
Saponins of Tomato Extract Improve Non-Alcoholic Fatty Liver Disease by Regulating Oxidative Stress and Lipid Homeostasis
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of STE
2.3. Determination of Total Saponins of Tomato Content
2.4. Determination of Esculeoside A Content
2.5. Animal Experimental Design
2.6. Determination of Biochemical Indicators
2.7. Oil Red Staining and HE Staining
2.8. Determination of Hepatic TG, TC, and Total Fat
2.9. Extraction of Liver Total Proteins and Nucleoproteins
2.10. Western Blot Analysis
2.11. Data Statistics
3. Results and Discussion
3.1. The Content of Total Saponins and Esculeoside A in STE
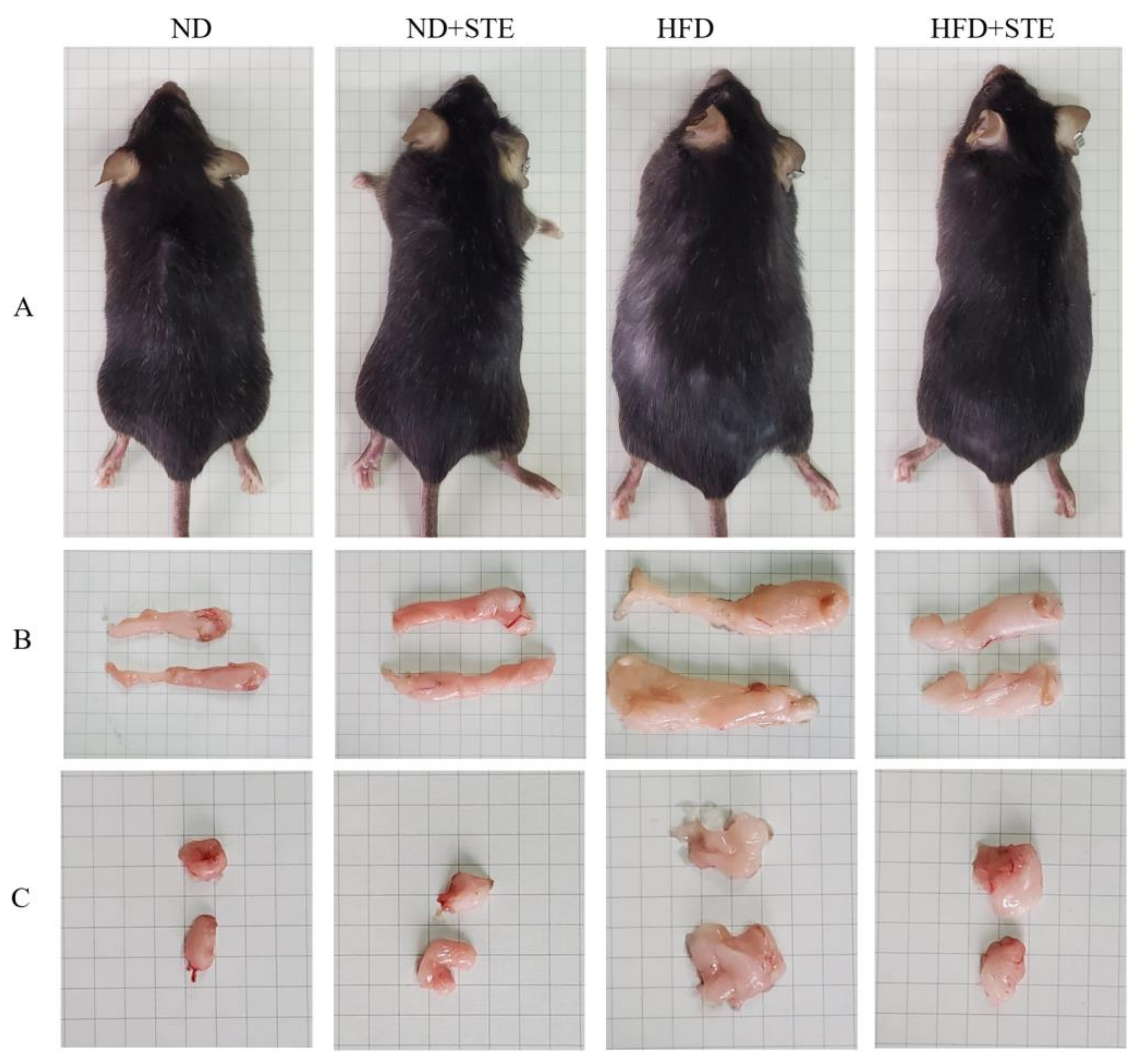
3.2. Effect of STE on Body Weight, Liver Weight, Kidney Weight, and Visceral Fat Weight of Mice
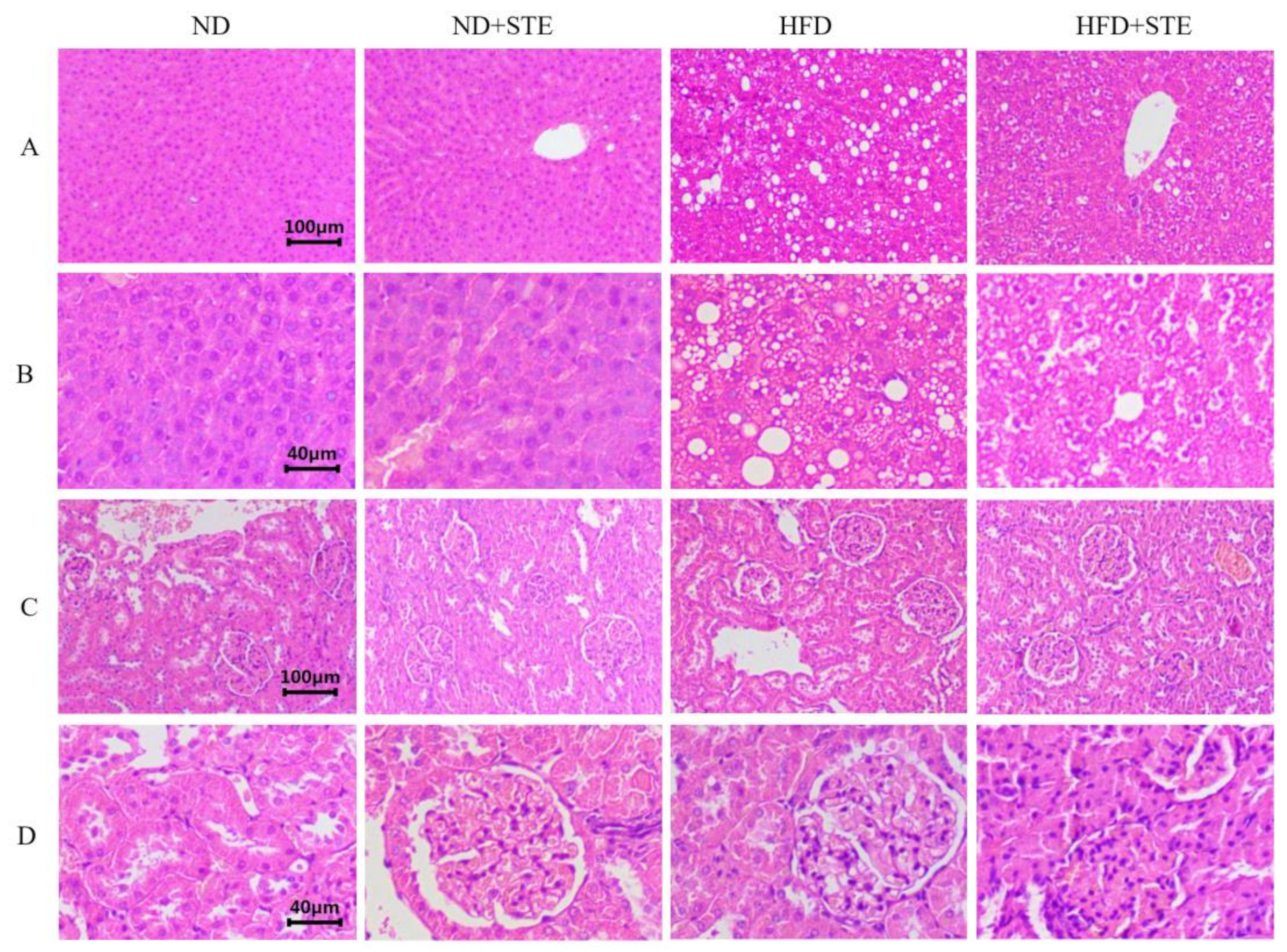
3.3. Effect of STE on Liver and Kidney Functions in Mice
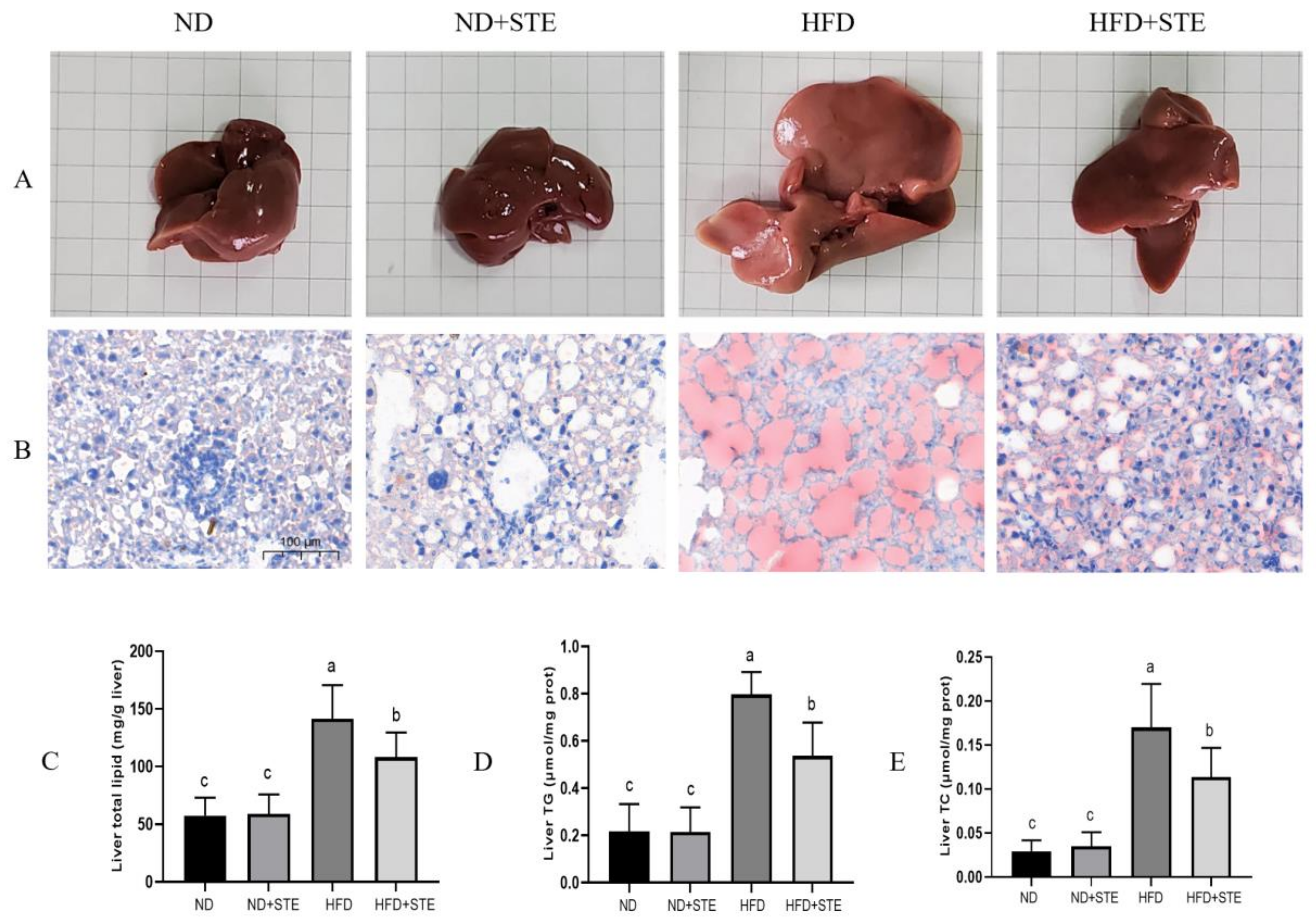
3.4. Effect of STE on Liver Fat in Mice
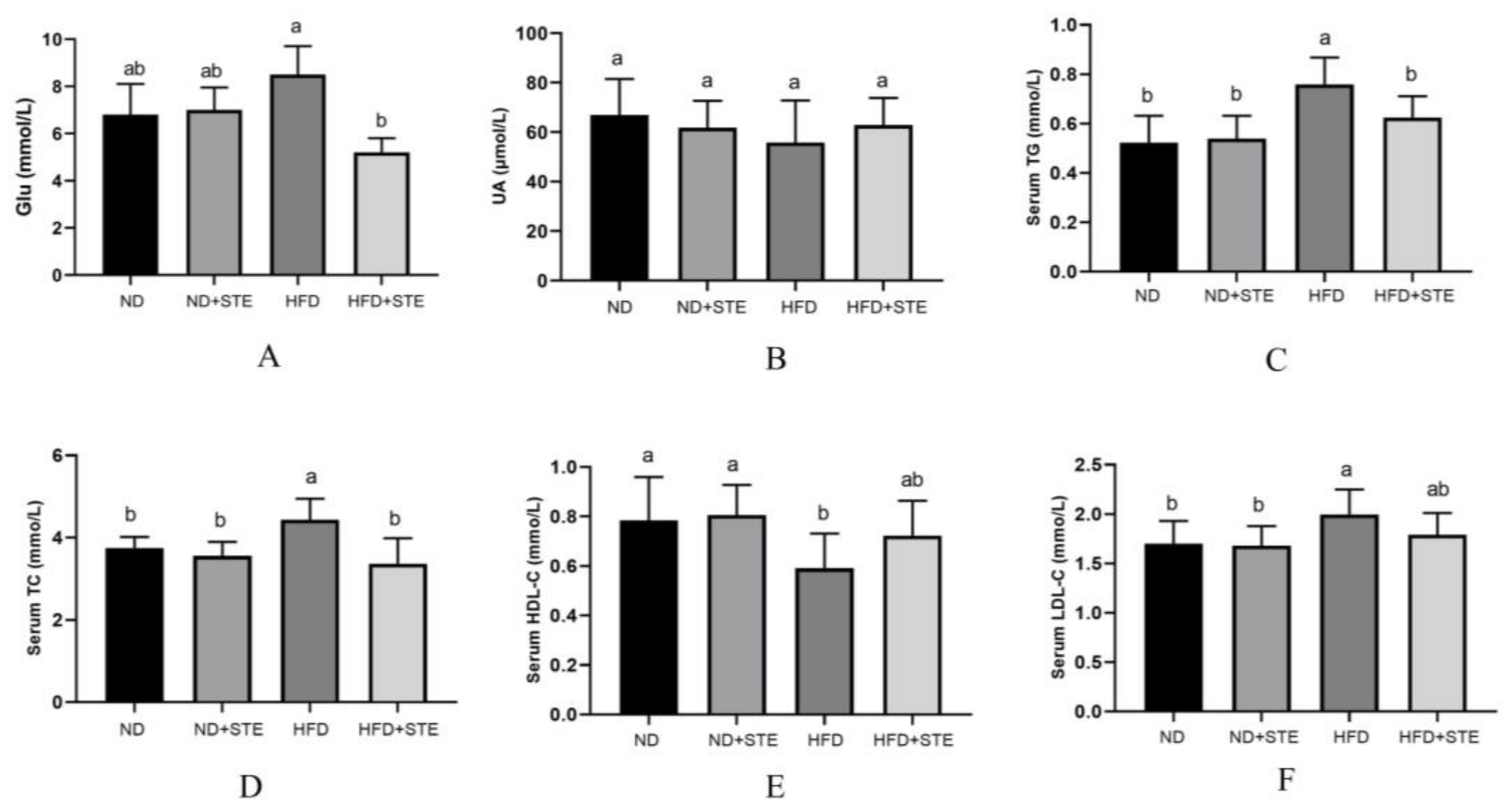
3.5. Effects of STE on Blood Glucose, UA, and Lipids in Mice
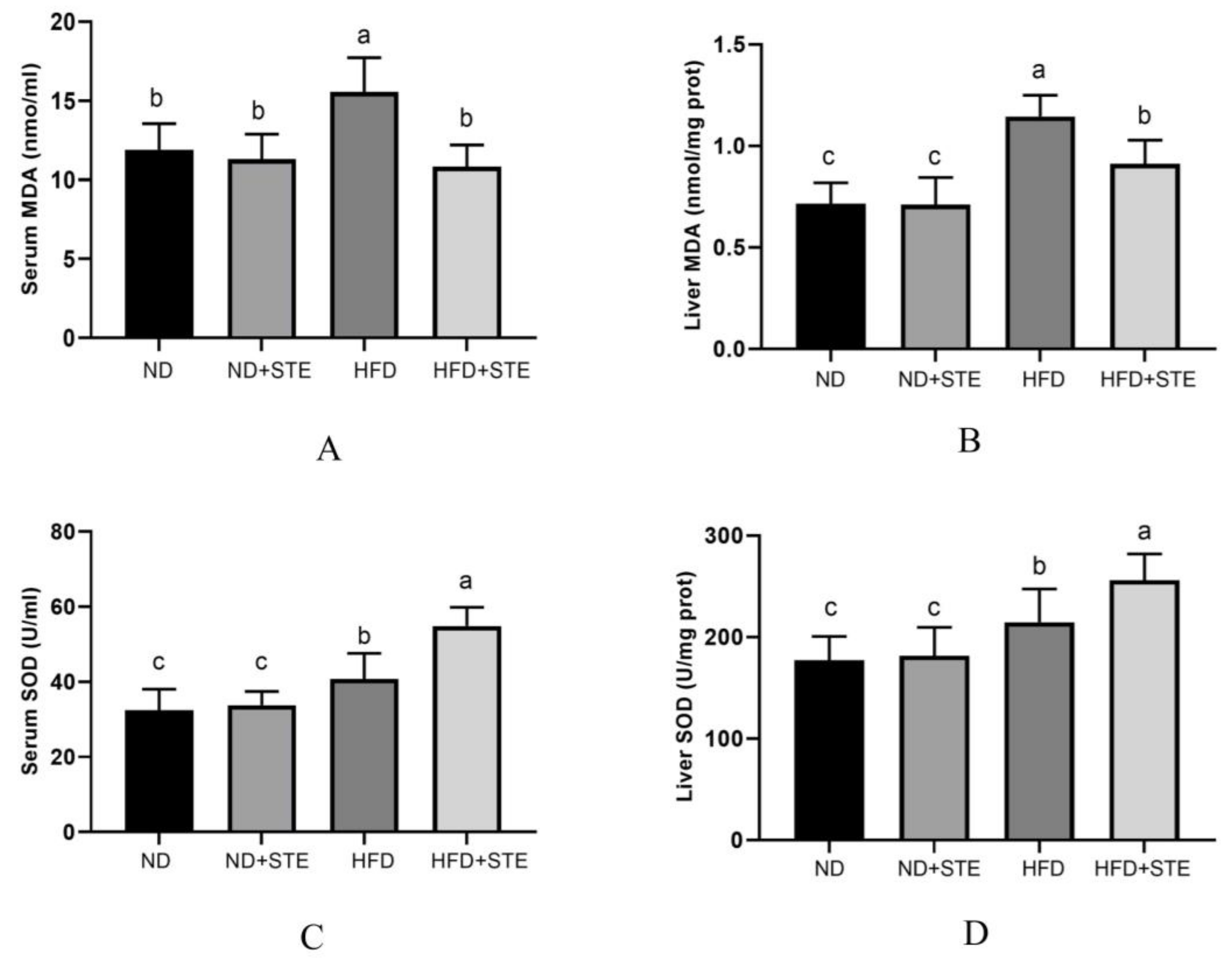
3.6. Effects of STE on SOD Activity and MDA Content in Livers and Blood of Mice
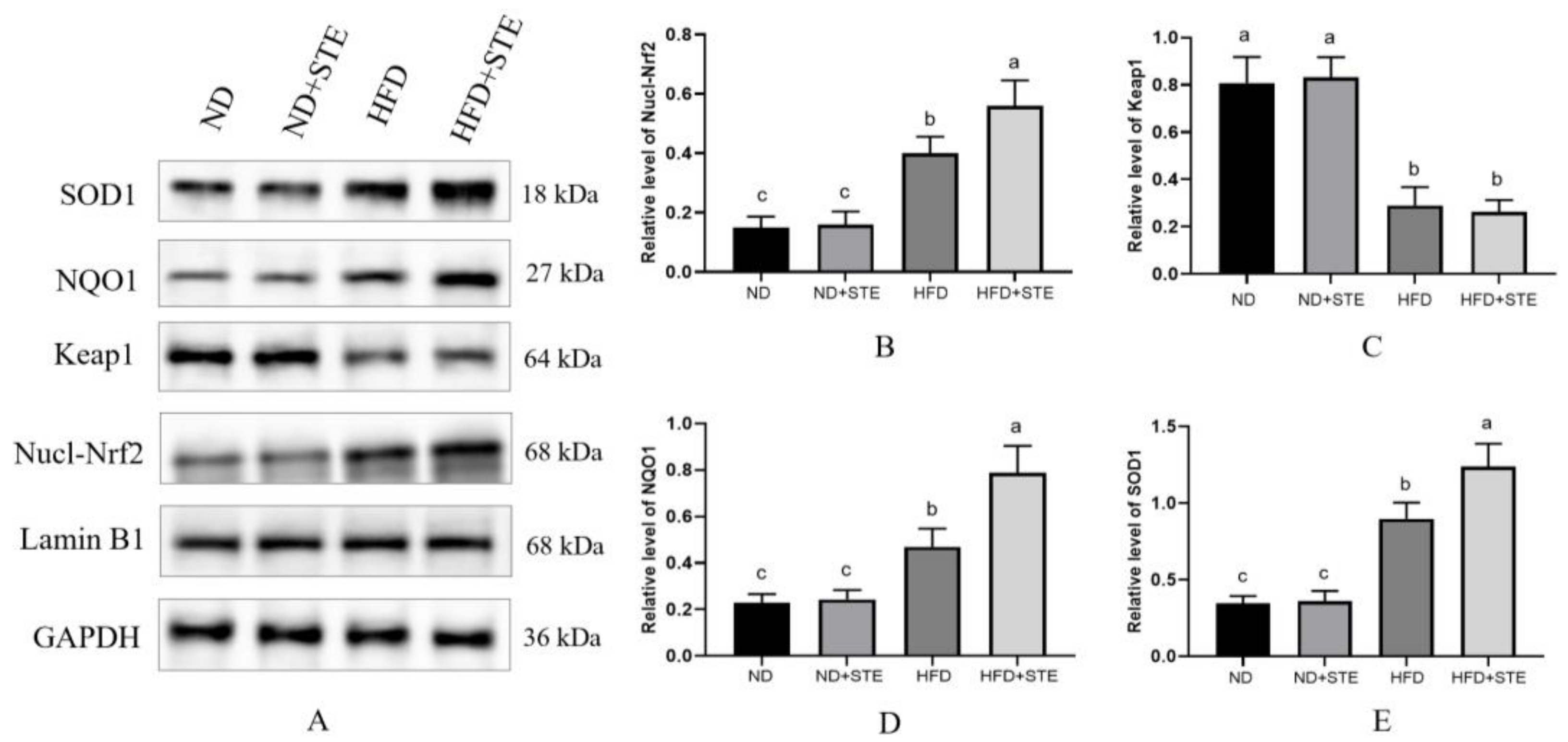
3.7. Effect of STE on the Nrf2 Pathway
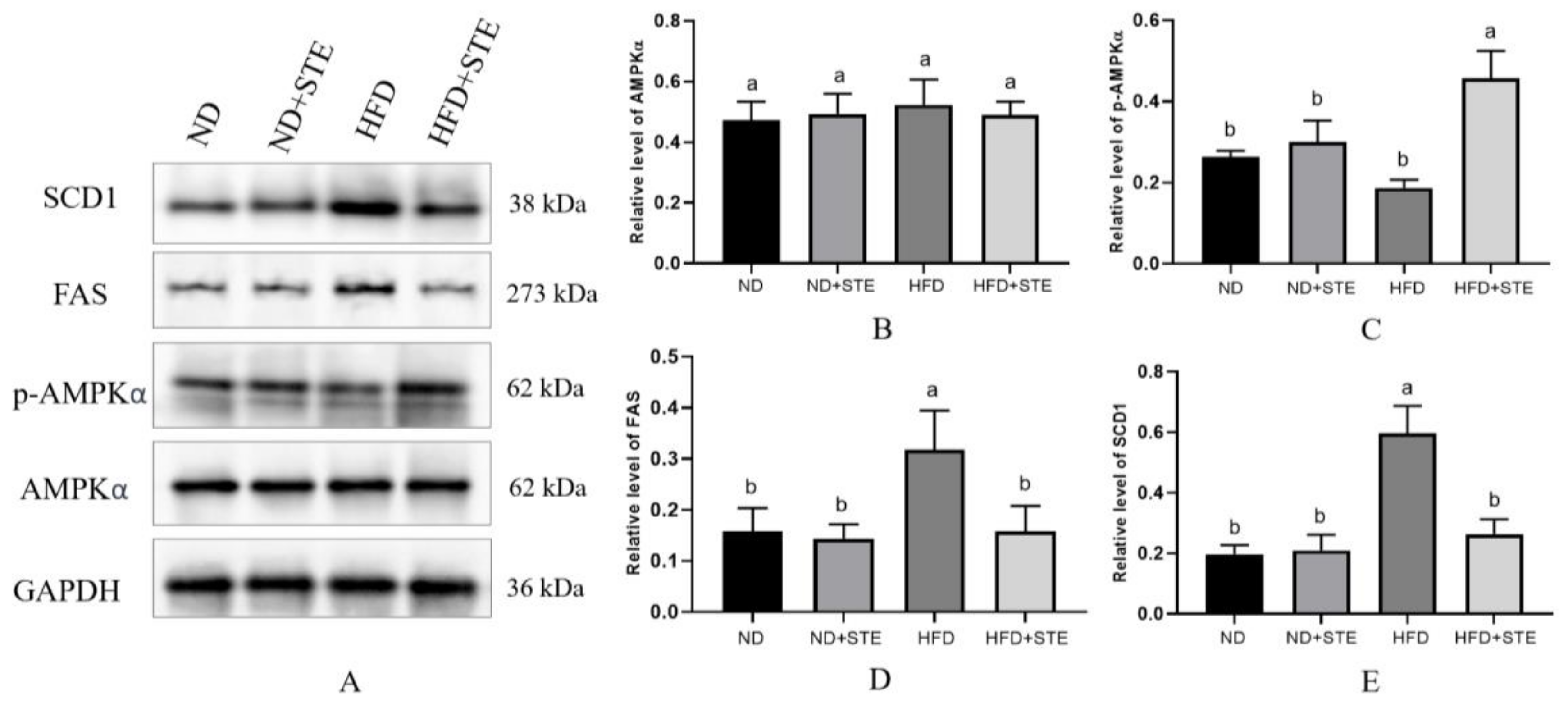
3.8. Effect of STE on AMPK Pathway
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ng, C.H.; Huang, D.Q.; Nguyen, M.H. Nonalcoholic fatty liver disease versus metabolic-associated fatty liver disease: Prevalence, outcomes and implications of a change in name. Clin. Exp. Hepatol. 2022, 28, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Finan, B.; Parlee, S.D.; Yang, B. Nuclear hormone and peptide hormone therapeutics for NAFLD and NASH. Mol. Metab. 2021, 46, 101153. [Google Scholar] [CrossRef] [PubMed]
