
The Azadirachta indica (Neem) Seed Oil Reduced Chronic Redox-Homeostasis Imbalance in a Mice Experimental Model on Ochratoxine A-Induced Hepatotoxicity
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Animals, Experimental Design, and Ethical Approval
2.3. Dissection Procedure
2.4. Histological Analysis for Visualization of Liver Changes
2.5. Analysis of Hydroxyproline (Hyp) in Liver Tissue
2.6. Determination of Glutathione S-Transferase (GST)
2.7. Experimental Protocol for Functional Markers Measurement of Hepatic Damage
2.8. Lipid Peroxidation Analysis
2.9. NO Radical Formations
2.10. 3-Maleimido proxyl (5-MSL) Protein Oxidation in the Liver
2.11. Immunoenzyme Assays
2.11.1. Determination of the Activity of Antioxidant Enzyme System and Products of Oxidation of Proteins and Lipids in the Liver
2.11.2. Measurement of Proinflammatory Markers in Liver Tissue and Blood
2.12. Statistical Analysis
3. Results
3.1. Physiological Status
3.2. Liver Histopathology
3.3. Functional Markers of Hepatic Damage
3.4. The Effects on SIRT1/PGC-1α Pathway
3.5. Analysis of Hydroxyproline (Hyp) and MDA in Liver Tissue
3.6. Effect of Antioxidant Enzymes
3.7. Determination of Oxidative Protein Remodeling in Liver Tissue
3.8. Parameters of Oxidative Damage in Liver Tissue
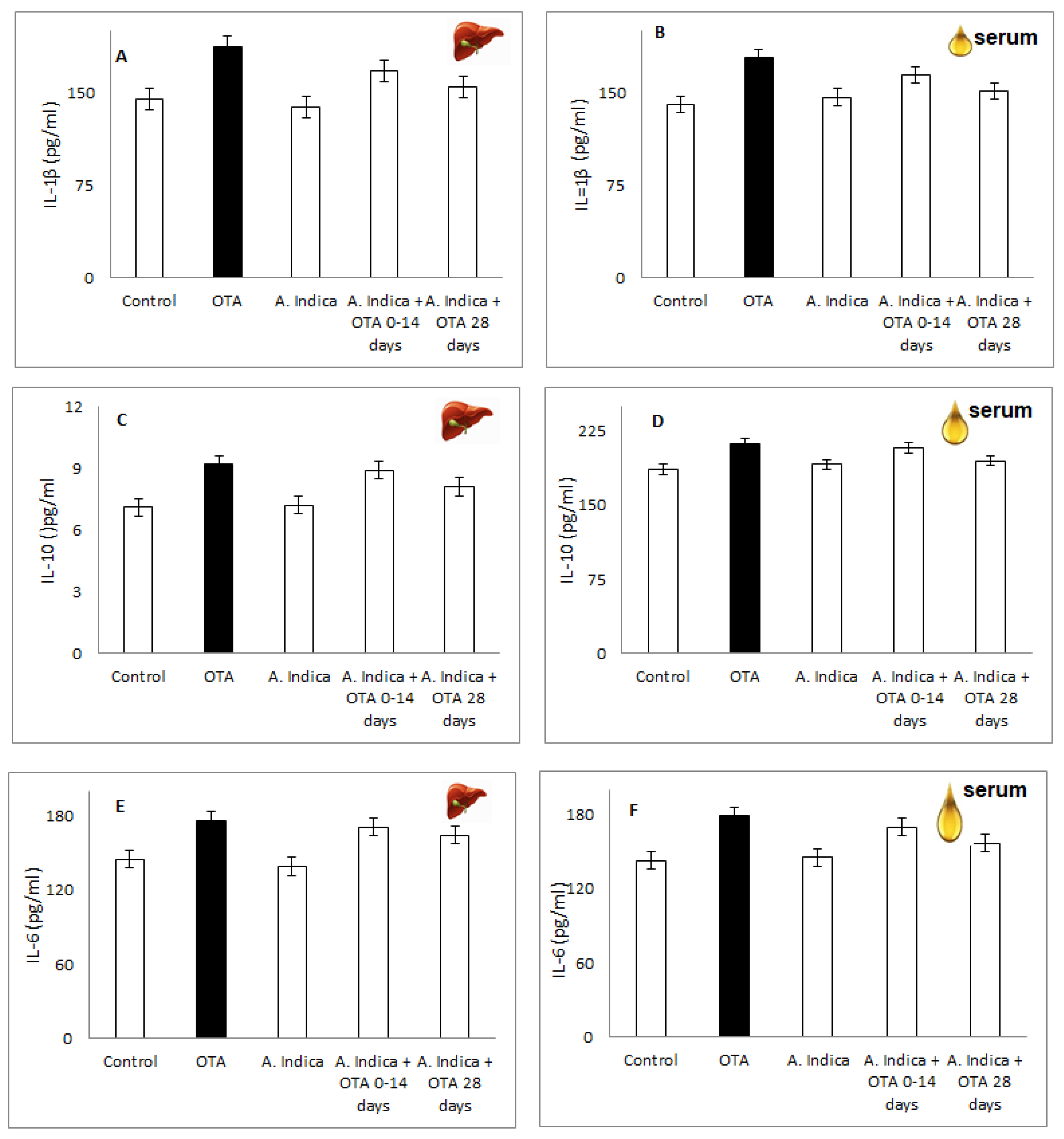
3.9. IL-1b, IL-6, IL-10 in Liver Homogenate/Serum of OTA-Injured Group, Compare to Group Protected with A. indica Oil Alone, Group with Combination Oil + OTA and Control
3.10. TNF-a and INF-γ Concentration of A. indica Oil Protected Group Treated with OTA, with A. indica oil Alone and OTA-Damaged Group Compared to Healthy Mice
3.11. Correlations between Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhukan, P.G.; Bangar, N.P.; Ramesh, D. Review on Hepatoprotective Medicinal. Plants 2020, 9, 531–549. [Google Scholar] [CrossRef]
- Huang, L.; Liu, J.; Bie, C.; Liu, H.; Ji, Y.; Chen, D.; Zhu, M.; Kuang, W. Advances in cell death-related signaling pathways in acute-on-chronic liver failure. Clin. Res. Hepatol. Gastroenterol. 2022, 46, 101783. [Google Scholar] [CrossRef] [PubMed]
- Atanasova-Penichon, V.; Legoahec, L.; Bernillon, S.; Deborde, C.; Maucourt, M.; Verdal-Bonnin, M.; Richard-Forget, F. Mycotoxin biosynthesis and central metabolism are two interlinked pathways in Fusarium graminearum, as demonstrated by the extensive metabolic changes induced by caffeic acid exposure. Appl. Environ. Microb. 2018, 84, 01705–01717. [Google Scholar] [CrossRef] [PubMed]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef]
- Wang, L.; Jin, J.; Liu, X.; Wang, Y.; Liu, Y.; Zhao, Y.; Xing, F. Effect of cinnamaldehyde on morphological alterations of Aspergillus ochraceus and expression of key genes involved in ochratoxin A biosynthesis. Toxins 2018, 10, 340. [Google Scholar] [CrossRef]
