
Effect of Encapsulation Material on Lipid Bioaccessibility and Oxidation during In Vitro Digestion of Black Seed Oil
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Chemicals and Materials
2.3. In Vitro Digestion of the Capsules
2.4. Extraction of the Lipids from the Digestates
2.5. 1H NMR Analysis
2.6. Nanoparticle Tracking Analysis
2.7. Sample Preparation for LC-MS Analysis
2.8. LC-MS Analysis of Free Oxylipins
2.9. Determination and Quantification of Tocols by HPLC-UV
2.10. Statistical Analysis
3. Results and Discussion
3.1. The Extent of Lipid Hydrolysis during In Vitro Digestion
3.2. Assessment of Lipid Oxidation of Encapsulated Black Seed Oil during In Vitro Digestion by 1H NMR
3.2.1. Changes in the Molar Percentages (%) of Black Seed Oil Acyl Groups and Fatty Acids
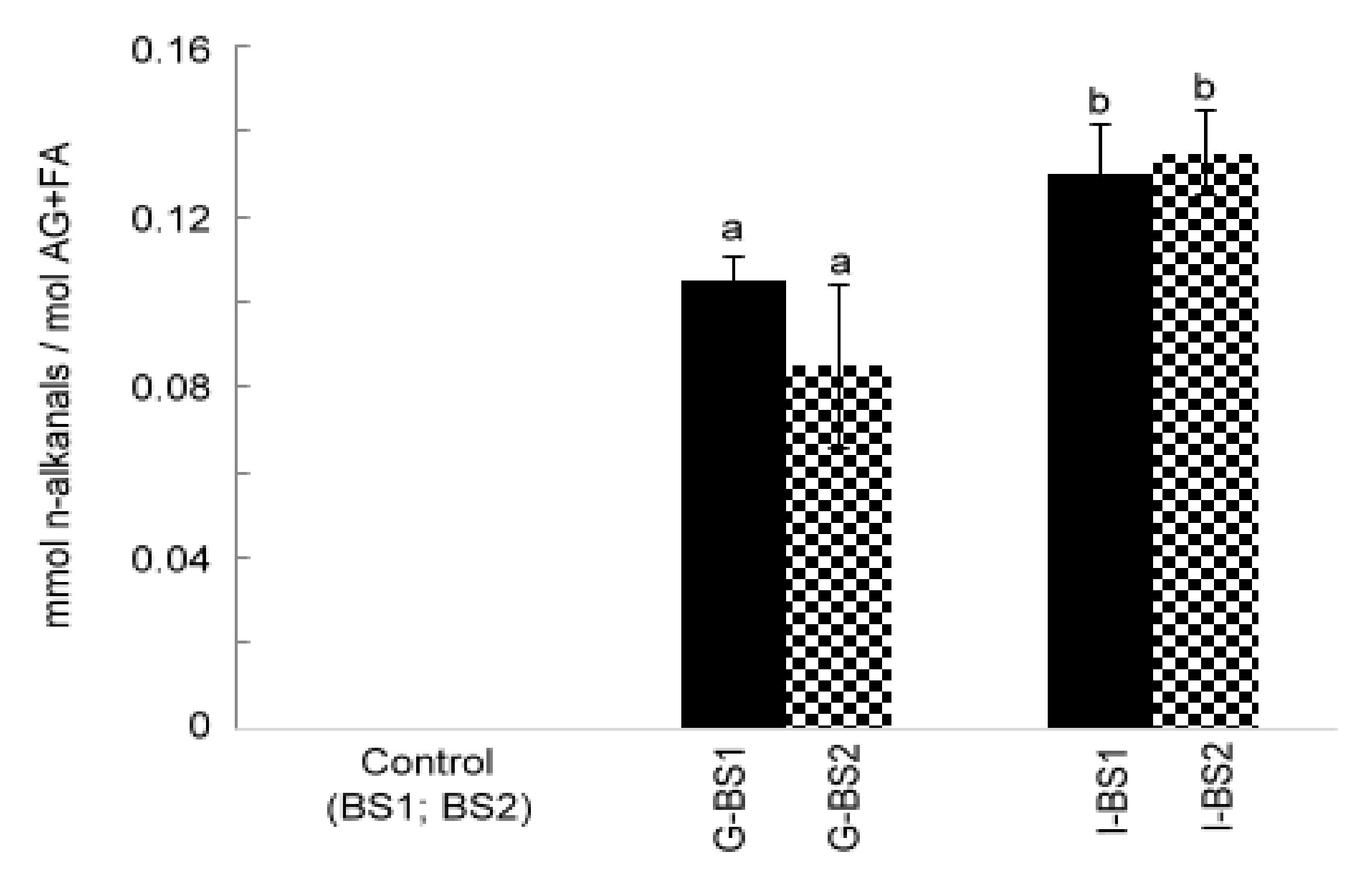
3.2.2. Formation of n-Alkanals
3.3. Assessment of Lipid Oxidation of Encapsulated Black Seed Oil during In Vitro Digestion by LC-MS: Formation and Detection of Free Oxylipins
3.4. In Vitro Assessment of the Bioaccessibility of Some Minor Compounds Present in Black Seed Oil
3.4.1. In Vitro Bioaccessibility of Tocols Present in Black Seed Oil
3.4.2. In Vitro Bioaccessibility of Other Minor Compounds Present in Black Seed Oil
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guillen, M.D.; Goicoechea, E. Formation of oxygenated α, β-unsaturated aldehydes and other toxic compounds in sunflower oil oxidation at room temperature in closed receptacles. Food Chem. 2008, 111, 157–164. [Google Scholar] [CrossRef]
- Martínez-Yusta, A.; Guillén, M.D. Monitoring compositional changes in sunflower oil-derived deep-frying media by 1H Nuclear Magnetic Resonance. Eur. J. Lipid Sci. Technol. 2016, 118, 984–996. [Google Scholar] [CrossRef]
- Nieva-Echevarría, B.; Goicoechea, E.; Guillén, M.D. Food lipid oxidation under gastrointestinal digestion conditions: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Alberdi-Cedeño, J.; Ibargoitia, M.L.; Guillén, M.D. Effect of the enrichment of corn oil with alpha- or gamma-tocopherol on its in vitro digestion studied by 1H NMR and SPME-GC/MS; formation of hydroperoxy-, hydroxy-, keto-dienes and keto-E-epoxy-E-monoenes in the more alpha-tocopherol enriched samples. Antioxidants 2020, 9, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberdi-Cedeño, J.; Ibargoitia, M.L.; Guillén, M.D. A global study by 1H NMR spectroscopy and SPME-GC/MS