
Evaluation of Hepatic Detoxification Effects of Enteromorpha prolifera Polysaccharides against Aflatoxin B1 in Broiler Chickens

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Growth Performance and Sampling
2.3. Serum Biochemical Analysis
2.4. Hepatic Histopathology and Apoptosis Analysis
2.5. Determination of Enzymatic Activity and AFBO-DNA Content of Liver
2.6. Gene Expression Analysis of Liver
2.7. Immunohistochemical Analysis and Western Blotting
2.8. Statistical Analysis
3. Results
3.1. Growth Performance, Serum Biochemical Indicators and Relative Liver Weight
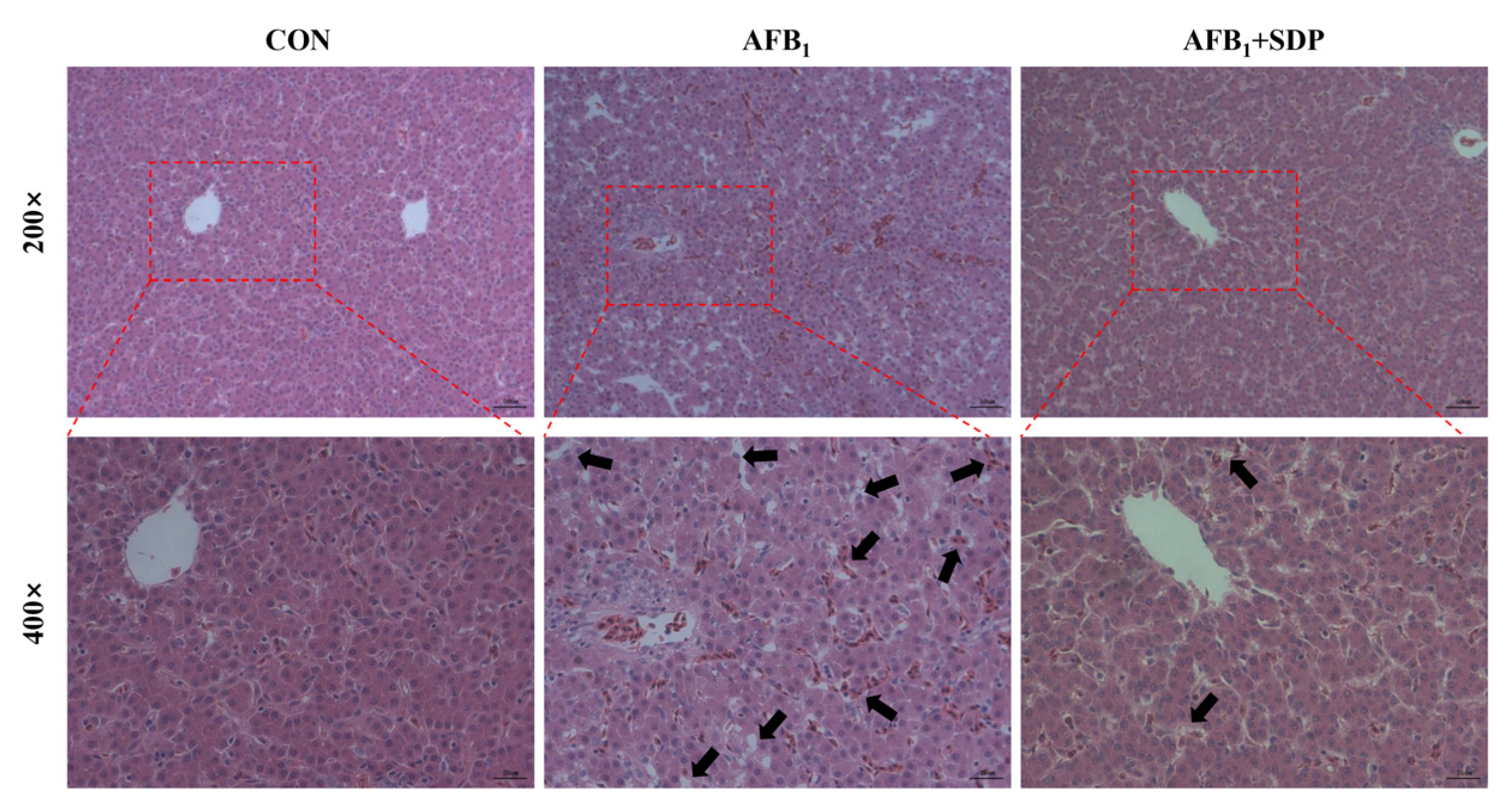
3.2. Histopathology and Apoptosis of Liver
3.3. Enzymatic Activity and AFBO-DNA level of Liver
3.4. Hepatic mRNA Expression of Phase I Detoxification Enzyme-Related Genes
3.5. Hepatic mRNA Expression of Antioxidant and Phase II Detoxification Enzyme Related Genes
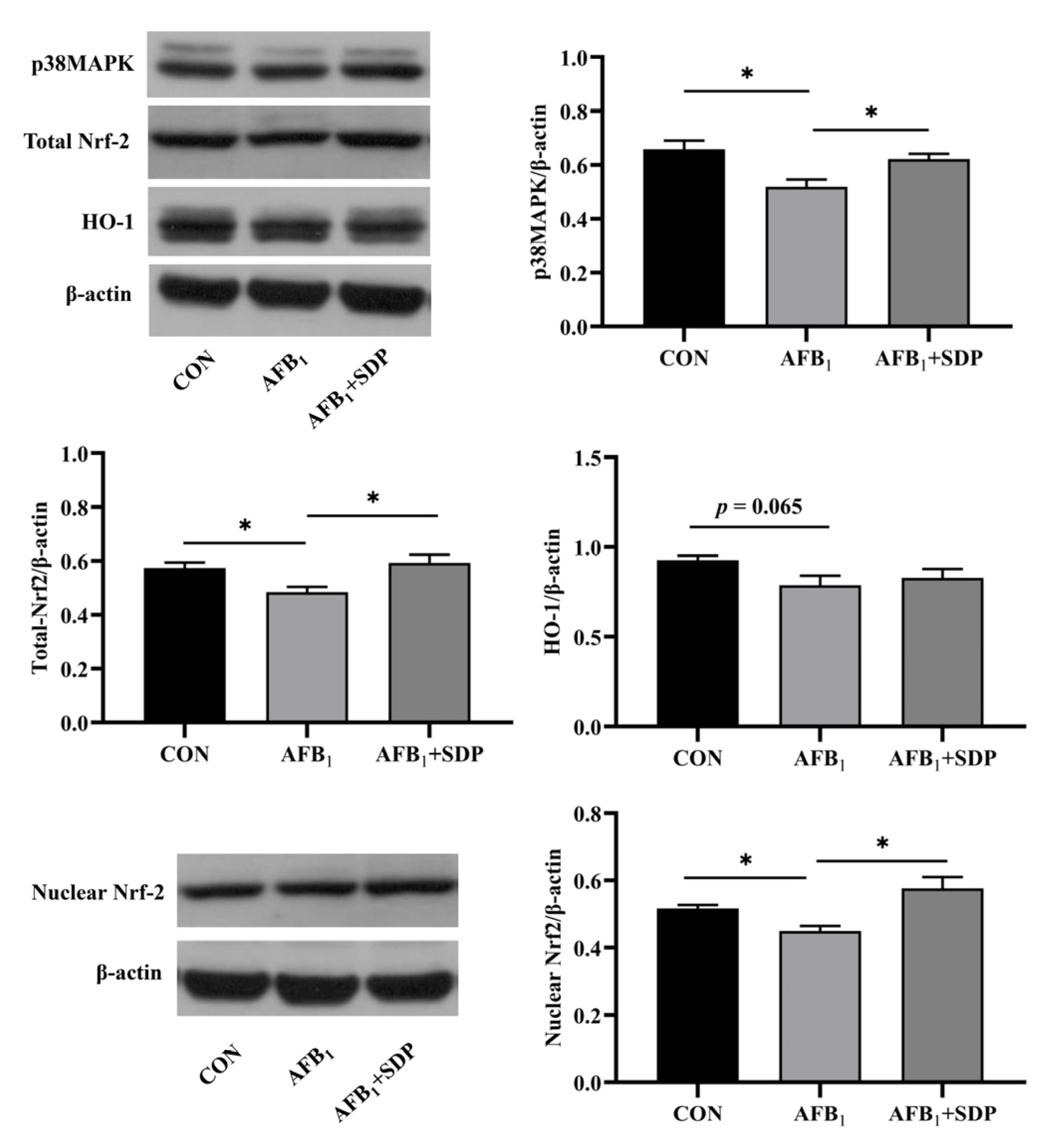
3.6. Protein Expression of p38MAPK/Nrf2 Signaling Pathway in the Liver
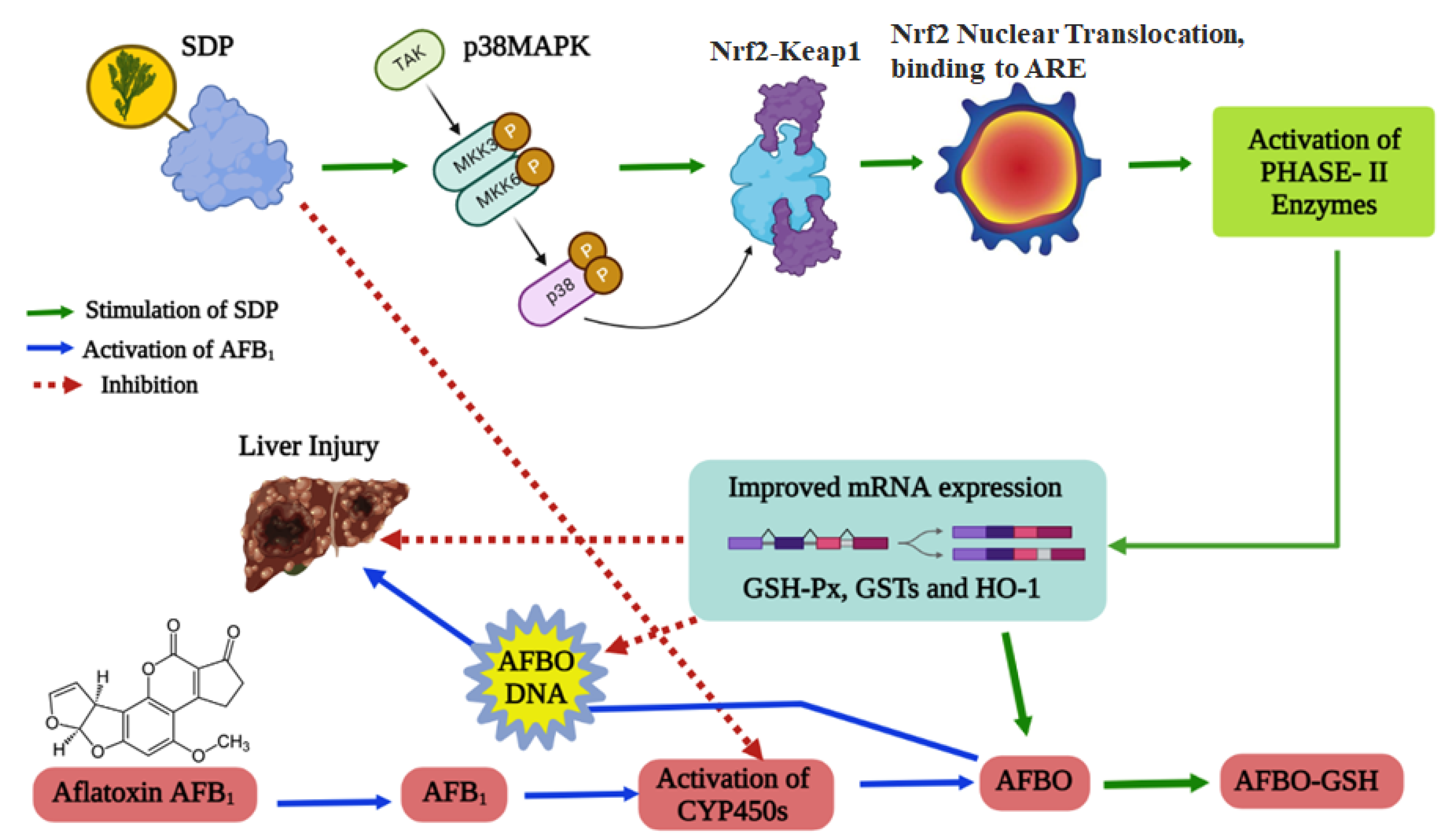
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maharjan, P.; Martinez, D.; Weil, J.; Suesuttajit, N.; Umberson, C.; Mullenix, G.; Hilton, K.; Beitia, A.; Coon, C. Physiological growth trend of current meat broilers and dietary protein and energy management approaches for sustainable broiler production. Animals 2021, 15, 100284. [Google Scholar] [CrossRef] [PubMed]
- Ochieng, P.E.; Scippo, M.-L.; Kemboi, D.C.; Croubels, S.; Okoth, S.; Kang’ethe, E.K.; Doupovec, B.; Gathumbi, J.K.; Lindahl, J.F.; Antonissen, G. Mycotoxins in Poultry Feed and Feed Ingredients from Sub-Saharan Africa and Their Impact on the Production of Broiler and Layer Chickens: A Review. Toxins 2021, 13, 633. [Google Scholar]
- den Hollander, D.; Croubels, S.; Lauwers, M.; Caekebeke, N.; Ringenier, M.; De Meyer, F.; Reisinger, N.; Van Immerseel, F.; Dewulf, J.; Antonissen, G. Applied Research Note: Biomonitoring of mycotoxins in blood serum and feed to assess exposure of broiler chickens. J. Appl. Poult. Res. 2021, 30, 100111. [Google Scholar] [CrossRef]
