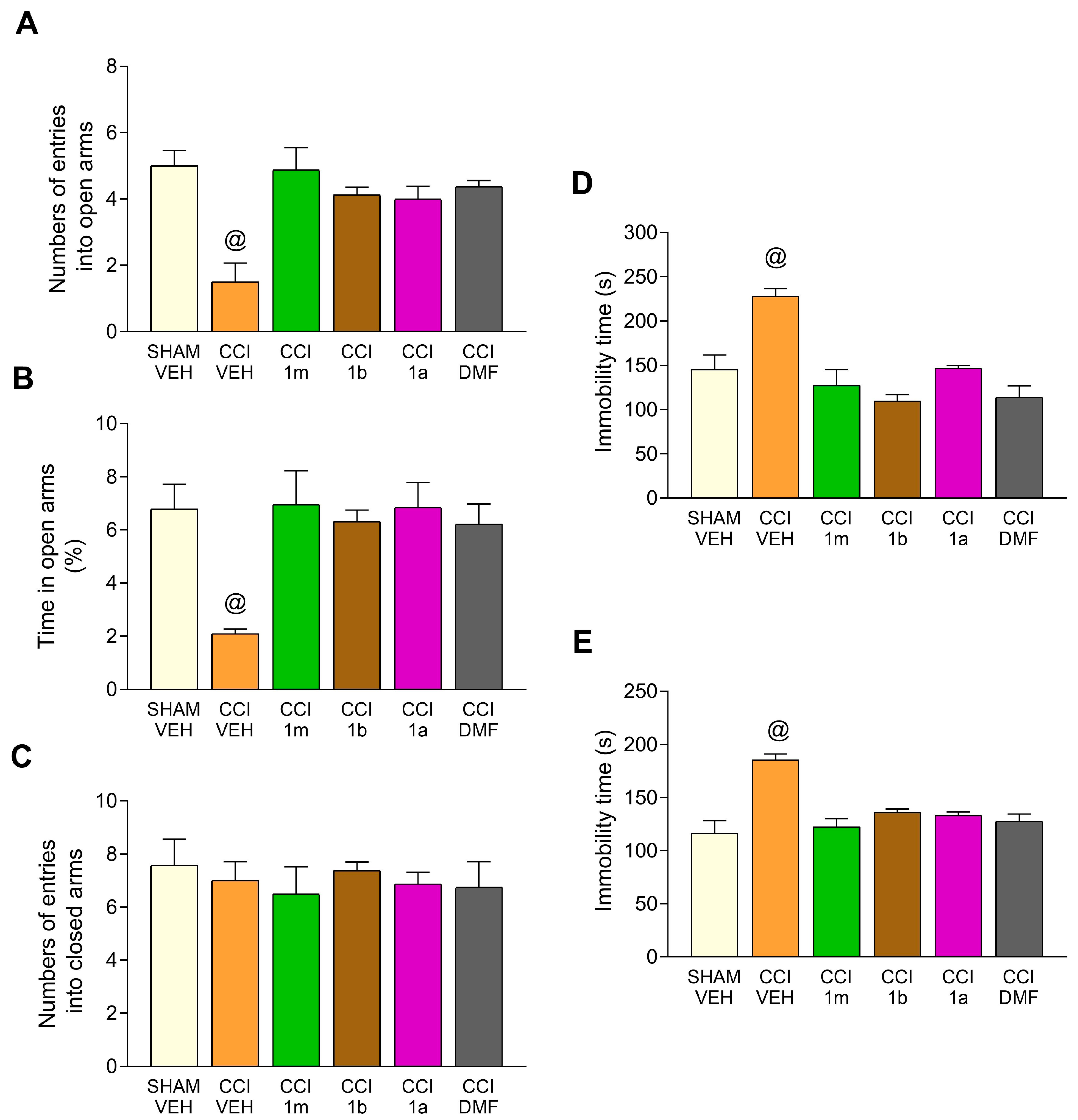
3.2. Impact of the Administration of 1m, 1b, 1a, and DMF on the Anxiety- and Depressive-Related Conducts Accompanying CCI-Provoked Neuropathy in Mice
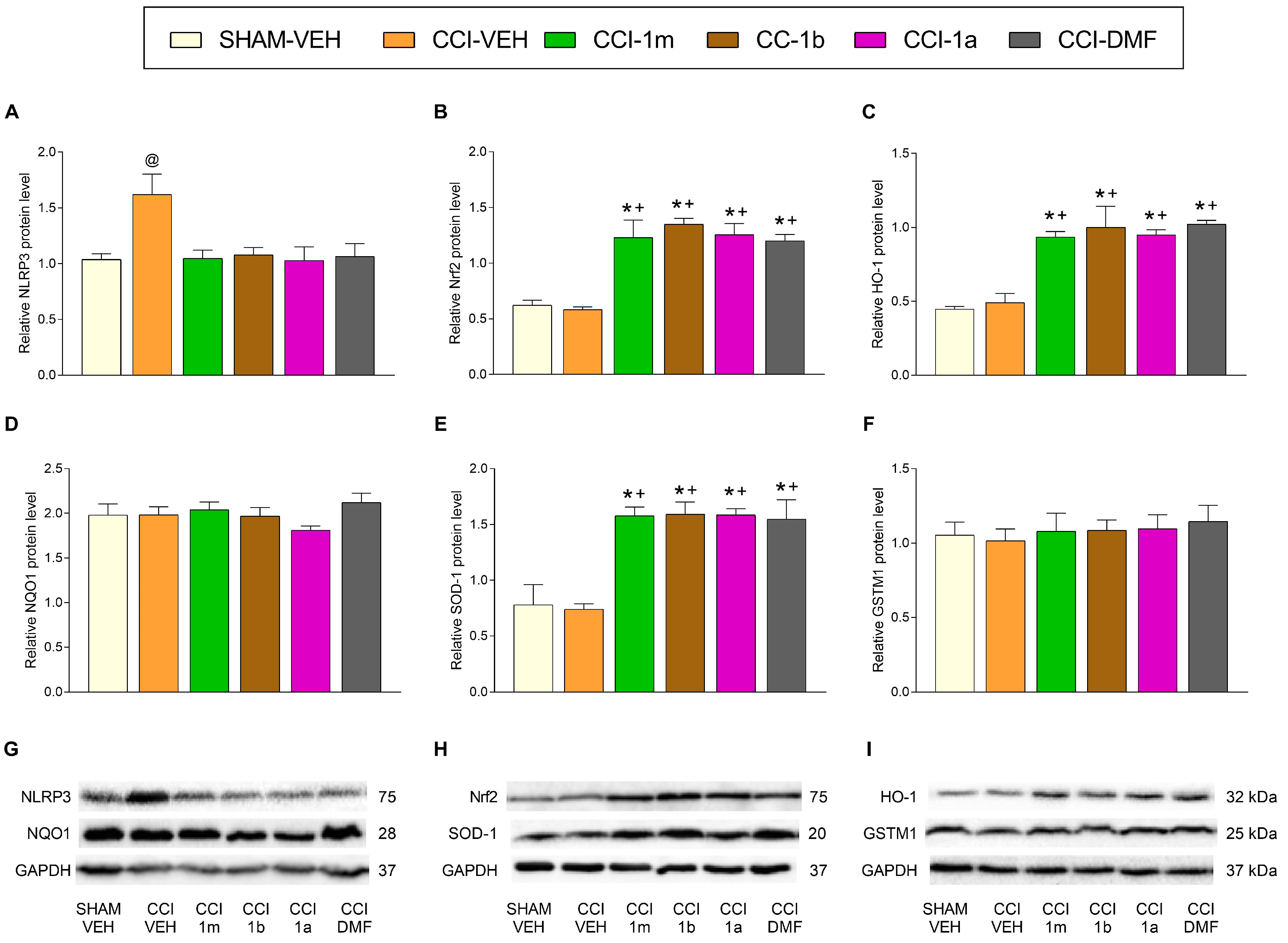
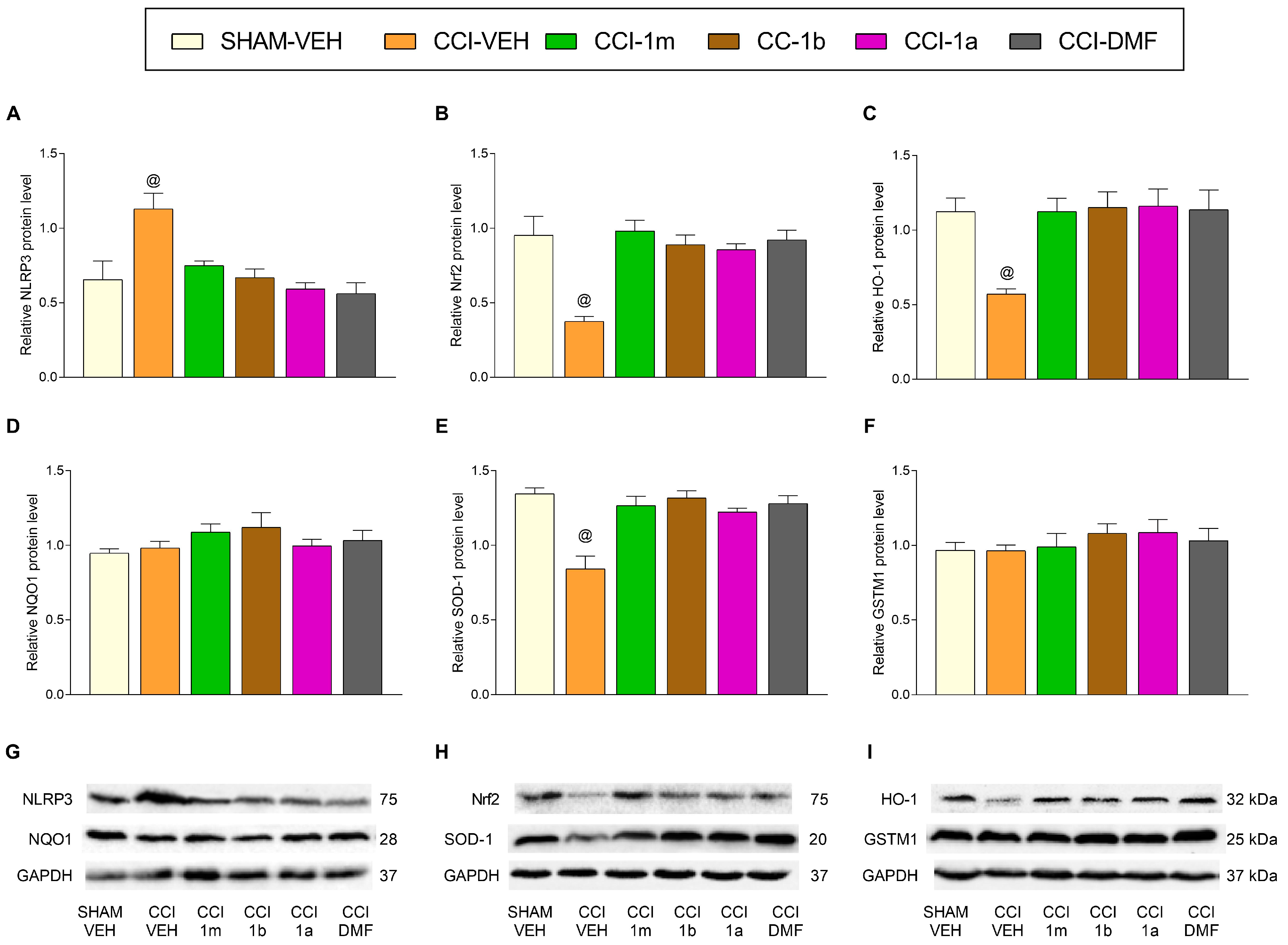
3.3. The Impact of 1m, 1b, 1a, and DMF on the Expression of Inflammatory and Antioxidant Proteins in the DRG and AMG of Animals with Neuropathic Pain
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Finnerup, N.B.; Kuner, R.; Jensen, T.S. Neuropathic Pain: From Mechanisms to Treatment. Physiol. Rev. 2021, 101, 259–301. [Google Scholar] [CrossRef]
- Borgonetti, V.; Galeotti, N. Microglia senescence is related to neuropathic pain-associated comorbidities in the spared nerve injury model. Pain 2023, 164, 1106–1117. [Google Scholar] [CrossRef]
- Gayman, M.D.; Brown, R.K.; Cui, M. Depressive symptoms and bodily pain: The role of physical disability and social stress. Stress. Health 2010, 27, 52–63. [Google Scholar] [CrossRef]
- Fornasari, D. Pharmacotherapy for Neuropathic Pain: A Review. Pain Ther. 2017, 6, 25–33. [Google Scholar] [CrossRef]
- Carrasco, C.; Naziroǧlu, M.; Rodríguez, A.B.; Pariente, J.A. Neuropathic Pain: Delving into the Oxidative Origin and the Possible Implication of Transient Receptor Potential Channels. Front. Physiol. 2018, 9, 95. [Google Scholar] [CrossRef]
- Doyle, T.M.; Salvemini, D. Mini-Review: Mitochondrial dysfunction and chemotherapy-induced neuropathic pain. Neurosci. Letters 2021, 760, 136087. [Google Scholar] [CrossRef]
- Rukavina, K.; Leta, V.; Sportelli, C.; Buhidma, Y.; Duty, S.; Malcangio, M.; Ray Chaudhuri, K. Pain in Parkinson’s disease: New concepts in pathogenesis and treatment. Curr. Opin. Neurology 2019, 32, 579–588. [Google Scholar] [CrossRef]
- Naik, A.K.; Tandan, S.K.; Dudhgaonkar, S.P.; Jadhav, S.H.; Kataria, M.; Prakash, V.R.; Kumar, D. Role of oxidative stress in pathophysiology of peripheral neuropathy and modulation by N-acetyl-L-cysteine in rats. Eur. J. Pain 2006, 10, 573–579. [Google Scholar] [CrossRef]
- Chen, S.M.; Wang, M.H.; Soung, H.S.; Tseng, H.C.; Fang, C.H.; Lin, Y.W.; Yang, C.C.; Tsai, C.C. Neuroprotective effect of l-theanine in a rat model of chronic constriction injury of sciatic nerve-induced neuropathic pain. J. Formos. Med. Assoc. 2022, 121, 802–814. [Google Scholar] [CrossRef]
