
Ethnopharmacological Effects of Urtica dioica, Matricaria chamomilla, and Murraya koenigii on Rotenone-Exposed D. melanogaster: An Attenuation of Cellular, Biochemical, and Organismal Markers


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Materials Used in the Study
2.2.1. Drying, Processing, and Extraction of Samples
2.2.2. Determination of Plant Yield
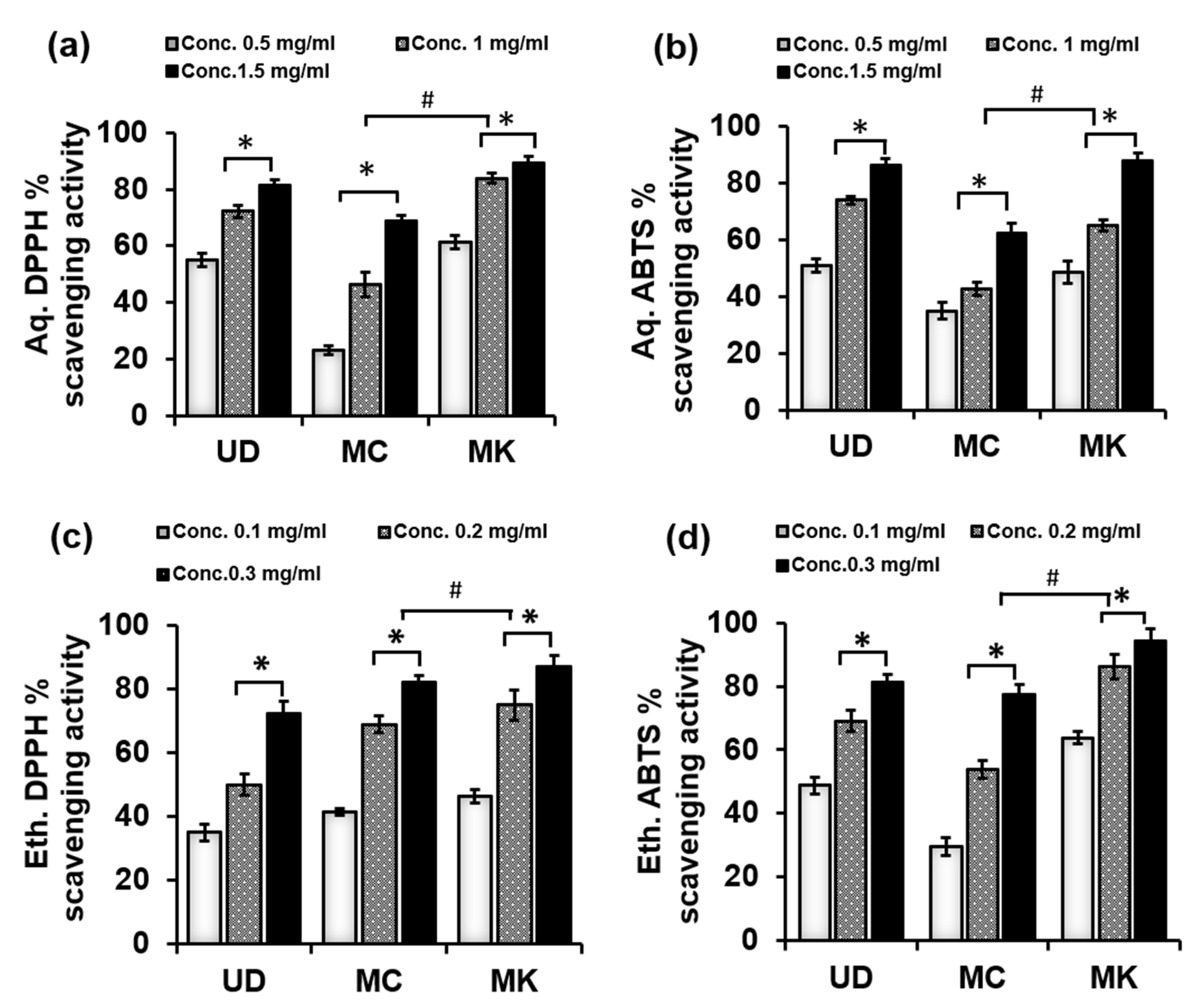
2.2.3. Free Radical Scavenging Activity Using the DPPH Radical Assay
2.2.4. ABTS Radical Cation Decolorization Assay
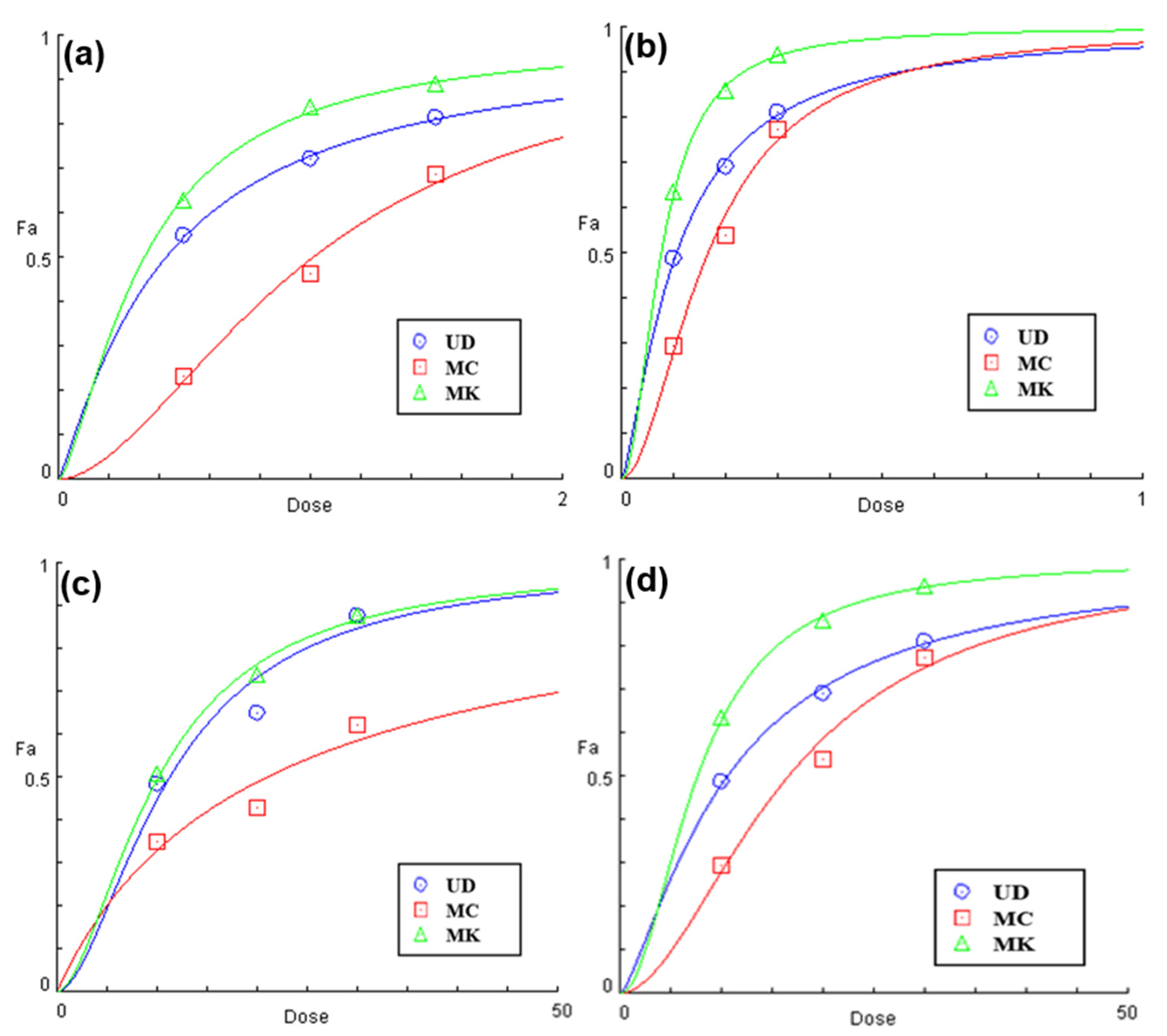
2.2.5. EC50 (Dose-Response Relationship)
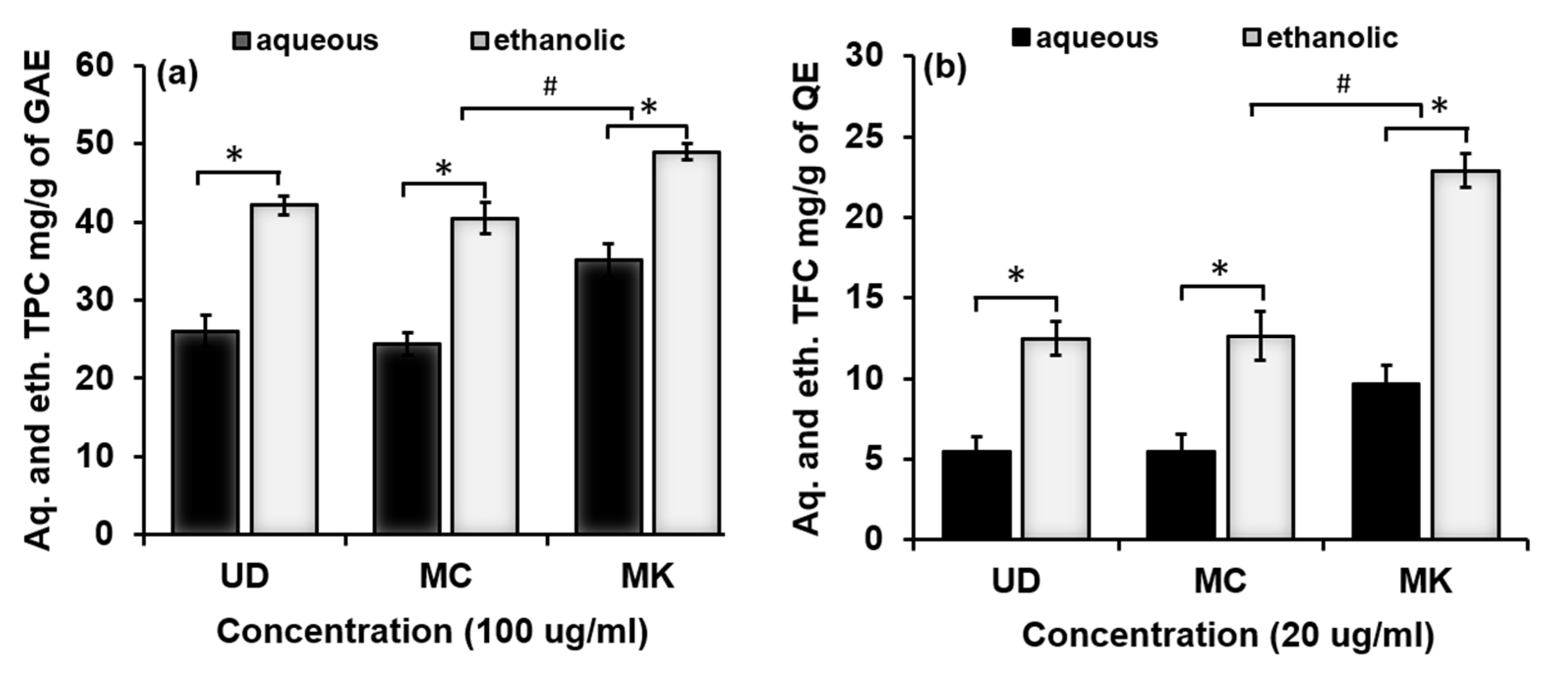
2.2.6. Determination of Total Phenolic Content (TPC)
2.2.7. Determination of Total Flavonoid Content (TFC)
2.2.8. Preliminary Qualitative Screening Analysis of Plant Extracts
- (a)
- Detection of phenols (ferric chloride test): An amount of 2 mL of plant extract was combined in a test tube with 2 mL of 5% ferric chloride aqueous solution. The presence of phenols was indicated by a deep blue-green solution [32].
- (b)
- Detection of flavonoids (Alkaline reagent test): A few drops of NaOH (20%) solution were added to 2 mL of plant extract, which displayed a yellowish red color within a second and turned transparent with the addition of diluted HCl, displaying a positive result [33].
- (c)
- Detection of alkaloids (Wagner’s test): To 4 mL of plant extract, 3 drops of Wagner’s reagent were added. The appearance of a reddish-brown precipitate indicated a positive outcome [33].