- Ogbuewu, I. Effects of mycotoxins in animal nutrition: A review. Asian J. Anim. Sci. 2011, 5, 1933. [Google Scholar]
- Agag, B. Mycotoxins in foods and feeds: 1-aflatoxins. Ass. Univ. Bull. Environ. Res. 2004, 7, 173–205. [Google Scholar]
- Guengerich, F.P.; Johnson, W.W.; Shimada, T.; Ueng, Y.-F.; Yamazaki, H.; Langouët, S. Activation and detoxication of aflatoxin B1. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 1998, 402, 121–128. [Google Scholar]
- Hussain, Z.; Khan, M.Z.; Khan, A.; Javed, I.; Saleemi, M.K.; Mahmood, S.; Asi, M.R. Residues of aflatoxin B1 in broiler meat: Effect of age and dietary aflatoxin B1 levels. Food Chem. Toxicol. 2010, 48, 3304–3307. [Google Scholar] [CrossRef]
- Yarru, L.; Settivari, R.; Antoniou, E.; Ledoux, D.; Rottinghaus, G. Toxicological and gene expression analysis of the impact of aflatoxin B1 on hepatic function of male broiler chicks. Poult. Sci. 2009, 88, 360–371. [Google Scholar] [CrossRef]
- Fouad, A.M.; Ruan, D.; El-Senousey, H.K.; Chen, W.; Jiang, S.; Zheng, C. Harmful effects and control strategies of aflatoxin b1 produced by Aspergillus flavus and Aspergillus parasiticus strains on poultry. Toxins 2019, 11, 176. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, L.; Liu, M.; Su, Y.-T.; Xie, W.-M.; Zhang, N.-Y.; Dai, J.-F.; Wang, Y.; Rajput, S.A.; Qi, D.-S. Individual and combined occurrence of mycotoxins in feed ingredients and complete feeds in China. Toxins 2018, 10, 113. [Google Scholar] [CrossRef]
- Li, S.; Muhammad, I.; Yu, H.; Sun, X.; Zhang, X. Detection of Aflatoxin adducts as potential markers and the role of curcumin in alleviating AFB1-induced liver damage in chickens. Ecotox. Environ. Saf. 2019, 176, 137–145. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.-G.; Zhao, L.-H.; Wei, H.; Duan, G.-X.; Zhang, J.-Y.; Ji, C. Effects of lipoic acid on immune function, the antioxidant defense system, and inflammation-related genes expression of broiler chickens fed aflatoxin contaminated diets. Int. J. Mol. Sci. 2014, 15, 5649–5662. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-H.; Zhang, N.-Y.; Zhu, M.-K.; Zhao, L.; Zhou, J.-C.; Qi, D.-S. Prevention of aflatoxin B1 hepatoxicity by dietary selenium is associated with inhibition of cytochrome P450 isozymes and up-regulation of 6 selenoprotein genes in chick liver. J. Nutr. 2015, 146, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Che, C.; Korolchuk, V.I.; Gan, F.; Pan, C.; Huang, K. Selenomethionine alleviates AFB1-induced damage in primary chicken hepatocytes by inhibiting CYP450 1A5 expression via upregulated SelW expression. J. Agric. Food Chem. 2017, 65, 2495–2502. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Feng, Y.; Deng, J.; Zhang, N.-Y.; Zhang, W.-P.; Liu, X.-L.; Rajput, S.A.; Qi, D.-S.; Sun, L.-H. Selenium deficiency aggravates aflatoxin B1–induced immunotoxicity in chick spleen by regulating 6 selenoprotein genes and redox/inflammation/apoptotic signaling. J. Nutr. 2019, 149, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Zhao, L.; Zhang, N.-Y.; Karrow, N.A.; Krumm, C.S.; Qi, D.-S.; Sun, L.-H. Aflatoxin B1 metabolism: Regulation by phase I and II metabolizing enzymes and chemoprotective agents. Mutat. Res. Rev. Mutat. Res. 2018, 778, 79–89. [Google Scholar]
- Murcia, H.W.; Diaz, G.J. Protective effect of glutathione S-transferase enzyme activity against aflatoxin B1 in poultry species: Relationship between glutathione S-transferase enzyme kinetic parameters, and resistance to aflatoxin B1. Poult. Sci. 2021, 100, 101235. [Google Scholar]
- Umaya, S.R.; Vijayalakshmi, Y.; Sejian, V. Exploration of plant products and phytochemicals against aflatoxin toxicity in broiler chicken production: Present status. Toxicon 2021, 200, 55–68. [Google Scholar] [CrossRef]
- Muhammad, I.; Wang, H.; Sun, X.; Wang, X.; Han, M.; Lu, Z.; Cheng, P.; Hussain, M.A.; Zhang, X. Dual role of dietary curcumin through attenuating AFB1-induced oxidative stress and liver injury via modulating liver phase-I and phase-II enzymes involved in AFB1 bioactivation and detoxification. Front. Pharmacol. 2018, 9, 554. [Google Scholar]