- Marinello, P.C.; Cella, P.S.; Testa, M.T.J.; Guirro, P.B.; Brito, W.A.D.S.; Padilha, C.S.; Cecchini, A.L.; Silva, R.P.; Duarte, J.A.R.; Deminice, R. Creatine supplementation protects against diet-induced non-alcoholic fatty liver but exacerbates alcoholic fatty liver. Life Sci. 2022, 310, 121064. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.; George, J. Nonalcoholic Fatty Liver Disease Management: Dietary and Lifestyle Modifications. Semin. Liver Dis. 2015, 35, 318–337. [Google Scholar] [CrossRef]
- Świderska, M.; Maciejczyk, M.; Zalewska, A.; Pogorzelska, J.; Flisiak, R.; Chabowski, A. Oxidative stress biomarkers in the serum and plasma of patients with non-alcoholic fatty liver disease (NAFLD). Can plasma AGE be a marker of NAFLD? Oxidative stress biomarkers in NAFLD patients. Free Radic. Res. 2019, 53, 3841–3850. [Google Scholar] [CrossRef] [PubMed]
- Preiser, J. Oxidative stress. JPEN J. Parenter. Enter. Nutr. 2012, 36, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Day, C.P.; James, O.F. Steatohepatitis: A tale of two “hits”? Gastroenterology 1998, 114, 842–845. [Google Scholar] [CrossRef]
- Ma, Y.; Lee, G.; Heo, S.-Y.; Roh, Y.-S. Oxidative Stress Is a Key Modulator in the Development of Nonalcoholic Fatty Liver Disease. Antioxidants 2021, 11, 91. [Google Scholar] [CrossRef]
- Cintolesi, A.; Rodríguez-Moyá, M.; Gonzalez, R. Fatty acid oxidation: Systems analysis and applications. Wiley Interdiscip. Rev. Syst. Biol. Med. 2013, 5, 575–585. [Google Scholar] [CrossRef]
- Hassanein, E.H.M.; Sayed, A.M.; Hussein, O.E.; Mahmoud, A.M. Coumarins as Modulators of the Keap1/Nrf2/ARE Signaling Pathway. Oxid. Med. Cell. Longev. 2020, 2020, 1675957. [Google Scholar] [CrossRef]
- Zakaria, Z.; Othman, Z.A.; Suleiman, J.B.; Mustaffa, K.M.F.; Jalil, N.A.C.; Ghazali, W.S.W.; Zulkipli, N.N.; Mohamed, M.; Kamaruzaman, K.A. Therapeutic Effects of Heterotrigona itama (Stingless Bee) Bee Bread in Improving Hepatic Lipid Metabolism through the Activation of the Keap1/Nrf2 Signaling Pathway in an Obese Rat Model. Antioxidants 2022, 11, 2190. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Yu, W.; Li, X.; Zhou, F.; Zhang, W.; Shen, Q.; Li, J.; Zhang, C.; Shen, P. Apigenin, a modulator of PPARγ, attenuates HFD-induced NAFLD by regulating hepatocyte lipid metabolism and oxidative stress via Nrf2 activation. Biochem. Pharmacol. 2017, 136, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ji, R.; Sun, H.; Peng, J.; Ma, X.; Wang, C.; Fu, Y.; Bao, L.; Jin, Y. Scutellarin ameliorates nonalcoholic fatty liver disease through the PPARγ/PGC-1α-Nrf2 pathway. Free Radic. Res. 2018, 52, 198–211. [Google Scholar] [CrossRef] [PubMed]
- Kosmalski, M.; Frankowski, R.; Ziółkowska, S.; Różycka-Kosmalska, M.; Pietras, T. What’s New in the Treatment of Non-Alcoholic Fatty Liver Disease (NAFLD). J. Clin. Med. 2023, 12, 1852. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.-L.; Cheng, S.-C.; Yan, X.-J.; Li, D.-P. Determination Result of Esculeoside A in Different Lycopersicon esculentum MILL Samples. Food Res. Dev. 2014, 35, 31–34. [Google Scholar]
- Fujiwara, Y.; Kiyota, N.; Hori, M.; Matsushita, S.; Iijima, Y.; Aoki, K.; Shibata, D.; Takeya, M.; Ikeda, T.; Nohara, T.; et al. Esculeogenin A, a New Tomato Sapogenol, Ameliorates Hyperlipidemia and Atherosclerosis in ApoE-Deficient Mice by Inhibiting ACAT. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2400–2406. [Google Scholar] [CrossRef]
- Zhang, L.; Li, D.; Yang, Z.; Liu, J.; Huang, Y.; Chen, Y. Effects of esculeoside A from Lycopersicon esculentum on serum lipid and liver lipid in mice. Guihaia 2018, 38, 583–589. [Google Scholar]
- Wu, J.-Z.; Li, C.-Y.; Liu, J.-L.; Lu, F.-L.; Chen, Y.-Y.; Li, D.-P. Effects of Aqueous Extract of Lycopersicon Esculentum on Atherosclerosis in Rats. Food Res. Dev. 2011, 35, 31–34. [Google Scholar]
- Yang, Z.; Li, W.; Li, D.; Chan, A.S.C. Evaluation of Nutritional Compositions, Bioactive Components, and Antioxidant Activity of Three Cherry Tomato Varieties. Agronomy 2023, 13, 637. [Google Scholar] [CrossRef]
