
Dietary Protein Levels Modulate the Antioxidant Capacity during Different Growth Stages in Huanjiang Mini-Pigs

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Experimental Management
2.2. Sample Collection
2.3. Plasma and Intestinal Mucosa Oxidation-Antioxidant Indices Analysis
2.4. RNA Extraction and Gene Expression Analysis of Small Intestinal Mucosa
2.5. Data Analysis
3. Results
3.1. Impacts of Dietary CP Levels on Plasma Oxidation-Antioxidant Indices of Pigs
3.2. Impacts of Dietary CP Levels on Jejunal Oxidation-Antioxidant Indices of Pigs
3.3. Impacts of Dietary CP Levels on Ileal Oxidation-Antioxidant Indices of Pigs
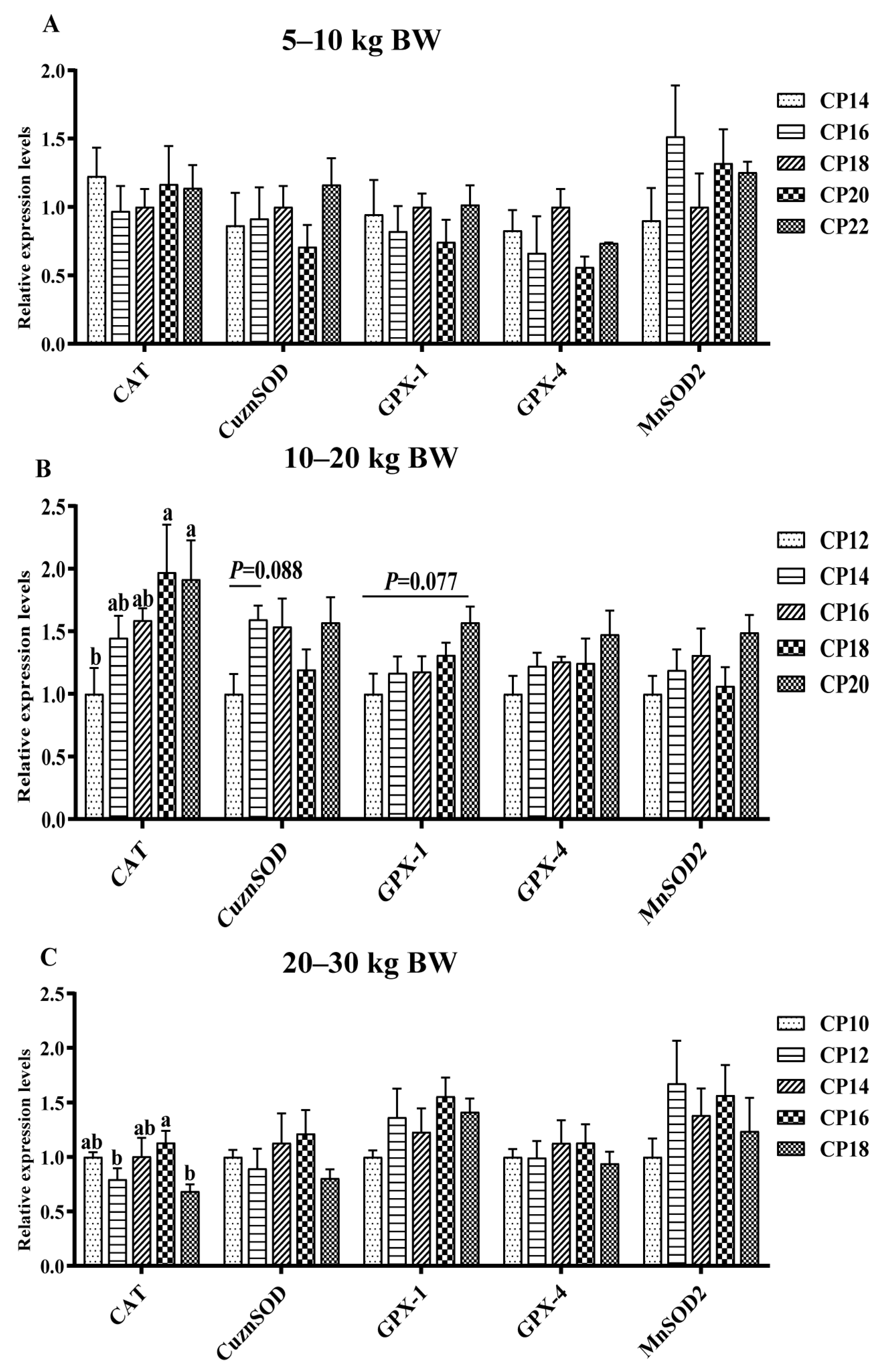
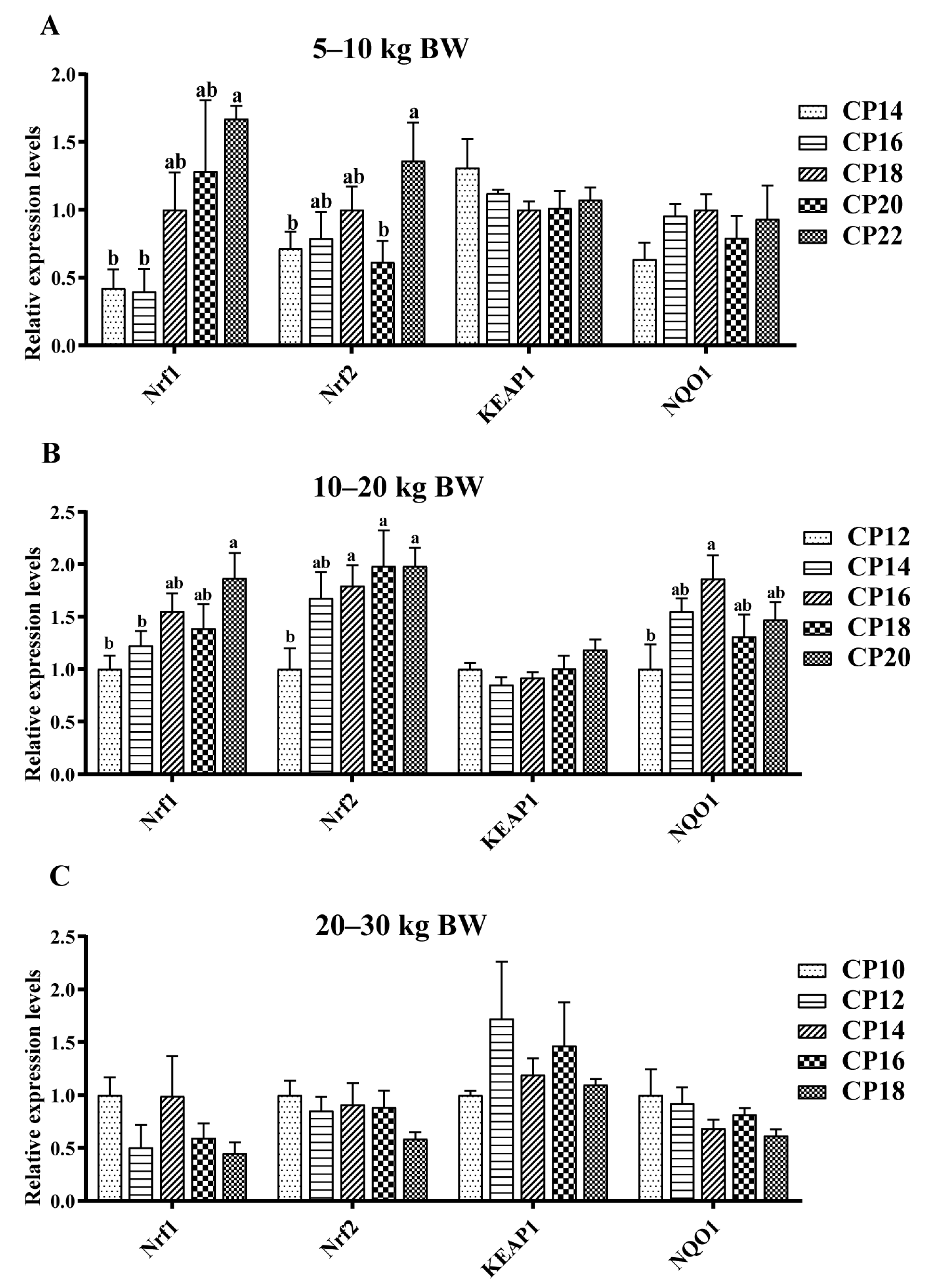
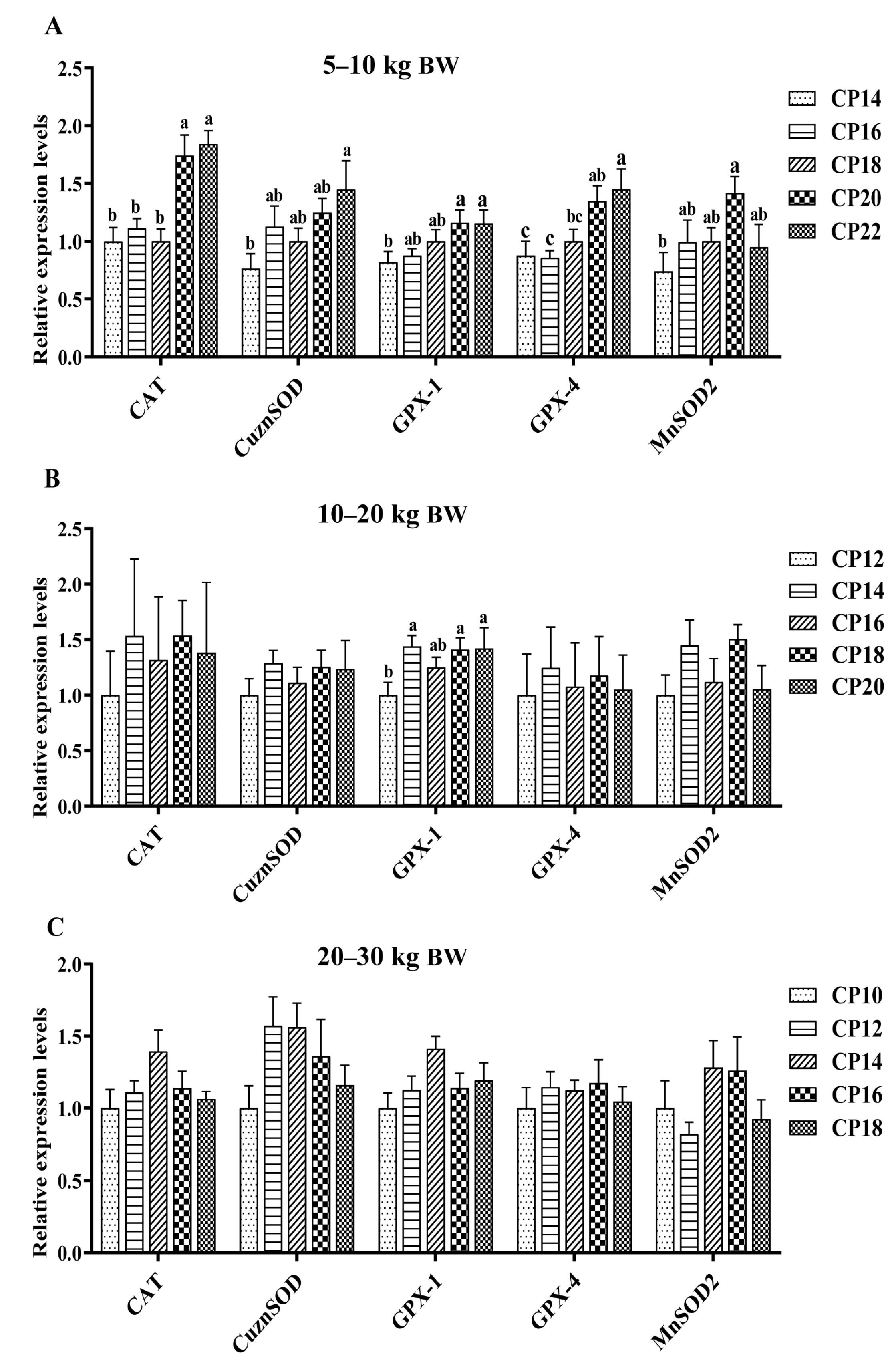
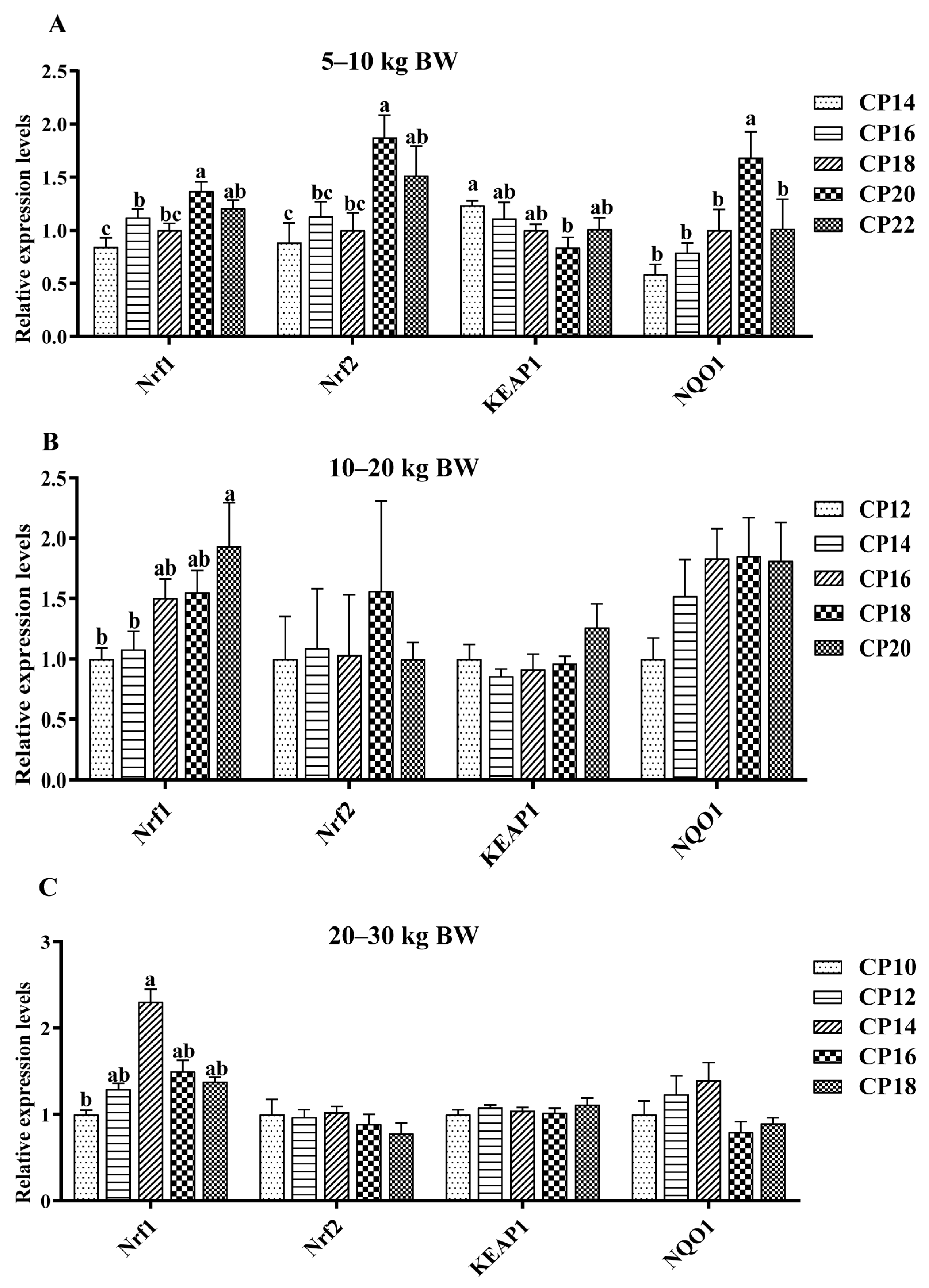
3.4. Impacts of Dietary CP Levels on Jejunal Oxidative Stress-Related Gene Expression of Pigs
3.5. Impacts of Dietary CP Levels on Ileal Oxidative Stress-Related Gene Expression of Pigs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Persson, T.; Popescu, B.O.; Cedazo-Minguez, A. Oxidative stress in alzheimer’s disease: Why did antioxidant therapy fail? Oxid. Med. Cell. Longev. 2014, 2014, 427318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borut, P.; Dusan, S.; Irina, M. Achieving the balance between ros and antioxidants: When to use the synthetic antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 956792. [Google Scholar]
- Stoyanova, S.; Geuns, J.; Hideg, É.; Ende, W. The food additives inulin and stevioside counteract oxidative stress. Int. J. Food Sci. Nutr. 2011, 62, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Tan, Y.Q.; Zhang, L.; Zhou, Y.M.; Gao, F.; Zhou, G.H. Effects of dietary supplementation with the combination of zeolite and attapulgite on growth performance, nutrient digestibility, secretion of digestive enzymes and intestinal health in broiler chickens. Asian Australas. J. Anim. Sci. 2014, 27, 1311–1318. [Google Scholar] [CrossRef]