- Zhang, N.-Y.; Qi, M.; Zhao, L.; Zhu, M.-K.; Guo, J.; Liu, J.; Gu, C.-Q.; Rajput, S.A.; Krumm, C.S.; Qi, D.-S. Curcumin prevents aflatoxin B1 hepatoxicity by inhibition of cytochrome P450 isozymes in chick liver. Toxins 2016, 8, 327. [Google Scholar]
- Gowda, N.K.; Ledoux, D.R.; Rottinghaus, G.E.; Bermudez, A.J.; Chen, Y.C. Antioxidant efficacy of curcuminoids from turmeric (Curcuma longa L.) powder in broiler chickens fed diets containing aflatoxin B1. Br. J. Nutr. 2009, 102, 1629–1634. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, F.; Liu, M.; Zhou, X.; Wang, M.; Cao, K.; Jin, S.; Shan, A.; Feng, X. Curcumin mitigates aflatoxin B1-induced liver injury via regulating the NLRP3 inflammasome and Nrf2 signaling pathway. Food Chem. Toxicol. 2022, 161, 112823. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, I.; Sun, X.; Wang, H.; Li, W.; Wang, X.; Cheng, P.; Li, S.; Zhang, X.; Hamid, S. Curcumin successfully inhibited the computationally identified CYP2A6 enzyme-mediated bioactivation of aflatoxin B1 in arbor acres broiler. Front. Pharmacol. 2017, 8, 143. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lin, X.; Zhang, S.; Guo, C.; Li, J.; Mi, Y.; Zhang, C. Lycopene ameliorates oxidative stress in the aging chicken ovary via activation of Nrf2/HO-1 pathway. Aging 2018, 10, 2016. [Google Scholar] [CrossRef]
- Xu, F.; Yu, K.; Yu, H.; Wang, P.; Song, M.; Xiu, C.; Li, Y. Lycopene relieves AFB1-induced liver injury through enhancing hepatic antioxidation and detoxification potential with Nrf2 activation. J. Funct. Foods 2017, 39, 215–224. [Google Scholar] [CrossRef]
- Wan, X.; Ji, H.; Ma, H.; Yang, Z.; Li, N.; Chen, X.; Chen, Y.; Yang, H.; Wang, Z. Lycopene alleviates aflatoxin B1 induced liver damage through inhibiting cytochrome 450 isozymes and improving detoxification and antioxidant systems in broiler chickens. Ital. J. Anim. Sci. 2022, 21, 31–40. [Google Scholar] [CrossRef]
