
Salinity Stress Ameliorates Pigments, Minerals, Polyphenolic Profiles, and Antiradical Capacity in Lalshak

 ,
,  ,
,  ,
, .jpg) and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Experimental Site, Plant Materials, and Experimental Conditions
2.2. Chemicals
2.3. Determination of Mineral Composition
2.4. Determination of Chlorophylls and Carotenoids
2.5. Betacyanins and Betaxanthins Content Measurement
2.6. Estimation of BC
2.7. Estimation of AsA
2.8. Sample Extraction and Determination of Folin–Ciocalteu Reducing Capacity, TF, and ARC
2.9. Sample Extraction and Determination of Polyphenolic Profiles by HPLC
2.10. Quantification of Polyphenolic Compounds
2.11. Statistical Analysis
3. Results and Discussion
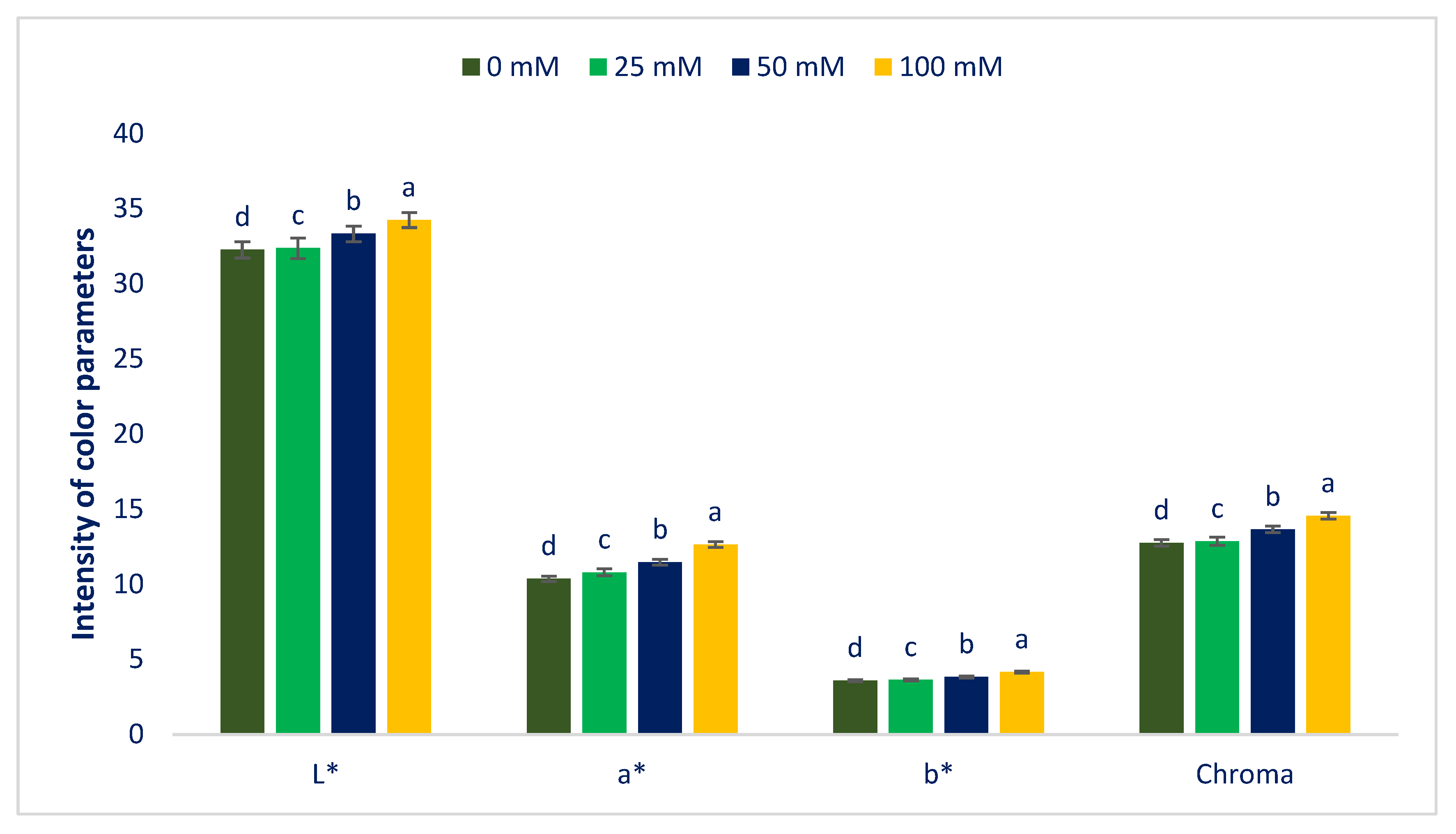
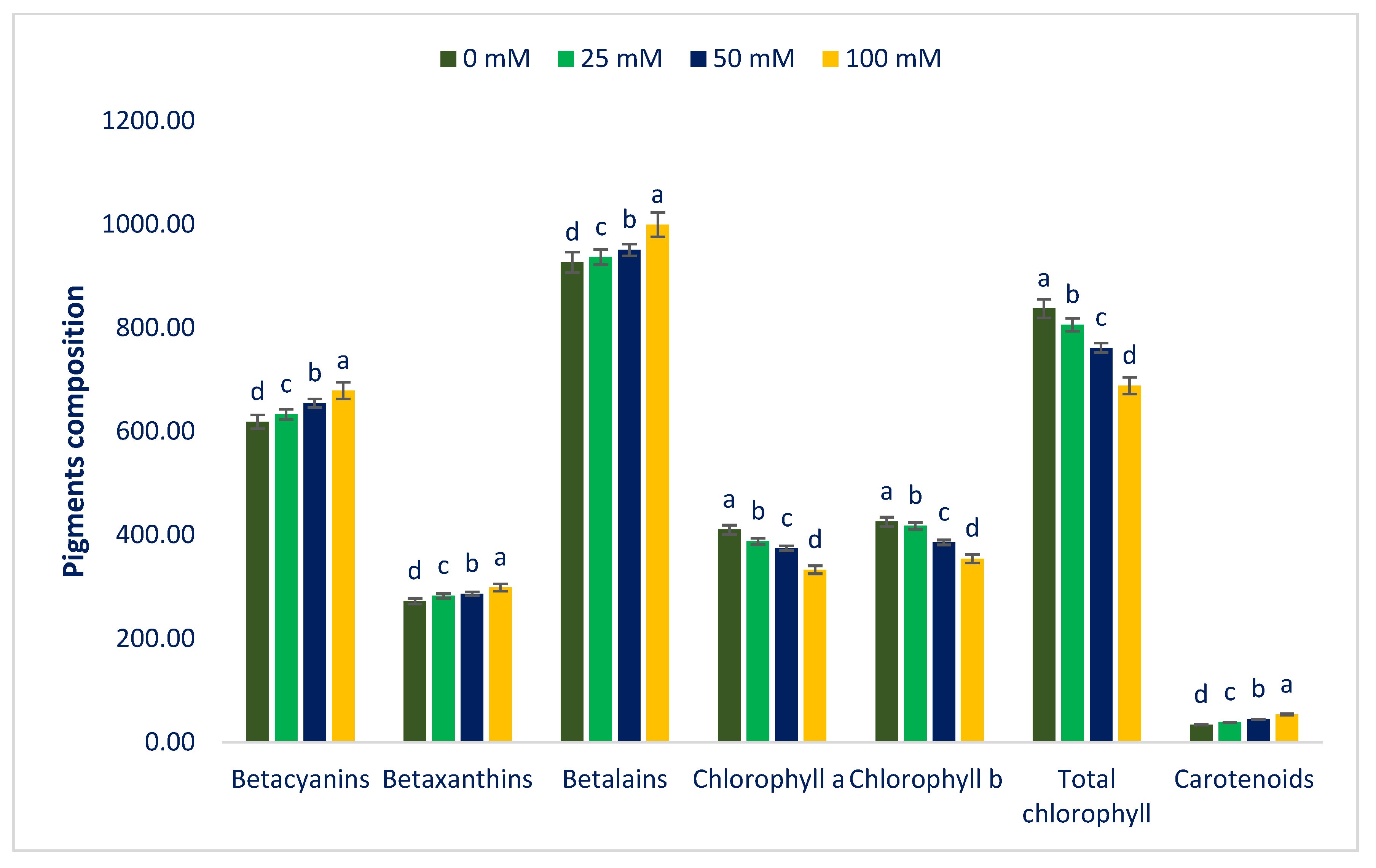
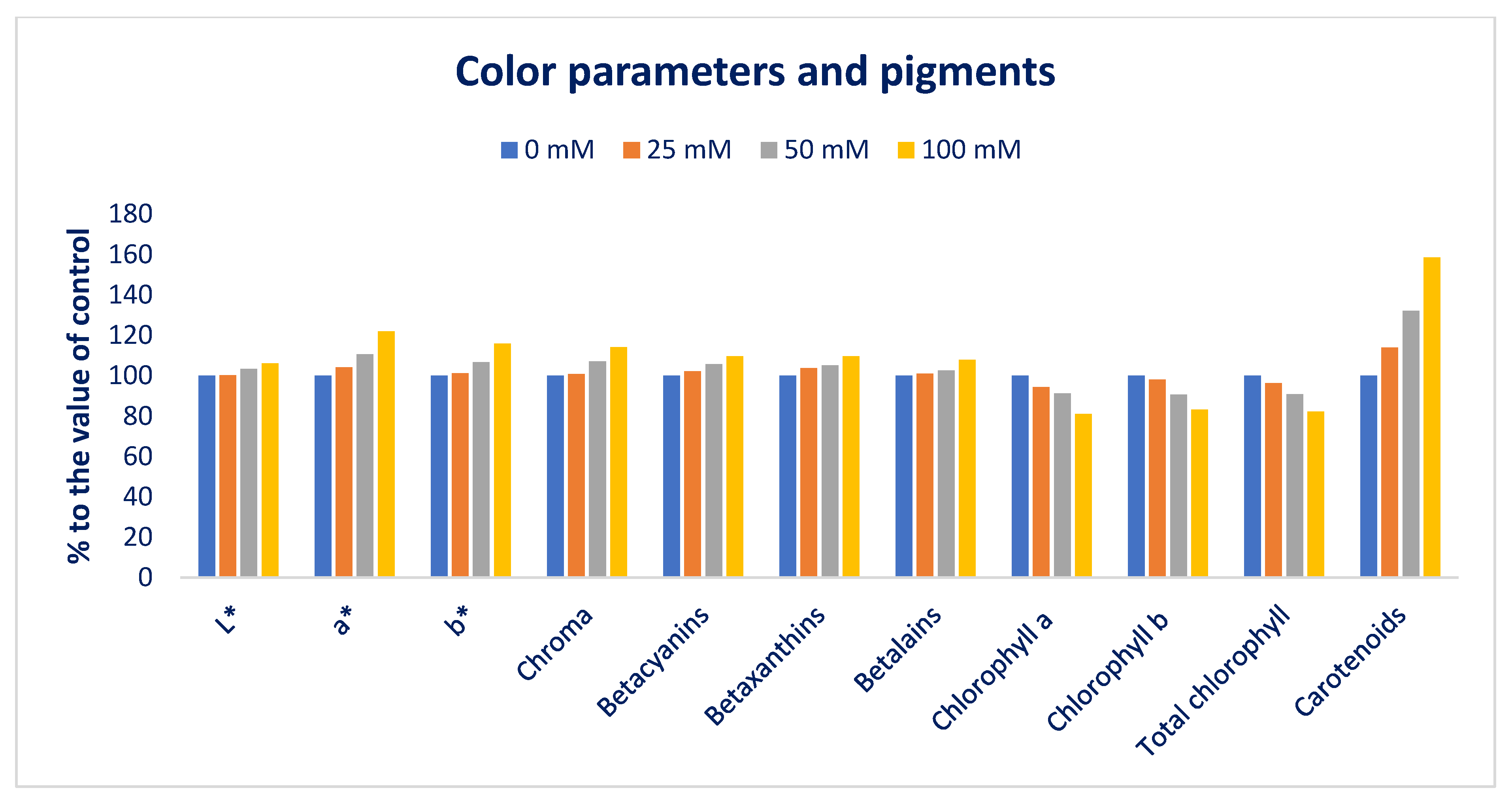
3.1. Influence of Sodium Chloride Stress on Color Parameters and Pigments
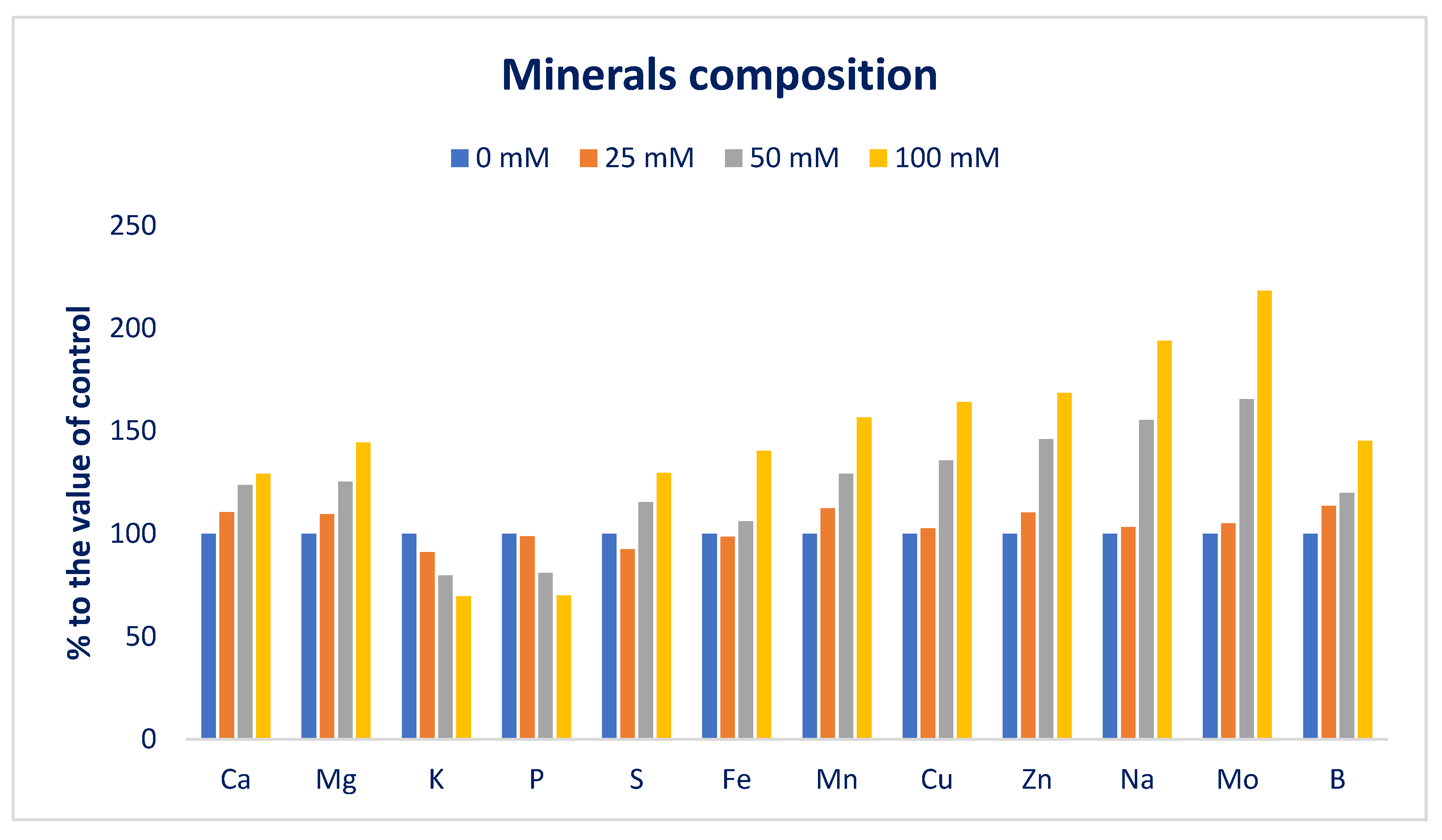
3.2. Sodium Chloride Impact on Minerals (Macroelements and Microelements)
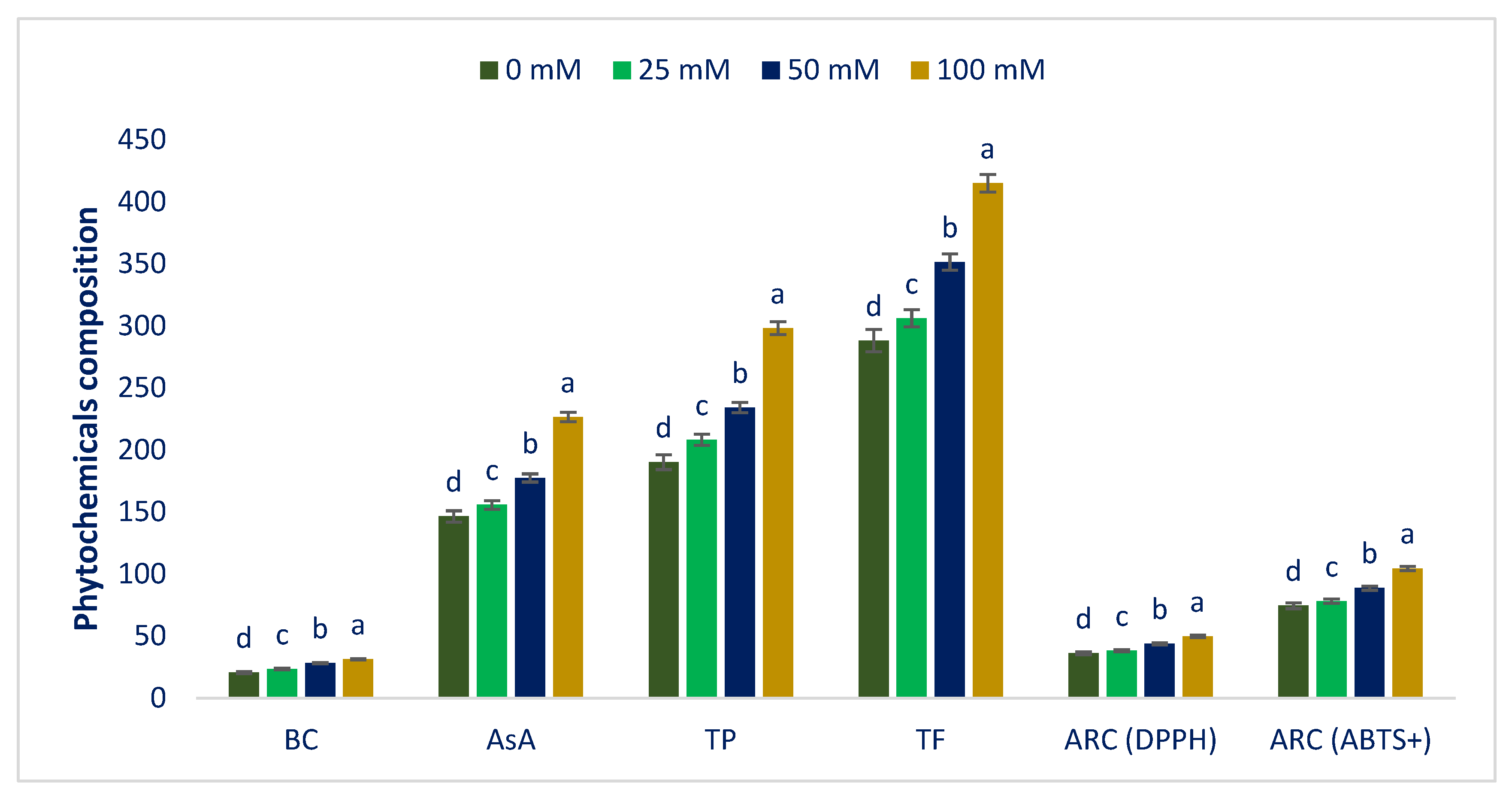
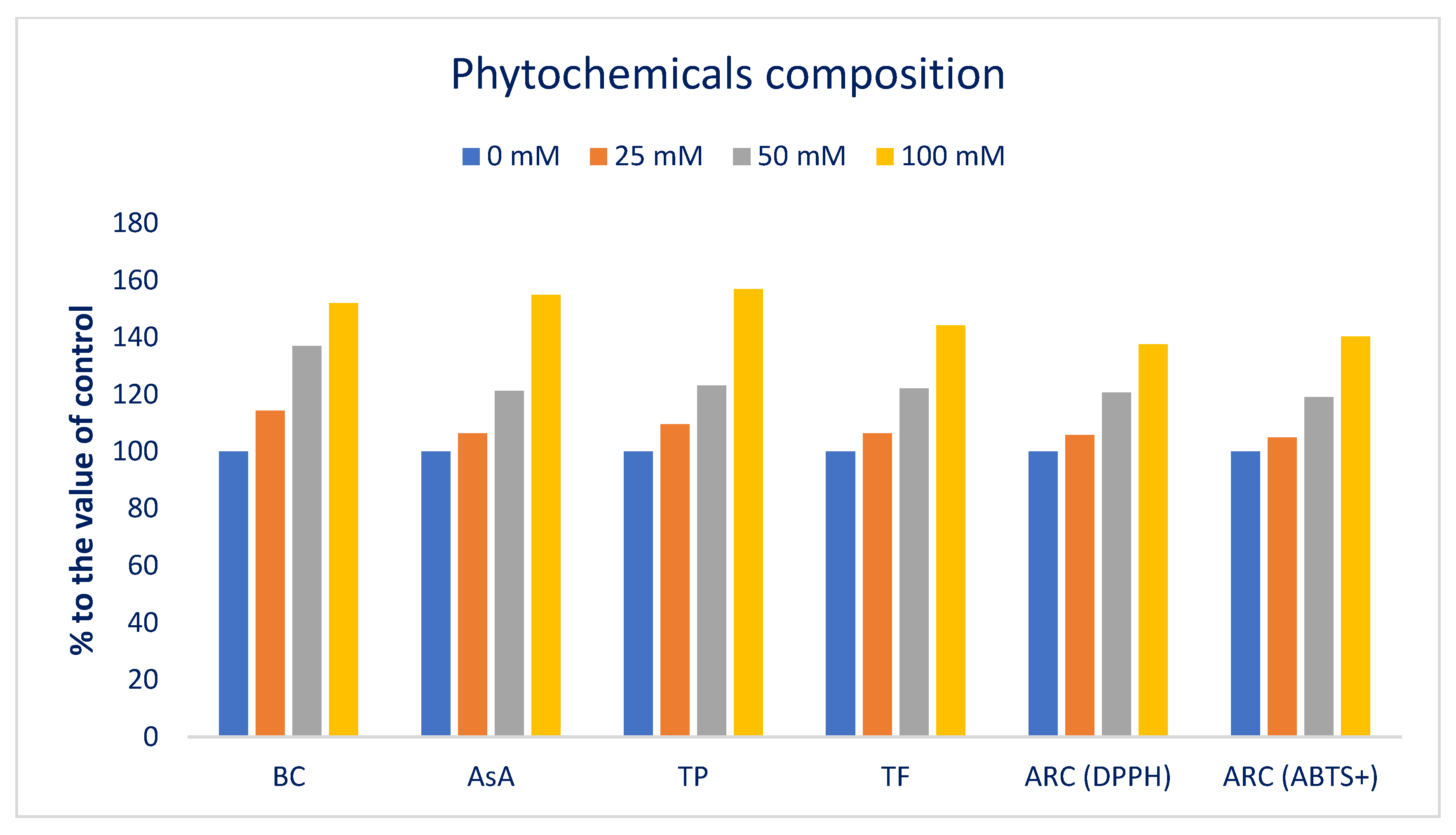
3.3. Influence of Sodium Chloride on Phytochemicals
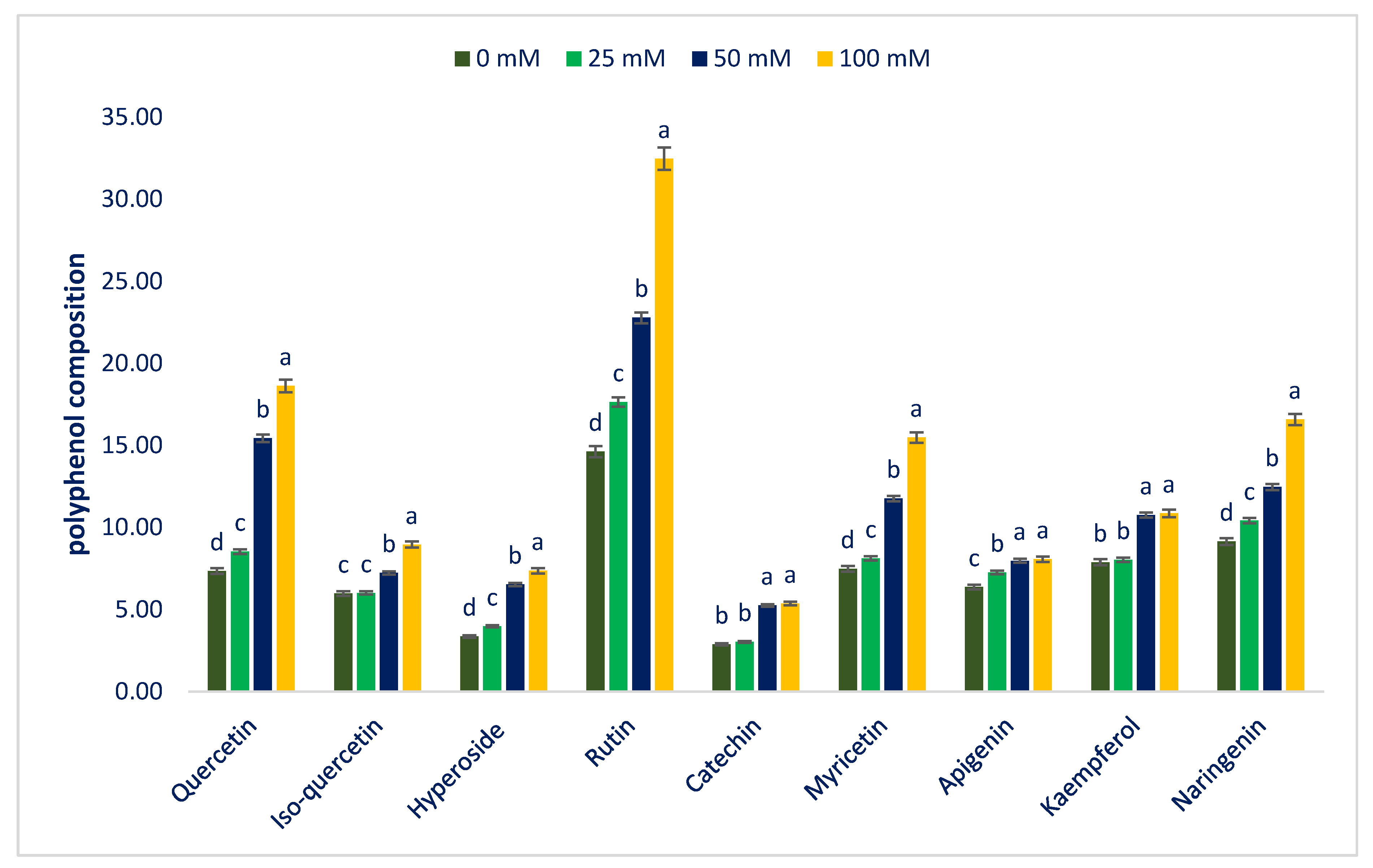
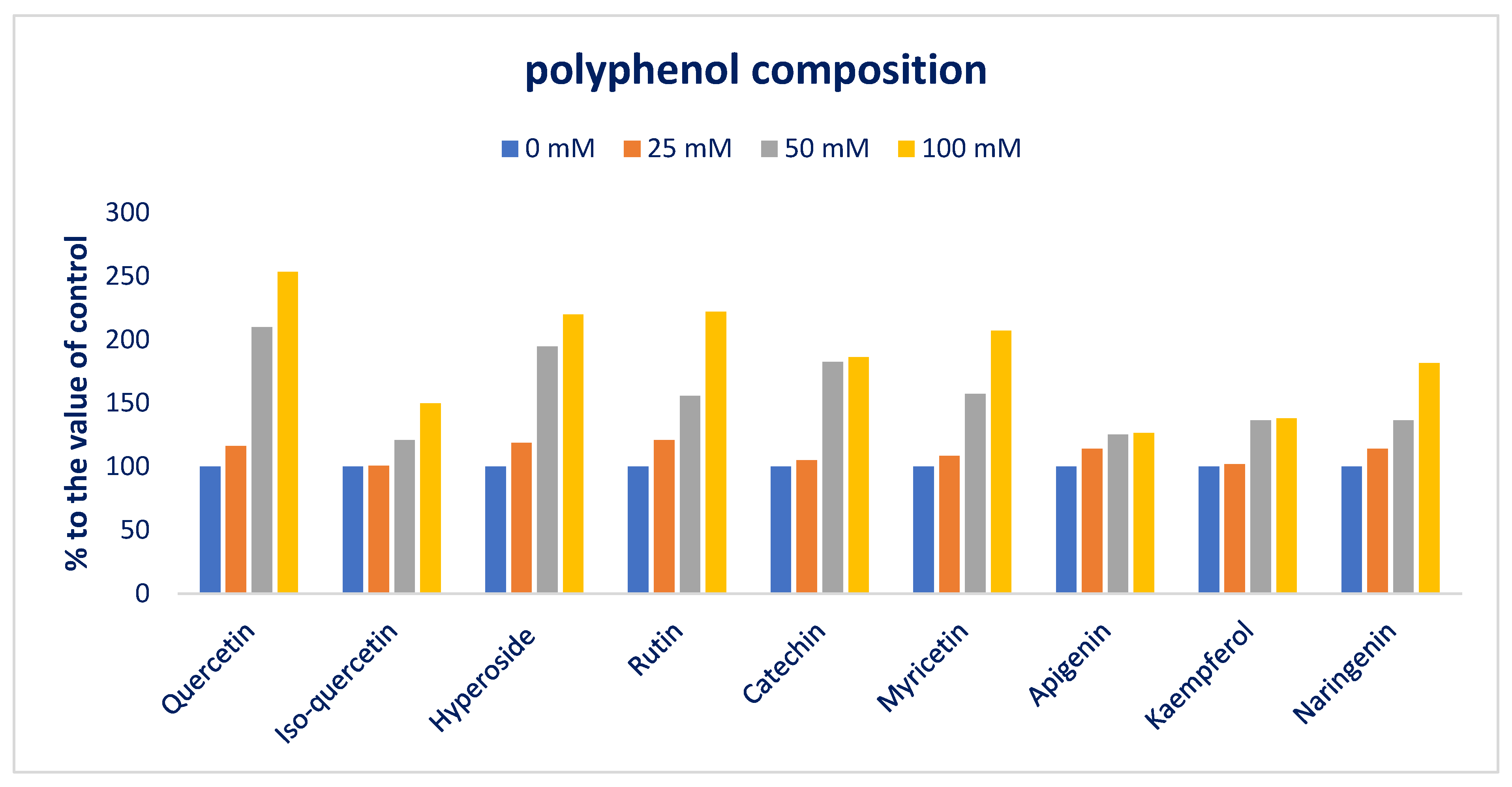
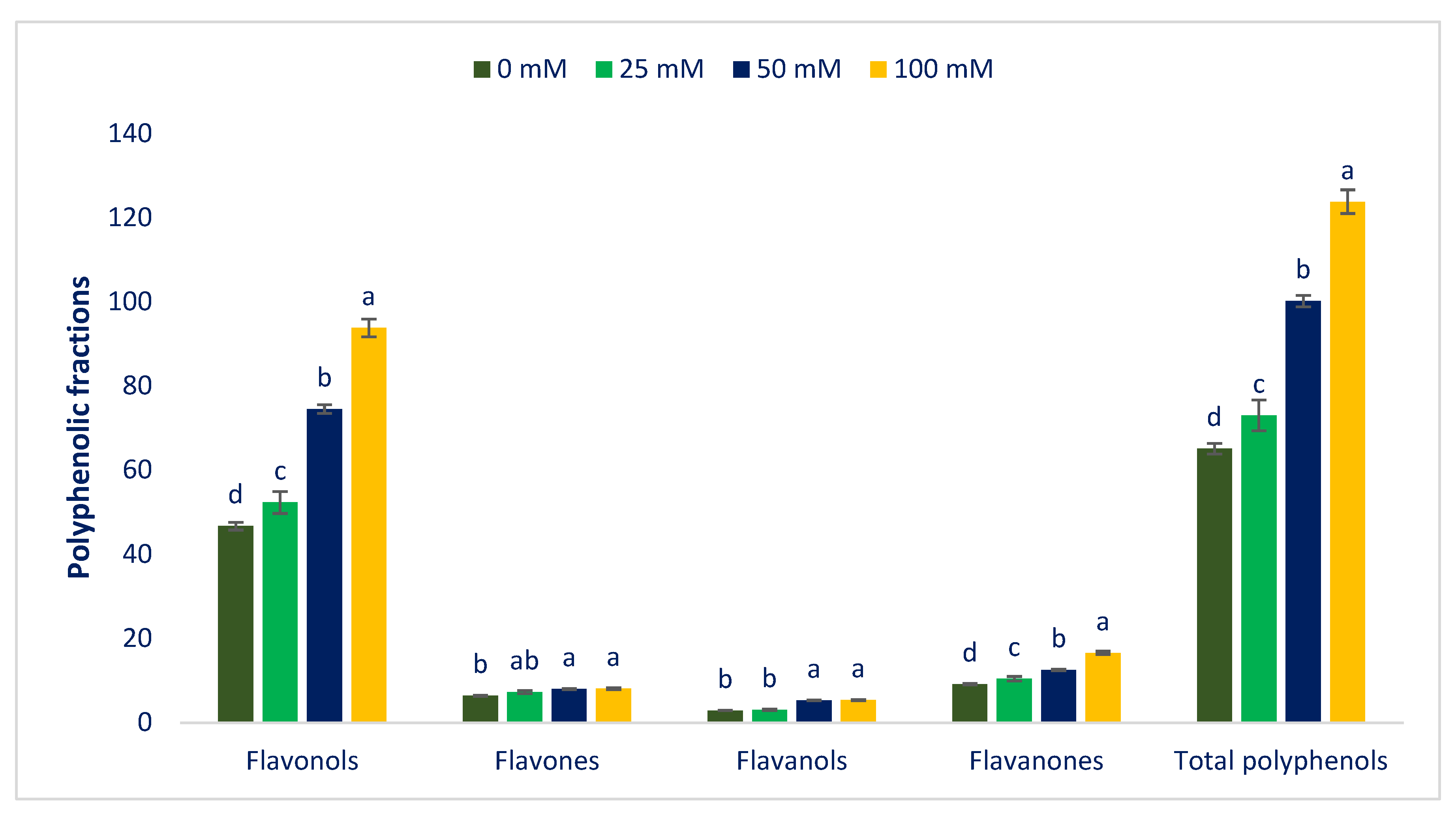
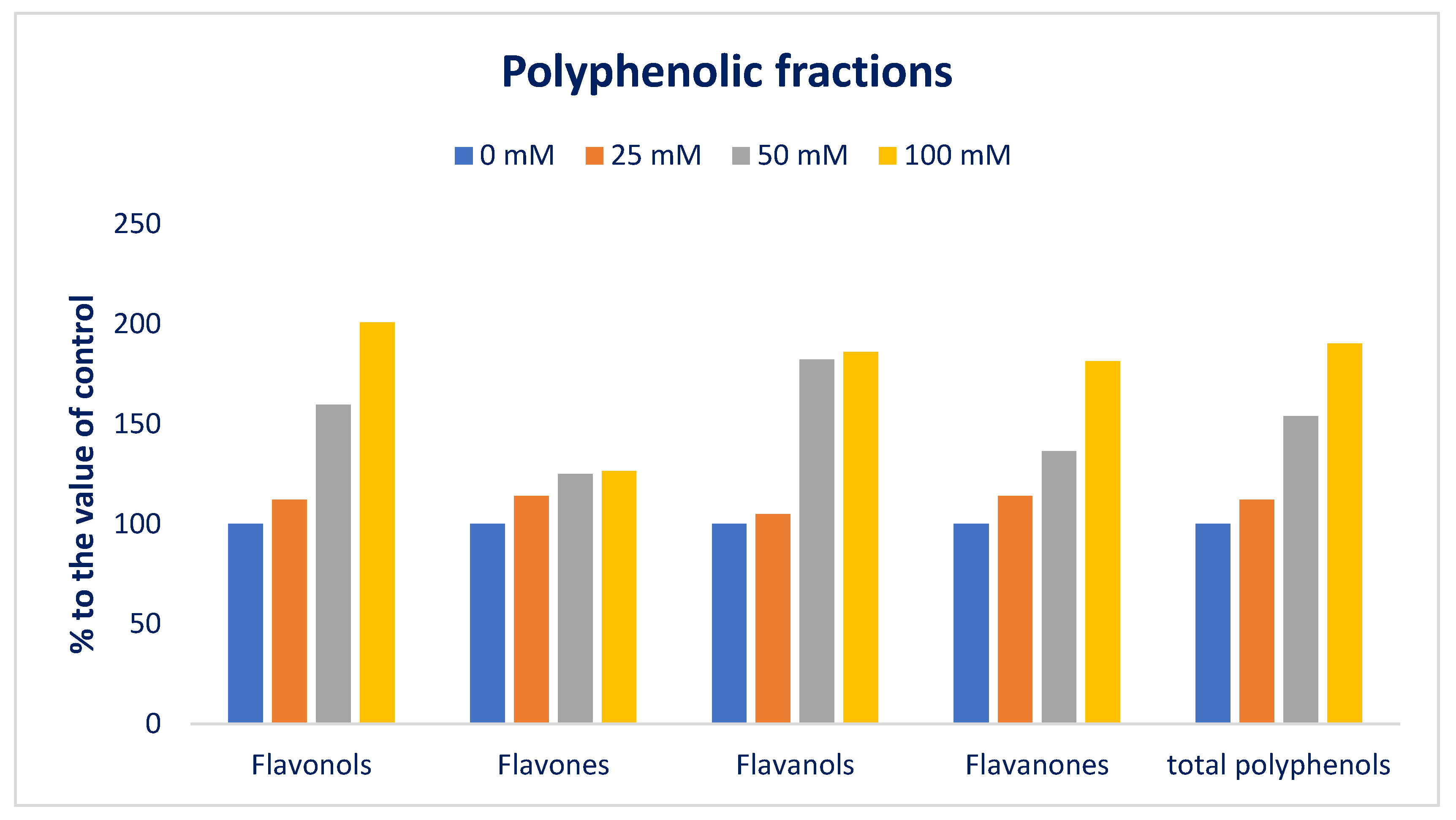
3.4. Response of Sodium Chloride Stress on Polyphenolic Compounds
3.5. The Coefficient of Correlation Study
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, Y.; Sun, M.; Corke, H. Antioxidant Activity of Betalains from Plants of the Amaranthaceae. J. Agril. Food Chem. 2003, 51, 2288–2294. [Google Scholar] [CrossRef]
- Stintzing, F.C.; Carle, R. Betalains—Emerging prospects for food scientists. Trends Food Sci. Technol. 2007, 18, 514–525. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Wagner, A.E.; Schini-Kerth, V.B. Rimbach, G. Betanin—A food colorant with biological activity. Mol. Nutr. Food Res. 2015, 59, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Bartosz, G. Biological Properties and Applications of Betalains. Molecules 2021, 26, 2520. [Google Scholar] [CrossRef] [PubMed]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Carotenoids: Potential allies of cardiovascular health? Food Nutr. Res. 2015, 59, 26762. [Google Scholar]
- Islam, F.; Muni, M.; Mitra, S.; Emran, T.B.; Chandran, D.; Das, R.; Rauf, A.; Safi, S.Z.; Chidambaram, K.; Dhawan, M.; et al. Recent advances in respiratory diseases: Dietary carotenoids as choice of therapeutics. Biomed. Pharmacother. 2022, 155, 113786. [Google Scholar] [CrossRef]
- Isabelle, M.; Lee, B.L.; Lim, M.T.; Koh, W.P.; Huang, D.; Ong, C.N. Antioxidant activity and profiles of common fruits in Singapore. Food Chem. 2010, 123, 77–84. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Corke, H. HPLC Characterization of Betalains from Plants in the Amaranthaceae. J. Chromatogr. Sci. 2005, 43, 454–460. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Leaf pigmentation, its profiles and radical scavenging activity in selected Amaranthus tricolor leafy vegetables. Sci. Rep. 2020, 10, 18617. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Color attributes, betacyanin, and carotenoid profiles, bioactive components, and radical quenching capacity in selected Amaranthus gangeticus leafy vegetables. Sci. Rep. 2021, 11, 11559. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Wagner, A.E.; Motafakkerazad, R.; Nakajima, Y.; Matsugo, S.; Rimbach, G. Free Radical Scavenging and Antioxidant Activity of Betanin: Electron Spin Resonance Spectroscopy Studies and Studies in Cultured Cells. Food Chem. Toxicol. 