- Bacou, E.; Walk, C.; Rider, S.; Litta, G.; Perez-Calvo, E. Dietary oxidative distress: A review of nutritional challenges as models for poultry, swine and fish. Antioxidants 2021, 10, 525. [Google Scholar] [CrossRef]
- D’Alessandro, E.; Arfuso, F.; Floridia, V.; Tardiolo, G.; Fazio, F.; Giannetto, C.; Piccione, G.; Zumbo, A. Different genotype and a liquid why-supplemented diet infludence the resilience of pigs through immune-modulation and anti-inflammatory response. Front. Vet. Sci. 2022, 9, 1046101. [Google Scholar] [CrossRef]
- Giannetto, C.; Arfuso, F.; Giudice, E.; Rizzo, M.; Piccione, G.; Mhalhel, K.; Levanti, M. Antioxidant and hepatoprotective effect of a nutritional supplement with silymarin phytosome, choline chloride, l-cysteine, artichoke, and vitamin E in dogs. Antioxidants 2022, 11, 2339. [Google Scholar] [CrossRef]
- Wang, W.; Zeng, X.; Mao, X.; Wu, G.; Qiao, S. Optimal dietary true ileal digestible threonin for supporting the mucosal barrier in small intestine of weanling pigs. J. Nutr. 2010, 140, 981–986. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhou, J.; Wang, G.; Cai, S.; Zeng, X.; Qiao, S. Advances in low-protein diets for swine. J. Anim. Sci. Biotechnol. 2018, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Vega, C.C.; Reyes-Castro, L.A.; Rodriguez-Gonzalez, G.L.; Bautista, C.J.; Vazquez-Martinez, M.; Larrea, F.; Chamorro-Cevallos, A.G.; Nathanielsz, P.W.; Zambrano, E. Resveratrol partially prevents oxidative stress and metabolic dysfunction in pregnant rats fed a low protein diet and their offspring. J. Physiol. 2016, 594, 1483–1499. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Xie, P.; Li, H.; Ma, C.; Zhang, W.; Yin, Y.; Kong, X. Fetal Huanjiang mini-pigs exhibit differences in nutrient composition according to body weight and gestational period. PLoS ONE 2018, 13, e0199939. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, Y.; Ding, H.; Huang, P.; Yin, Y.; Deng, J.; Kong, X. Effects of different dietary protein levels on the growth performance, serum biochemical parameters, fecal nitrogen, and carcass traits of Huanjiang mini-pigs. Front. Vet. Sci. 2021, 8, 777671. [Google Scholar] [CrossRef]
- Liu, Y.; Azad, M.A.K.; Zhao, X.; Zhu, Q.; Kong, X. Dietary crude protein levels alter diarrhea incidence, immunity, and intestinal barrier function of Huanjiang mini-pigs during different growth stages. Front. Immunol. 2022, 13, 908753. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Macias, H.; Romieu, I. Effects of antioxidant supplements and nutrients on patients with asthma and allergies. J. Allergy Clin. Immunol. 2014, 133, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (NRC). Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Northwest Washington, DC, USA, 2012. [Google Scholar]
- GB, NY/T 65-2004; Ministry of Agriculture of the People’s Republic of China Feeding Standard of Swine. Agriculture Press: Beijing, China, 2004.
- Wang, K.; Zhu, Q.; Kong, X.; Song, M.; Azad, M.A.K.; Xiong, L.; Zheng, Y.; He, Q. Dietary probiotics or synbiotics supplementation during gestation, lactation, and nursery periods modifies colonic microbiota, antioxidant capacity, and immune function in weaned piglets. Front. Vet. Sci. 2020, 7, 597832. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wu, X.; Yin, Y.; Zhang, C.; He, L. Preventive oral supplementation with glutamine and arginine has beneficial effects on the intestinal mucosa and inflammatory cytokines in endotoxemic rats. Amino Acids 2012, 43, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Kong, X.; Azad, M.A.K.; Zhu, Q.; Xiong, L.; Zheng, Y.; Hu, Z.; Yin, Y.; He, Q. Maternal probiotic or synbiotic supplementation modulates jejunal and colonic antioxidant capacity, mitochondrial function, and microbial abundance in Bama mini-piglets. Oxid. Med. Cell. Longev. 