1. Introduction
Aging is a complicated process that involves morphological and metabolic changes in individual cells as well as the whole organism. The oxidative stress theory is one of the most prevalent ideas for how aging happens at the molecular level [
1]. During the aging process in laying hens, harmful reactive oxygen species (ROS) are gradually produced, resulting in oxidative stress damage, including a decrease in liver antioxidant capacity [
2,
3], changes in intestinal oxidative state [
4], and the aging of reproductive organs [
5,
6], which lead to a decline in egg production performance. Oxidative stress reaches its peak when the hen is about 65 to 70 weeks old [
7]. After 75 weeks of age, laying hens’ performance often declines significantly [
8], which is connected with oxidative damage and lower immunity [
9,
10,
11,
12,
13]. The gut, as a key organ of diet digestion, absorption, and transformation, is easily impacted by oxidants and antioxidants in diet, resulting in redox balance shifts. Oxidative stress is a major contributor to the onset of inflammatory disorders [
14]. Furthermore, intestinal inflammation decreases nutritional absorption and egg production, and inflammatory cytokines in turn can cause oxidative stress [
15]. Changes in intestinal morphology, antioxidant status, and inflammation lead to poor intestinal health, which is one of the key causes of poor laying performance [
16]. Changing components of the diet can quickly affect the makeup and activity of the gut microbiota [
17], which plays a significant role in the development of the immune system in mammals and poultry, as well as maintaining immune system homeostasis [
18,
19].
Polyphenols, which are critical micronutrients in the diet, are one of the most abundant and widespread groups in the plant world, with over 8000 polyphenolic chemicals discovered. Polyphenols (phenolic acids, flavonoids, stilbenes, and lignans) have been extensively explored in recent decades due to their numerous health advantages [
20,
21]. Polyphenols have anti-inflammatory, antibacterial, and antioxidant activities that can affect egg production and health in older laying hens by improving gut integrity and function, lowering inflammation, or modifying microbial communities [
22,
23]. In older laying hens, resveratrol administration improved performance, lipid-related markers, and antioxidant activity [
24]. Tea polyphenols can improve serum and liver antioxidant capability while increasing egg production [
25,
26,
27]. Flavonoids have been used as feed additive to promote laying hens health and productive performance [
28]. Rutin is a flavonoid that is found in abundance in plants. Rutin has a wide range of pharmacological effects on biological systems, including analgesic, anti-inflammatory, and anti-arthritis properties [
29,
30,
31]. Rutin supplementation at doses of 0.50 and 1.00 g/kg substantially increased superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GSH-Px) activity and decreased malondialdehyde (MDA) concentrations in serum, showing rutin’s potential to transport electrons, activate antioxidant enzymes, and thereby reduce oxidative stress [
32]. Among the immunological parameters studied, lysozyme activity, nitric oxide concentrations, and IgM synthesis were considerably greater in rutin-fed birds than in control birds; however, rutin at any quantity had no influence on IgG and IgA concentrations or lymphoid organ weight [
33]. Previous studies suggested that rutin is a promising feed additive for broilers. Considering that aged laying hens suffer from significant oxidative damage and lower immunity, the use of antioxidants is crucial; however, there is limited study on the effects of antioxidant rutin on aging laying hens. The purpose of this research was to investigate rutin’s antioxidant capacity, immunomodulatory effects, and gut microbiome impacts in an aged laying hen.
2. Materials and Methods
2.1. Animals, Diets, and Experimental Design
All procedures were conducted under the guidelines of Nanjing Agricultural University Institutional Animal Care and Use Committee (Certification No.: SYXK(Su)2017-0007).
A total of 216 Hyline Brown aged laying hens (560 days of age) were obtained from a local commercial laying hens farm in this study. Laying hens were randomly divided into three treatments with six replicates per group. After a two-week adjustment, laying hens were separately fed with one of three kind of diets: basal diet; basal diet +500 mg/kg rutin; basal diet +1000 mg/kg rutin, and the trial lasted eight weeks. Rutin was provided by Jiangsu Bison Biotechnology Co., Ltd. (Taizhou, Jiangsu, China). The content of rutin was 96%, determined by high-performance liquid chromatography (HPLC) diffraction. These diets were formulated to meet or exceed the National Research Council (NRC, 1994) recommendation, and the composition and nutrient levels are shown in
Table 1. Hens were free to drink water throughout the experiment and were exposed to a 16-h/8-h light/dark cycle.