2014, 73, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. Structural Implications on Color, Fluorescence, and Antiradical Activity in Betalains. Planta 2010, 232, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Peiretti, P.G.; Meineri, G.; Gai, F.; Longato, E.; Amarowicz, R. Antioxidative Activity and Phenolic Compounds of Pumpkin (Cucurbita pepo) Seeds and Amaranth (Amaranthus caudatus) Grain Extracts. Nat. Prod. Res. 2017, 31, 2178–2182. [Google Scholar] [CrossRef] [PubMed]
- Stintzing, F.C.; Kammerer, D.; Schieber, A.; Adama, H.; Nacoulma, O.G.; Carle, R. Betacyanins and Phenolic Compounds from Amaranthus spinosus L. and Boerhavia erecta L. Z. Naturforsch. C 2004, 59, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Xiao, Y.; Tang, Z.; Jin, W.; Wang, Y.; Chen, H.; Yao, H.; Shan, Z.; Bu, T.; Wang, X. Extraction of Polysaccharides from Amaranthus hybridus L. by Hot Water and Analysis of Their Antioxidant Activity. PeerJ 2019, 7, e7149. [Google Scholar] [CrossRef] [Green Version]
- Zeashan, H.; Amresh, G.; Singh, S.; Rao, C.V. Hepatoprotective and Antioxidant Activity of Amaranthus spinosus Against CCl4 Induced Toxicity. J. Ethnopharmacol. 2009, 125, 364–366. [Google Scholar] [CrossRef]
- Karamac, M.; Gai, F.; Longato, E.; Meineri, G.; Janiak, M.A.; Amarowicz, R.; Peiretti, P.G. Antioxidant Activity and Phenolic Composition of Amaranth (Amaranthus caudatus) During Plant Growth. Antioxidants 2019, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Kraujalis, P.; Venskutonis, P.R.; Kraujalienė, V.; Pukalskas, A. Antioxidant properties and preliminary evaluation of phytochemical composition of different anatomical parts of amaranth. Plant Foods Hum. Nutr. 2013, 68, 322–328. [Google Scholar] [CrossRef]
- Repo-Carrasco-Valencia, R.; Peña, J.; Kallio, H.; Salminen, S. Dietary Fiber and Other Functional Components in Two Varieties of Crude and Extruded Kiwicha (Amaranthus caudatus). J. Cereal Sci. 2009, 49, 219–224. [Google Scholar] [CrossRef]
- Amornrit, W.; Santiyanont, R. Neuroprotective Effect of Amaranthus lividus and Amaranthus tricolor and Their Effects on Gene Expression of RAGE During Oxidative Stress in SH-SY5Y Cells. Genet. Mol. Res. 2016, 15, gmr15027562. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Hanif, S.; Iftikhar, T. Phytochemical Profiling with Antioxidant and Antimicrobial Screening of Amaranthus viridis L. Leaf and Seed Extracts. Open J. Med. Microbiol. 2013, 3, 16–171. [Google Scholar]
- Vardhana, H. In vitro Antibacterial Activity of Amaranthus spinosus Root Extracts. Pharmacophore. 2011, 2, 266–270. [Google Scholar]
- Guo, L.; Wang, Y.; Bi, X.; Duo, K.; Sun, Q.; Yun, X.; Zhang, Y.; Fei, P.; Han, J. Antimicrobial Activity and Mechanism of Action of the Amaranthus tricolor Crude Extract Against Staphylococcus aureus and Potential Application in Cooked Meat. Foods 2020, 9, 359. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.J.; Pong, L.Y.; Hassan, S.S.; Choo, W.S. Antiviral Activity of Betacyanins from Red Pitahaya (Hylocereus polyrhizus) and Red Spinach (Amaranthus dubius) against Dengue Virus Type 2 (GenBank accession no. MH488959). Access Microbiol. 2020, 2, acmi000073. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Z.; Amresh, G.; Singh, S.; Rao, C.V. Antidiarrheal and Antiulcer Activity of Amaranthus spinosus in Experimental Animals. Pharm. Biol. 2009, 47, 932–939. [Google Scholar] [CrossRef] [Green Version]