- Tanabe, M.; Nagatani, Y.; Saitoh, K.; Takasu, K.; Ono, H. Pharmacological assessments of nitric oxide synthase isoforms and downstream diversity of NO signaling in the maintenance of thermal and mechanical hypersensitivity after peripheral nerve injury in mice. Neuropharmacology 2009, 56, 702–708. [Google Scholar] [CrossRef]
- Teixeira-Santos, L.; Albino-Teixeira, A.; Pinho, D. Neuroinflammation, oxidative stress and their interplay in neuropathic pain: Focus on specialized pro-resolving mediators and NADPH oxidase inhibitors as potential therapeutic strategies. Pharmacol. Res. 2020, 162, 105280. [Google Scholar] [CrossRef] [PubMed]
- Tonkin, R.S.; Bowles, C.; Perera, C.J.; Keating, B.A.; Makker, P.G.S.; Duffy, S.S.; Lees, J.G.; Tran, C.; Don, A.S.; Fath, T.; et al. Attenuation of mechanical pain hypersensitivity by treatment with Peptide5, a connexin-43 mimetic peptide, involves inhibition of NLRP3 inflammasome in nerve-injured mice. Exp. Neurol. 2018, 300, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, Q.; Jiang, W.; Yu, S.; Zhang, S. MiR-34c Ameliorates Neuropathic Pain by Targeting NLRP3 in a Mouse Model of Chronic Constriction Injury. Neuroscience 2019, 399, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, T.; Lu, M.; El-Kenawy, A.E. Potential anxiolytic and antidepressant-like effects of luteolin in a chronic constriction injury rat model of neuropathic pain: Role of oxidative stress, neurotrophins, and inflammatory factors. Int. Immunopharmacol. 2023, 122, 110520. [Google Scholar] [CrossRef]
- Shan, W.; Liao, X.; Tang, Y.; Liu, J. Dexmedetomidine alleviates inflammation in neuropathic pain by suppressing NLRP3 via Nrf2 activation. Exp. Ther. Med. 2021, 22, 1046. [Google Scholar] [CrossRef]
- Khan, N.; Kuo, A.; Brockman, D.A.; Cooper, M.A.; Smith, M.T. Pharmacological inhibition of the NLRP3 inflammasome as a potential target for multiple sclerosis induced central neuropathic pain. Inflammopharmacology 2018, 26, 77–86. [Google Scholar] [CrossRef]
- Zhai, Y.; Meng, X.; Ye, T.; Xie, W.; Sun, G.; Sun, X. Inhibiting the NLRP3 Inflammasome Activation with MCC950 Ameliorates Diabetic Encephalopathy in db/db Mice. Molecules 2018, 23, 522. [Google Scholar] [CrossRef]