of the in vitro digestion of virgin flaxseed oil enriched or not with mono-, di-or tri-phenolic derivatives. Antioxidant efficiency of these compounds. Antioxidants 2020, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Alberdi-Cedeño, J.; Ibargoitia, M.L.; Guillén, M.D. Study of the in vitro digestion of olive oil enriched or not with antioxidant phenolic compounds. Relationships between bioaccessibility of main components of different oils and their composition. Antioxidants 2020, 9, 543. [Google Scholar] [CrossRef]
- Alberdi-Cedeño, J.; Ibargoitia, M.L.; Guillén, M.D. Oxylipins associated to current diseases detected for the first time in the oxidation of corn oil as a model system of oils rich in omega-6 polyunsaturated groups. A global, broad and in-depth study by 1H NMR spectroscopy. Antioxidants 2020, 9, 544. [Google Scholar] [CrossRef]
- Fruehwirth, S.; Zehentner, S.; Salim, M.; Sterneder, S.; Tiroch, J.; Lieder, B.; Pignitter, M. In vitro digestion of grape seed oil inhibits phospholipid-regulating effects of oxidized lipids. Biomolecules 2020, 10, 708. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Guillén, M.D.; Goicoechea, E. Toxic oxygenated α, β-unsaturated aldehydes and their study in foods: A review. Crit. Rev. Food Sci. Nutr. 2008, 48, 119–136. [Google Scholar] [CrossRef]
- Deol, P.; Fahrmann, J.; Yang, J.; Evans, J.R.; Rizo, A.; Grapov, D.; Salemi, M.; Wanichthanarak, K.; Fiehn, O.; Phinney, B.; et al. Omega-6 and omega-3 oxylipins are implicated in soybean oil-induced obesity in mice. Sci. Rep. 2017, 7, 12488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsden, C.E.; Domenichiello, A.F.; Yuan, Z.-X.; Sapio, M.R.; Keyes, G.S.; Mishra, S.K.; Gross, J.R.; Majchrzak-Hong, S.; Zamora, D.; Horowitz, M.S.; et al. A systems approach for discovering linoleic acid derivatives that potentially mediate pain and itch. Sci. Sig. 2017, 10, eaal5241. [Google Scholar] [CrossRef] [Green Version]
- Schulze, M.B.; Minihane, A.M.; Saleh, R.N.M.; Risérus, U. Intake and metabolism of omega-3 and omega-6 polyunsaturated fatty acids: Nutritional implications for cardiometabolic diseases. Lancet Diabetes Endocrinol. 2020, 8, 915–930. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.V.; Santos, M.R.C.; Silva, I.R.A.; Viana, E.B.M.; Dos Anjos, D.A.; Santos, I.A.; de Lima, N.G.B.; Wobeto, C.; Jorge, N.; Lannes, S.C.D.S. Synthetic and natural antioxidants used in the oxidative stability of edible oils: An overview. Food Rev. Int. 2021, 38, 349–372. [Google Scholar] [CrossRef]