- (d)
- Detection of tannins (FeCl3 solution test): An alcoholic ferric chloride (10%) solution was added to 2 mL of plant extract. The appearance of the blue/green color suggested a positive outcome [33].
- (e)
- Detection of carbohydrates (Molisch’s test): An amount of 3 mL of extract and 3 mL of H2SO4 were placed in a test tube; a few drops of Molisch’s reagent were added (conc.). Allowing it to stand 3 min, the appearance of a red/dull violet tone at the interphase of the two layers showed a successful outcome [32].
- (f)
- Detection of saponins (saponin foam test): Five milliliters of distilled water was combined with 500 µL of plant fractions. The presence of saponins is indicated by foaming (formation of creamy tiny bubbles) [34].
- (g)
- Detection of terpenoids (Salkowski test): An amount of 2 mL of extract and a few drops of conc. H2SO4 was mixed with 1 mL of chloroform. The appearance of a reddish-brown precipitate indicated a positive outcome [32].
- (h)
- Detection of steroids (Liebermann-Burchard test): An amount of 3 mL of acetic anhydride was mixed with 5 mL of plant extract. Then, 2 mL of H2SO4 was added to it. The presence of steroids was indicated by a shift in color from violet to bluish green [34].
- (i)
- Detection of glycosides (Kellar-Kiliani test): An amount of 1 mL of glacial acetic acid was mixed with 2 mL of plant extract. Then, 1 mL of FeCl3 and 1 mL of (conc.) H2SO4 was mixed into it. The appearance of glycosides was confirmed by the solution’s greenish-blue color [34].
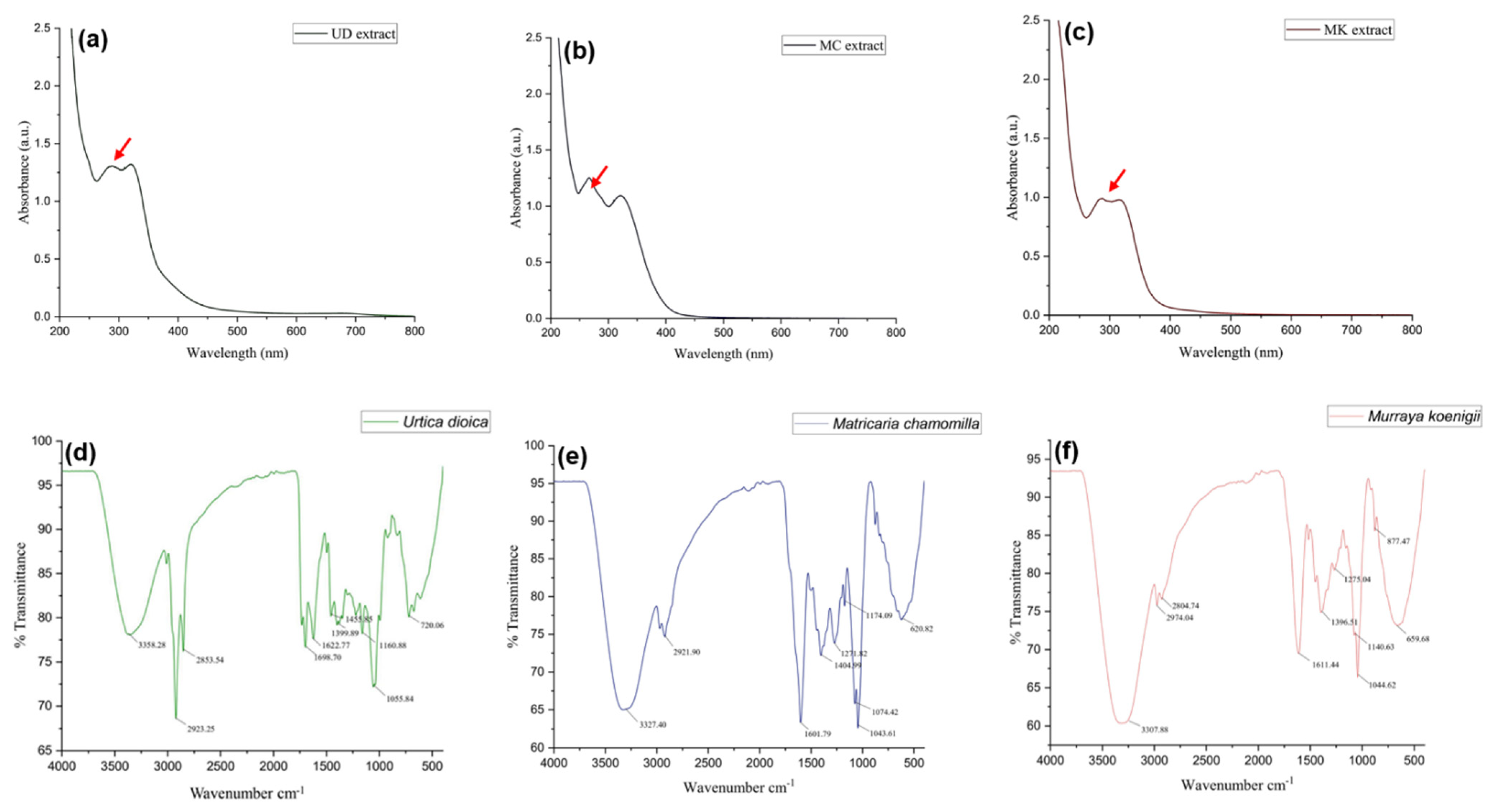
2.2.9. UV-Visible Spectroscopic Analysis
2.2.10. FT-IR Spectroscopic Analysis
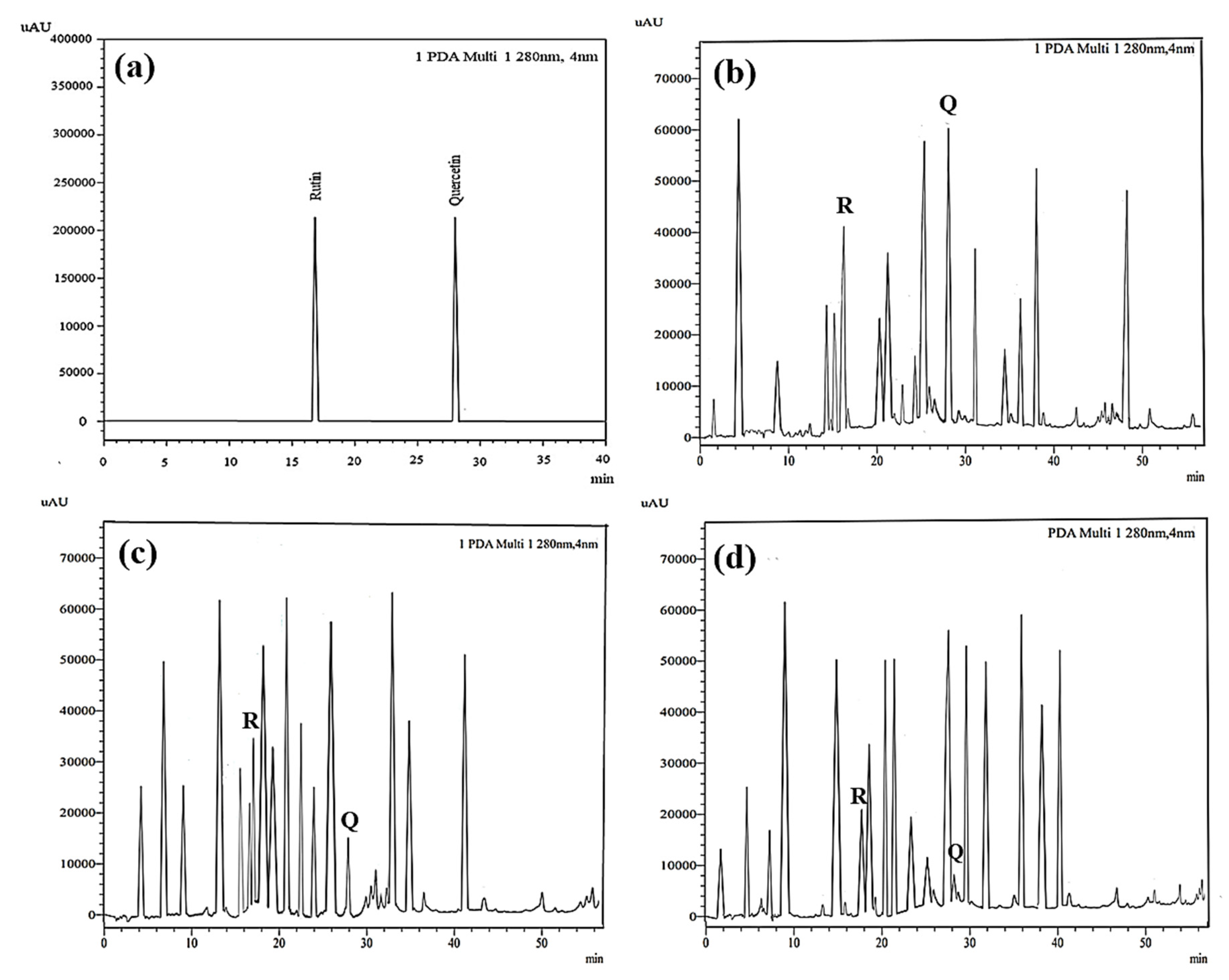
2.2.11. HPLC Chromatographic Analysis
2.3. Fly Strain and Maintenance of Culture
2.3.1. Plant Concentration and Rotenone Exposure
2.3.2. Treatment Schedule
2.3.3. Trypan Blue Dye Exclusion Assay
2.3.4. Homogenate Preparation
2.3.5. Protein Estimation
2.3.6. Acetylcholinesterase (AChE) Enzymatic Assay
2.3.7. Measurement of Locomotor Deficits: Climbing Assay