- Zeidan, R.; Ul-Hassan, Z.; Al-Thani, R.; Migheli, Q.; Jaoua, S. In-Vitro Application of a Qatari Burkholderia cepacia strain (QBC03) in the Biocontrol of Mycotoxigenic Fungi and in the Reduction of Ochratoxin A biosynthesis by Aspergillus carbonarius. Toxins 2019, 11, 700. [Google Scholar] [CrossRef]
- Benkerroum, N. Chronic and Acute Toxicities of Aflatoxins: Mechanisms of Action. Int. J. Environ. Res. Public Health 2020, 17, 423. [Google Scholar] [CrossRef]
- Khaneghah, M.A.; Ismail, E.; Raeisi, S.; Fakhri, Y. Aflatoxins in cereals: State of the art. J. Food Saf. 2018, 38, e12532. [Google Scholar] [CrossRef]
- Petzinger, E.; Ziegler, K. Ochratoxin A from a toxicological perspective. J. Vet. Pharmacol. Ther. 2000, 23, 91–98. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A.; Bartsc, H.; Azémar, B. MESNA protects rats against nephrotoxicity but not carcinogenicity induced by ochratoxin A, implicating two separate pathways. Facta Univ. Ser. Med. Biol. 2002, 9, 57–63. [Google Scholar]
- Petkova-Bocharova, T.; Castegnaro, M. Ochratoxin A contamination of cereals in an area of high incidence of Balkan endemic nephropathy in Bulgaria. Food Addit. Contam. 1985, 2, 267–270. [Google Scholar] [CrossRef]
- Imaoka, T.; Yang, J.; Wang, L.; McDonald, M.; Afsharinejad, Z.; Bammler, T.; Kelly, E. Microphysiological system modeling of ochratoxin A-associated nephrotoxicity. Toxicology 2020, 444, 152582. [Google Scholar] [CrossRef]
- Gagliano, N.; Donne, I.D.; Torr, C.; Migliori, M.; Grizzi, F.; Milzani, A.; Filippi, C.; Annoni, G.; Colombo, P.; Costa, F. Early cytotoxic effects of ochratoxin A in rat liver: A morphological, biochemical and molecular study. Toxicology 2006, 15, 214–224. [Google Scholar] [CrossRef]
- Biney, E.E.; Nkoom, M.; Darkwah, W.K.; Puplampu, J.B. High-Performance Liquid Chromatography Analysis and Antioxidant Activities of Extract of Azadirachta indica (Neem) Leaves. Pharmacogn. Res. 2021, 12, 29–34. [Google Scholar] [CrossRef]
- Bakr, A. Changes of hemoglobin content and glucose levels in the blood of Rattus norvegicus by water extracts of Azadirachta indica. Chin. J. Nat. Med. 2012, 10, 135–137. [Google Scholar] [CrossRef]
- Sen, S.; Chakraborty, R. The role of antioxidants in human health. In Oxidative Stress: Diagnostics, Prevention, and Therapy; ACS Publications: Washington, DC, USA, 2011; pp. 1–37. [Google Scholar] [CrossRef]
- Parcheta, M.; Świsłocka, R.; Orzechowska, S.; Akimowicz, M.; Choińska, R.; Lewandowski, W. Recent Developments in Effective Antioxidants: The Structure and Antioxidant Properties. Materials 2021, 14, 1984. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxidative Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Smirne, C.; Croce, E.; Di Benedetto, D.; Cantaluppi, V.; Comi, C.; Sainaghi, P.P.; Minisini, R.; Grossini, E.; Pirisi, M. Oxidative Stress in Non-Alcoholic Fatty Liver Disease. Livers 2022, 2, 30–76. [Google Scholar] [CrossRef]