- Chen, C.; Li, S.-X.; Wang, S.-M.; Liang, S.-W. Investigation into the anti-thrombosis effect and contents of total saponins and flavonoids in the bioactive fraction of Naodesheng prescription. J. Ethnopharmacol. 2012, 144, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Liu, J.; Chen, Y.; Wu, J.; Li, D. Determination of esculeoside A in Lycopersicon esculentum MILL by high performance liquid chromatography with l evaporative light scattering detection. Chin. J. Chromatogr. 2010, 28, 908–911. [Google Scholar]
- Tung, Y.-C.; Shih, Y.-A.; Nagabhushanam, K.; Ho, C.-T.; Cheng, A.-C.; Pan, M.-H. Coleus forskohlii and Garcinia indica extracts attenuated lipid accumulation by regulating energy metabolism and modulating gut microbiota in obese mice. Food Res. Int. 2021, 142, 110143. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual hematoxylin and eosin staining of mouse tissue sections. Cold Spring Harb. Perspect. Biol. 2014, 2014, 655–658. [Google Scholar] [CrossRef]
- Tong, L.-T.; Zhong, K.; Liu, L.; Guo, L.; Cao, L.; Zhou, S. Oat oil lowers the plasma and liver cholesterol concentrations by promoting the excretion of faecal lipids in hypercholesterolemic rats. Food Chem. 2014, 142, 129–134. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, J.; Guo, Y.; Ye, H.; Yu, C.; Xu, C.; Xu, L.; Wu, S.; Sun, W.; Wei, H.; et al. Functional proteomic analysis of nonalcoholic fatty liver disease in rat models: Enoyl-coenzyme a hydratase down-regulation exacerbates hepatic steatosis. Hepatology 2010, 51, 1190–1199. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1956, 226, 497–509. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, L.; Liu, J.; Wang, L.; Chen, Y.; Li, D. Hypoglycemic effects of esculeoside A are mediated via activation of AMPK and upregulation of IRS-1. BMC Complement. Altern. Med. 2019, 19, 136. [Google Scholar] [CrossRef] [PubMed]
- Haberka, M.; Stolarz-Skrzypek, K.; Biedroń, M.; Szóstak-Janiak, K.; Partyka, M.; Olszanecka-Glinianowicz, M.; Gąsior, Z. Obesity, Visceral Fat, and Hypertension-Related Complications. Metab. Syndr. Relat. Disord. 2018, 16, 521–529. [Google Scholar] [CrossRef]