2.3.8. Jumping Assay
2.4. Statistical Analysis
3. Results
3.1. Analytical Assays
3.1.1. Percentage Yield of Bioactive Compounds
3.1.2. Antioxidant Potential of UD, MC, and MK
3.1.3. EC50 Prediction Using Statistical Models
3.1.4. Phenolic and Flavonoid Potential of UD, MC, and MK
3.1.5. Preliminary Qualitative Screening Analysis of Plant Extracts
3.1.6. UV-Visible Analysis
3.1.7. Fourier Transform Infrared Spectrophotometer (FTIR)
3.1.8. High-Performance Liquid Chromatography (HPLC)
3.2. Cellular Assays
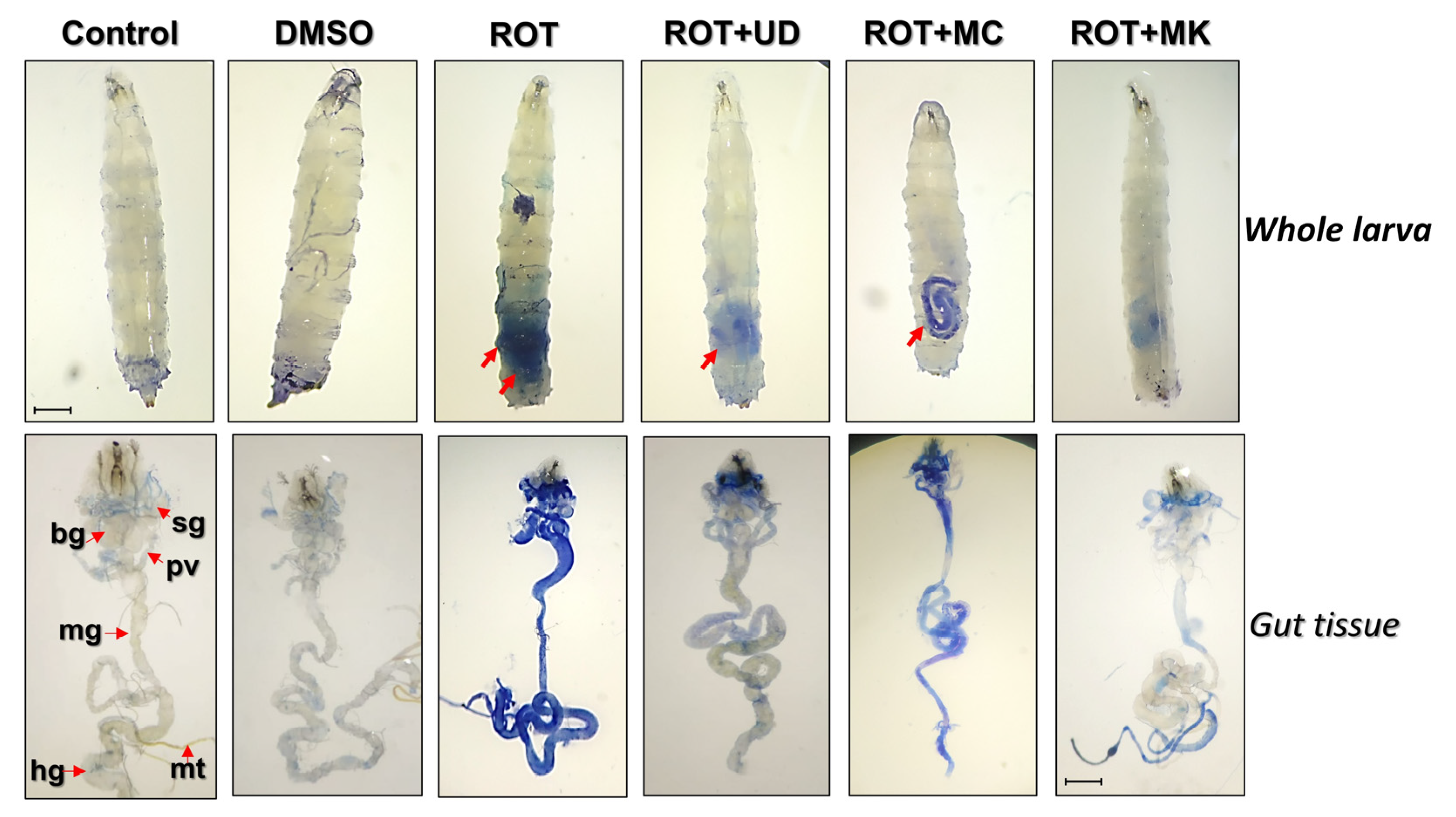
Cytotoxicity of Rotenone and Amelioration of Cytotoxicity through Bioactive Compounds UD, MC, and MK Determined by a Dye Exclusion Test (Trypan Blue) in Whole Larvae and Tissues of ROT-Exposed Organisms
3.3. Biochemical Assays
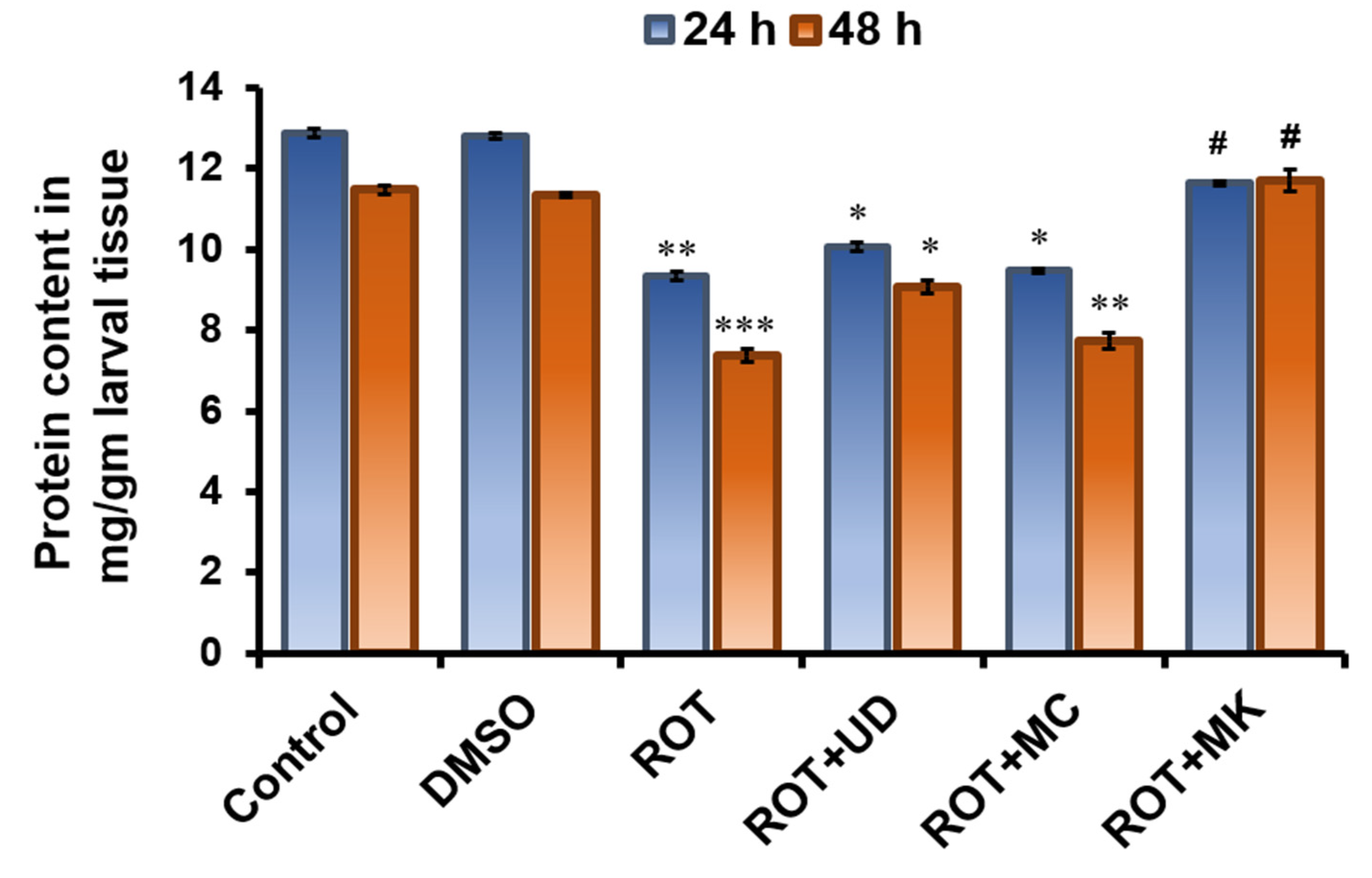
3.3.1. Decreased Protein Content in D. melanogaster Treated with Rotenone after 24 and 48 h
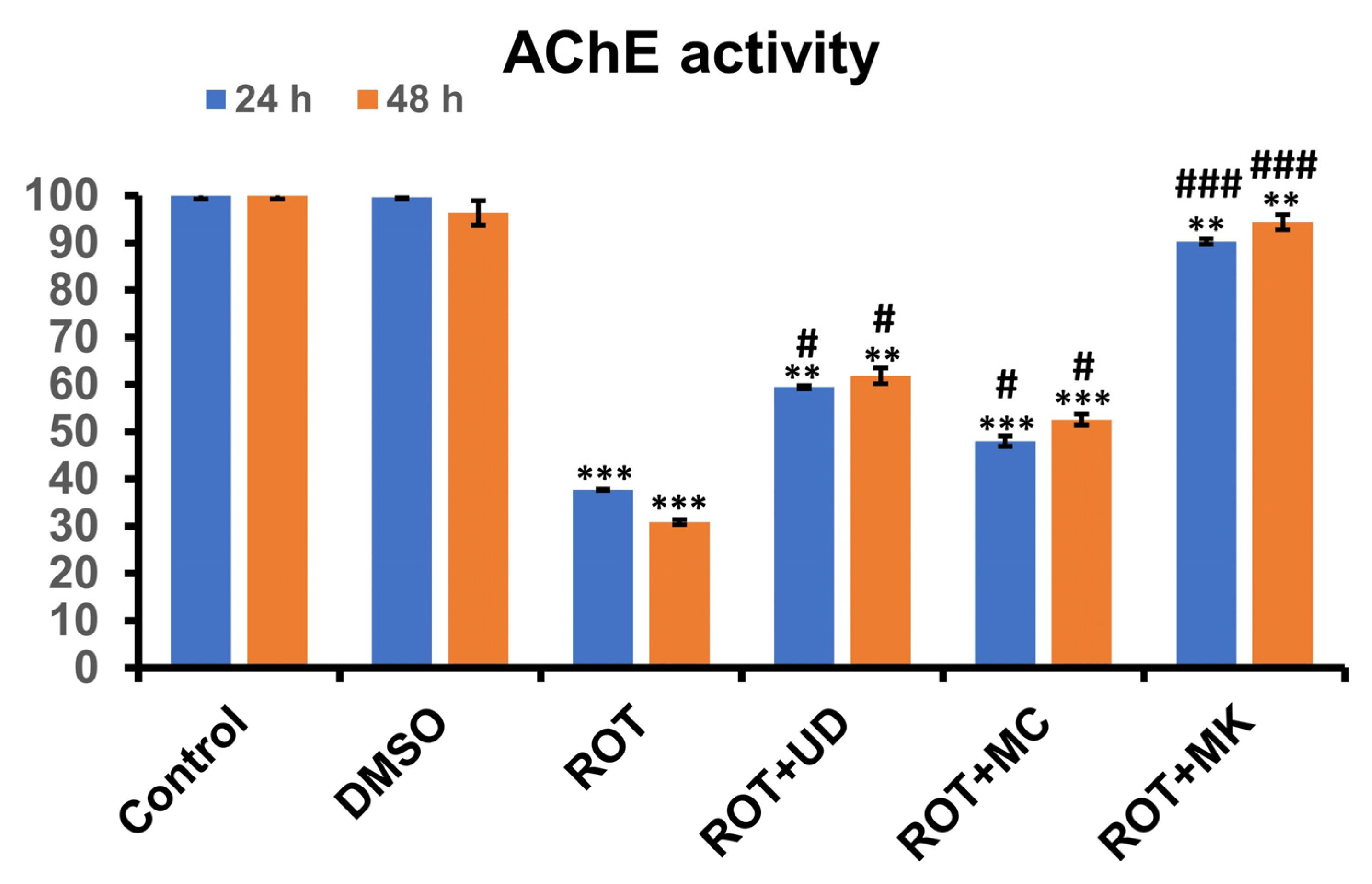
3.3.2. Rotenone Inhibits AChE Activity in D. melanogaster, and This Effect Is Reversed by Phytoextraction
3.4. Behavioral Assays
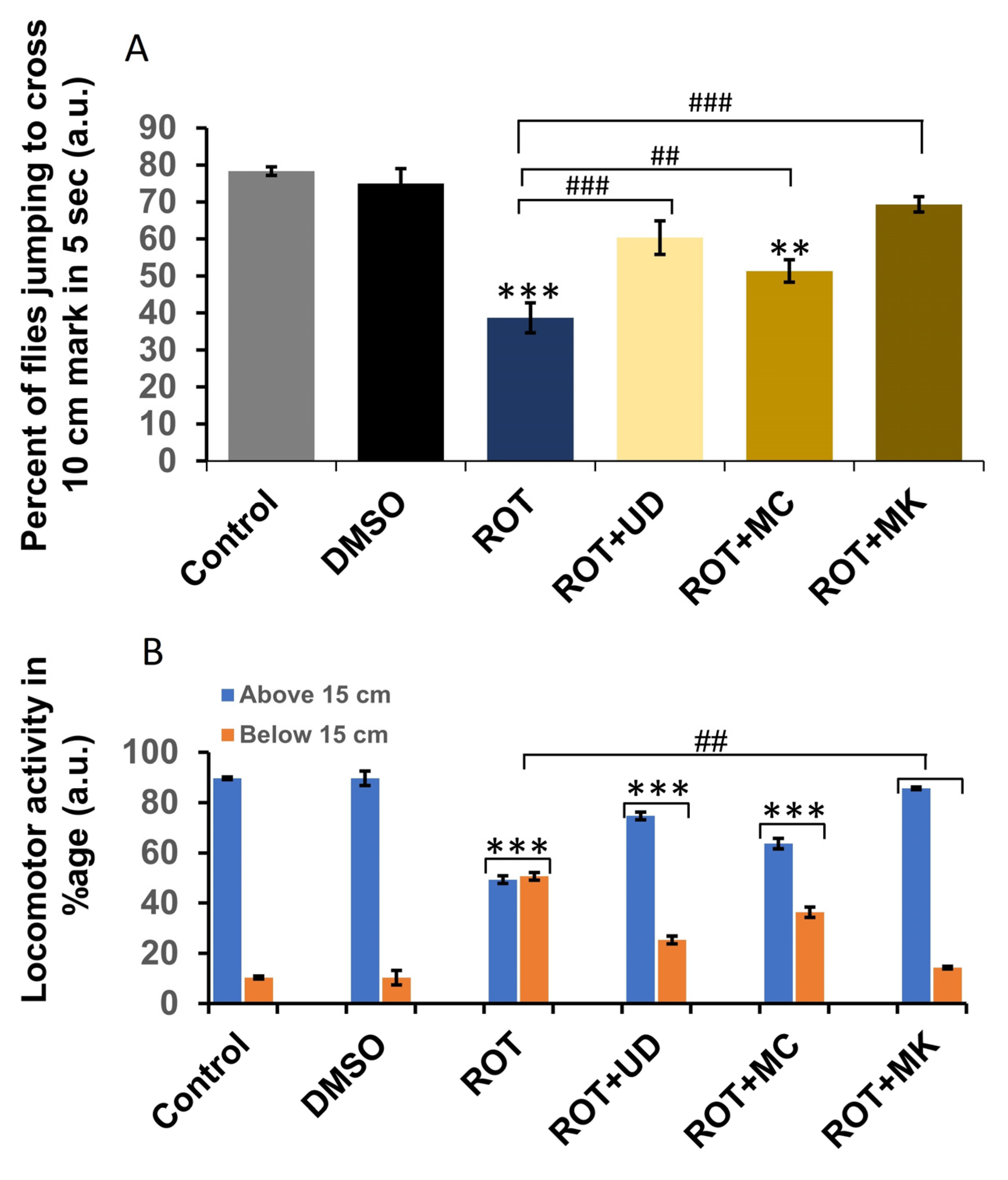
3.4.1. Rotenone Affects Locomotor Behavior in D. melanogaster
3.4.2. Significant Changes in the Jumping Activity of ROT-Exposed Flies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miliaraki, M.; Briassoulis, P.; Ilia, S.; Michalakakou, K.; Karakonstantakis, T.; Polonifi, A.; Bastaki, K.; Briassouli, E.; Vardas, K.; Pistiki, A.; et al. Oxidant/Antioxidant Status Is Impaired in Sepsis and Is Related to Anti-Apoptotic, Inflammatory, and Innate Immunity Alterations. Antioxidants 2022, 11, 231. [Google Scholar] [CrossRef]
- Sies, H.; Belousov, V.V.; Chandel, N.S.; Davies, M.J.; Jones, D.P.; Mann, G.E.; Murphy, M.P.; Yamamoto, M.; Winterbourn, C. Defining roles of specific reactive oxygen species (ROS) in cell biology and physiology. Nat. Rev. Mol. Cell Boil. 2022, 23, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, B.M.; Banik, B.K.; Borah, P.; Jain, A. Reactive Oxygen Species (ROS): Key Components in Cancer Therapies. Anticancer Agents Med. Chem. 2022, 22, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Abramov, A.Y. Redox biology in neurodegenerative disorders. Free Rad. Boil. Med. 2022, 188, 24–25. [Google Scholar] [CrossRef]