- Timilsena, Y.P.; Adhikari, R.; Barrow, C.J.; Adhikari, B. Digestion behaviour of chia seed oil encapsulated in chia seed protein-gum complex coacervates. Food Hydrocoll. 2017, 66, 71–81. [Google Scholar] [CrossRef]
- Fu, J.; Song, L.; Liu, Y.; Bai, C.; Zhou, D.; Zhu, B.; Wang, T. Improving oxidative stability and release behavior of docosahexaenoic acid algae oil by microencapsulation. J. Sci. Food Agric. 2020, 100, 2774–2781. [Google Scholar] [CrossRef]
- Mu, H.; Song, Z.; Wang, X.; Wang, D.; Zheng, X.; Li, X. Microencapsulation of algae oil by complex coacervation of chitosan and modified starch: Characterization and oxidative stability. Int. J. Biol. Macromol. 2022, 194, 66–73. [Google Scholar] [CrossRef]
- Abang, S.; Chan, E.S.; Poncelet, D. Effects of process variables on the encapsulation of oil in ca-alginate capsules using an inverse gelation technique. J. Microencapsul. 2012, 29, 417–428. [Google Scholar] [CrossRef]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Nieva-Echevarría, B.; Goicoechea, E.; Manzanos, M.J.; Guillén, M.D. A study by 1H NMR on the influence of some factors affecting lipid in vitro digestion. Food Chem. 2016, 211, 17–26. [Google Scholar] [CrossRef]
- Augustin, M.A.; Sanguansri, L. Challenges in developing delivery systems for food additives, nutraceuticals and dietary supplements. In Encapsulation Technologies and Delivery Systems for Food Ingredients and Nutraceuticals; Woodhead Publishing: Sawston, UK, 2012; pp. 19–48. [Google Scholar]
- Mun, S.; Decker, E.A.; McClements, D.J. Influence of emulsifier type on in vitro digestibility of lipid droplets by pancreatic lipase. Food Res. Int. 2007, 40, 770–781. [Google Scholar] [CrossRef]
- Sathasivam, T.; Muniyandy, S.; Chuah, L.H.; Janarthanan, P. Encapsulation of red palm oil in carboxymethyl sago cellulose beads by emulsification and vibration technology: Physicochemical characterization and in vitro digestion. J. Food Eng. 2018, 231, 10–21. [Google Scholar] [CrossRef]