- Shao, S.; Xu, C.B.; Chen, C.J.; Shi, G.N.; Guo, Q.L.; Zhou, Y.; Wei, Y.Z.; Wu, L.; Shi, J.G.; Zhang, T.T. Divanillyl sulfone suppresses NLRP3 inflammasome activation via inducing mitophagy to ameliorate chronic neuropathic pain in mice. J. Neuroinflammation 2021, 18, 142. [Google Scholar] [CrossRef]
- Pol, O. CO and Pain Management. In Carbon Monoxide in Drug Discovery: Basics, Pharmacology, and Therapeutic Potential, 1st ed.; Chapter 29; Wang, B., Otterbein, L., Eds.; John Wiley & Sons, Inc.: Ney York, NY, USA, 2022; pp. 497–509. [Google Scholar]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of inflammation by the antioxidant haem oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; León, R.; López, M.G.; Oliva, B.; et al. Transcription Factor NRF2 as a Therapeutic Target for Chronic Diseases: A Systems Medicine Approach. Pharmacol. Rev. 2018, 70, 348–383. [Google Scholar] [CrossRef]
- Li, J.; Ma, J.; Lacagnina, M.J.; Lorca, S.; Odem, M.A.; Walters, E.T.; Kavelaars, A.; Grace, P.M. Oral dimethyl fumarate reduces peripheral neuropathic pain in rodents via NFE2L2 antioxidant signaling. Anesthesiology 2020, 132, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Thapliyal, S.; Kumar, A.; Paul, P.; Kumar, N.; Bisht, M.; Naithani, M.; Rao, S.; Handu, S.S. Dimethyl Fumarate Ameliorates Paclitaxel-Induced Neuropathic Pain in Rats. Cureus 2022, 14, e28818. [Google Scholar] [CrossRef]
- Shen, Y.; Zhang, Z.J.; Zhu, M.D.; Jiang, B.C.; Yang, T.; Gao, Y.J. Exogenous induction of HO-1 alleviates vincristine-induced neuropathic pain by reducing spinal glial activation in mice. Neurobiol. Dis. 2015, 79, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Riego, G.; Redondo, A.; Leánez, S.; Pol, O. Mechanism implicated in the anti-allodynic and anti-hyperalgesic effects induced by the activation of HEME oxygenase 1/carbon monoxide signaling pathway in the central nervous system of mice with neuropathic pain. Biochem. Pharmacol. 2018, 148, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Pol, O. The role of carbon monoxide, heme oxygenase 1, and the Nrf2 transcription factor in the modulation of chronic pain and their interactions with opioids and cannabinoids. Med. Res. Rev. 2021, 41, 136–155. [Google Scholar] [CrossRef] [PubMed]