- Deepashree, S.; Shivanandappa, T.; Ramesh, S.R. Genetic repression of the antioxidant enzymes reduces the lifespan in Drosophila melanogaster. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2022, 192, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef] [PubMed]
- Beckers, M.; Bloem, B.R.; Verbeek, M.M. Mechanisms of peripheral levodopa resistance in Parkinson’s disease. NPJ Park. Dis. 2022, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Bandopadhyay, R.; Mishra, N.; Rana, R.; Kaur, G.; Ghoneim, M.M.; Alshehri, S.; Mustafa, G.; Ahmad, J.; Alhakamy, N.A.; Mishra, A. Molecular Mechanisms and Therapeutic Strategies for Levodopa-Induced Dyskinesia in Parkinson’s Disease: A Perspective Through Preclinical and Clinical Evidence. Front. Pharmacol. 2022, 13, 805388. [Google Scholar] [CrossRef] [PubMed]
- Gulcan, H.O. Selected Natural and Synthetic Agents Effective against Parkinson’s Disease with Diverse Mechanisms. Curr. Top. Med. Chem. 2022, 22, 199–208. [Google Scholar] [CrossRef]
- Tan, M.A.; Sharma, N.; An, S. Phyto-Carbazole Alkaloids from the Rutaceae Family as Potential Protective Agents against Neurodegenerative Diseases. Antioxidants 2022, 11, 493. [Google Scholar] [CrossRef]
- Nath, M.; Debnath, P. Therapeutic role of traditionally used Indian medicinal plants and spices in combating COVID-19 pandemic situation. J. Biomol. Str. Dynam. 2022, 40, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Anand, U.; Tudu, C.K.; Nandy, S.; Sunita, K.; Tripathi, V.; Loake, G.J.; Dey, A.; Proćków, J. Ethnodermatological use of medicinal plants in India: From ayurvedic formulations to clinical perspectives—A review. J. Ethnopharmacol. 2022, 284, 114744. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, K.K.; Magar, S.K.; Thapa, R.; Lamsal, A.; Bhandari, S.; Shrestha, J. Nutritional and pharmacological importance of stinging nettle (Urtica dioica L.): A review. Heliyon 2022, e09717. [Google Scholar] [CrossRef] [PubMed]