2.2. Sample Collection
The small intestine was dissected and sampled on a refrigerated stainless-steel tray, clear of the mesentery. Middle sections of the jejunum and ileum from all hens were harvested, flushed several times with ice-cold phosphate-buffered saline (pH 7.4), fixed with 4% paraformaldehyde-PBS, and kept at 4 C for evaluation of the mucosal morphology. The jejunum and ileum mucosas were quickly scraped and fully washed with ice-cold phosphatebuffered saline before being frozen in liquid nitrogen and stored at −80 C, for intestinal antioxidant capacity and immune parameters, as well as mRNA expression analysis. Meanwhile, the contents of the cecum were collected aseptically, put in a centrifugal tube, quickly frozen in liquid nitrogen, and kept at −80 C for microbiota analysis.
2.3. Intestinal Morphological Analysis
The middle sections of ileum and jejunum from all laying hens were removed from 4% paraformaldehyde-PBS and rinsed with water. Then, they were dehydrated with a series of different ethanol concentrations, cleared with xylene until saturation, and embedded with paraffin. A section of 5 was cut for histological analysis. The sections were deparaffinized and hydrated, and then stained with H&E. Villus height and crypt depth were measured.
2.4. Intestinal Antioxidant Capacity
The SOD activity and MDA, total antioxidant capacity (T-AOC) levels in jejunal mucosa and ileal mucosa were measured in accordance with the manufacturer’s instructions using a commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.5. Intestinal Immune Parameters
Protein concentration was tested in the supernatant of the jejunum mucosa and ileum mucosa. Meanwhile, sIgA, IgG, IgM, IL-1, TNF-, IFN-, IL-4, IL-6, and IL-10 were tested by using ELISA kits in accordance with the manufacturer’s instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.6. Real-Time Quantitative (RT-PCR)
For total RNA extraction, a sample of 50.0–60.0 mg of tissue (intestinal mucosa) was homogenized in 1.00 mL of RNAiso Plus (TaKaRa, Dalian, China). The RNA quality and quantity were then assessed using a Nano Drop ND-1000 (Nano Drop Technologies, Wilmington, DE, USA) of optical density at 260 and 280 nm. The concentration of RNA samples was diluted with diethyl pyrocarbonate-treated water to 0.5
g/
L, then reverse-transcribed into cDNA using the Prime Script RT Master Mix reagentkit in accordance with the manufacturer’s instructions (TaKaRa, Dalian, China) immediately. The total RNA of the samples was processed to synthesize complementary DNA with a PrimeScript
TM RT reagent kit according to the manufacturer’s instructions (TaKaRa, Dalian, China). The reverse transcription was conducted at 37
C for 15 min, 85
C for 5 s. The relative mRNA expression of target genes was assessed using quantitative real-time PCR. The primer sequences, including IL-1
, TNF-
, IFN-
, IL-4, IL-10, Occludin, Claudin-1, Claudin-2, ZO-1,
-actin, and their gene bank ID numbers are presented in
Table 2.