- Reyad-ul-Ferdous, M.; Shahjahan, D.S.; Tanvir, S. Present Biological Status of Potential Medicinal Plant of Amaranthus viridis: A comprehensive review. Am. Mukti M.J. Clin. Exp. Med. 2015, 3, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Baral, M.; Chakraborty, S.; Chakraborty, P. Evaluation of Anthelmintic and Anti-Inflammatory Activity of Amaranthus spinosus L. Int. J. Curr. Pharm. Res. 2010, 2, 2–5. [Google Scholar]
- Kumar, B.S.A.; Lakshman, K.; Jayaveera, K.N.; Velmurugan, C.; Manoj, B.; Sridhar, S.M. Anthelmintic Activity of Methanol Extract of Amaranthus caudatus L. Internet J. Food Saf. 2010, 12, 127–129. [Google Scholar]
- Kumar, A.; Lakshman, K.; Jayaveera, K.N.; Nandeesh, R.; Manoj, B.; Ranganayakulu, D. Comparative in vitro Anthelmintic Activity of Three Plants from Amaranthaceae Family. Archive Biol. Sci. 2010, 62, 185–189. [Google Scholar]
- Zeashan, H.; Amresh, G.; Singh, S.; Rao, C.V. Hepatoprotective Activity of Amaranthus spinosus in Experimental Animals. Food Chem. Toxicol. 2008, 46, 3417–3421. [Google Scholar] [CrossRef]
- Aneja, S.; Vats, M.; Aggarwal, S.; Sardana, S. Phytochemistry and Hepatoprotective Activity of Aqueous Extract of Amaranthus tricolor L. Roots. J. Ayurveda Integr. Med. 2013, 4, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeashan, H.; Amresh, G.; Singh, S.; Rao, C.V. Protective Effect of Amaranthus spinosus Against D-Galactosamine/Lipopolysaccharide-Induced Hepatic Failure. Pharm. Biol. 2010, 48, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.S.A.; Lakshman, K.; Kumar, P.A.A.; Viswantha, G.L.S.; Veerapur, V.P.; Thippeswamy, B.S.; Manoj, B. Hepatoprotective Activity of Methanol Extract of Amaranthus caudatus Linn. Against Paracetamol-Induced Hepatic Injury in Rats. J. Chin. Integr. Med. 2011, 9, 194–200. [Google Scholar] [CrossRef]
- House, N.C.; Puthenparampil, D.; Malayil, D.; Narayanankutty, A. Variation in The Polyphenol Composition, Antioxidant, and Anticancer Activity Among Different Amaranthus Species. S. Afr. J. Bot. 2020, 135, 408–412. [Google Scholar] [CrossRef]
- Amornrit, W.; Santiyanont, R. Effect of Amaranthus on Advanced Glycation End-Products Induced Cytotoxicity and Proinflammatory Cytokine Gene Expression in SH-SY5Y Cells. Molecules 2015, 20, 17288. [Google Scholar] [CrossRef] [Green Version]
- Olajide, O.; Ogunleye, B.; Erinle, T. Anti-inflammatory Properties of Amaranthus spinosus Leaf Extract. Pharm. Biol. 2004, 42, 521–525. [Google Scholar] [CrossRef]
- Jin, Y.; Xuan, Y.; Chen, M.; Chen, J.; Jin, Y.; Piao, J.; Tao, J. Antioxidant, Anti-inflammatory and Anticancer Activities of Amaranthus viridis L. Extracts. Asian J. Chem. 2013, 25, 8901–8904. [Google Scholar] [CrossRef]
- Allegra, M.; Tesoriere, L.; Livrea, M.A. Betanin Inhibits the Myeloperoxidase/Nitrite-Induced Oxidation of Human Low-Density Lipoproteins. Free Radic. Res. 2007, 41, 335–341. [Google Scholar] [CrossRef]
- Balakrishnan, S.; Pandhare, R. Antihyperglycemic and Antihyperlipidemic Activities of Amaranthus spinosus L. Extract on Alloxan Induced Diabetic Rats. Malays. J. Pharm. Sci. 2010, 8, 13–22. [Google Scholar]
- Krishnamurthy, G.; Lakshman, K.; Pruthvi, N.; Chandrika, P.U. Antihyperglicemic and Hypolipidemic Activity of Methanolic Extract of Amaranthus viridis Leaves in Experimental Diabetes. Indian J. Pharmacol. 2011, 43, 450–454. [Google Scholar]