- Versantvoort, C.H.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.; Sips, A.J. Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Aracama, A.; Goicoechea, E.; Guillén, M.D. Direct study of minor extra-virgin olive oil components without any sample modification. 1H NMR multisupression experiment: A powerful tool. Food Chem. 2017, 228, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Goryainov, S.V.; Khromov, A.V.; Bakureza, G.; Cesar, E.; Ivlev, V.A.; Vorobyev, A.N.; Abramovich, R.A.; Potanina, O.G.; Novikov, O.O. Results of a comparative study of Nigella Sativa L. seeds oils composition. Sci. Pract. J. 2020, 8, 29–39. [Google Scholar] [CrossRef]
- Nieva-Echevarría, B.; Goicoechea, E.; Manzanos, M.J.; Guillén, M.D. A method based on 1H NMR spectral data useful to evaluate the hydrolysis level in complex lipid mixtures. Food Res. Int. 2014, 66, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Emami, S.; Zhang, Z.; Taha, A.Y. Quantitation of oxylipins in fish and algae oil supplements using optimized hydrolysis procedures and ultra-high performance liquid chromatography coupled to tandem mass-spectrometry. J. Agric. Food Chem. 2020, 68, 9329–9344. [Google Scholar] [CrossRef]
- Nieva-Echevarría, B.; Goicoechea, E.; Manzanos, M.J.; Guillén, M.D. 1H NMR and SPME-GC/MS study of hydrolysis, oxidation and other reactions occurring during in vitro digestion of non-oxidized and oxidized sunflower oil. Formation of hydroxy-octadecadienoates. Food Res. Int. 2017, 91, 171–182. [Google Scholar] [CrossRef]
- Martin-Rubio, A.S.; Sopelana, P.; Guillén, M.D. The key role of ovalbumin in lipid bioaccessibility and oxidation product profile during the in vitro digestion of slightly oxidized soybean oil. Food Funct. 2019, 10, 4440–4451. [Google Scholar] [CrossRef] [Green Version]
- Nieva-Echevarría, B.; Goicoechea, E.; Guillén, M.D. Effect of adding alpha-tocopherol on the oxidation advance during in vitro gastrointestinal digestion of sunflower and flaxseed oils. Food Res. Int. 2019, 125, 108558. [Google Scholar] [CrossRef]
- Choe, E.; Min, D.B. Mechanisms and factors for edible oil oxidation. Compr. Rev. Food Sci. Food Saf. 2006, 5, 169–186. [Google Scholar] [CrossRef]
- Matthaus, B.; Özcan, M.M. Fatty acids, tocopherol, and sterol contents of some Nigella species seed oil. Czech J. Food Sci. 2011, 29, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Mazaheri, Y.; Torbati, M.; Azadmard-Damirchi, S.; Savage, G.P. A comprehensive review of the physicochemical, quality and nutritional properties of Nigella sativa oil. Food Rev. Int. 2019, 35, 342–362. [Google Scholar] [CrossRef]
- Ermumcu, M.Ş.K.; Şanlıer, N. Black cumin (Nigella sativa) and its active component of thymoquinone: Effects on health. Food Health 2017, 3, 170–183. [Google Scholar] [CrossRef]
- Hannan, M.A.; Rahman, M.A.; Sohag, A.A.M.; Uddin, M.J.; Dash, R.; Sikder, M.H.; Rahman, M.S.; Timalsina, B.; Munni, Y.A.; Sarker, P.P.; et al. Black cumin (Nigella sativa L.): A comprehensive review on phytochemistry, health benefits, molecular pharmacology, and safety. Nutrients 2021, 13, 1784. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Rauf, A.; Khan, I.A.; Shahbaz, M.; Qaisrani, T.B.; Fatmawati, S.; Abu-Izneid, T.; Imran, A.; Rahman, K.U.; Gondal, T.A. Thymoquinone: A novel strategy to combat cancer: A review. Biomed. Pharmacother. 2018, 106, 390–402. [Google Scholar] [CrossRef]
- Abraham, A.; Kattoor, A.J.; Saldeen, T.; Mehta, J.L. Vitamin E and its anticancer effects. Crit. Rev. Food Sci. Nutr. 2019, 59, 2831–2838. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, R.; Chang, M.; Jin, Q.; Zhang, H.; Wang, X. Health benefits of 4,4-dimethyl phytosterols: An exploration beyond 4-desmethyl phytosterols. Food Funct. 2020, 11, 93–110. [Google Scholar] [CrossRef]
- Dufour, C.; Loonis, M. Regio- and stereoselective oxidation of linoleic acid bound to serum albumin: Identification by ESI–mass spectrometry and NMR of the oxidation products. Chem. Phys. Lipids 2005, 138, 60–68. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Acylglycerol Species (Molar %) | LB (%) | ||||||
|---|---|---|---|---|---|---|---|
| Samples | TG% | 1,2-DG% | 1,3%-DG% | 1-MG% | 2-MG% | Gol% | |
| BS1 | 76.9 ± 0.19 a | 5.29 ± 0.11 a | - | 1.50 ± 0.04 a | 0.17 ± 0.01 a | 16.2 ± 0.34 a | 19.6 ± 0.26 a |
| G-BS1 | 42.6 ± 1.10 b | 27.9 ± 0.36 b | 2.40 ± 0.29 a | 0.84 ± 0.04 b | 6.34 ± 0.39 b | 20.0 ± 0.64 b | 37.2 ± 1.03 b |
| I-BS1 | 24.0 ± 5.41 c | 14.2 ± 1.41 c | 4.83 ± 1.14 b | 6.82 ± 2.37 c | 17.1 ± 4.10 c | 33.0 ± 2.80 c | 63.3 ± 7.05 c |
| BS2 | 82.7 ± 0.22 d | 4.09 ± 0.10 d | - | 1.39 ± 0.11 a | 0.12 ± 0.00 d | 11.8 ± 0.23 d | 14.6 ± 0.15 d |
| G-BS2 | 64.2 ± 4.01 e | 17.1 ± 2.41 c | 4.57 ± 0.41 b | 0.87 ± 0.09 b | 2.12 ± 0.48 e | 11.1 ± 1.45 e | 21.3 ± 2.65 e |
| I-BS2 | 25.8 ± 3.93 c | 16.7 ± 1.04 c | 4.21 ± 0.47 b | 5.26 ± 0.44 c | 18.8 ± 1.75 c | 29.2 ± 2.37 c | 60.2 ± 3.70 c |
| Oxylipins | BS1 | G-BS1 | I-BS1 | BS2 | G-BS2 | I-BS2 |
|---|---|---|---|---|---|---|
| HpODE | 3.00 ± 0.57 a | 61.6 ± 9.70 b | 147 ± 28.8 c | 4.47 ± 0.72 a | 24.3 ± 8.02 d | 85.6 ± 6.11 e |