2021, 2021, 6618874. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparitive C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Zhu, L.H.; Zhao, K.L.; Chen, X.L.; Xu, J.X. Impact of weaning and an antioxidant blend on intestinal barrier function and antioxidant status in pigs. J. Anim. Sci. 2012, 90, 2581–2589. [Google Scholar] [CrossRef]
- Seifried, H.E.; Anderson, D.E.; Fisher, E.I.; Milner, J.A. A review of the interaction among dietary antioxidants and reactive oxygen species. J. Nutr. Biochem. 2007, 18, 567–579. [Google Scholar] [CrossRef]
- Bai, K.; Feng, C.; Jiang, L.; Zhang, L.; Zhang, J.; Zhang, L.; Wang, T. Dietary effects of Bacillus subtilis fmbj on growth performance, small intestinal morphology, and its antioxidant capacity of broilers. Poult. Sci. 2018, 97, 2312–2321. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Piao, X. Different dietary protein sources influence growth performance, antioxidant capacity, immunity, fecal microbiota and metabolites in weaned piglets. Anim. Nutr. 2022, 8, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Guo, X.; Xie, C.; Zhang, T.; Gao, P.; Gao, T.; Yin, Y. Effects of a two-meal daily feeding pattern with varied crude protein levels on growth performance and antioxidant indexes in pigs. Anim. Nutr. 2016, 2, 267–270. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Q.; Sun, S.; Huang, B.; Zhang, Y.; Xu, Y.; Zhang, S.; Xiang, H. Probiotics-fermented Massa Medicata Fermentata ameliorates weaning stress in piglets related to improving intestinal homeostasis. Appl. Microbiol. Biotechnol. 2018, 102, 10713–10727. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, Z.; Li, L.; Song, W. Resveratrol affects the expression of glutamate cysteine ligase in the kidneys of aged rats. Exp. Ther. Med. 2014, 7, 1762–1766. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, L.A.; El-Hindawy, M.M.; Alagawany, M.; Salah, A.S.; El-Sayed, S.A.A. Effect of low- or high-CP diet with cold-pressed oil supplementation on growth, immunity and antioxidant indices of growing quail. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1380–1387. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Park, E.M.; Bok, S.H.; Jung, U.J.; Kim, J.Y.; Park, Y.B.; Huh, T.L.; Kwon, O.S.; Choi, M.S. Two cinnamate derivatives produce similar alteration in mRNA expression and activity of antioxidant enzymes in rats. J. Biochem. Mol. Toxicol. 2003, 17, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, K.; Motohashi, H.; Yamamoto, M. Molecular mechanisms of the Keap1–Nrf2 pathway in stress response and cancer evolution. Genes Cells 2011, 16, 123–140. [Google Scholar] [CrossRef]
- González-Maán, D.; D’Espessailles, A.; Dossi, C.G.; San, M.M.; Mancilla, R.A.; Tapia, G.S. Rosa mosqueta oil prevents oxidative stress and inflammation through the upregulation of PPAR-α and Nrf2 in C57BL/6J mice fed a high-fat diet. J. Nutr. 2017, 147, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of Nrf2 in oxidative stress and toxicity. Ann. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Dietary CP Levels (%) | p-Values | ||||
|---|---|---|---|---|---|---|
| 5–10 kg BW | 14 | 16 | 18 | 20 | 22 | |
| CAT (U/mL) | 6.44 ± 0.32 b | 7.89 ± 0.68 ab | 7.02 ± 0.53 b | 8.98 ± 0.44 a | 7.67 ± 0.68 ab | 0.026 |
| GSH (μg/mL) | 58.80 ± 1.90 b | 75.13 ± 6.60 ab | 86.47 ± 8.57 a | 98.28 ± 12.66 a | 94.44 ± 8.49 a | 0.020 |
| GSH-Px (U/mL) | 78.94 ± 4.32 b | 85.39 ± 4.28 b | 83.88 ± 5.76 b | 100.39 ± 3.63 a | 92.30 ± 3.38 ab | 0.049 |