- Hilou, A.; Nacoulma, O.G.; Guiguemde, T.R. In vivo Antimalarial Activities of Extracts from Amaranthus spinosus L. and Boerhaavia erecta L. in Mice. J. Ethnopharmacol. 2006, 103, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, L.W.; Tsay, G.J.; Mong, M.C.; Liu, W.H.; Yin, M.C. Aqueous Extract Prepared from Steamed Red Amaranth (Amaranthus gangeticus L.) Leaves Protected Human Lens Cells Against High Glucose Induced Glycative and Oxidative Stress. J. Food Sci. 2021, 86, 3686–3697. [Google Scholar] [CrossRef] [PubMed]
- Prajitha, V.; Thoppil, J.E. Cytotoxic and Apoptotic Activities of Extract of Amaranthus spinosus L. in Allium cepa and Human Erythrocytes. Cytotechnology 2017, 69, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, A.; Ghazanfar, A. Possible involvement of some secondary metabolites in salt tolerance of sugarcane. J. Plant Physiol. 2006, 163, 723–730. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress effects on growth, ROS markers, compatible solutes, phenolics, flavonoids, and antioxidant activity in Amaranthus tricolor. Appl. Biochem. Biotechnol. 2018, 186, 999–1016. [Google Scholar] [CrossRef]
- Sarker, U.; Islam, M.T.; Oba, S. Salinity Stress Accelerates Nutrients, Dietary Fiber, Minerals, Phytochemicals and Antioxidant Activity in Amaranthus tricolor Leaves. PLoS ONE 2018, 13, 0206388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Oba, S. The Response of Salinity Stress-Induced A. tricolor to Growth, Anatomy, Physiology, Non-Enzymatic and Enzymatic Antioxidants. Front. Plant Sci. 2020, 11, 559876. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Response of Nutrients, Minerals, Antioxidant Leaf Pigments, Vitamins, Polyphenol, Flavonoid and Antioxidant Activity in Selected Vegetable Amaranth under Four Soil Water Content. Food Chem. 2018, 252, 72–83. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought Stress Enhances Nutritional and Bioactive Compounds, Phenolic Acids and Antioxidant Capacity of Amaranthus Leafy Vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [Green Version]
- Selmar, D.; Kleinwachter, M. Influencing the product quality by deliberately applying drought stress during the cultivation of medicinal plants. Industrial Crop Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Levizou, E.; Ntatsi, G.; Fernandes, Â.; Petrotos, K.; Akoumianakis, K.; Barros, L.; Ferreira, I.C. Salinity effect on nutritional value, chemical composition, and bioactive compounds content of Cichorium spinosum L. Food Chem. 2017, 214, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.H.; Park, K.J.; Kim, B.K.; Jeong, J.W.; Kim, H.J. Effect of salinity stress on phenolic compounds and carotenoids in buckwheat (Fagopyrum esculentum M.) sprout. Food Chem. 2012, 135, 1065–1070. [Google Scholar] [CrossRef]
- Alam, M.A.; Juraimi, A.S.; Rafii, M.Y.; Hamid, A.A.; Aslani, F.; Alam, M.Z. Effects of salinity and salinity-induced augmented bioactive compounds in purslane (Portulaca oleracea L.) for possible economical use. Food Chem. 2015, 169, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.M.; Cao, F.; Han, Y.; Nadira, U.A.; Zhang, G.; Wu, F. Differential changes in grain ultrastructure, amylase, protein and amino acid profiles between Tibetan wild and cultivated barleys under drought and salinity alone and combined stress. Food Chem. 2013, 141, 2743–2750. [Google Scholar] [CrossRef]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Genotypic variability for nutrient, antioxidant, yield and yield contributing traits in vegetable amaranth. J. Food Agri. Environ. 2014, 12, 168–174. [Google Scholar]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Variability, heritability and genetic association in vegetable amaranth (Amaranthus tricolor). Span. J. Agric. Res. 2015, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Genotype variability in composition of antioxidant vitamins and minerals in vegetable amaranth. Genetika 2015, 47, 85–96. [Google Scholar] [CrossRef]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Genetic variation and interrelationship among antioxidant, quality and agronomic traits in vegetable amaranth. Turk. J. Agric. For. 2016, 40, 526–535. [Google Scholar] [CrossRef]