- Spector, A. Oxidative stress and disease. J. Ocular Pharmacol. Ther. 2000, 16, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Huang, J.; Nisar, M.F.; Wan, C.; Huang, W. The beneficial roles of SIRT1 in drug-induced liver injury. Oxidative Med. Cell. Longev. 2019, 2019, 8506195. [Google Scholar] [CrossRef] [PubMed]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Auwerx, J. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; He, H.; Zuo, Z.; Xu, Z.; Wei, Z.; Deng, J. The role of different SIRT1-mediated signaling pathways in toxic injury. Cell. Mol. Biol. Lett. 2019, 24, 36. [Google Scholar] [CrossRef]
- Deng, Y.X.; Cao, M.; Shi, D.X.; Yin, Z.Q.; Jia, R.Y.; Xu, J.; Zhao, J. Toxicological evaluation of neem (Azadirachta indica) oil: Acute 690 and subacute toxicity. Environ. Toxicol. Pharmacol. 2013, 35, 240–246. [Google Scholar] [CrossRef]
- Stoev, S.D.; Dimitrov, K.; Zarkov, I.; Mircheva, T.; Zapryanova, D.; Valchev, I.; Arora, R. Some Indian herbs have protective effects against deleterious effects of ochratoxin A in broiler chicks. World Mycotoxin J. 2021, 14, 525–538. [Google Scholar] [CrossRef]
- Woessner, J.B. The determination of hidroxyproline in tissue and protein samples containing small proportions of this amino acid. Arch. Biochem. Biophys. 1961, 93, 440–447. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Novaes, R.D.; Gonçalves, R.V.; Marques, D.C.; Cupertino, M.C.; Peluzio, M.C.; Leite, J.P.; Maldonado, I.R. Effect of bark extract of Bathysa cuspidata on hepatic oxidative damage and blood glucose kinetics in rats exposed to paraquat. Toxicol. Pathol. 2012, 40, 62–70. [Google Scholar] [CrossRef]
- Novaes, R.D.; Santos, E.C.; Cupertino, M.C.; Bastos, D.S.; Oliveira, J.M.; Carvalho, T.V.; Neves, M.M.; Oliveira, L.L.; Talvani, A. Trypanosoma cruzi infection and benznidazole therapy independently stimulate oxidative status and structural pathological remodeling of the liver tissue in mice. Parasitol. Res. 2015, 114, 2873–2881. [Google Scholar] [CrossRef]
- Shi, H.; Sui, Y.; Wang, X.; Luo, Y.; Ji, L. Hydroxyl radical production and oxidative damage induced by cadmium and naphthalene in liver of Carassius auratus. Comp. Biochem. Phys. Part C Toxicol. Pharmacol. 2005, 140, 115–121. [Google Scholar] [CrossRef]
- Yokoyama, K.; Hashiba, K.; Wakabayashi, H.; Hashimoto, K.; Saton, K.; Kurihara, T.; Motohashi, N.; Sakagami, H. Inhibition of LPS-stimulated NO production in mouse macrophage-like cells by Tropolones. Anticancer Res. 2004, 24, 3917–3922. [Google Scholar]
- Yoshioka, T.; Iwamoto, N.; Lto, K. An applicationof Electron Paramagnetic Resonance to evaluate nitric oxide and its quenchers. J. Am. Soc. Nephrol. 1996, 7, 961–965. [Google Scholar] [CrossRef]
- Takeshita, K.; Saito, K.; Ueda, J.I.; Anzai, K.; Ozawa, T. Kinetic study on ESR signal decay of nitroxyl radicals, potent redox probes for in vivo ESR spectroscopy, caused by reactive oxygen species. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2002, 1573, 156–164. [Google Scholar] [CrossRef]