- Uyar, A.; Doğan, A.; Yaman, T.; Keleş, Ö.F.; Yener, Z.; Çelik, İ.; Alkan, E.E. The Protective Role of Urtica dioica Seed Extract Against Azoxymethane-Induced Colon Carcinogenesis in Rats. Nutr. Cancer 2022, 74, 306–319. [Google Scholar] [CrossRef]
- Chehri, A.; Yarani, R.; Yousefi, Z.; Novin Bahador, T.; Shakouri, S.K.; Ostadrahimi, A.; Mobasseri, M.; Pociot, F.; Araj-Khodaei, M. Anti-diabetic potential of Urtica Dioica: Current knowledge and future direction. J. Diabetes Metab. Disord. 2022, 21, 931–940. [Google Scholar] [CrossRef]
- Ahmadipour, B.; Khajali, F. Expression of antioxidant genes in broiler chickens fed nettle (Urtica dioica) and its link with pulmonary hypertension. Anim. Nutr. 2019, 5, 264–269. [Google Scholar] [CrossRef]
- Samakar, B.; Mehri, S.; Hosseinzadeh, H. A review of the effects of Urtica dioica (nettle) in metabolic syndrome. Iranian J. Basic Med. Sci. 2022, 25, 543. [Google Scholar] [CrossRef]
- Perestrelo, B.O.; Carvalho, P.M.; Souza, D.N.; Carneiro, M.J.; Cirino, J.; Carvalho, P.O.; Sawaya, A.; Oyama, L.M.; Nogueira, F.N. Antioxidant effect of chamomile tea on the salivary glands of streptozotocin-induced diabetic rats. Braz. Oral Res. 2022, 36, e034. [Google Scholar] [CrossRef]
- Shoara, R.; Hashempur, M.H.; Ashraf, A.; Salehi, A.; Dehshahri, S.; Habibagahi, Z. Efficacy and safety of topical Matricaria chamomilla L. (chamomile) oil for knee osteoarthritis: A randomized controlled clinical trial. Complement. Ther. Clin. Pr. 2022, 21, 181–187. [Google Scholar] [CrossRef]
- Mojibi, R.; Morad Jodaki, H.; Mehrzad, J.; Khosravi, A.R.; Sharifzadeh, A.; Nikaein, D. Apoptotic Effects of Caffeic Acid Phenethyl Ester and Matricaria chamomilla essential oil on A549 Non-small Cell Lung Cancer Cells. Iranian J. Vet. Med. 2022. [Google Scholar] [CrossRef]