- Sarker, U.; Islam, M.T.; Rabbani, M.G.; Oba, S. Genotypic diversity in vegetable amaranth for antioxidant, nutrient and agronomic traits. Indian J. Genet. Pl. Breed. 2017, 77, 173–176. [Google Scholar] [CrossRef]
- Chakrabarty, T.; Sarker, U.; Hasan, M.; Rahman, M.M. Variability in mineral compositions, yield, and yield contributing traits of stem amaranth (Amaranthus lividus). Genetika 2018, 50, 995–1010. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Rabbani, M.G.; Oba, S.; Eldehna, W.M.; Al-Rashood, S.T.; Mostafa, N.M.; Eldahshan, O.A. Phytonutrients, Colorant Pigments, Phytochemicals, and Antioxidant Potential of Orphan Leafy Amaranthus Species. Molecules 2022, 27, 2899. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Azam, M.G.; Talukder, M.Z.A. Genetic Variation in Mineral Profiles, Yield Contributing Agronomic Traits, and Foliage Yield of Stem Amaranth. Genetika 2022, 54, 91–108. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Ercisli, S.; Assouguem, A.; Alotaibi, A.; Ullah, R. Bioactive Phytochemicals and Quenching Activity of Radicals in Selected Drought-Resistant Amaranthus tricolor Vegetable Amaranth. Antioxidants 2022, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S.; Alsanie, W.F.; Gaber, A. Characterization of Phytochemicals, Nutrients, and Antiradical Potential in Slim Amaranth. Antioxidants 2022, 11, 1089. [Google Scholar] [CrossRef]
- Sarker, U.; Ercisli, S. Salt Eustress Induction in Red Amaranth (Amaranthus gangeticus) Augments Nutritional, Phenolic Acids and Antiradical Potential of Leaves. Antioxidants 2022, 11, 2434. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.N.; Sarker, U.; Raihan, M.S.; Al-Huqail, A.A.; Siddiqui, M.H.; Oba, S. Influence of Salinity Stress on Color Parameters, Leaf Pigmentation, Polyphenol and Flavonoid Contents, and Antioxidant Activity of Amaranthus lividus Leafy Vegetables. Molecules 2022, 27, 1821. [Google Scholar] [CrossRef]
- Sarker, U.; Iqbal, M.A.; Hossain, M.N.; Oba, S.; Ercisli, S.; Muresan, C.C.; Marc, R.A. Colorant Pigments, Nutrients, Bioactive Components, and Antiradical Potential of Danta Leaves (Amaranthus lividus). Antioxidants 2022, 11, 1206. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutraceuticals, antioxidant pigments, and phytochemicals in the leaves of Amaranthus spinosus and Amaranthus viridis weedy species. Sci. Rep 2019, 9, 20413. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Protein, dietary fiber, minerals, antioxidant, pigments and phytochemicals, and antioxidant activity in selected red morph Amaranthus leafy vegetable. PLoS ONE 2019, 14, 0222517. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Nutraceuticals, phytochemicals, and radical quenching ability of selected drought-tolerant advance lines of vegetable amaranth. BMC Plant Biol. 2020, 20, 564. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutritional and bioactive constituents and scavenging capacity of radicals in Amaranthus hypochondriacus. Sci. Rep. 2020, 10, 19962. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S.; Daramy, M.A. Nutrients, minerals, antioxidant pigments and phytochemicals, and antioxidant capacity of the leaves of stem amaranth. Sci. Rep. 2020, 10, 3892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Hossain, M.M.; Oba, S. Nutritional and antioxidant components and antioxidant capacity in green morph Amaranthus leafy vegetable. Sci. Rep. 2020, 10, 1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarker, U.; Hossain, M.N.; Iqbal, M.A.; Oba, S. Bioactive components and radical scavenging activity in selected advance lines of salt-tolerant vegetable amaranth. Front. Nutr. 2020, 7, 587257. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Nutrients, minerals, pigments, phytochemicals, and radical scavenging activity in Amaranthus blitum leafy vegetables. Sci. Rep. 2020, 10, 3868. [Google Scholar] [CrossRef] [Green Version]
- Sarker, U.; Oba, S. Phenolic profiles and antioxidant activities in selected drought-tolerant leafy vegetable amaranth. Sci. Rep. 2020, 10, 18287. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Polyphenol and flavonoid profiles and radical scavenging activity in selected leafy vegetable Amaranthus gangeticus. BMC Plant Biol. 2020, 20, 499. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Antioxidant constituents of three selected red and green color Amaranthus leafy vegetable. Sci. Rep. 2019, 9, 18233. [Google Scholar] [CrossRef]
- Faysal, A.S.M.; Ali, L.; Azam, M.G.; Sarker, U.; Ercisli, S.; Golokhvast, K.S.; Marc, R.A. Genetic Variability, Character Association, and Path Coefficient Analysis in Transplant Aman Rice Genotypes. Plants 2022, 11, 2952. [Google Scholar] [CrossRef]
- Hassan, J.; Jahan, F.; Rajib, M.M.; Sarker, U.; Miyajima, I.; Ozaki, Y.; Ercisli, S.; Golokhvast, K.S.; Marc, R.A. Color and physiochemical attributes of pointed gourd (Trichosanthes dioica Roxb.) Influenced by modified atmosphere packaging and postharvest treatment during storage. Front. Plant Sci. 2022, 13, 1016324. [Google Scholar] [CrossRef]
- Kulsum, U.; Sarker, U.; Rasul, M.G. Genetic variability, heritability and interrelationship in salt-tolerant lines of T. Aman rice. Genetika 2022, 54, 761–776. [Google Scholar] [CrossRef]
- Hasan, M.J.; Kulsum, M.U.; Sarker, U.; Matin, M.Q.I.; Shahin, N.H.; Kabir, M.S.; Ercisli, S.; Marc, R.A. Assessment of GGE, AMMI, Regression, and Its Deviation Model to Identify Stable Rice Hybrids in Bangladesh. Plants 2022, 11, 2336. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; Sarker, U.; Swapan, M.A.H.; Raihan, M.S.; Oba, S.; Alamri, S.; Siddiqui, M.H. Combining Ability Analysis and Marker-Based Prediction of Heterosis in Yield Reveal Prominent Heterotic Combinations from Diallel Population of Rice. Agronomy 2022, 12, 1797. [Google Scholar] [CrossRef]
- Azam, M.G.; Sarker, U.; Hossain, M.A.; Iqbal, M.S.; Islam, M.R.; Hossain, M.F.; Ercisli, S.; Kul, R.; Assouguem, A.; AL-Huqail, A.A.; et al. Genetic Analysis in Grain Legumes [Vigna radiata (L.) Wilczek] for Yield Improvement and Identifying Heterotic Hybrids. Plants 2022, 11, 1774. [Google Scholar] [CrossRef] [PubMed]
- Hassan, J.; Rajib, M.; Rahman, M.; Sarker, U.; Akter, M.; Khan, M.; Azam, N.E.; Khandaker, S.; Khalid, F.; Rahman, G.K.; et al. Optimizing textile dyeing wastewater for tomato irrigation through physiochemical, plant nutrient uses and pollution load index of irrigated soil. Sci. Rep. 2022, 12, 10088. [Google Scholar] [CrossRef]