- Rajput, S.A.; Sun, L.; Zhang, N.-Y.; Khalil, M.M.; Ling, Z.; Chong, L.; Wang, S.; Rajput, I.R.; Bloch, D.M.; Khan, F.A. Grape seed proanthocyanidin extract alleviates aflatoxinB1-induced immunotoxicity and oxidative stress via modulation of NF-κB and Nrf2 signaling pathways in broilers. Toxins 2019, 11, 23. [Google Scholar] [CrossRef]
- Lin, A.; Shen, S.; Wang, J.; Yan, B. Reproduction diversity of Enteromorpha prolifera. J. Integr. Plant Biol. 2008, 50, 622–629. [Google Scholar] [CrossRef]
- Zhong, R.; Wan, X.; Wang, D.; Zhao, C.; Liu, D.; Gao, L.; Wang, M.; Wu, C.; Nabavid, S.M.; Daglia, M. Polysaccharides from marine Enteromorpha: Structure and function. Trends Food Sci. Technol. 2020, 99, 11–20. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, Z.-H.; Pan, Z.-Y.; An, L.-L.; Balasubramanian, B.; Liu, W.-C. New insights into the role of dietary marine-derived polysaccharides on productive performance, egg quality, antioxidant capacity, and jejunal morphology in late-phase laying hens. Poult. Sci. 2020, 99, 2100–2107. [Google Scholar] [CrossRef]
- Liu, W.-C.; Zhuang, D.-P.; Zhao, Y.; Balasubramanian, B.; Zhao, Z.-H. Seaweed-Derived Polysaccharides Attenuate Heat Stress-Induced Splenic Oxidative Stress and Inflammatory Response via Regulating Nrf2 and NF-κB Signaling Pathways. Mar. Drugs 2022, 20, 358. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Balasubramanian, B.; Guo, Y.; Qiu, S.-J.; Jha, R.; Liu, W.-C. Dietary Enteromorpha polysaccharides supplementation improves breast muscle yield and is associated with modification of mrna transcriptome in broiler chickens. Front. Vet. Sci. 2021, 8, 337. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-C.; Ou, B.-H.; Liang, Z.-L.; Zhang, R.; Zhao, Z.-H. Algae-derived polysaccharides supplementation ameliorates heat stress-induced impairment of bursa of Fabricius via modulating NF-κB signaling pathway in broilers. Poult. Sci. 2021, 100, 101139. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.-C.; Guo, Y.; Zhihui, Z.; Jha, R.; Balasubramanian, B. Algae-derived polysaccharides promote growth performance by improving antioxidant capacity and intestinal barrier function in broiler chickens. Front. Vet. Sci. 2020, 7, 601336. [Google Scholar] [CrossRef]