| MDA (nmol/mL) | 8.84 ± 0.45 | 9.17 ± 0.28 | 9.14 ± 0.31 | 9.07 ± 0.37 | 9.88 ± 0.38 | 0.400 |
| SOD (U/mL) | 108.78 ± 9.08 | 96.92 ± 7.48 | 102.57 ± 7.64 | 108.23 ± 7.56 | 113.09 ± 7.68 | 0.679 |
| 10–20 kg BW | 12 | 14 | 16 | 18 | 20 | |
| CAT (U/mL) | 7.90 ± 0.52 ab | 9.41 ± 0.44 a | 9.15 ± 0.68 a | 9.52 ± 0.66 a | 7.36 ± 0.45 b | 0.030 |
| GSH (μg/mL) | 95.92 ± 8.18 b | 90.48 ± 6.65 b | 91.97 ± 5.71 b | 117.97 ± 5.42 a | 79.57 ± 4.82 b | 0.002 |
| GSH-Px (U/mL) | 94.94 ± 9.13 | 96.56 ± 3.78 | 96.38 ± 7.78 | 111.94 ± 6.07 | 90.98 ± 6.31 | 0.312 |
| MDA (nmol/mL) | 9.63 ± 0.44 a | 9.62 ± 0.62 a | 9.45 ± 0.27 a | 8.03 ± 0.47 b | 10.34 ± 0.38 a | 0.029 |
| SOD (U/mL) | 99.28 ± 7.85 ab | 121.42 ± 8.76 a | 124.34 ± 4.95 a | 118.88 ± 8.20 ab | 95.18 ± 11.18 b | 0.049 |
| 20–30 kg BW | 10 | 12 | 14 | 16 | 18 | |
| CAT (U/mL) | 9.57 ± 0.84 ab | 9.02 ± 0.34 b | 7.88 ± 0.41 b | 10.90 ± 0.67 a | 9.66 ± 0.67 ab | 0.020 |
| GSH (μg/mL) | 104.66 ± 4.54 a | 101.71 ± 7.66 a | 111.60 ± 7.14 a | 115.14 ± 2.20 a | 78.14 ± 3.30 b | 0.004 |
| GSH-Px (U/mL) | 113.45 ± 10.00 | 115.74 ± 6.98 | 105.39 ± 9.59 | 115.31 ± 7.63 | 110.39 ± 7.46 | 0.709 |
| MDA (nmol/mL) | 8.12 ± 0.67 | 7.94 ± 0.56 | 7.67 ± 0.54 | 8.31 ± 0.59 | 8.68 ± 0.34 | 0.801 |
| SOD (U/mL) | 149.56 ± 10.52 | 151.91 ± 6.33 | 146.50 ± 7.88 | 141.69 ± 8.26 | 135.80 ± 14.32 | 0.805 |
| Items | Dietary CP Levels (%) | p-Values | ||||
|---|---|---|---|---|---|---|
| 5–10 kg BW | 14 | 16 | 18 | 20 | 22 | |
| CAT (U/mL) | 3.92 ± 0.20 | 4.33 ± 0.27 | 4.44 ± 0.23 | 4.41 ± 0.30 | 4.13 ± 0.30 | 0.606 |
| GSH (μg/mL) | 29.22 ± 2.52 b | 40.95 ± 1.78 a | 40.53 ± 2.52 a | 31.81 ± 2.15 b | 33.93 ± 2.30 ab | 0.008 |
| GSH-Px (U/mL) | 43.22 ± 3.50 | 53.91 ± 2.64 | 44.83 ± 2.49 | 51.68 ± 2.30 | 46.08 ± 1.50 | 0.058 |
| MDA (nmol/mL) | 3.67 ± 0.16 | 3.20 ± 0.22 | 3.91 ± 0.30 | 3.43 ± 0.24 | 3.35 ± 0.20 | 0.257 |