- Prodhan, M.M.; Sarker, U.; Hoque, M.A.; Biswas, M.S.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almutairi, M.H.; Mohamed, H.R.H.; Najda, A. Foliar Application of GA3 Stimulates Seed Production in Cauliflower. Agronomy 2022, 12, 1394. [Google Scholar] [CrossRef]
- Azad, A.K.; Sarker, U.; Ercisli, S.; Assouguem, A.; Ullah, R.; Almeer, R.; Sayed, A.A.; Peluso, I. Evaluation of Combining Ability and Heterosis of Popular Restorer and Male Sterile Lines for the Development of Superior Rice Hybrids. Agronomy 2022, 12, 965. [Google Scholar] [CrossRef]
- Hasan, M.J.; Kulsum, M.U.; Majumder, R.R.; Sarker, U. Genotypic Variability for Grain Quality Attributes in Restorer Lines of Hybrid Rice. Genetika 2020, 52, 973–989. [Google Scholar] [CrossRef]
- Hasan-Ud-Daula, M.; Sarker, U. Variability, Heritability, Character Association, and Path Coefficient Analysis in Advanced Breeding Lines of Rice (Oryza sativa L.). Genetika 2020, 52, 711–726. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Shimizu, S. Chlorophyll Metabolism in Higher Plants VI. Involvement of Peroxidase in Chlorophyll Degradation. Plant Cell Physiol. 1985, 26, 1291–1301. [Google Scholar]
- Parida, A.K.; Das, A.B.; Sanada, Y.; Mohanty, P. Effects of Salinity on Biochemical Components of the Mangrove, Aegiceras corniculatum. Aquat. Bot. 2004, 80, 77–87. [Google Scholar] [CrossRef]
- Jain, G.; Schwinn, K.E.; Gould, K.S. Functional Role of Betalains in Disphyma australe under Salinity Stress. Environ. Exp. Bot. 2015, 109, 131–140. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced Responses of Photosynthesis and Antioxidant Metabolism in Higher Plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Bhargava, A.; Chatterjee, A.; Srivastava, J.; Singh, N.; Singh, S.P. Mineral profile and variability in vegetable amaranth (Amaranthus tricolor). Plant Food. Hum. Nutri. 2006, 61, 23–28. [Google Scholar] [CrossRef]
- Madruga, M.S.; Camara, F.S. The chemical composition of “Multimistura” as a food supplement. Food Chem. 2000, 68, 41–44. [Google Scholar] [CrossRef]
- Shahidi, F.; Chavan, U.D.; Bal, A.K.; McKenzie, D.B. Chemical composition of beach pea (Lathyrus maritimus L.) plant parts. Food Chem. 1999, 64, 39–44. [Google Scholar] [CrossRef]
- Jimenez-Aguilar, D.M.; Grusak, M.A. Minerals, vitamin C, phenolics, flavonoids and antioxidant activity of Amaranthus leafy vegetables. J. Food Compos Anal. 2017, 58, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Menezes, R.V.; Azevedo Neto, A.D.; Oliveira Ribeiro, M.; Watanabe Cova, A.M. Growth and Contents of Organic and Inorganic Solutes in Amaranth under Salt Stress. Agropec. Trop. Goiania. 2017, 47, 22–30. [Google Scholar] [CrossRef]
- Odjegba, V.J.; Chukwunwike, I.C. Physiological Responses of Amaranthus hybridus L. under Salinity Stress. Indian J. Innov. Dev. 2012, 1, 742–748. [Google Scholar]
- Koksal, N.; Alkan-Torun, A.; Kulahlioglu, I.; Ertargin, E.; Karalar, E. Ion uptake of marigold under saline growth conditions. Springer Plus 2016, 5, 139. [Google Scholar] [CrossRef] [Green Version]
- Anower-Rokebul, M.; Mott, I.W.; Peel, M.D.; Wu, Y. Characterization of Physiological Responses of Two Alfalfa Half-sib Families with Improved Salt Tolerance. Plant Physiol. Biochem. 2013, 71, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Asgari, H.R.; Cornelis, W.; Van Damm, P. Salt Stress Effect on Wheat (Triticum aestivum L.) Growth and Leaf Ion Concentration. Int. J. Agron. Plant Prod. 2012, 6, 195–208. [Google Scholar]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M. Biotechnological Approach of Improving Plant Salt Tolerance Using Antioxidants as Markers. Biotech. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef]