- Cazuza, R.A.; Batallé, G.; Bai, X.; Leite-Panissi, C.R.A.; Pol, O. Effects of treatment with a carbon monoxide donor and an activator of heme oxygenase 1 on the nociceptive, apoptotic and/or oxidative alterations induced by persistent inflammatory pain in the central nervous system of mice. Brain Res. Bull. 2022, 188, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Casili, G.; Lanza, M.; Filippone, A.; Campolo, M.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. Dimethyl fumarate alleviates the nitroglycerin (NTG)-induced migraine in mice. J. Neuroinflammation 2020, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Liu, S.; Wang, Y.; Cui, R.; Zhang, X. The Link between Depression and Chronic Pain: Neural Mechanisms in the Brain. Neural Plast. 2017, 2017, 9724371. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; Vanella, L.; Platania, C.B.M.; Greish, K.; Bucolo, C.; Pittalà, V.; Salerno, L. Novel Heme Oxygenase-1 (HO-1) Inducers Based on Dimethyl Fumarate Structure. Int. J. Mol. Sci. 2020, 21, 9541. [Google Scholar] [CrossRef]
- Pittalà, V.; Vanella, L.; Maria Platania, C.B.; Salerno, L.; Raffaele, M.; Amata, E.; Marrazzo, A.; Floresta, G.; Romeo, G.; Greish, K.; et al. Synthesis, in vitro and in silico studies of HO-1 inducers and lung antifibrotic agents. Future Med. Chem. 2019, 11, 1523–1536. [Google Scholar] [CrossRef]
- Bennett, G.J.; Xie, Y.K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef] [PubMed]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.; Dubner, R.; Brown, F.; Flores, C.; Joris, J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain 1988, 32, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Porsolt, R.D.; Le Pichon, M.; Jalfre, M. Depression: A new animal model sensitive to antidepressant treatments. Nature 1977, 266, 730–732. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Franklin, K.B. Paxinos and Franklin’s, the Mouse Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 2008. [Google Scholar]
- Gao, S.J.; Li, D.Y.; Liu, D.Q.; Sun, J.; Zhang, L.Q.; Wu, J.Y.; Song, F.H.; Zhou, Y.Q.; Mei, W. Dimethyl Fumarate Attenuates Pain Behaviors in Osteoarthritis Rats via Induction of Nrf2-Mediated Mitochondrial Biogenesis. Mol. Pain 2022, 18, 17448069221124920. [Google Scholar] [CrossRef]
- Casili, G.; Lanza, M.; Filippone, A.; Cucinotta, L.; Paterniti, I.; Repici, A.; Capra, A.P.; Cuzzocrea, S.; Esposito, E.; Campolo, M. Dimethyl Fumarate (DMF) Alleviated Post-Operative (PO) Pain through the N-Methyl-d-Aspartate (NMDA) Receptors. Antioxidants 2022, 11, 1774. [Google Scholar] [CrossRef] [PubMed]