2.7. Gut Microbiota Sequencing
Total genome DNA was isolated from the cecum using the QIAamp DNA Stool Mini Kit (QIAGEN, Dusseldorf, Germany), and DNA concentration and purity were measured on 1% agarose gels. The v3-v4 region of the 16S rDNA gene sequences were amplified by using 338F (ACTCCTACGGGAGGCAGCAG) and 806R (GGACTACHVGGGTWTCTAAT).Then, the elution with Tris-HCl buffer and detection by 2% agarose electrophoresis were done, and all amplicons were purified using an AxyPrep DNA gel extraction kit (Axygen Biosciences, Union City, CA, USA). After being quantified and purified, amplicons were sequenced on an Illumina MiSeq Sequencer. The data were all analyzed using the free online platform of Majorbio Cloud Platform (
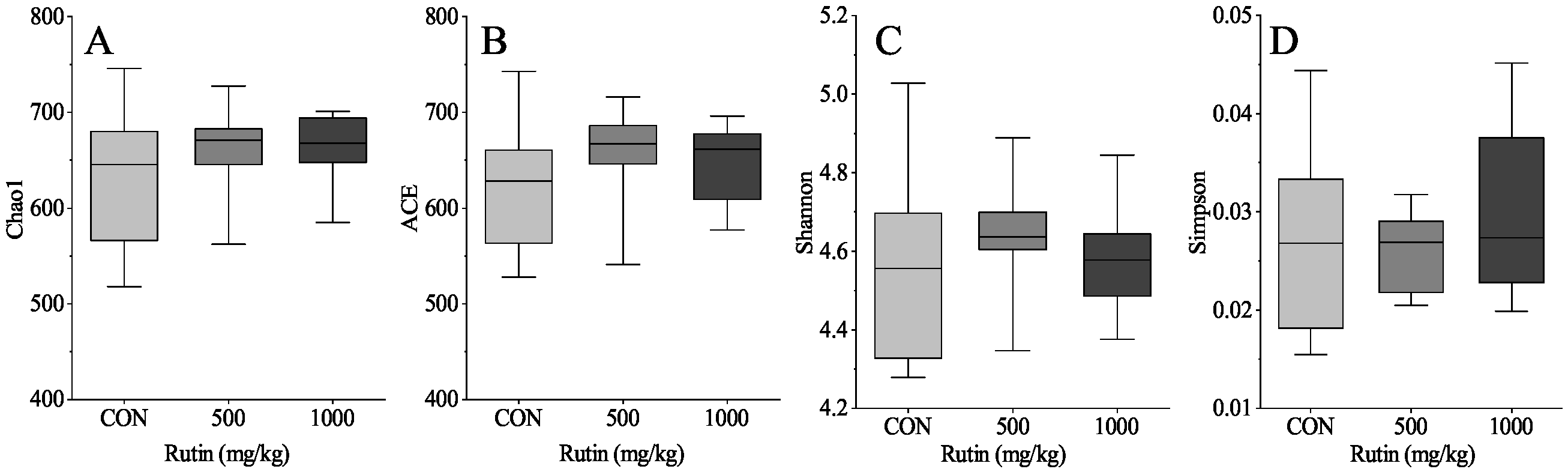
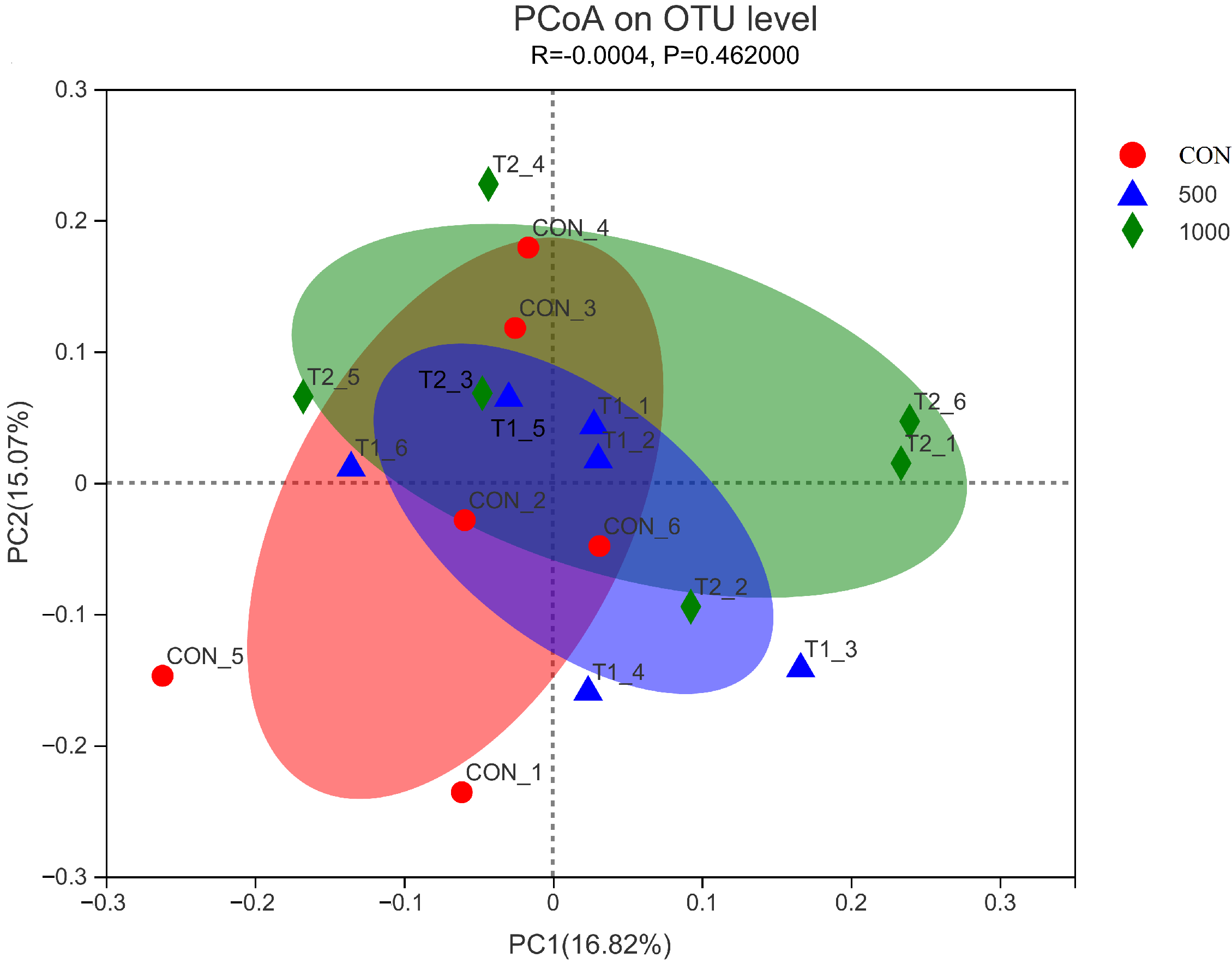
www.majorbio.com). The sequences were analyzed and assigned to operational taxonomic units (OTUs; 97% identity). Alpha diversity included calculation of ACE, Chao1, Shannon, and Simpson indices, beta diversity using principal coordinate analysis (PCoA) and unweighted Unifrac cluster tree and environmental factor correlation analysis.
5. Discussion
Intestinal integrity and intestinal morphology play a crucial role in intestinal function, such as the absorption of nutrient, and form a major physical barrier to protect against pathogens and toxic compounds [
23,
34]. An increase in the height or the width of the villus indicates an increase in the surface area, which can better absorb the available nutrients, thereby regulating the nutritional status and improving the development and health of the poultry [