- Mondal, P.; Natesh, J.; Penta, D.; Meeran, S.M. Extract of Murraya koenigii selectively causes genomic instability by altering redox-status by targeting PI3K/AKT/Nrf2/caspase-3 signaling pathway in human non-small cell lung cancer. Phytomedicine 2022, 104, 154272. [Google Scholar] [CrossRef] [PubMed]
- Aniqa, A.; Kaur, S.; Negi, A.; Sadwal, S.; Bharati, S. Phytomodulatory effects of Murraya koenigii in DMBA/TPA induced angiogenesis, hepatotoxicity and renal toxicity during skin carcinogenesis in mice. J. Adv. Sci. Res. 2022, 13, 153–165. [Google Scholar] [CrossRef]
- Sanmugarajah, V.; Rajkumar, G. A Review of Anti-hyperglycemic Effects of Curry Leaf Tree (Murraya koenigii). Borneo J. Pharm. 2022, 5, 104–114. [Google Scholar] [CrossRef]
- Markom, M.; Hasan, M.; Daud, W.R.W.; Singh, H.; Jahim, J.M. Extraction of hydrolyzable tannins from Phylla nthusniruri Linn: Effects of solvents and extraction methods. Sep. Purify. Technol. 2007, 52, 487–496. [Google Scholar] [CrossRef]
- Adams, C.; Thapa, S.; Kimura, E. Determination of a plant population density threshold for optimizing cotton lint yield: A synthesis. Field Crops Res. 2019, 230, 11–16. [Google Scholar] [CrossRef]
- Baliyan, S.; Mukherjee, R.; Priyadarshini, A.; Vibhuti, A.; Gupta, A.; Pandey, R.P.; Chang, C.M. Determination of Antioxidants by DPPH Radical Scavenging Activity and Quantitative Phytochemical Analysis of Ficus religiosa. Molecules 2022, 27, 1326. [Google Scholar] [CrossRef]
- Wołosiak, R.; Drużyńska, B.; Derewiaka, D.; Piecyk, M.; Majewska, E.; Ciecierska, M.; Worobiej, E.; Pakosz, P. Verification of the Conditions for Determination of Antioxidant Activity by ABTS and DPPH Assays-A Practical Approach. Molecules 2021, 27, 50. [Google Scholar] [CrossRef]
- Gudimella, K.K.; Gedda, G.; Kumar, P.S.; Babu, B.K.; Yamajala, B.; Rao, B.V.; Singh, P.P.; Kumar, D.; Sharma, A. Novel synthesis of fluorescent carbon dots from biobased Carica Papaya Leaves: Optical and structural properties with antioxidant and anti-inflammatory activities. Environ. Res. 2022, 204, 111854. [Google Scholar] [CrossRef]