- Momchilova, S.; Antonova, D.; Marekov, I.; Kuleva, L.; Nikolova-Damyanova, B. Fatty acids, triacylglycerols, and sterols in neem oil (Azadirachta indica a. juss) as determined by a combination of chromatographic and spectral techniques. J. Liq. Chromatogr. Relat. Technol. 2007, 30, 11–25. [Google Scholar] [CrossRef]
- Mavrommatis, A.; Giamouri, E.; Tavrizelou, S.; Zacharioudaki, M.; Danezis, G.; Simitzis, P.E.; Feggeros, K. Impact of mycotoxins on animals’ oxidative status. Antioxidants 2021, 10, 214. [Google Scholar] [CrossRef]
- Chienwichai, P.; Reamtong, O.; Pisitkun, T.; Somparn, P.; Tharnpoophasiam, P.; Worakhunpiset, S.; Topanurak, S. Hepatic protein Carbonylation profiles induced by lipid accumulation and oxidative stress for investigating cellular response to non-alcoholic fatty liver disease in vitro. Proteome Sci. 2019, 17, 1–13. [Google Scholar] [CrossRef]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Yuan, Z. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef]
- Gray, J.P.; Mishin, V.; Heck, D.E.; Laskin, D.L.; Laskin, J.D. Inhibition of NADPH cytochrome P450 reductase by the model sulfur mustard vesicant 2-chloroethyl ethyl sulfide is associated with increased production of reactive oxygen species. Toxicol. Appl. Pharmacol. 2010, 247, 76–82. [Google Scholar] [CrossRef]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell Molecimmunol. 2016, 13, 267–276. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, L.; Sandhu, J.K.; Harper, M.E.; Cuperlovic-Culf, M. Role of glutathione in cancer: From mechanisms to therapies. Biomolecules 2020, 10, 1429. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.A.; Scheff, S.W. Oxidative stress in the progression of Alzheimer disease in the frontal cortex. J. Neuropathol. Exp. Neurol. 2010, 69, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Dhanasekaran, M.; Ignacimuthu, S.; Agastian, P. Potential hepatoprotective activity of ononitol monohydrate isolated from Cassia tora L. on carbon tetrachloride induced hepatotoxicity in wistar rats. Phytomedicine 2009, 16, 891–895. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxidative Med. Cell Longev. 2019, 5, 6175804. [Google Scholar] [CrossRef]
- Nabi, S.; Tanveer, S.; Ganie, S.A. Glutathione-S-transferase, Superoxide Dismutase (GST, SOD) Levels, Protein Content and Lipid Peroxidation in Schizothorax plagiostomus under the Infection of Pomphorhynchus in Nallah Sukhnag of Kashmir Valley. Pak. J. Biol. Sci. 2017, 20, 442–446. [Google Scholar] [CrossRef]
- Zhu, L.; Yu, T.; Qi, X.; Gao, J.; Huang, K.; He, X.; Luo, H.; Xu, W. Limited link between oxidative stress and ochratoxin A—Induced renal injury in an acute toxicity rat model. Toxins 2016, 8, 373. [Google Scholar] [CrossRef]
- Marin-Kuan, M.; Ehrlich, V.; Delatour, T.; Cavin, C.; Schilter, B. Evidence for a role of oxidative stress in the carcinogenicity of ochratoxin A. J. Toxicol. 2011, 2011, 645361. [Google Scholar] [CrossRef]
- Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Bignami, M. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, e06113. [Google Scholar] [CrossRef]
- Baligar, N.S.; Aladakatti, R.H.; Ahmed, M.; Hiremath, M.B. Hepatoprotective activity of the neem-based constituent azadirachtin-A in carbon tetrachloride intoxicated Wistar rats. Can. J. Physiol. Pharmacol. 2014, 92, 267–277. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Al-Quraishy, S.; Aref, A.M.; Othman, M.S.; El-Deib, K.M.; Abdel Moneim, A.E. The potential role of Azadirachta indica treatment on cisplatin-induced hepatotoxicity and oxidative stress in female rats. Oxidative Med. Cell Longev. 2013, 2013, 741817. [Google Scholar] [CrossRef] [Green Version]
- Yanpallewar, S.U.; Sen, S.; Tapas, S.; Kumar, M.; Raju, S.S.; Acharya, S.B. Effect of Azadirachta indica on paracetamol-induced hepatic damage in albino rats. Phytomedicine 2003, 10, 391–396. [Google Scholar] [CrossRef]
- Bhanwra, S.; Singh, J.; Khosla, P. Effect of Azadirachta indica (neem) leaf aqueous extract on paracetamol induced liver damage in rats. Ind. J. Physiol. Pharmacol. 2000, 44, 64–68. [Google Scholar]
- Alzohairy, M.A. Therapeutics Role of Azadirachta indica (Neem) and Their Active Constituents in Diseases Prevention and Treatment. Evid.-Based Complementary Altern. Med. 2016, 2016, 7382506. [Google Scholar] [CrossRef]
- Rao, N.G.; Rao, P.P.G.; Satyanarayana, A. Chemical, fatty acid, volatile oil composition and antioxidant activity of shade dried neem (Azadirachta indica L.) flower powder. Int. Food Res. J. 2014, 21, 807–813. [Google Scholar]
- Gossé, B.; Amissa, A.A.; Adjé, A.F.; Niamké, B.F.; Ollivier, D.; Ito, Y. Analysis of components of neem (Azadirachta indica) oil by diverse chromatographic techniques. J. Liquid Chromatogr. Relat. Technol. 2005, 28, 2225–2233. [Google Scholar] [CrossRef]
- Papackova, Z.; Cahova, M. Fatty acid signaling: The new function of intracellular lipases. Int. J. Mol. Sci. 2015, 16, 3831–3855. [Google Scholar] [CrossRef]
- Baligar, N.; Aladakatti, R.; Ahmed, M.; Hiremath, M.B. Evaluation of acute toxicity of neem active constituent, nimbolide and its hepatoprotective activity against acute dose of carbon tetrachloride treated albino rats. Int. J. Pharm. Sci. Res. 2014, 5, 3455–3466. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Stela, M.; Saluk-Bijak, J.; Siadkowski, A.; Bijak, M. Molecular Aspects of Mycotoxins-A Serious Problem for Human Health. Int. J. Mol. Sci. 2020, 21, 8187. [Google Scholar] [CrossRef]
- Niaz, K.; Shah, S.Z.A.; Khan, F.; Bule, M. Ochratoxin A–induced genotoxic and epigenetic mechanisms lead to Alzheimer disease: Its modulation with strategies. Environ. Sci. Pollut. Res. 2020, 27, 44673–44700. [Google Scholar] [CrossRef]
- Shin, M.-R.; Lee, J.A.; Kim, M.; Lee, S.; Oh, M.; Moon, J.; Nam, J.-W.; Choi, H.; Mun, Y.-J.; Roh, S.-S. Gardeniae Fructus Attenuates Thioacetamide-Induced Liver Fibrosis in Mice via Both AMPK/SIRT1/NF-κB Pathway and Nrf2 Signaling. Antioxidants 2021, 10, 1837. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A. Colombo R Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Shin, H.S.; Lee, H.J.; Pyo, M.C.; Ryu, D.; Lee, K.-W. Ochratoxin A-Induced Hepatotoxicity through Phase I and Phase II Reactions Regulated by AhR in Liver Cells. Toxins 2019, 11, 377. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.P.; Astoreca, A.L.; Oliveira, Á.A.D.; Salvato, L.A.; Biscoto, G.L.; Keller, L.A.M.; Keller, K.M. In vitro activity of neem (Azadirachta indica) oil on growth and ochratoxin A production by Aspergillus carbonarius Isolates. Toxins 2019, 11, 579. [Google Scholar] [CrossRef]
- Schumacher, M.; Cerella, C.; Reuter, S.; Dicato, M.; Diederich, M. Anti-inflammatory, pro-apoptotic, and anti-proliferative effects of a methanolic neem (Azadirachta indica) leaf extract are mediated via modulation of the nuclear factor-κB pathway. Genes Nutr. 2011, 6, 149–160. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Administration | BW (g) | BW (g) | BW (g) | BW (g) |
|---|---|---|---|---|
| (n = 6) | 5th day | 8th day | 14th day | 28th day |
| Control | 205.3 ± 2.98 | 221.7 ± 1.87 | 233.1 ± 2.11 | 255.4 ± 2.82 |
| OTA | 189.1± 1.99 * | 175.4± 2.14 * | 165.4 ± 2.03 * | 157.8 ± 3.01 * |
| A. indica oil | 204.8 ± 3.11 | 224.4 ± 2.01 | 232.1 ± 2.25 | 254.9 ± 2.14 |
| A. indica oil + OTA | 199.1 ± 188 | 218.1 ± 2.31 ** | 231.9 ± 2.09 ** | 252.9 ± 1.05 ** |
| Controls | A. indica | A. indica + OTA | OTA | |
|---|---|---|---|---|
| Fat degeneration | 0 | 0 | 0 | 0 |
| Hyaline drops | 0 | 0 | 0 | 0 |
| Inflammation | 0 | 0 | 1 | 2 |
| Necrosis | 0 | 0 | 0 | 1 |
| Vascular congestion | 0 | 0 | 1 | 2 |
| Karyomegaly | 0 | 0 | 1 | 2 |
| Aneuploidy | 0 | 0 | 0 | 1 |
| Polychromasia | 0 | 0 | 0 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolova, G.; Ananiev, J.; Ivanov, V.; Petkova-Parlapanska, K.; Georgieva, E.; Karamalakova, Y. The Azadirachta indica (Neem) Seed Oil Reduced Chronic Redox-Homeostasis Imbalance in a Mice Experimental Model on Ochratoxine A-Induced Hepatotoxicity. Antioxidants 2022, 11, 1678. https://doi.org/10.3390/antiox11091678
Nikolova G, Ananiev J, Ivanov V, Petkova-Parlapanska K, Georgieva E, Karamalakova Y. The Azadirachta indica (Neem) Seed Oil Reduced Chronic Redox-Homeostasis Imbalance in a Mice Experimental Model on Ochratoxine A-Induced Hepatotoxicity. Antioxidants. 2022; 11(9):1678. https://doi.org/10.3390/antiox11091678
Chicago/Turabian StyleNikolova, Galina, Julian Ananiev, Veselin Ivanov, Kamelia Petkova-Parlapanska, Ekaterina Georgieva, and Yanka Karamalakova. 2022. "The Azadirachta indica (Neem) Seed Oil Reduced Chronic Redox-Homeostasis Imbalance in a Mice Experimental Model on Ochratoxine A-Induced Hepatotoxicity" Antioxidants 11, no. 9: 1678. https://doi.org/10.3390/antiox11091678