35]. Fideles et al. [
36] pointed out that pretreatment with rutin (100 and 200 mg/kg) showed a marked reversal in villi shortening in the duodenum and the jejunum in mice of 5-Fluorouracil (FU) induced experimental intestinal mucositis; however, all rutin doses failed to reverse the villi shortening caused by 5-FU in the ileum. Additionally, Zhang et al. [
37] reported that dietary rutin (250 mg/kg) and rutin (1000 mg/kg) in weaned mice for 21 days increased jejunum villus height, indicating that rutin improved intestinal morphology. The current study found that the villus height and villus-crypt ratio were longer in a rutin-supplemented diet in jejunal compared with the control group. Rutin had no influence on ileal villus height, crypt depth, and the villi-crypt ratio. Rutin has a greater impact on gut morphology in the jejunum than in the ileum. Previous research has suggested that rutin may reduce inflammation and oxidative stress via the cyclooxygenase 2 (COX-2) pathway, while also improving intestinal morphology [
36]. However, the detailed mechanism of rutin on intestinal morphology needs further evaluation. The diet with 500 mg/kg rutin dramatically improved claudin-2 expression in the jejunum. Claudin-2 is expressed in the tight junctions of leaky epithelia, where it forms cation-selective and water permeable paracellular channels [
38]. The results suggested that rutin is beneficial for improving intestinal integrity.