| SOD (U/mL) | 19.45 ± 1.83 b | 32.16 ± 2.27 a | 31.69 ± 1.53 a | 30.81 ± 2.46 a | 25.78 ± 2.87 ab | 0.004 |
| 10–20 kg BW | 12 | 14 | 16 | 18 | 20 | |
| CAT (U/mL) | 4.02 ± 0.16 b | 4.69 ± 0.36 ab | 5.18 ± 0.28 a | 5.04 ± 0.29 a | 4.75 ± 0.36 ab | 0.045 |
| GSH (μg/mL) | 35.13 ± 1.35 b | 40.71 ± 2.51 ab | 45.80 ± 1.71 a | 39.68 ± 1.98 ab | 34.07 ± 2.43 b | 0.011 |
| GSH-Px (U/mL) | 41.86 ± 2.94 b | 57.57 ± 3.44 a | 56.23 ± 3.65 a | 57.74 ± 2.30 a | 53.22 ± 2.34 a | 0.004 |
| MDA (nmol/mL) | 3.21 ± 0.34 | 3.18 ± 0.07 | 2.92 ± 0.25 | 3.15 ± 0.26 | 3.47 ± 0.23 | 0.633 |
| SOD (U/mL) | 23.37 ± 2.87 b | 35.54 ± 4.85 a | 34.79 ± 1.92 a | 37.76 ± 2.71a | 30.10 ± 3.01 ab | 0.045 |
| 20–30 kg BW | 10 | 12 | 14 | 16 | 18 | |
| CAT (U/mL) | 4.62 ± 0.30 | 4.76 ± 0.27 | 4.65 ± 0.29 | 4.71 ± 0.21 | 4.73 ± 0.51 | 0.997 |
| GSH (μg/mL) | 47.77 ± 3.40 | 40.78 ± 2.26 | 50.56 ± 0.71 | 42.04 ± 3.68 | 42.56 ± 3.42 | 0.102 |
| GSH-Px (U/mL) | 58.05 ± 3.78 | 56.40 ± 3.99 | 56.27 ± 4.56 | 55.25 ± 2.93 | 63.19 ± 3.28 | 0.647 |
| MDA (nmol/mL) | 3.24 ± 0.23 a | 2.82 ± 0.24 ab | 2.31 ± 0.16 b | 2.27 ± 0.23 b | 2.60 ± 0.21 ab | 0.023 |
| SOD (U/mL) | 41.65 ± 4.99 | 37.29 ± 1.99 | 42.91 ± 2.25 | 41.20 ± 1.88 | 45.52 ± 4.31 | 0.538 |
| Items | Dietary CP Levels (%) | p-Values | ||||
|---|---|---|---|---|---|---|
| 5–10 kg BW | 14 | 16 | 18 | 20 | 22 | |
| CAT (U/mL) | 3.46 ± 0.20 | 4.06 ± 0.26 | 4.03 ± 0.34 | 3.97 ± 0.32 | 4.07 ± 0.22 | 0.539 |
| GSH (μg/mL) | 37.10 ± 2.41 b | 40.58 ± 0.82 ab | 44.12 ± 1.85 a | 36.08 ± 3.29 b | 29.66 ± 1.26 c | 0.001 |
| GSH-Px (U/mL) | 40.87 ± 2.70 | 38.87 ± 2.35 | 41.98 ± 2.41 | 45.06 ± 3.57 | 40.20 ± 2.21 | 0.513 |
| MDA (nmol/mL) | 3.27 ± 0.28 | 2.94 ± 0.15 | 2.72 ± 0.12 | 2.63 ± 0.13 | 2.86 ± 0.19 | 0.177 |
| SOD (U/mL) | 29.66 ± 2.00 | 29.96 ± 1.76 | 25.27 ± 2.96 | 31.97 ± 3.99 | 22.79 ± 2.34 | 0.157 |
| 10–20 kg BW | 12 | 14 | 16 | 18 | 20 | |