- Hervera, A.; Leánez, S.; Motterlini, R.; Pol, O. Treatment with carbon monoxide-releasing molecules and an HO-1 inducer enhances the effects and expression of µ-opioid receptors during neuropathic pain. Anesthesiology 2013, 118, 1180–1197. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, C.; Leánez, S.; Pol, O. The Inhibitory Effects of Cobalt Protoporphyrin IX and Cannabinoid 2 Receptor Agonists in Type 2 Diabetic Mice. Int. J. Mol. Sci. 2017, 18, 2268. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Khor, T.O.; Xu, C.; Shen, G.; Jeong, W.S.; Yu, S.; Kong, A.N. Activation of Nrf2-antioxidant signaling attenuates NFkappaB-inflammatory response and elicits apoptosis. Biochem. Pharmacol. 2008, 76, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wang, Y.; Liu, M.; Chen, H. Hydrogen sulfide attenuates renal I/R-induced activation of the inflammatory response and apoptosis via regulating Nrf2-mediated NLRP3 signaling pathway inhibition. Mol. Med. Rep. 2021, 24, 518. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Smith, M.T. The NLRP3 inflammasome: Role in the pathobiology of chronic pain. Inflammopharmacology 2023, 31, 1589–1603. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, W.; Zhang, X.; Lu, P.; Du, Q.; Tao, L.; Ding, Y.; Wang, Y.; Hu, R. Dimethyl fumarate ameliorates dextran sulfate sodium-induced murine experimental colitis by activating Nrf2 and suppressing NLRP3 inflammasome activation. Biochem. Pharmacol. 2016, 112, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Shang, H.; Chen, Y.Q.; Pan, L.L.; Bhatia, M.; Sun, J. Sulforaphane protects pancreatic acinar cell injury by modulating Nrf2-Mediated oxidative stress and NLRP3 inflammatory pathway. Oxid. Med. Cell Longev. 2016, 2016, 7864150. [Google Scholar] [CrossRef]
- Raison, V. Neurobiology of depression, fibromyalgia and neuropathic pain. Front Biosci. 2009, 14, 5291–5338. [Google Scholar]
- Humo, M.; Lu, H.; Yalcin, I. The molecular neurobiology of chronic pain-induced depression. Cell Tissue Res. 2019, 377, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Ferreira-Chamorro, P.; Redondo, A.; Riego, G.; Leánez, S.; Pol, O. Sulforaphane Inhibited the Nociceptive Responses, Anxiety- and Depressive-Like Behaviors Associated with Neuropathic Pain and Improved the Anti-allodynic Effects of Morphine in Mice. Front. Pharmacol. 