In vitro, phenolic compounds have ideal structural chemistry for free radical-scavenging activities, high levels of 2,2-diphenyl-1-picrylhydrazyl (DPPH) free radical scavenging activity, and have been shown in vitro to be more effective antioxidants than vitamins E and C on a molar basis [
39,
40]. In the animal antioxidant system, SOD removes superoxide radicals by accelerating their conversion to hydrogen peroxide [
41]. MDA is a byproduct of polyunsaturated fatty acid peroxidation in cells, and MDA levels are routinely used to assess oxidative stress and antioxidant status [
42]. T-AOC is usually applied as a comprehensive measure to assess an animal’s antioxidant capability [
43]. A previous study have indicated that dietary rutin supplementation enhanced the serum antioxidant capacity by increasing SOD, CAT, and GSH-Px activity while decreasing MDA levels in serum of broilers [
32]. In this study, rutin supplementation in the diet had no effect on the levels of MDA, and T-AOC, or SOD activity in the jejunum and ileum. This suggests that the antioxidant effect of rutin differs significantly in vivo and in vitro, and that differences in antioxidant ability of rutin in vivo studies may be related to animal species, age, tissue, and other factors.
The gut is the biggest immunological organ [
44]. Secretory immunoglobulin A (sIgA) is a crucial line of defense on the mucosal surface of the intestine that protects the intestinal epithelium from toxins and pathogenic bacteria [
45]. Our findings revealed that a diet with 500 mg/kg rutin increased the sIgA content but had no effect on the IgG and IgM content. Furthermore, a diet with 1000 mg/kg rutin can enhance sIgA and IgM concentrations. Ileal immunoglobulins were more responsive to rutin addition than those in the jejunum. The addition of two doses of rutin, 500 mg/kg and 1000 mg/kg, increased the concentrations of sIgA, IgM and IgG in the ileal mucosa. The results showed that rutin increases the function of the intestinal immune barrier by promoting the synthesis and release of immune globulin in the intestinal mucosa of aged laying hens. Previous investigations have reported that rutin inhibits the production of proinflammatory cytokines in microglia by lowering TNF-
and IL-1
levels in human neuroblastoma ( SH-SY5Y ) cells [
46]. Furthermore, rutin supplementation 10 or 20 days after trimethyltin (TMT) injection reduced the TMT-induced elevation of mRNA expression levels of reactive microglia marker and pro-inflammatory cytokines in rats [
47]. In this study, we discovered that supplementing with rutin reduces TNF-
in jejunal and ileal mucosa, the effect on IFN-
in the ileum was greater than that in the jejunum, and high-dose rutin supplementation can significantly decrease the IL-1
, TNF-
, and IFN-
and increase IL-10 content. The research showed that dietary rutin supplementation had an influence on intestinal cytokines, which was dose-dependent. The effect on the ileum is stronger than that on the jejunum, which may be associated with the intestinal metabolism mechanism of rutin. When compared to the control group, dietary supplementation of rutin substantially reduced IL-1
, IFN-
, and IL-4 mRNA expression in the jejunal mucosa. In the ileal mucosa, both groups treated with rutin dramatically reduced TNF-
mRNA expression while significantly boosting IFN-
, IL-4, and IL-10 mRNA expression. In view of the above data, we speculated that rutin can regulate cytokine content via modulating cytokine gene expression, and it can also function as an immunomodulator to reduce inflammation.