- Katzmarzyk, P.T.; Brown, J.C.; Yang, S.; Mire, E.F.; Wu, X.-C.; Miele, L.; Ochoa, A.C.; Zabaleta, J. Association of Abdominal Visceral Adiposity and Total Fat Mass with Cancer Incidence and Mortality in White and Black Adults. Cancer Epidemiol. Biomarkers Prev. 2022, 31, 1532–1538. [Google Scholar] [CrossRef]
- Beek, J.H.D.A.V.; Moor, M.H.M.D.; Geus, E.J.C.D.; Lubke, G.H.; Vink, J.M.; Willemsen, G.; Boomsma, D.I. The genetic architecture of liver enzyme levels: GGT, ALT and AST. Behav. Genet. 2013, 43, 329–339. [Google Scholar] [CrossRef]
- Yang, C.-H.; Kuo, W.-H.; Wang, J.-S.; Hsiang, Y.-P.; Lin, Y.-M.; Wang, Y.-T.; Tsai, F.-H.; Lee, C.-T.; Chou, J.-H.; Chang, H.-Y.; et al. Improvement in the Blood Urea Nitrogen and Serum Creatinine Using New Cultivation of Cordyceps militaris. Evid. Based Complement. Altern. Med. 2022, 2022, 4321298. [Google Scholar] [CrossRef] [PubMed]
- Okuda, M.; Asakura, K.; Sasaki, S. Protein Intake Estimated from Brief-Type Self-Administered Diet History Questionnaire and Urinary Urea Nitrogen Level in Adolescents. Nutrients 2019, 11, 319. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Q.-Y.; Zeng, L.-T.; Wang, J.-J.; Liu, Z.; Fan, G.-Q.; Li, J.; Cai, J.-P. Long-Term High-Fat High-Fructose Diet Induces Type 2 Diabetes in Rats through Oxidative Stress. Nutrients 2022, 14, 2181. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.; Fang, Y.; Chen, S.; Zhu, X.; Shan, C.; Su, J.; Yu, J.; Li, B.; Yang, Y.; Chen, B.; et al. Gypenosides Inhibits Xanthine Oxidoreductase and Ameliorates Urate Excretion in Hyperuricemic Rats Induced by High Cholesterol and High Fat Food (Lipid Emulsion). Med. Sci. Monit. 2017, 23, 1129–1140. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Tzeng, T.-F.; Liou, S.-S.; Chang, Y.-S.; Liu, I.-M. Regulation of lipid disorders by ethanol extracts from Zingiber zerumbet in high-fat diet-induced rats. Food Chem. 2012, 132, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Borgstahl, G.E.O.; Oberley-Deegan, R.E. Superoxide Dismutases (SODs) and SOD Mimetics. Antioxidants 2018, 7, 156. [Google Scholar] [CrossRef]