- Clarke, G.; Ting, K.N.; Wiart, C.; Fry, J. High correlation of 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging, ferric reducing activity potential, and total phenolic content indicates redundancy in the use of all three assays to screen for antioxidant activity of extracts of plants from the Malaysian rainforest. Antioxidants 2013, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Do, Q.D.; Angkawijaya, A.E.; Tran-Nguyen, P.L.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.H. Effect of extraction solvent on total phenol content, total flavonoid content, and antioxidant activity of Limnophila aromatic. J. Food Drug Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Sreenivasulu, N.; Fernie, A.R. Diversity: Current and prospective secondary metabolites for nutrition and medicine. Curr. Opin. Biotechnol. 2022, 74, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Sofowora, A. Medicinal Plants and Traditional Medicine in Africa; Spectrum Books Ltd.: Ibadan, Nigeria, 1993; pp. 191–289. [Google Scholar] [CrossRef]
- Kumar, G.S.; Jayaveera, K.N.; Kumar, C.K.; Sanjay, U.P.; Swamy, B.M.; Kumar, D.V. Antimicrobial effects of Indian medicinal plants against acne-inducing bacteria. Trop. J. Pharm. Res. 2007, 6, 717–723. [Google Scholar] [CrossRef]
- Olugbenga, O.O.; Adebola, S.S.; Friday, A.D.; Mercy, A.T.; Keniokpo, O.S. Effect of dietary tomato powder on growth performance and blood characteristics of heat-stressed broiler chickens. Trop. Anim. Heal. Prod. 2022, 54, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mannu, A.; Poddighe, M.; Garroni, S.; Malfatti, L. Application of IR and UV-Vis spectroscopies and multivariate analysis for the classification of waste vegetable oils. Resour. Conserv. Recycl. 2022, 178, 106088. [Google Scholar] [CrossRef]
- Tian, W.; Chen, G.; Gui, Y.; Zhang, G.; Li, Y. Rapid quantification of total phenolics and ferulic acid in whole wheat using UV-Vis spectrophotometry. Food Control 2021, 123, 107691. [Google Scholar] [CrossRef]
- Ismail, M.M.; Morsy, G.M.; