
Development of Dietary Thiol Antioxidant via Reductive Modification of Whey Protein and Its Application in the Treatment of Ischemic Kidney Injury
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Whey Protein and Its Preparation
2.3. Mice
2.4. Induction of Renal Ischemia Reperfusion Injury
2.5. Western Blot Analysis
2.6. Modification of Whey Protein and Albumin with Reductive Chemical DTT
2.7. Maleimide-Labeling Assay
2.8. Measurement of Free Sulfhydryl Groups (-SH) in Proteins Using Ellman’s Reagent
2.9. Enzyme-Linked Immunosorbent Assay and Renal Function Analysis
2.10. Kidney Histology Analysis
2.11. Detection of Sulfenic Acid
2.12. Cell Culture
2.13. Calcein-AM and Propidium Iodide (PI) Staining
2.14. Assessment of Cell Viability with WST Reagent
2.15. Analysis of Hydrogen-Peroxide-Quenching Capacity of the Modified WP
2.16. GSH/GSSG Assay
2.17. Measurement of BUN, Creatinine
2.18. Statistical Analysis
3. Results
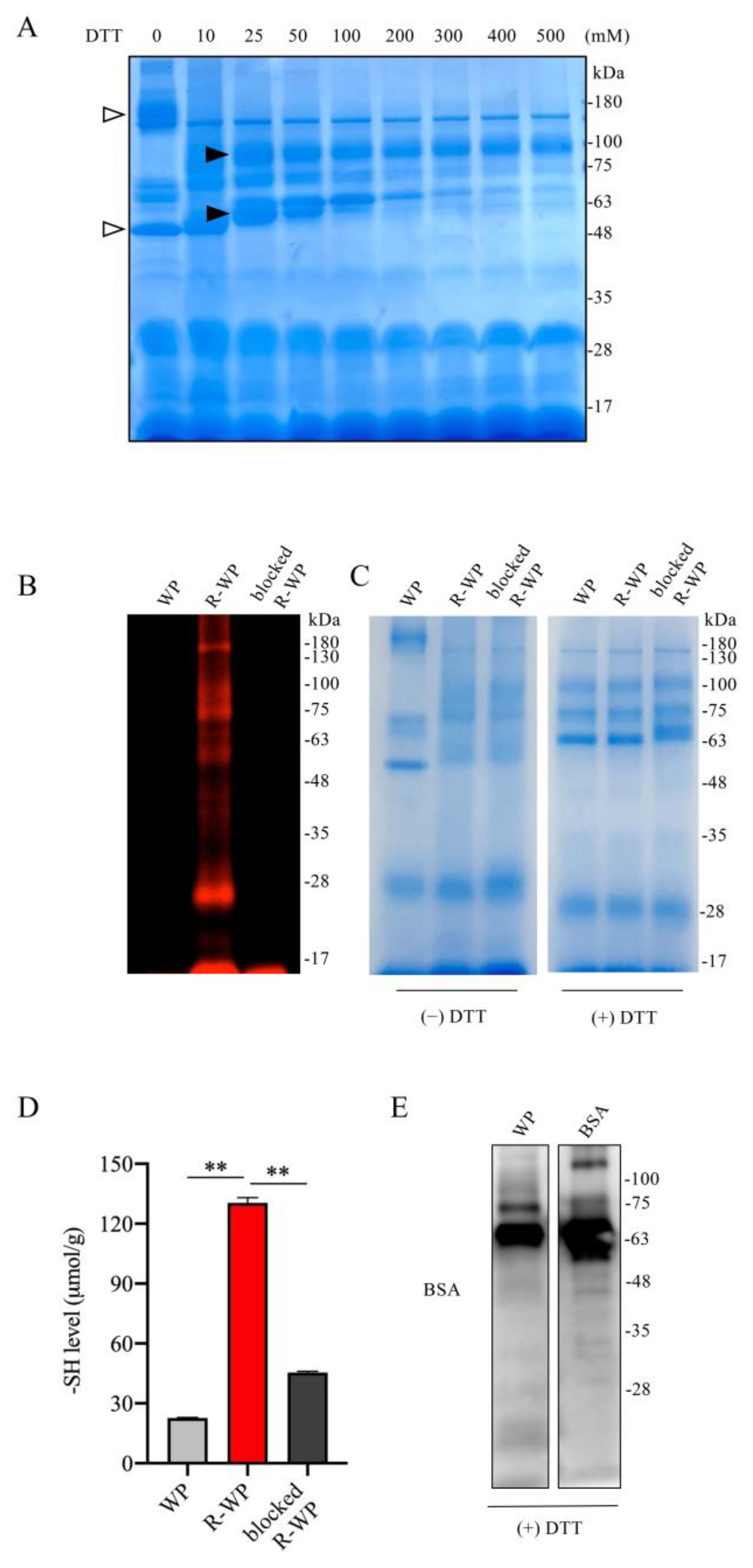
3.1. Reductive Modification of WP Increases Its -SH Activity
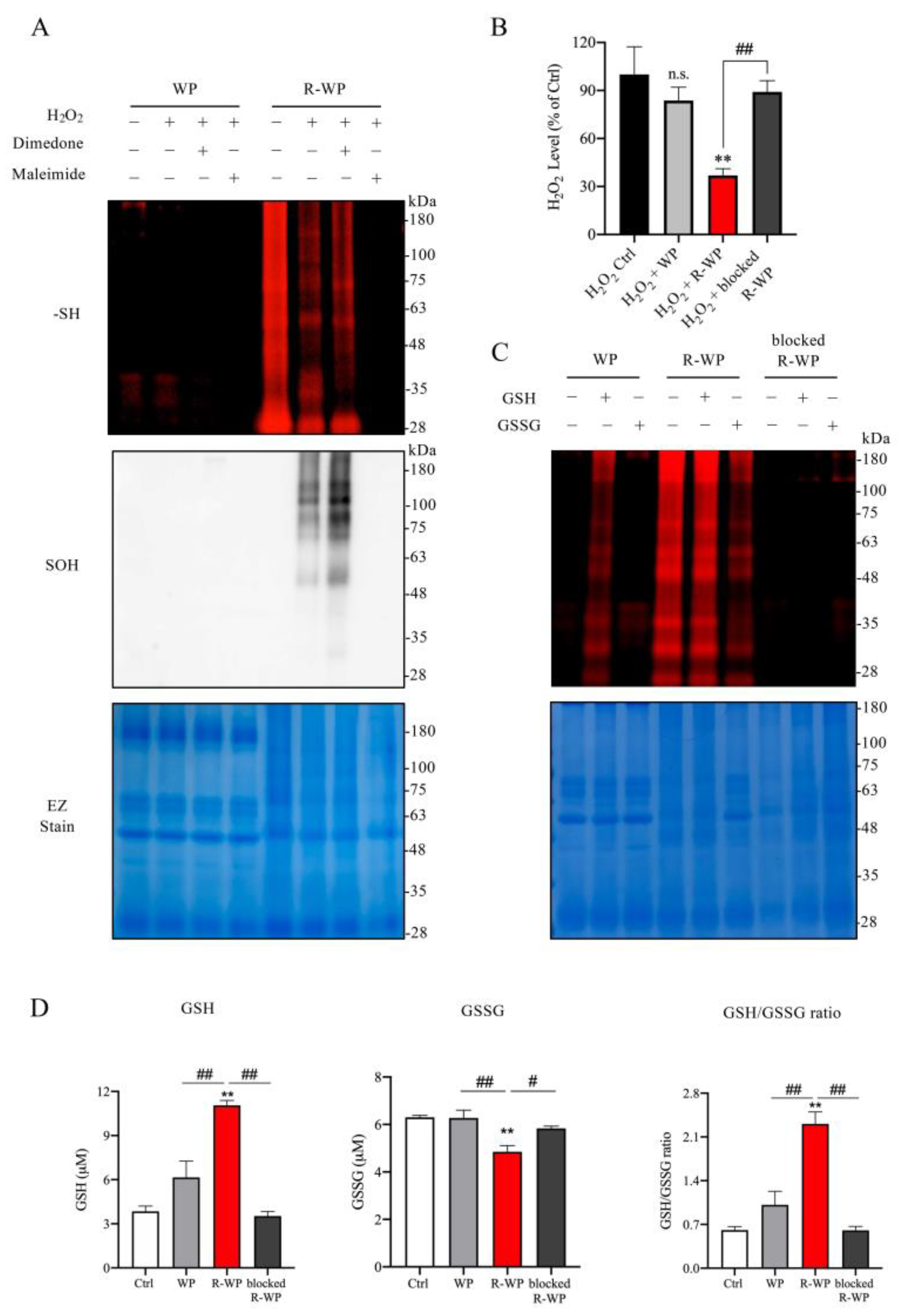
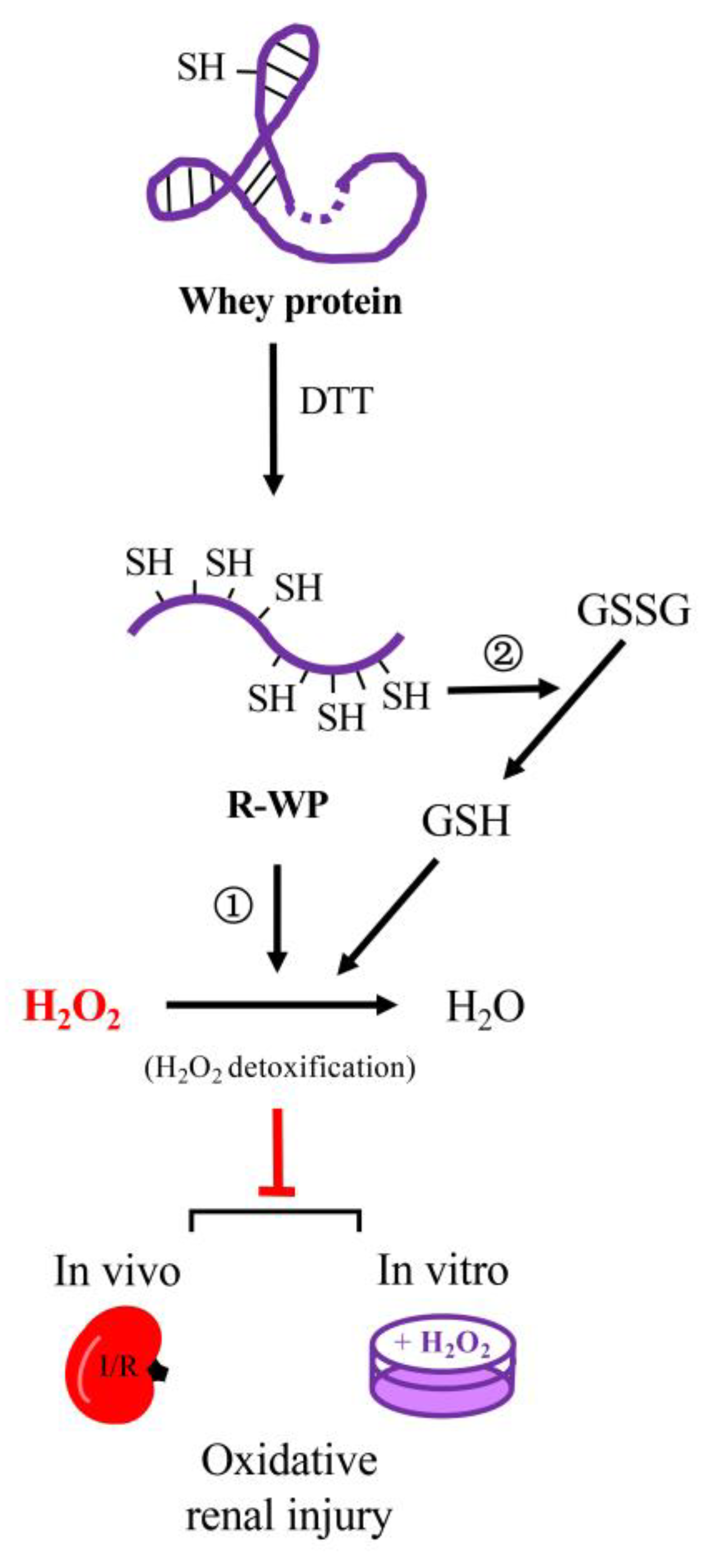
3.2. R-WP Reacts with H2O2 and GSH/GSSG via Thiol–Disulfide Exchange
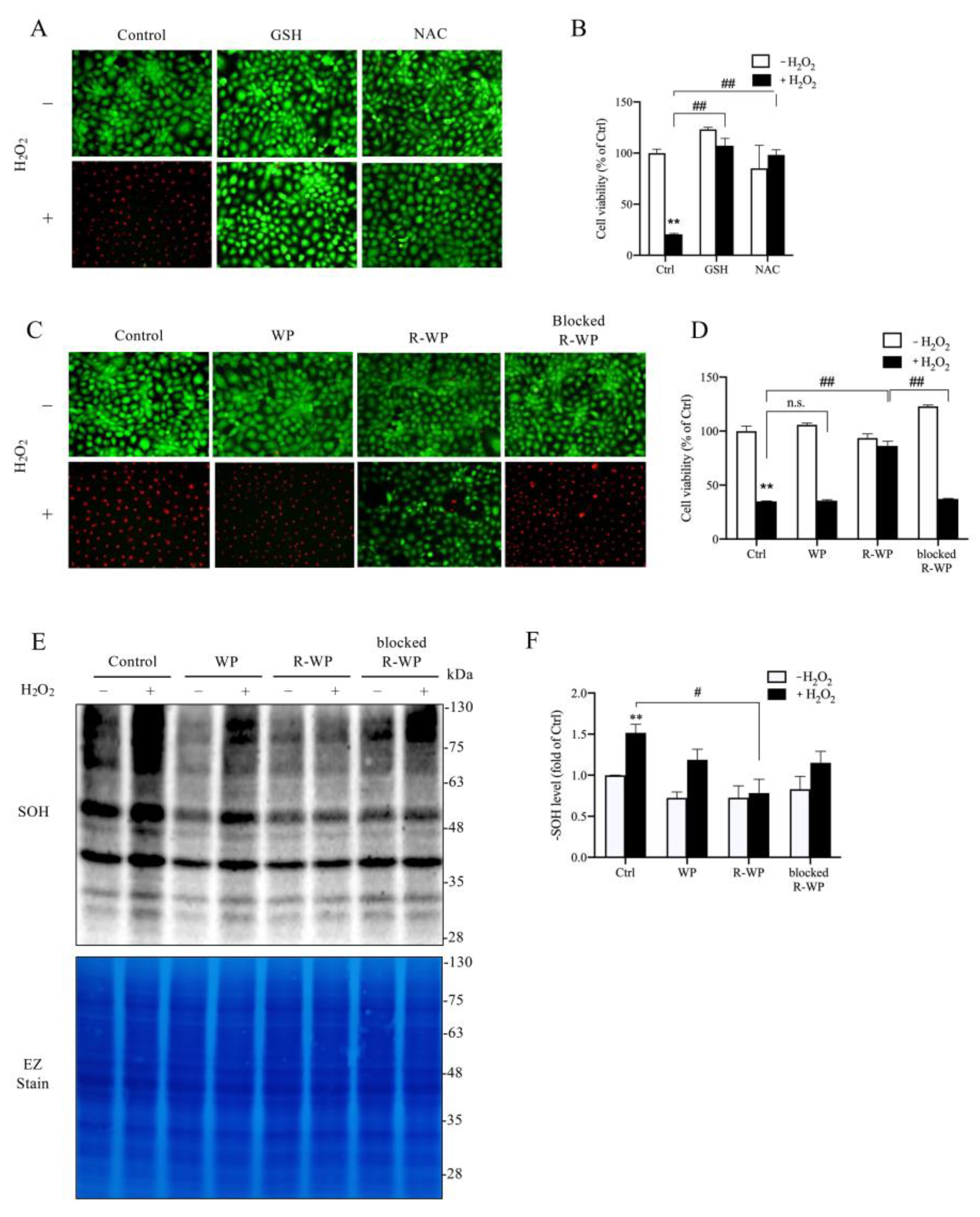
3.3. R-WP Protects Renal Tubular Cells from H2O2-Elicited Cell Injury
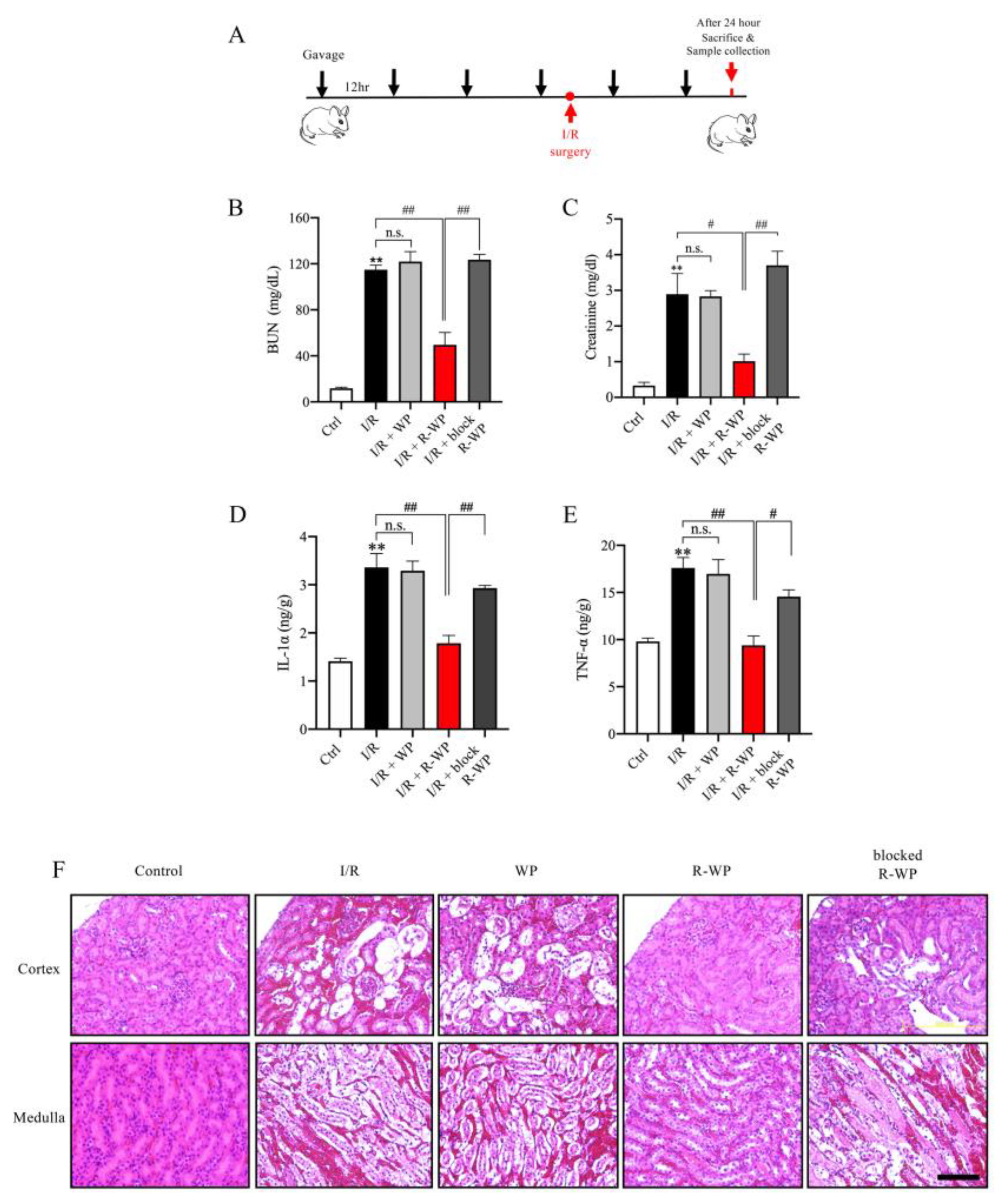
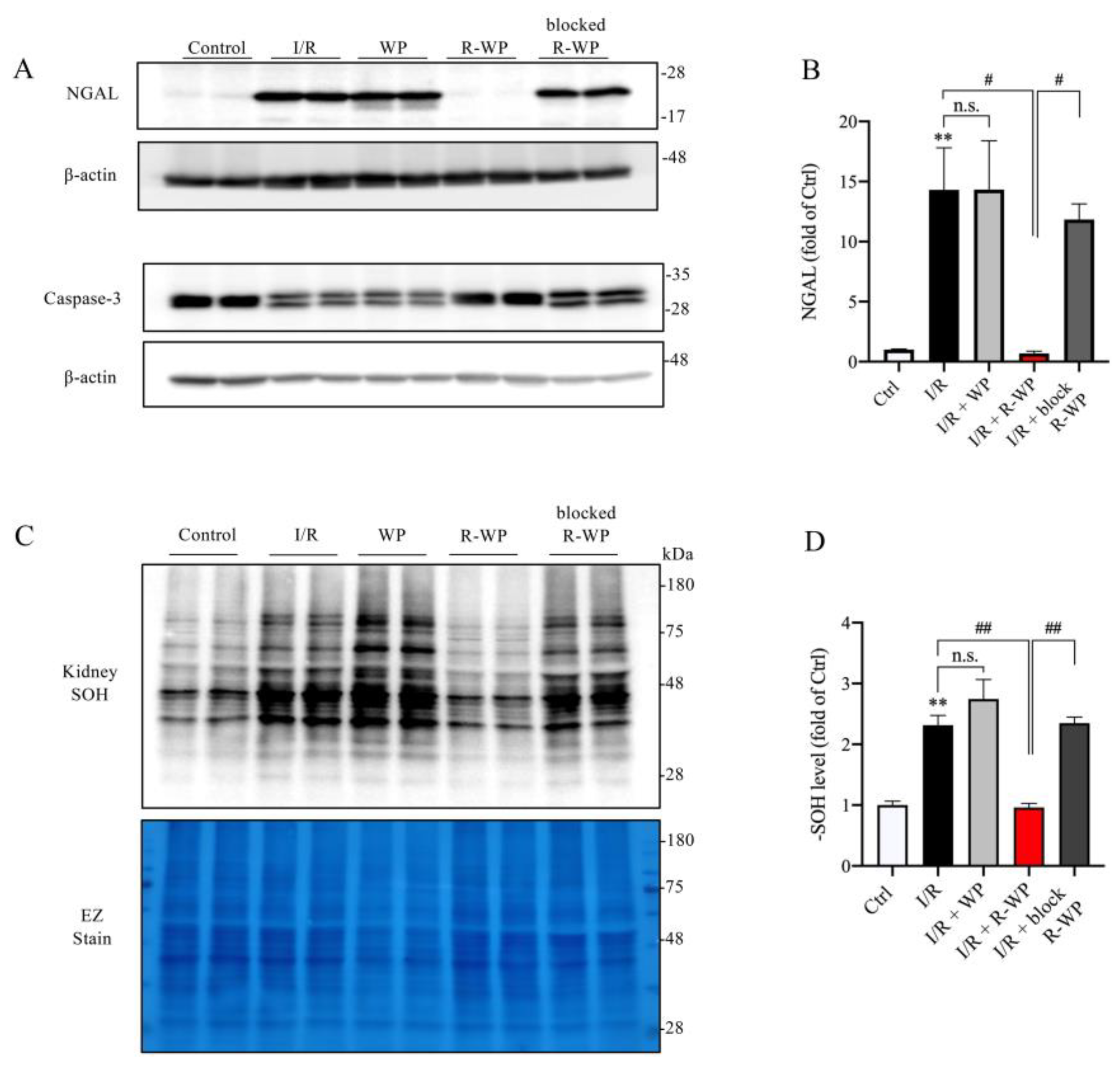
3.4. R-WP Protects Kidney from I/R-Induced Injury
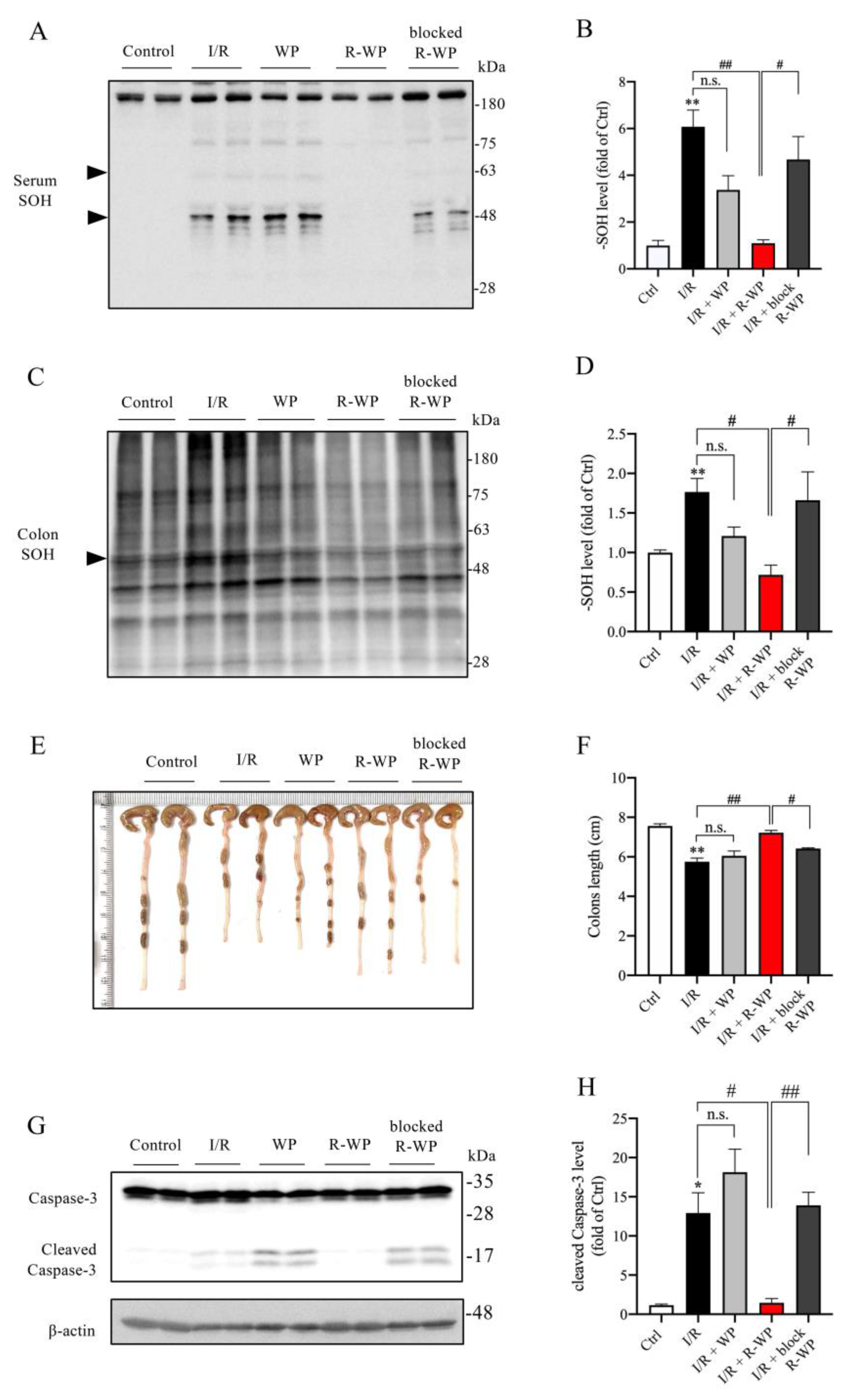
3.5. Administration of R-WP Leads to an Improvement in Systemic Redox Status
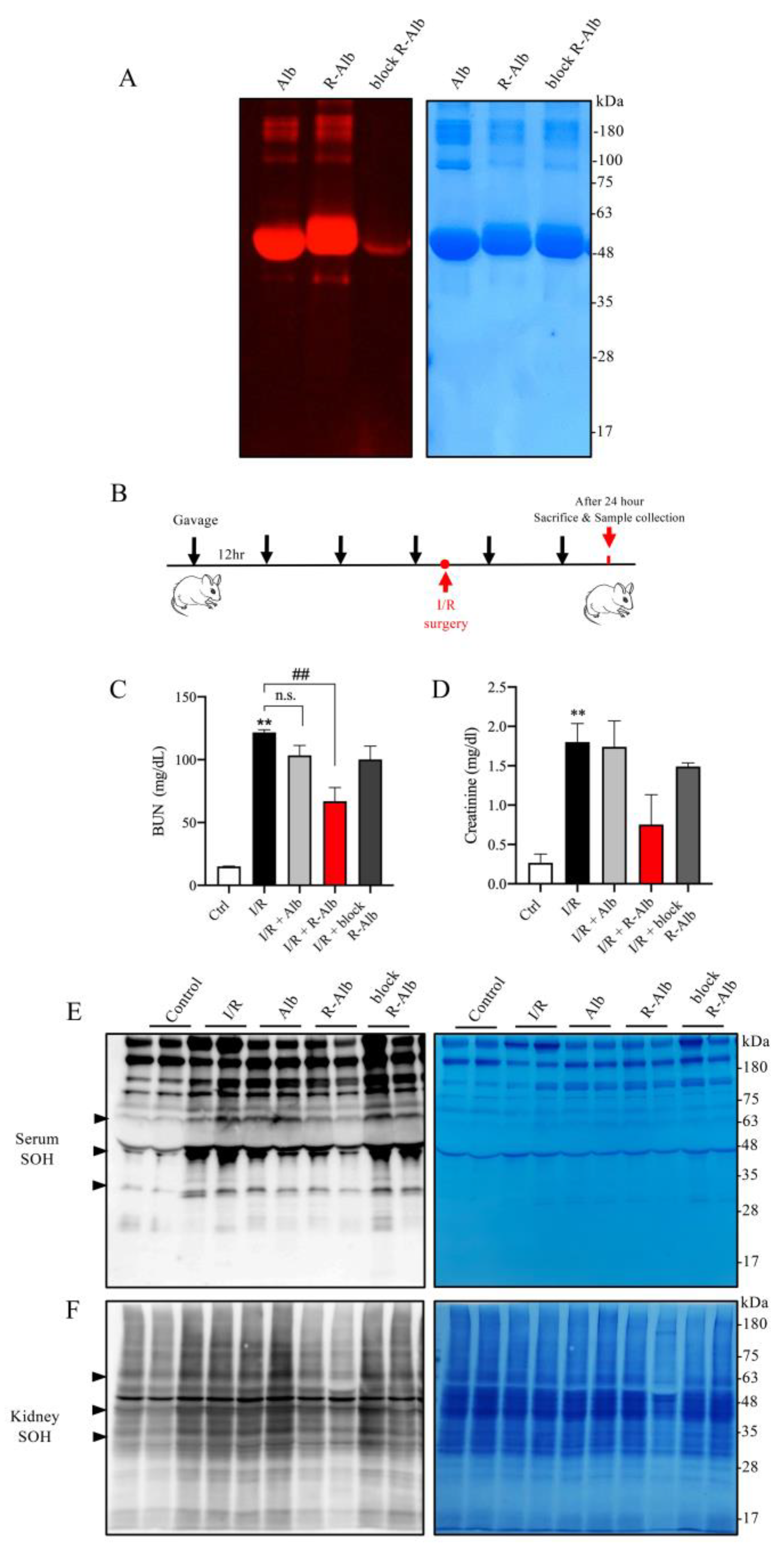
3.6. Reductively Modified Albumin Similarly Alleviates Renal IRI
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug. Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Deneke, S.M. Thiol-based antioxidants. Curr. Top. Cell. Regul. 2001, 36, 151–180. [Google Scholar]
- Bonanata, J.; Turell, L.; Antmann, L.; Ferrer-Sueta, G.; Botasini, S.; Mendez, E.; Alvarez, B.; Coitino, E.L. The thiol of human serum albumin: Acidity, microenvironment and mechanistic insights on its oxidation to sulfenic acid. Free Radic. Biol. Med. 2017, 108, 952–962. [Google Scholar] [CrossRef]
- Turell, L.; Radi, R.; Alvarez, B. The thiol pool in human plasma: The central contribution of albumin to redox processes. Free Radic. Biol. Med. 2013, 65, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Colombo, G.; Clerici, M.; Giustarini, D.; Rossi, R.; Milzani, A.; Dalle-Donne, I. Redox albuminomics: Oxidized albumin in human diseases. Antioxid. Redox Signal. 2012, 17, 1515–1527. [Google Scholar] [CrossRef]
- Carballal, S.; Radi, R.; Kirk, M.C.; Barnes, S.; Freeman, B.A.; Alvarez, B. Sulfenic acid formation in human serum albumin by hydrogen peroxide and peroxynitrite. Biochemistry 2003, 42, 9906–9914. [Google Scholar] [CrossRef] [PubMed]
- Nagumo, K.; Tanaka, M.; Chuang, V.T.G.; Setoyama, H.; Watanabe, H.; Yamada, N.; Kubota, K.; Tanaka, M.; Matsushita, K.; Yoshida, A. Cys34-cysteinylated human serum albumin is a sensitive plasma marker in oxidative stress-related chronic diseases. PLoS ONE 2014, 9, e85216. [Google Scholar] [CrossRef]
- King, T.P. On the sulfhydryl group of human plasma albumin. J. Biol. Chem. 1961, 236, PC5. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S. Role of human serum albumin and oxidative stress in diabetes. J. Appl. Biotechnol. Bioeng. 2017, 3, 00057. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Mao, Z.; Huang, Y.; Yan, H.; Yan, Q.; Hong, J.; Fan, J.; Yao, J. Reductively modified albumin attenuates DSS-Induced mouse colitis through rebalancing systemic redox state. Redox. Biol. 2021, 41, 101881. [Google Scholar] [CrossRef] [PubMed]
- Deeth, H.C.; Bansal, N. Whey Proteins: From Milk to Medicine; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Corrochano, A.R.; Buckin, V.; Kelly, P.M.; Giblin, L. Invited review: Whey proteins as antioxidants and promoters of cellular antioxidant pathways. J. Dairy Sci. 2018, 101, 4747–4761. [Google Scholar] [CrossRef] [Green Version]
- Quig, D. Cysteine metabolism and metal toxicity. Altern. Med. Rev. 1998, 3, 262–270. [Google Scholar] [PubMed]
- Chitapanarux, T.; Tienboon, P.; Pojchamarnwiputh, S.; Leelarungrayub, D. Open-labeled pilot study of cysteine-rich whey protein isolate supplementation for nonalcoholic steatohepatitis patients. J. Gastroenterol. Hepatol. 2009, 24, 1045–1050. [Google Scholar] [CrossRef]
- Doi, K.; Suzuki, Y.; Nakao, A.; Fujita, T.; Noiri, E. Radical scavenger edaravone developed for clinical use ameliorates ischemia/reperfusion injury in rat kidney. Kidney Int. 2004, 65, 1714–1723. [Google Scholar] [CrossRef] [Green Version]
- Malek, M.; Nematbakhsh, M. Renal ischemia/reperfusion injury; from pathophysiology to treatment. J. Ren. Inj. Prev. 2015, 4, 20. [Google Scholar] [PubMed]
- Rodrigo, R.; Fernandez-Gajardo, R.; Gutierrez, R.; Manuel Matamala, J.; Carrasco, R.; Miranda-Merchak, A.; Feuerhake, W. Oxidative stress and pathophysiology of ischemic stroke: Novel therapeutic opportunities. CNS Neurol. Disord. 2013, 12, 698–714. [Google Scholar] [CrossRef]
- Jayatilake, S.; Arai, K.; Kumada, N.; Ishida, Y.; Tanaka, I.; Iwatsuki, S.; Ohwada, T.; Ohnishi, M.; Tokuji, Y.; Kinoshita, M. The effect of oral intake of low-temperature-processed whey protein concentrate on colitis and gene expression profiles in mice. Foods 2014, 3, 351–368. [Google Scholar] [CrossRef]
- Kijima, Y.; Iwatsuki, S.; Akamatsu, H.; Terato, K.; Kuwabara, Y.; Ueda, S.; Shionoya, H. Natural antibodies to pathogenic bacteria and their toxins in whey protein concentrate. J. Jpn. Soc. Food Sci. Technol. 2009, 56, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Katayama, K.; Matsuno, T.; Waritani, T.; Terato, K.; Shionoya, H. Supplemental treatment of rheumatoid arthritis with natural milk antibodies against enteromicrobes and their toxins: Results of an open-labelled pilot study. Nutr. J. 2011, 10, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Cao, C.; Wu, Z.; Zhang, H.; Sun, Z.; Wang, M.; Xu, H.; Zhao, Z.; Wang, Y.; Pei, G. The probiotic L. casei Zhang slows the progression of acute and chronic kidney disease. Cell. Metab. 2021, 33, 1926–1942.e8. [Google Scholar] [CrossRef] [PubMed]
- Saurin, A.T.; Neubert, H.; Brennan, J.P.; Eaton, P. Widespread sulfenic acid formation in tissues in response to hydrogen peroxide. Proc. Natl. Acad. Sci. USA. 2004, 101, 17982–17987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, C.K.; Packer, L. Thiol homeostasis and supplements in physical exercise. Am. J. Clin. Nutr. 2000, 72, 653S–669S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitz, R.; Kregel, K.C. Thiol supplementation in aged animals alters antioxidant. J. Appl. Physiol. 2005, 99, 2271–2277. [Google Scholar]
- Benedikter, B.J.; Weseler, A.R.; Wouters, E.F.; Savelkoul, P.H.; Rohde, G.G.; Stassen, F.R. Redox-dependent thiol modifications: Implications for the release of extracellular vesicles. Cell. Mol. Life Sci. 2018, 75, 2321–2337. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, P.V.; Laurindo, F.R. Implications of plasma thiol redox in disease. Clin. Sci. 2018, 132, 1257–1280. [Google Scholar] [CrossRef]
- Erel, O.; Erdogan, S. Thiol-disulfide homeostasis: An integrated approach with biochemical and clinical aspects. Turk. J. Med. Sci. 2020, 50, 1728–1738. [Google Scholar] [CrossRef]
- Ghezzi, P.; Bonetto, V.; Fratelli, M. Thiol-disulfide balance: From the concept of oxidative stress to that of redox regulation. Antioxid. Redox Signal. 2005, 7, 964–972. [Google Scholar] [CrossRef]
- Gould, R.L.; Pazdro, R. Impact of supplementary amino acids, micronutrients, and overall diet on glutathione homeostasis. Nutrients 2019, 11, 1056. [Google Scholar] [CrossRef] [Green Version]
- Kerasioti, E.; Stagos, D.; Priftis, A.; Aivazidis, S.; Tsatsakis, A.M.; Hayes, A.W.; Kouretas, D. Antioxidant effects of whey protein on muscle C2C12 cells. Food Chem. 2014, 155, 271–278. [Google Scholar] [CrossRef]
- Liu, Q.; Cheng, J.; Sun, X.; Guo, M. Preparation, characterization, and antioxidant activity of zein nanoparticles stabilized by whey protein nanofibrils. Int. J. Biol. Macromol. 2021, 167, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Vavrusova, M.; Pindstrup, H.; Johansen, L.B.; Andersen, M.L.; Andersen, H.J.; Skibsted, L.H. Characterisation of a whey protein hydrolysate as antioxidant. Int. Dairy J. 2015, 47, 86–93. [Google Scholar] [CrossRef]
- Rodzik, A.; Pomastowski, P.; Sagandykova, G.N.; Buszewski, B. Interactions of whey proteins with metal ions. Int. J. Mol. Sci. 2020, 21, 2156. [Google Scholar] [CrossRef] [Green Version]
- Stobiecka, M.; Krol, J.; Brodziak, A. Antioxidant activity of milk and dairy products. Animals. 2022, 12, 41. [Google Scholar] [CrossRef]
- Cuzzocrea, S.; Riley, D.P.; Caputi, A.P.; Salvemini, D. Antioxidant therapy: A new pharmacological approach in shock, inflammation, and ischemia/reperfusion injury. Pharmacol. Rev. 2001, 53, 135–159. [Google Scholar]
- Ishihara, T.; Tanaka, K.-I.; Tasaka, Y.; Namba, T.; Suzuki, J.; Ishihara, T.; Okamoto, S.; Hibi, T.; Takenaga, M.; Igarashi, R. Therapeutic effect of lecithinized superoxide dismutase against colitis. J. Pharmacol. Exp. Ther. 2009, 328, 152–164. [Google Scholar] [CrossRef] [Green Version]
- Munro, D.; Treberg, J.R. A radical shift in perspective: Mitochondria as regulators of reactive oxygen species. J. Pharmacol. Exp. Ther. 2017, 220, 1170–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledo, A.; Fernandes, E.; Salvador, A.; Laranjinha, J.; Barbosa, R. In vivo hydrogen peroxide diffusivity in brain tissue supports volume signaling activity. Redox Biol. 2022, 50, 102250. [Google Scholar] [CrossRef] [PubMed]
- Sousa, T.; Gouveia, M.; Travasso, R.D.; Salvador, A. How abundant are superoxide and hydrogen peroxide in the vasculature lumen, how far can they reach? Redox Biol. 2022, 58, 102527. [Google Scholar] [CrossRef]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- Turell, L.; Carballal, S.; Botti, H.; Radi, R.; Alvarez, B. Oxidation of the albumin thiol to sulfenic acid and its implications in the intravascular compartment. Braz. J. Med. Biol. Res. 2009, 42, 305–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.Y.; Liou, S.Y.; Kuo, W.W.; Wu, H.C.; Chang, Y.L.; Chen, T.S. Chemiluminescence analysis of antioxidant capacity for serum albumin isolated from healthy or uremic volunteers. Luminescence 2016, 31, 1474–1478. [Google Scholar] [CrossRef] [PubMed]
- Kunduzova, O.R.; Bianchi, P.; Parini, A.; Cambon, C. Hydrogen peroxide production by monoamine oxidase during ischemia/reperfusion. Eur. J. Pharmacol. 2002, 448, 225–230. [Google Scholar] [CrossRef]
- Gonzalez-Flecha, B.; Boveris, A. Mitochondrial sites of hydrogen peroxide production in reperfused rat kidney cortex. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 1995, 1243, 361–366. [Google Scholar] [CrossRef]
- Chang, E.; Lee, T.; Mun, K.; Kim, H.; Suh, S.; Bae, J.; Kim, S.; Cho, K.; Hwang, J. Effects of polyhemoglobin-antioxidant enzyme complex on ischemia-reperfusion in kidney. Transplant. Proc. 2004, 36, 1952–1954. [Google Scholar] [CrossRef]
- Nitescu, N.; Ricksten, S.-E.; Marcussen, N.; Haraldsson, B.; Nilsson, U.; Basu, S.; Guron, G. N-acetylcysteine attenuates kidney injury in rats subjected to renal ischaemia-reperfusion. Nephrol. Dial. Transplant. 2006, 21, 1240–1247. [Google Scholar] [CrossRef] [Green Version]
- Heppner, D.E.; Janssen-Heininger, Y.M.; van der Vliet, A. The role of sulfenic acids in cellular redox signaling: Reconciling chemical kinetics and molecular detection strategies. Arch. Biochem. Biophys. 2017, 616, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Croguennec, T.; Bouhallab, S.d.; Mollé, D.; O’kennedy, B.T.; Mehra, R. Stable monomeric intermediate with exposed Cys-119 is formed during heat denaturation of β-lactoglobulin. Biochem. Biophys. Res. Commun. 2003, 301, 465–471. [Google Scholar] [CrossRef] [Green Version]
- Krissansen, G.W. Emerging health properties of whey proteins and their clinical implications. J. Am. Coll. Nutr. 2007, 26, 713S–723S. [Google Scholar] [CrossRef]
- Sprong, R.; Schonewille, A.; Van der Meer, R. Dietary cheese whey protein protects rats against mild dextran sulfate sodium–induced colitis: Role of mucin and microbiota. J. Dairy Sci. 2010, 93, 1364–1371. [Google Scholar] [CrossRef]
- Teixeira, F.J.; Santos, H.O.; Howell, S.L.; Pimentel, G.D. Whey protein in cancer therapy: A narrative review. Pharmacol. Res. 2019, 144, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.; Dupont, C.; Lemieux, P. Whey proteins and peptides: Beneficial effects on immune health. Clin. Pract. 2006, 3, 69. [Google Scholar] [CrossRef] [PubMed]
- Phull, A.-R.; Nasir, B.; ul Haq, I.; Kim, S.J. Oxidative stress, consequences and ROS mediated cellular signaling in rheumatoid arthritis. Chem. Biol. Interact. 2018, 281, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sui, Y.; Jiang, R.; Niimi, M.; Hong, J.; Yan, Q.; Shi, Z.; Yao, J. Development of Dietary Thiol Antioxidant via Reductive Modification of Whey Protein and Its Application in the Treatment of Ischemic Kidney Injury. Antioxidants 2023, 12, 193. https://doi.org/10.3390/antiox12010193
Sui Y, Jiang R, Niimi M, Hong J, Yan Q, Shi Z, Yao J. Development of Dietary Thiol Antioxidant via Reductive Modification of Whey Protein and Its Application in the Treatment of Ischemic Kidney Injury. Antioxidants. 2023; 12(1):193. https://doi.org/10.3390/antiox12010193
Chicago/Turabian StyleSui, Yang, Rui Jiang, Manabu Niimi, Jingru Hong, Qiaojing Yan, Zhuheng Shi, and Jian Yao. 2023. "Development of Dietary Thiol Antioxidant via Reductive Modification of Whey Protein and Its Application in the Treatment of Ischemic Kidney Injury" Antioxidants 12, no. 1: 193. https://doi.org/10.3390/antiox12010193