- Weismann, D.; Hartvigsen, K.; Lauer, N.; Bennett, K.L.; Scholl, H.P.N.; Issa, P.C.; Cano, M.; Brandstätter, H.; Tsimikas, S.; Skerka, C.; et al. Complement factor H binds malondialdehyde epitopes and protects from oxidative stress. Nature 2011, 478, 76–81. [Google Scholar] [CrossRef]
- Young, D.; Fan, M.Z.; Mine, Y. Egg yolk peptides up-regulate glutathione synthesis and antioxidant enzyme activities in a porcine model of intestinal oxidative stress. J. Agric. Food Chem. 2010, 58, 7624–7633. [Google Scholar] [CrossRef]
- Tkachev, V.O.; Menshchikova, E.B.; Zenkov, N.K. Mechanism of the Nrf2/Keap1/ARE signaling system. Biochemistry 2011, 76, 407–422. [Google Scholar] [CrossRef]
- Kabaria, S.; Choi, D.C.; Chaudhuri, A.D.; Jain, M.R.; Li, H.; Junn, E. MicroRNA-7 activates Nrf2 pathway by targeting Keap1 expression. Free Radic. Biol. Med. 2015, 89, 548–556. [Google Scholar] [CrossRef]
- Li, H.; Wu, M.; Guo, C.; Zhai, R.; Chen, J. Tanshinone IIA Regulates Keap1/Nrf2 Signal Pathway by Activating Sestrin2 to Restrain Pulmonary Fibrosis. Am. J. Chin. Med. 2022, 50, 2125–2151. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huangfu, M.; Li, X.; Han, M.; Liu, G.; Yu, D.; Zhou, L.; Dou, T.; Liu, Y.; Guan, X.; et al. Osthole Induces Apoptosis and Caspase-3/GSDME-Dependent Pyroptosis via NQO1-Mediated ROS Generation in HeLa Cells. Oxid. Med. Cell. Longev. 2022, 2022, 8585598. [Google Scholar] [CrossRef]
- Poovitha, S.; Parani, M. Protein extract from the fruit pulp of, Momordica charantia, potentiate glucose uptake by up-regulating GLUT4 and AMPK. J. Funct. Foods 2017, 37, 507–512. [Google Scholar] [CrossRef]
- Xu, N.; Luo, H.; Li, M.; Wu, J.; Wu, X.; Chen, L.; Gan, Y.; Guan, F.; Li, M.; Su, Z.; et al. β-patchoulene improves lipid metabolism to alleviate non-alcoholic fatty liver disease via activating AMPK signaling pathway. Biomed. Pharmacother. 2021, 134, 111104. [Google Scholar] [CrossRef]