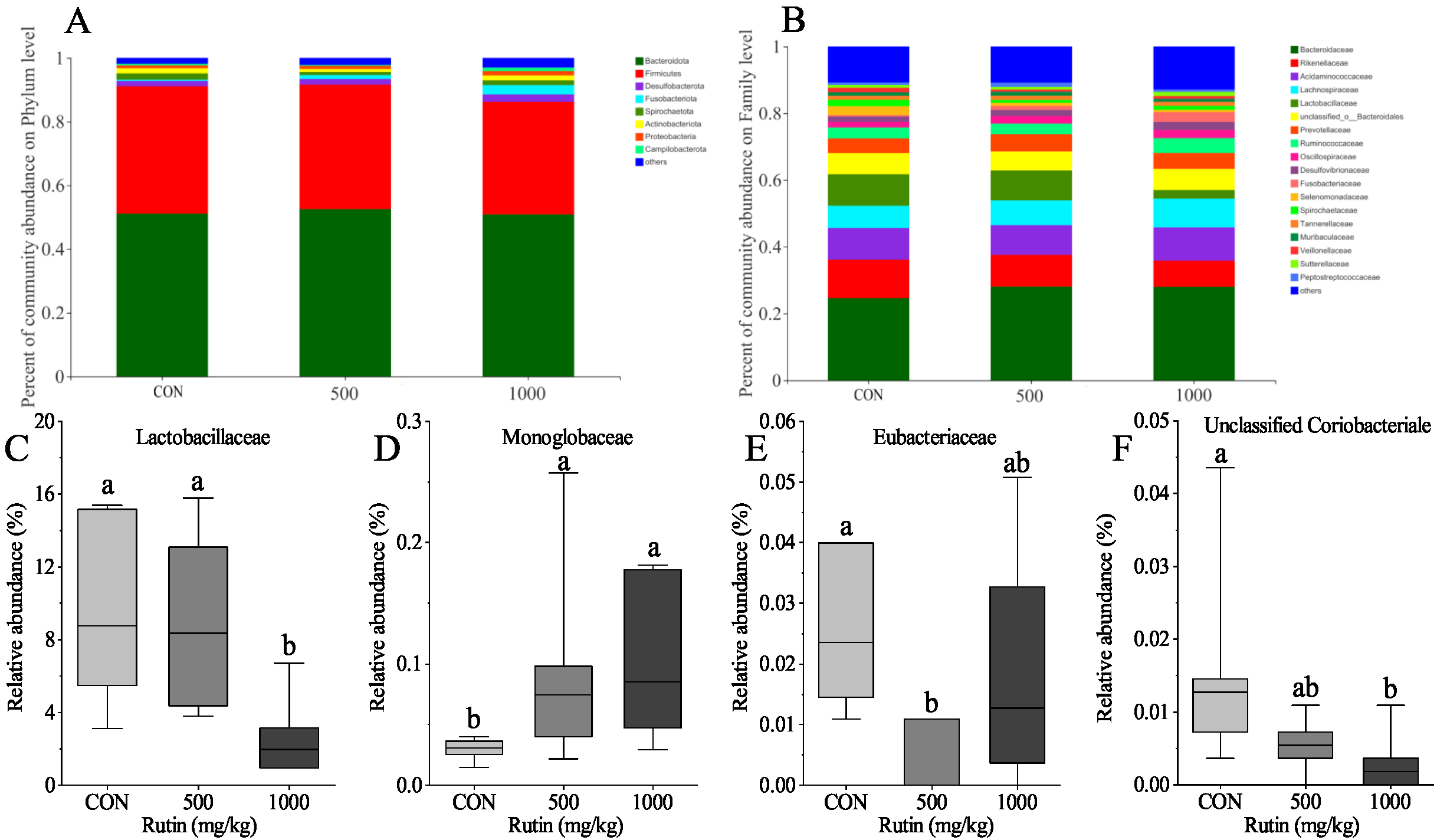
Gut bacteria play an important role in rutin function [
48]. The majority of dietary rutin passes through the large intestine, where the colonic bacteria breaks down rutin into quercetin aglycone. Enterobacteria can then absorb or further degrade the quercetin aglycone to form different ring-fission products [
49]. Rutin treatments all modulated the gut microbiota in rats during high diet fat intake [
50]. The influence of rutin on the alpha and beta diversity of the gut microbiota in laying hens was not seen in this investigation. Rutin, on the other hand, increased the relative abundance of Lactobacillaceae. Lactobacillus has been demonstrated in several studies to increase the amount of Treg cells, which are key in controlling inflammation [
51,
52]. Meanwhile, rutin treatment reduced the relative abundance of Monoglobaceae, Eubacteriaceae, and Unclassified Coriobacteriale in the current research. Based on these findings, we infer that rutin may alter gut microbial composition and thereby change the intestinal immune system function in aged laying hens.





{kind=link}
{kind=link}
{kind=link}