| CAT (U/mL) | 4.69 ± 0.41 ab | 4.12 ± 0.22 b | 4.97 ± 0.22 a | 5.07 ± 0.15 a | 4.13 ± 0.43 b | 0.049 |
| GSH (μg/mL) | 35.30 ± 3.53 | 38.56 ± 2.97 | 48.79 ± 3.06 | 42.94 ± 4.74 | 42.47 ± 4.79 | 0.219 |
| GSH-Px (U/mL) | 48.66 ± 2.99 b | 45.61 ± 2.64 b | 60.49 ± 1.95 a | 47.19 ± 3.79 b | 50.00 ± 4.55 b | 0.049 |
| MDA (nmol/mL) | 3.12 ± 0.16 a | 2.78 ± 0.17 ab | 2.88 ± 0.21 ab | 2.44 ± 0.21 b | 2.43 ± 0.12 b | 0.045 |
| SOD (U/mL) | 24.54 ± 1.85 b | 27.14 ± 3.53 b | 41.08 ± 2.76 a | 37.85 ± 3.96 a | 27.67 ± 3.64 b | 0.004 |
| 20–30 kg BW | 10 | 12 | 14 | 16 | 18 | |
| CAT (U/mL) | 4.22 ± 0.18 | 4.48 ± 0.25 | 4.88 ± 0.37 | 4.71 ± 0.17 | 4.43 ± 0.30 | 0.433 |
| GSH (μg/mL) | 43.56 ± 3.23 | 43.66 ± 3.61 | 46.76 ± 2.79 | 41.74 ± 3.36 | 38.54 ± 4.79 | 0.638 |
| GSH-Px (U/mL) | 55.22 ± 3.30 | 53.97 ± 2.57 | 53.73 ± 3.01 | 55.50 ± 3.51 | 47.28 ± 2.78 | 0.316 |
| MDA (nmol/mL) | 2.56 ± 0.17 a | 2.47 ± 0.08 ab | 1.89 ± 0.22 c | 1.99 ± 0.19 bc | 2.30 ± 0.21 abc | 0.049 |
| SOD (U/mL) | 28.89 ± 3.23 b | 46.53 ± 1.15 a | 47.25 ± 5.36 a | 49.19 ± 2.59 a | 32.55 ± 2.22 b | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Azad, M.A.K.; Zhao, X.; Zhu, Q.; Kong, X. Dietary Protein Levels Modulate the Antioxidant Capacity during Different Growth Stages in Huanjiang Mini-Pigs. Antioxidants 2023, 12, 148. https://doi.org/10.3390/antiox12010148
Liu Y, Azad MAK, Zhao X, Zhu Q, Kong X. Dietary Protein Levels Modulate the Antioxidant Capacity during Different Growth Stages in Huanjiang Mini-Pigs. Antioxidants. 2023; 12(1):148. https://doi.org/10.3390/antiox12010148
Chicago/Turabian StyleLiu, Yating, Md. Abul Kalam Azad, Xichen Zhao, Qian Zhu, and Xiangfeng Kong. 2023. "Dietary Protein Levels Modulate the Antioxidant Capacity during Different Growth Stages in Huanjiang Mini-Pigs" Antioxidants 12, no. 1: 148. https://doi.org/10.3390/antiox12010148