2018, 9, 1332. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. Essential Role of Keap1-Nrf2 Signaling in Mood Disorders: Overview and Future Perspective. Front. Pharmacol. 2018, 9, 1182. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Zhang, J.C.; Ishima, T.; Ren, Q.; Yang, C.; Dong, C.; Ma, M.; Saito, A.; Honda, T.; Hashimoto, K. Antidepressant effects of TBE-31 and MCE-1, the novel Nrf2 activators, in an inflammation model of depression. Eur. J. Pharmacol. 2016, 793, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ullah, R.; Wang, J.; Du, Y.; Huang, S.; Meng, L.; Gao, Y.; Gong, M.; Galaj, E.; Yin, X.; et al. Exogenous Carbon Monoxide Produces Rapid Antidepressant- and Anxiolytic-Like Effects. Front. Pharmacol. 2021, 12, 757417. [Google Scholar] [CrossRef]
- Zuo, C.; Cao, H.; Song, Y.; Gu, Z.; Huang, Y.; Yang, Y.; Miao, J.; Zhu, L.; Chen, J.; Jiang, Y.; et al. Nrf2: An all-rounder in depression. Redox Biol. 2022, 58, 102522. [Google Scholar] [CrossRef]
- Martín-Hernández, D.; Caso, J.R.; Javier Meana, J.; Callado, L.F.; Madrigal, J.L.M.; García-Bueno, B.; Leza, J.C. Intracellular inflammatory and antioxidant pathways in postmortem frontal cortex of subjects with major depression: Effect of antidepressants. J. Neuroinflamm. 2018, 15, 251. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Batallé, G.; Pol, O. The Anxiolytic and Antidepressant Effects of Diallyl Disulfide and GYY4137 in Animals with Chronic Neuropathic Pain. Antioxidants 2021, 10, 1074. [Google Scholar] [CrossRef] [PubMed]
- Kremer, M.; Becker, L.J.; Barrot, M.; Yalcin, I. How to study anxiety and depression in rodent models of chronic pain? Eur. J. Neurosci. 2021, 53, 236–270. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, L.; Silva, R.; Pinto-Ribeiro, F.; Pêgo, J.M.; Bessa, J.M.; Pertovaara, A.; Sousa, N.; Almeida, A. Neuropathic pain is associated with depressive behaviour and induces neuroplasticity in the amygdala of the rat. Exp. Neurol. 2008, 213, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Tastan, B.; Arioz, B.I.; Genc, S. Targeting NLRP3 Inflammasome With Nrf2 Inducers in Central Nervous System Disorders. Front. Immunol. 2022, 13, 865772. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Ji, R.R. Chemokines, neuronal-glial interactions, and central processing of neuropathic pain. Pharmacol. Ther. 2010, 126, 56–68. [Google Scholar] [CrossRef]
- Fiore, N.T.; Austin, P.J. Are the emergence of affective disturbances in neuropathic pain states contingent on supraspinal neuroinflammation? Brain Behav. Immun. 2016, 56, 397–411. [Google Scholar] [CrossRef]
- Barcelon, E.E.; Cho, W.H.; Jun, S.B.; Lee, S.J. Brain Microglial Activation in Chronic Pain-Associated Affective Disorder. Front. Neurosci. 2019, 13, 213. [Google Scholar] [CrossRef] [PubMed]
- Yirmiya, R.; Rimmerman, N.; Reshef, R. Depression as a microglial disease. Trends Neurosci. 2015, 38, 637–658. [Google Scholar] [CrossRef]
- Bortolato, B.; Carvalho, A.F.; Soczynska, J.K.; Perini, G.I.; McIntyre, R.S. The Involvement of TNF-α in Cognitive Dysfunction Associated with Major Depressive Disorder: An Opportunity for Domain Specific Treatments. Curr. Neuropharmacol. 2015, 13, 558–576. [Google Scholar] [CrossRef]
- McGeough, M.D.; Wree, A.; Inzaugarat, M.E.; Haimovich, A.; Johnson, C.D.; Peña, C.A.; Goldbach-Mansky, R.; Broderick, L.; Feldstein, A.E.; Hoffman, H.M. TNF regulates transcription of NLRP3 inflammasome components and inflammatory molecules in cryopyrinopathies. J. Clin. Investig. 2017, 127, 4488–4497. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.G.; Lopes, I.S.; Filho, A.J.M.C.; Cavalcante, T.M.B.; Oliveira, J.V.S.; de Carvalho, M.A.J.; de Lima, K.A.; Jucá, P.M.; Mendonça, S.S.; Mottin, M.; et al. Neuroprotective effects of dimethyl fumarate against depression-like behaviors via astrocytes and microglia modulation in mice: Possible involvement of the HCAR2/Nrf2 signaling pathway. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2022, 395, 1029–1045. [Google Scholar] [CrossRef]
- Micó, J.A.; Prieto, R. Elucidating the mechanism of action of pregabalin: α2δ as a therapeutic target in anxiety. CNS Drugs. 2012, 26, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Grégoire, S.; Michaud, V.; Chapuy, E.; Eschalier, A.; Ardid, D. Study of emotional and cognitive impairments in mononeuropathic rats: Effect of duloxetine and gabapentin. Pain 2012, 153, 1657–1663. [Google Scholar] [CrossRef]