- Joo, M.S.; Kim, W.D.; Lee, K.Y.; Kim, J.H.; Koo, J.H.; Kim, S.G. AMPK Facilitates Nuclear Accumulation of Nrf2 by Phosphorylating at Serine 550. Mol. Cell. Biol. 2016, 36, 1931–1942. [Google Scholar] [CrossRef]
- Ceylan, C.; Yahşi, S.; Doğan, S.; ÖztÜrk, E.; Ceylan, G. Emphasis of FAS/FASL gene polymorphism in patients with non-muscle invasive bladder cancer. Ir. J. Med. Sci. 2018, 187, 1115–1119. [Google Scholar] [CrossRef]
- Iida, T.; Ubukata, M.; Mitani, I.; Nakagawa, Y.; Maeda, K.; Imai, H.; Ogoshi, Y.; Hotta, T.; Sakata, S.; Sano, R.; et al. Discovery of potent liver-selective stearoyl-CoA desaturase-1 (SCD1) inhibitors, thiazole-4-acetic acid derivatives, for the treatment of diabetes, hepatic steatosis, and obesity. Eur. J. Med. Chem. 2018, 158, 832–852. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | XT304 Normal Control Diet | XT301 High-Fat Diet |
|---|---|---|
| Ingredient | amounts (g) | amounts (g) |
| Casein | 200.00 | 200.00 |
| L-Cystine | 3.00 | 3.00 |
| Corn starch | 504.00 | - |
| Fructose | - | 200.00 |
| Maltodextrin | 100.00 | 100.00 |
| Sucrose | 100.00 | 100.00 |
| Cellulose | 50.00 | 50.00 |
| Soybean oil | 25.00 | 25.00 |
| Lard | 20.00 | 20.00 |
| Primex Shortening | - | 135.00 |
| Mineral mix S10026B | 50.00 | 50.00 |
| Vitamin mix V10001C | 1.00 | 1.00 |
| Choline bitartrate | 2.00 | 2.00 |
| Cholesterol | - | 18.00 |
| Formula | proportion (%) | proportion (%) |
| Protein | 19.24 | 22.45 |
| Carbohydrate | 66.82 | 44.36 |
| Fat | 4.27 | 19.91 |
| Energy | proportion (%) | proportion (%) |
| Protein | 20.11 | 20.12 |
| Carbohydrate | 69.85 | 39.74 |
| Fat | 10.04 | 40.14 |
| kal/g | kal/g | |
| Diet | 3.83 | 4.46 |
| Item | STE |
|---|---|
| Total saponins of tomato (g/100 g) | 43.12 ± 1.31 |
| Esculeoside A (g/100 g) | 15.63 ± 0.42 |
| Item | ND | ND + STE | HFD | HFD + STE |
|---|---|---|---|---|
| Initial weight (g) | 30.32 ± 0.89 b | 30.33 ± 0.91 b | 37.63 ± 0.92 a | 37.61 ± 1.05 a |
| Final weight (g) | 34.74 ± 1.13 c | 33.92 ± 0.92 c | 45.41 ± 3.21 a | 39.53 ± 2.62 b |
| Weight gain (g) | 4.44 ± 0.72 b | 3.58 ± 0.83 b | 7.89 ± 0.81 a | 2.01 ± 0.33 c |
| Liver (g) | 1.30 ± 0.09 c | 1.29 ± 0.10 c | 2.01 ± 0.26 a | 1.66 ± 0.20 b |
| Liver index (mg/g) | 36.87 ± 1.59 b | 37.24 ± 1.48 b | 44.51 ± 3.56 a | 41.25 ± 3.18 a |
| Kidney (g) | 0.40 ± 0.02 a | 0.42 ± 0.03 a | 0.40 ± 0.06 a | 0.45 ± 0.07 a |
| Kidney index (mg/g) | 11.28 ± 0.38 a | 11.45 ± 0.51 a | 8.80 ± 1.05 b | 11.34 ± 0.02 a |
| Perirenal fat pad (g) | 0.23 ± 0.06 c | 0.24 ± 0.06 c | 1.10 ± 0.21 a | 0.52 ± 0.15 b |
| Epididymal fat pad (g) | 0.90 ± 0.20 c | 0.92 ± 0.23 c | 2.51 ± 0.57 a | 1.74 ± 0.47 b |
| Visceral fat (g) | 1.13 ± 0.26 c | 1.19 ± 0.33 c | 3.61 ± 0.43 a | 2.26 ± 0.57 b |
| Visceral fat index (mg/g) | 32.17 ± 6.82 c | 34.15 ± 4.41 c | 80.23 ± 7.64 a | 56.60 ± 10.69 b |
| Item | ND | ND + STE | HFD | HFD + STE |
|---|---|---|---|---|
| ALT (IU/L) | 5.41 ± 2.14 c | 5.22 ± 2.32 c | 19.87 ± 4.88 a | 11.68 ± 3.52 b |
| AST (IU/L) | 4.12 ± 1.53 c | 4.56 ± 1.38 c | 15.01 ± 3.25 a | 9.83 ± 2.12 b |
| BUN (mmol/L) | 11.36 ± 2.18 a | 11.13 ± 1.98 a | 8.74 ± 0.88 b | 9.89 ± 0.95 ab |
| Cre (μmol/L) | 4.34 ± 0.51 a | 4.21 ± 0.38 a | 4.75 ± 0.86 a | 4.66 ± 0.45 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Zhang, L.; Liu, J.; Chan, A.S.C.; Li, D. Saponins of Tomato Extract Improve Non-Alcoholic Fatty Liver Disease by Regulating Oxidative Stress and Lipid Homeostasis. Antioxidants 2023, 12, 1848. https://doi.org/10.3390/antiox12101848
Yang Z, Zhang L, Liu J, Chan ASC, Li D. Saponins of Tomato Extract Improve Non-Alcoholic Fatty Liver Disease by Regulating Oxidative Stress and Lipid Homeostasis. Antioxidants. 2023; 12(10):1848. https://doi.org/10.3390/antiox12101848
Chicago/Turabian StyleYang, Ziming, Li Zhang, Jinlei Liu, Albert S. C. Chan, and Dianpeng Li. 2023. "Saponins of Tomato Extract Improve Non-Alcoholic Fatty Liver Disease by Regulating Oxidative Stress and Lipid Homeostasis" Antioxidants 12, no. 10: 1848. https://doi.org/10.3390/antiox12101848