- Khanam, U.K.S.; Oba, S.; Yanase, E.; Murakami, Y. Phenolic acids, flavonoids and total antioxidant capacity of selected leafy vegetables. J. Functional Foods. 2012, 4, 979–987. [Google Scholar] [CrossRef]
- Khanam, U.K.S.; Oba, S. Bioactive substances in leaves of two amaranth species, Amaranthus tricolor and A. hypochondriacus. Canadian J. Plant Sci. 2013, 93, 47–58. [Google Scholar] [CrossRef]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of Secondary Metabolites and Brassinosteroids in Plant Defense Against Environmental Stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Espinoza, A.; Martína, A.S.; Lopez-Climentb, M.; Ruiz-Laraa, S.; Gomez-Cadenasb, A.; Casarettoa, J. Engineered Drought-Induced Biosynthesis of α-Tocopherol Alleviates Stress-Induced Leaf Damage in Tobacco. J. Plant Physiol. 2013, 170, 1285–1294. [Google Scholar] [CrossRef]
- Lin, K.H.; Chao, P.Y.; Yang, C.M.; Cheng, W.C.; Lo, H.F.; Chang, T.R. The Effects of Flooding and Drought Stresses on The Antioxidant Constituents in Sweet Potato Leaves. Bot. Stud. 2006, 47, 417–426. [Google Scholar]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Goli, S.A.H. Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Appl. Biochem. Biotechnol. 2016, 178, 796–809. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
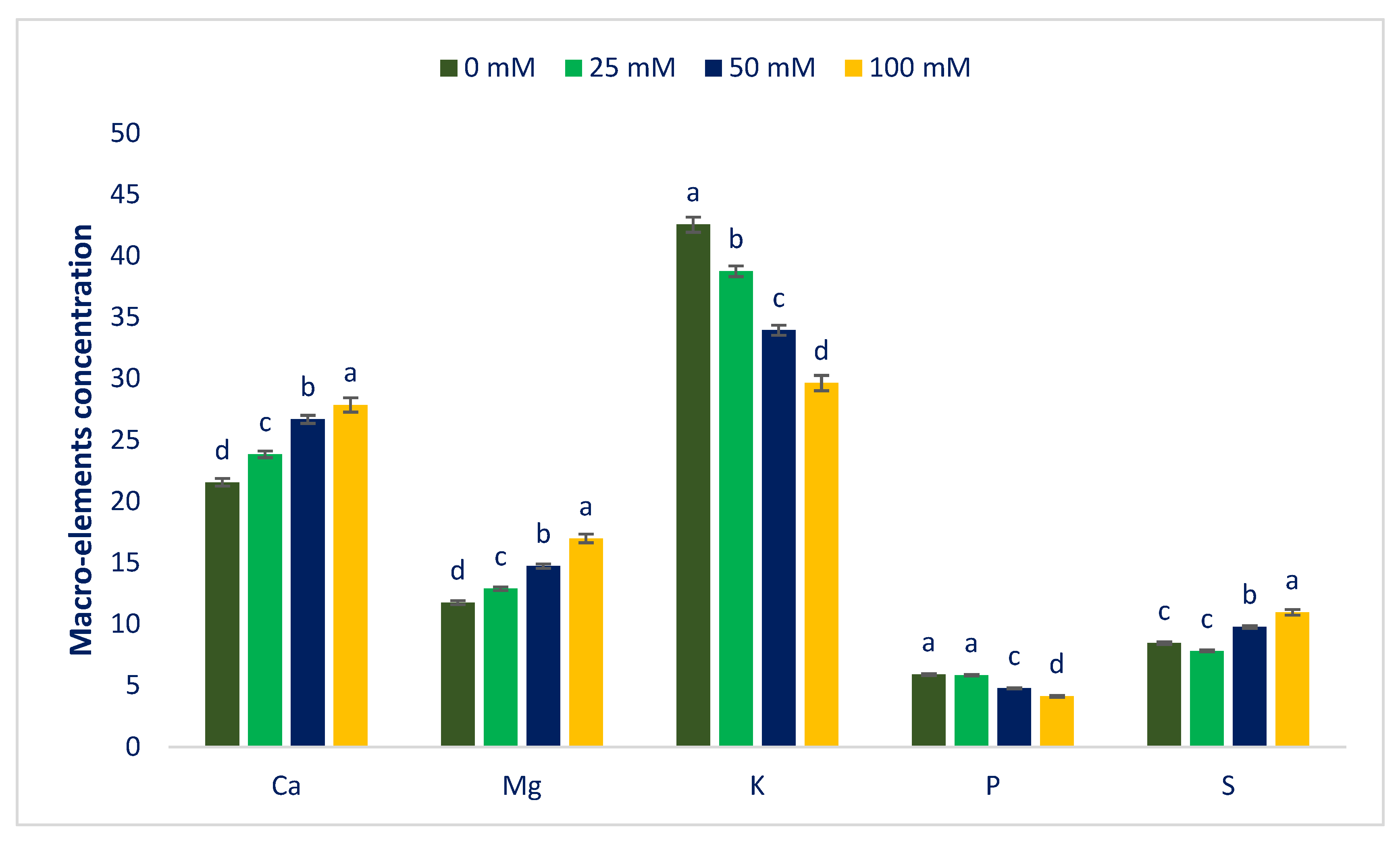{kind=link}
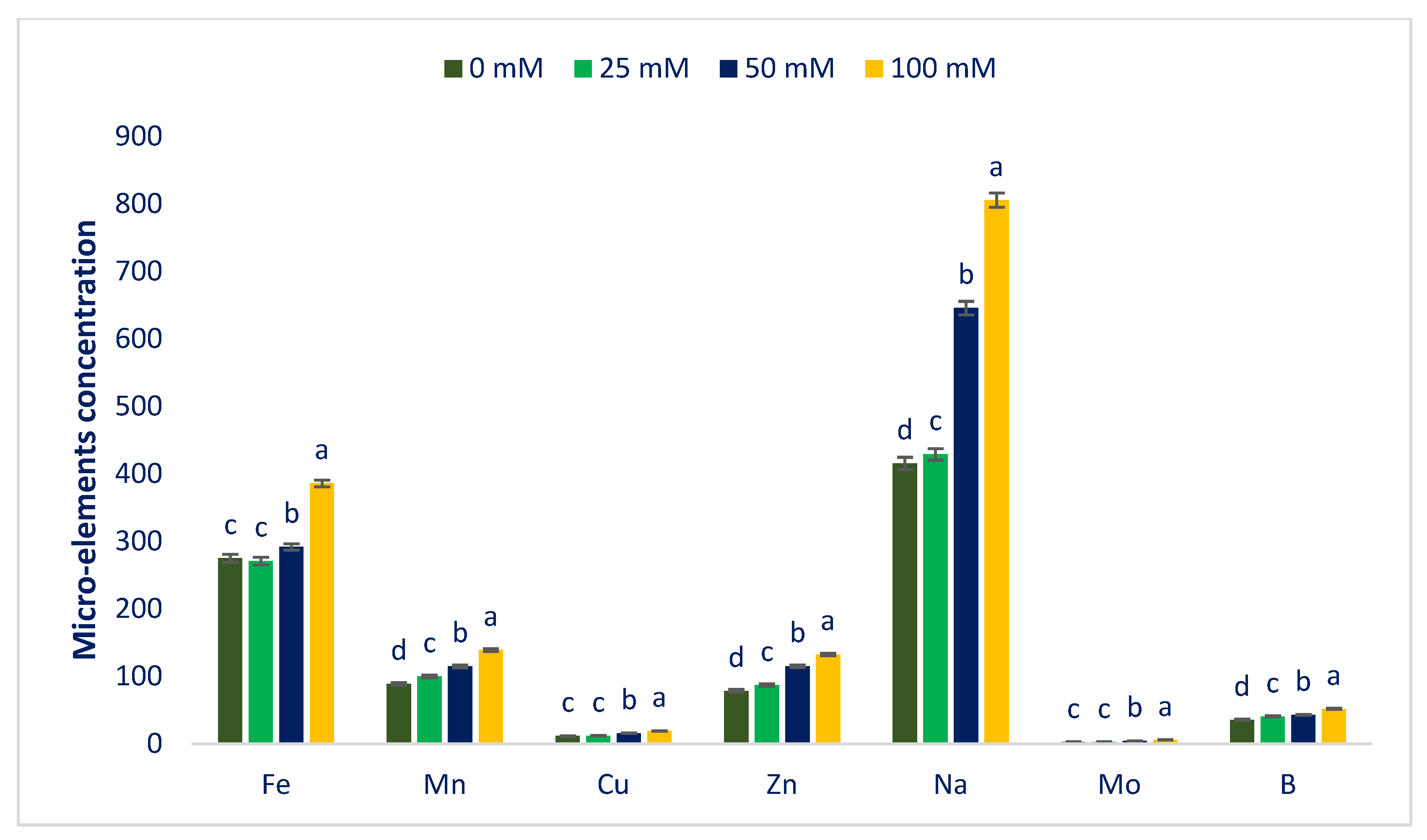{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt (min) | λmax (nm) | Molecular Ion [M − H]− (m/z) | MS2 (m/z) | Identity of Tentative Compounds |
|---|---|---|---|---|
| 4.6 | 370 | 626.2468 | 626.3216 | Myricetin-3-O-rutinoside |
| 7.5 | 370 | 301.0348 | 301.2267 | 2-(3,4-dihydroxy phenyl)-3,5,7-trihydroxychromene-4-one |
| 15.4 | 370 | 270.2344 | 270.3221 | 4′,5,7-Trihydroxyflavone, 5,7-Dihydroxy-2-(4-hydroxyphenyl)-4-benzopyrone |
| 17.8 | 370 | 593.4253 | 593.3687 | kaempferol-3-O-rutinoside |
| 23.9 | 280 | 290.2463 | 290.1238 | (2R-3S)-2-(3,4-dihydroxyphenyl)-3,4-dihydro-2-chromene-3,5,7-triol |
| 26.7 | 280 | 271.0622 | 271.2448 | Naringenin |
| 53.0 | 360 | 609.3874 | 609.4265 | Quercetin-3-O-rutinoside |
| 53.3 | 360 | 463.4358 | 463.5125 | Quercetin-3-O-galactoside |
| 54.3 | 360 | 463.2875 | 463.3124 | Quercetin-3-O-glucoside |
| AsA | FCRC | TF | ARC (DPPH) | ARC (ABTS+) | |
|---|---|---|---|---|---|
| BC | 0.82 ** | 0.81 ** | 0.96 ** | 0.79 * | 0.84 * |
| AsA | 0.88 ** | 0.91 ** | 0.92 ** | 0.96 ** | |
| FCRC | 0.81 * | 0.93 ** | 0.94 ** | ||
| TF | 0.82 * | 0.78 * | |||
| ARC (DPPH) | 0.97 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarker, U.; Hossain, M.N.; Oba, S.; Ercisli, S.; Marc, R.A.; Golokhvast, K.S. Salinity Stress Ameliorates Pigments, Minerals, Polyphenolic Profiles, and Antiradical Capacity in Lalshak. Antioxidants 2023, 12, 173. https://doi.org/10.3390/antiox12010173
Sarker U, Hossain MN, Oba S, Ercisli S, Marc RA, Golokhvast KS. Salinity Stress Ameliorates Pigments, Minerals, Polyphenolic Profiles, and Antiradical Capacity in Lalshak. Antioxidants. 2023; 12(1):173. https://doi.org/10.3390/antiox12010173
Chicago/Turabian StyleSarker, Umakanta, Md. Nazmul Hossain, Shinya Oba, Sezai Ercisli, Romina Alina Marc, and Kirill S. Golokhvast. 2023. "Salinity Stress Ameliorates Pigments, Minerals, Polyphenolic Profiles, and Antiradical Capacity in Lalshak" Antioxidants 12, no. 1: 173. https://doi.org/10.3390/antiox12010173