| HODE | 2.25 ± 0.26 a | 27.8 ± 1.14 b | 24.5 ± 1.98 c | 2.32 ± 0.17 a | 6.19 ± 0.83 d | 5.82 ± 0.44 d |
| oxoODE | 1.14 ± 0.10 a | 10.6 ± 0.73 b | 8.72 ± 0.77 c | 1.06 ± 0.06 a | 2.92 ± 0.39 d | 2.71 ± 0.24 d |
| EpOME | 0.42 ± 0.07 a | 5.21 ± 0.22 b | 4.48 ± 0.41 c | 0.46 ± 0.05 a | 1.21 ± 0.20 d | 1.13 ± 0.09 d |
| DiHOME | 2.67 ± 0.40 a | 13.1 ± 1.47 b | 9.23 ± 0.93 c | 1.10 ± 0.10 d | 1.83 ± 0.09 ad | 1.19 ± 0.09 d |
| TriHOME | 0.28 ± 0.09 a | 5.89 ± 0.66 b | 1.98 ± 0.36 c | 0.25 ± 0.03 a | 1.28 ± 0.14 d | 0.63 ± 0.06 e |
| Total | 9.76 ± 1.50 a | 124 ± 13.9 b | 196 ± 33.2 c | 9.68 ± 1.13 a | 37.7 ± 9.69 d | 97.0 ± 7.04 e |
| Minor Compounds | BS1 | G-BS1 | I-BS1 | BS2 | G-BS2 | I-BS2 |
|---|---|---|---|---|---|---|
| α-Tocopherol (μg/g oil) | 92.3 ± 3.19 a | 64.7 ± 12.8 b | - | 54.2 ± 9.74 b | 18.8 ± 3.22 c | - |
| γ-Tocopherol (μg/g oil) | 388 ± 77.4 a | 318 ± 19.0 a | 243 ± 26.0 b | 112 ± 14.7 c | 117 ± 8.72 c | 37.2 ± 8.50 d |
| β-Tocotrienol (μg/g oil) | 363 ± 12.8 a | 326 ± 12.3 a | 243 ± 29.2 b | 394 ± 35.8 a | 334 ± 17.6 a | 226 ± 13.7 b |
| Δ7-Avenasterol (mmol/mol AG + FA) | 0.10 ± 0.00 a | 0.10 ± 0.01 a | 0.07 ± 0.01 b | 0.14 ± 0.01 c | 0.14 ± 0.00 c | 0.09 ± 0.01 d |
| Esters of cycloartenol (mmol/mol AG + FA) | 0.29 ± 0.01 a | 0.28 ± 0.01 a | 0.27 ± 0.02 a | 0.32 ± 0.02 a | 0.28 ± 0.01 a | 0.25 ± 0.02 a |
| Sitostanol (mmol/mol AG + FA) | 0.05 ± 0.00 a | 0.04 ± 0.01 a | - | 0.05 ± 0.00 a | 0.05 ± 0.00 a | - |
| Thymoquinone (mmol/mol AG + FA) | 1.75 ± 0.01 a | 1.26 ± 0.06 b | 1.14 ± 0.06 c | 2.77 ± 0.04 d | 1.88 ± 0.11 e | 1.66 ± 0.18 f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alberdi-Cedeño, J.; Aichner, M.; Mistlberger-Reiner, A.; Shi, A.; Pignitter, M. Effect of Encapsulation Material on Lipid Bioaccessibility and Oxidation during In Vitro Digestion of Black Seed Oil. Antioxidants 2023, 12, 191. https://doi.org/10.3390/antiox12010191
Alberdi-Cedeño J, Aichner M, Mistlberger-Reiner A, Shi A, Pignitter M. Effect of Encapsulation Material on Lipid Bioaccessibility and Oxidation during In Vitro Digestion of Black Seed Oil. Antioxidants. 2023; 12(1):191. https://doi.org/10.3390/antiox12010191
Chicago/Turabian StyleAlberdi-Cedeño, Jon, Martha Aichner, Agnes Mistlberger-Reiner, Aimin Shi, and Marc Pignitter. 2023. "Effect of Encapsulation Material on Lipid Bioaccessibility and Oxidation during In Vitro Digestion of Black Seed Oil" Antioxidants 12, no. 1: 191. https://doi.org/10.3390/antiox12010191