- La Porta, C.; Lara-Mayorga, I.M.; Negrete, R.; Maldonado, R. Effects of pregabalin on the nociceptive, emotional and cognitive manifestations of neuropathic pain in mice. Eur. J. Pain 2016, 20, 1454–1466. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Doods, H.; Treede, R.D.; Ceci, A. Duloxetine and 8-OH-DPAT, but not fluoxetine, reduce depression-like behaviour in an animal model of chronic neuropathic pain. Neurosci. Lett. 2016, 619, 162–167. [Google Scholar] [CrossRef]
- Kappos, L.; Gold, R.; Miller, D.H.; Macmanus, D.G.; Havrdova, E.; Limmroth, V.; Polman, C.H.; Schmierer, K.; Yousry, T.A.; Yang, M.; et al. Efficacy and safety of oral fumarate in patients with relapsing-remitting multiple sclerosis: A multicentre, randomised, double-blind, placebo-controlled phase IIb study. Lancet 2008, 372, 1463–1472. [Google Scholar] [CrossRef]






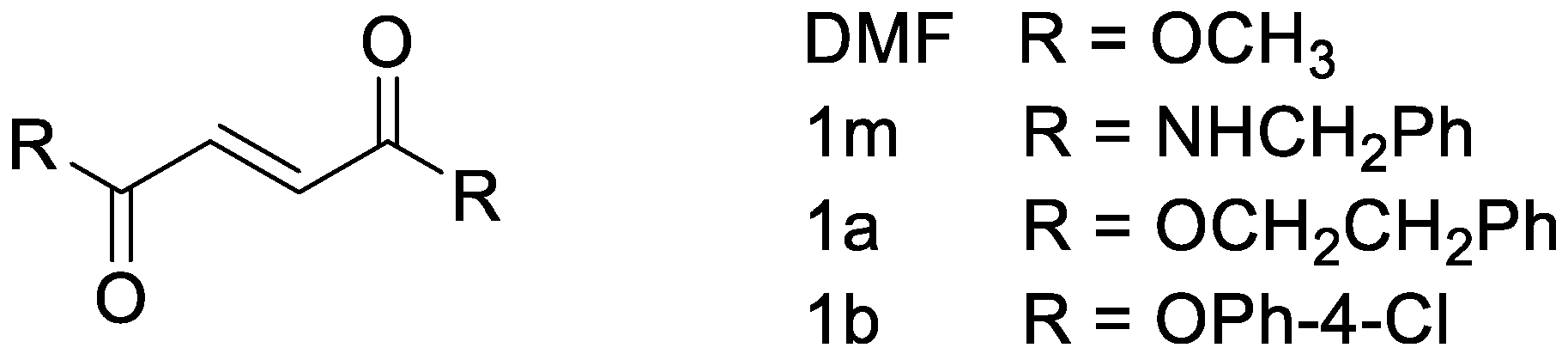{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days of Treatment | |||||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |
| Mechanical | F9,50 = 46.38 | F9,50 = 59.07 | F9,50 = 47.26 | F9,50 = 28.31 | F9,50 = 36.74 |
| Allodynia | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Thermal | F9,50 = 49.80 | F9,50 = 49.10 | F9,50 = 37.35 | F9,50 = 30.71 | F9,50 = 26.40 |
| Hyperalgesia | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Cold | F9,50 = 52.36 | F9,50 = 46.45 | F9,50 = 23.81 | F9,50 = 37.03 | F9,50 = 45.64 |
| Allodynia | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez-Rojas, I.; Pérez-Fernández, M.; Bai, X.; Martínez-Martel, I.; Intagliata, S.; Pittalà, V.; Salerno, L.; Pol, O. The Inhibition of Neuropathic Pain Incited by Nerve Injury and Accompanying Mood Disorders by New Heme Oxygenase-1 Inducers: Mechanisms Implicated. Antioxidants 2023, 12, 1859. https://doi.org/10.3390/antiox12101859
Suárez-Rojas I, Pérez-Fernández M, Bai X, Martínez-Martel I, Intagliata S, Pittalà V, Salerno L, Pol O. The Inhibition of Neuropathic Pain Incited by Nerve Injury and Accompanying Mood Disorders by New Heme Oxygenase-1 Inducers: Mechanisms Implicated. Antioxidants. 2023; 12(10):1859. https://doi.org/10.3390/antiox12101859
Chicago/Turabian StyleSuárez-Rojas, Irene, Montse Pérez-Fernández, Xue Bai, Ignacio Martínez-Martel, Sebastiano Intagliata, Valeria Pittalà, Loredana Salerno, and Olga Pol. 2023. "The Inhibition of Neuropathic Pain Incited by Nerve Injury and Accompanying Mood Disorders by New Heme Oxygenase-1 Inducers: Mechanisms Implicated" Antioxidants 12, no. 10: 1859. https://doi.org/10.3390/antiox12101859