- Liu, W.-C.; Zhu, Y.-R.; Zhao, Z.-H.; Jiang, P.; Yin, F.-Q. Effects of Dietary Supplementation of Algae-Derived Polysaccharides on Morphology, Tight Junctions, Antioxidant Capacity and Immune Response of Duodenum in Broilers under Heat Stress. Animals 2021, 11, 2279. [Google Scholar] [CrossRef]
- Guo, Y.; Balasubramanian, B.; Zhao, Z.-H.; Liu, W.-C. Marine algal polysaccharides alleviate aflatoxin B1-induced bursa of Fabricius injury by regulating redox and apoptotic signaling pathway in broilers. Poult. Sci. 2021, 100, 844–857. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rui, X.; Chen, S.; Li, C. Protective effects of selenium on aflatoxin B1 induced hepatic oxidative stress in broilers. Chin. J. Anim. Nutr. 2014, 26, 2281–2288. [Google Scholar]
- Zhao, L.; Deng, J.; Xu, Z.-J.; Zhang, W.-P.; Khalil, M.M.; Karrow, N.A.; Sun, L.-H. Mitigation of aflatoxin B1 hepatoxicity by dietary hedyotis diffusa is associated with activation of NRF2/ARE signaling in chicks. Antioxidants 2021, 10, 878. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Fu, P.; Chen, N.; Gao, N.; Cao, Q.; Yue, K.; Xu, T.; Zhang, C.; Zhang, C.; Liu, F. Total flavonoids of Rhizoma Drynariae protect hepatocytes against aflatoxin B1-induced oxidative stress and apoptosis in broiler chickens. Ecotox. Environ. Saf. 2022, 230, 113148. [Google Scholar] [CrossRef]
- Solis-Cruz, B.; Hernandez-Patlan, D.; Petrone, V.M.; Pontin, K.P.; Latorre, J.D.; Beyssac, E.; Hernandez-Velasco, X.; Merino-Guzman, R.; Owens, C.; Hargis, B.M. Evaluation of cellulosic polymers and curcumin to reduce aflatoxin B1 toxic effects on performance, biochemical, and immunological parameters of broiler chickens. Toxins 2019, 11, 121. [Google Scholar] [CrossRef]
- Van Deursen, V.; Damman, K.; Hillege, H.; Van Beek, A.; Van Veldhuisen, D.; Voors, A. Abnormal liver function in relation to hemodynamic profile in heart failure patients. J. Card. Fail. 2010, 16, 84–90. [Google Scholar] [CrossRef]
- Ashry, A.; Taha, N.M.; Lebda, M.A.; Abdo, W.; El-Diasty, E.M.; Fadl, S.E.; Morsi Elkamshishi, M. Ameliorative effect of nanocurcumin and Saccharomyces cell wall alone and in combination against aflatoxicosis in broilers. BMC Vet. Res. 2022, 18, 178. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, X.; Zou, K.; Xie, J.; Zhao, S.; Liu, J.; Liu, H.; Wang, J.; Wang, Y. Seabuckthorn berry polysaccharide protects against carbon tetrachloride-induced hepatotoxicity in mice via anti-oxidative and anti-inflammatory activities. Food Funct. 2017, 8, 3130–3138. [Google Scholar] [CrossRef]
- Gan, F.; Yang, Y.; Chen, Y.; Che, C.; Pan, C.; Huang, K. Bush sophora root polysaccharide could help prevent aflatoxin B1-induced hepatotoxicity in the primary chicken hepatocytes. Toxicon 2018, 150, 180–187. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Yamamoto, T.; Suzuki, T.; Kobayashi, A.; Wakabayashi, J.; Maher, J.; Motohashi, H.; Yamamoto, M. Physiological significance of reactive cysteine residues of Keap1 in determining Nrf2 activity. Mol. Cell Biol. 2008, 28, 2758–2770. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, H.; Yue, H.; Wu, S.; Yang, H.; Qi, G.; Wang, Z. Low-current & high-frequency electrical stunning increased oxidative stress, lipid peroxidation, and gene transcription of the mitogen-activated protein kinase/nuclear factor-erythroid 2-related factor 2/antioxidant responsive element (MAPK/Nrf2/ARE) signaling pathway in breast muscle of broilers. Food Chem. 2018, 242, 491–496. [Google Scholar]
- Bao, J.; Ding, R.; Zou, L.; Zhang, C.; Wang, K.; Liu, F.; Li, P.; Chen, M.; Wan, J.-B.; Su, H. Forsythiae fructus inhibits B16 melanoma growth involving MAPKs/Nrf2/HO-1 mediated anti-oxidation and anti-inflammation. Am. J. Chin. Med. 2016, 44, 1043–1061. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, Z.; Zhang, K.; Ding, X.; Bai, S.; Zeng, Q.; Peng, H.; Celi, P. Epigallocatechin-3-gallate protected vanadium-induced eggshell depigmentation via P38MAPK-Nrf2/HO-1 signaling pathway in laying hens. Poult. Sci. 2018, 97, 3109–3118. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | CON | AFB1 | AFB1 + SDP | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|---|
| ANOVA | CON vs. AFB1 | AFB1 vs. AFB1 + SDP | CON vs. AFB1 + SDP | |||||
| ADG, g | 21.34 | 20.77 | 21.06 | 0.41 | 0.629 | 0.348 | 0.626 | 0.640 |
| ADFI, g | 75.05 | 75.42 | 73.96 | 1.31 | 0.732 | 0.846 | 0.458 | 0.579 |
| F/G | 3.51 | 3.63 | 3.54 | 0.04 | 0.152 | 0.089 | 0.098 | 0.928 |
| Items | CON | AFB1 | AFB1 + SDP | SEM | p-Value | |||
|---|---|---|---|---|---|---|---|---|
| ANOVA | CON vs. AFB1 | AFB1 vs. AFB1 + SDP | CON vs. AFB1 + SDP | |||||
| ALT activity, U/L | 6.68 | 8.05 | 6.82 | 0.35 | 0.038 | 0.020 | 0.033 | 0.782 |
| AST activity, U/L | 232.72 | 272.59 | 249.48 | 6.60 | 0.005 | 0.002 | 0.032 | 0.103 |
| TP content, g/L | 40.55 | 44.28 | 43.01 | 1.49 | 0.243 | 0.106 | 0.558 | 0.269 |
| Relative liver weight, % | 2.06 | 2.35 | 2.12 | 0.07 | 0.049 | 0.020 | 0.045 | 0.683 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.-C.; Yang, Y.-Y.; Pushparaj, K.; Balasubramanian, B. Evaluation of Hepatic Detoxification Effects of Enteromorpha prolifera Polysaccharides against Aflatoxin B1 in Broiler Chickens. Antioxidants 2022, 11, 1757. https://doi.org/10.3390/antiox11091757
Liu W-C, Yang Y-Y, Pushparaj K, Balasubramanian B. Evaluation of Hepatic Detoxification Effects of Enteromorpha prolifera Polysaccharides against Aflatoxin B1 in Broiler Chickens. Antioxidants. 2022; 11(9):1757. https://doi.org/10.3390/antiox11091757
Chicago/Turabian StyleLiu, Wen-Chao, Yu-Ying Yang, Karthika Pushparaj, and Balamuralikrishnan Balasubramanian. 2022. "Evaluation of Hepatic Detoxification Effects of Enteromorpha prolifera Polysaccharides against Aflatoxin B1 in Broiler Chickens" Antioxidants 11, no. 9: 1757. https://doi.org/10.3390/antiox11091757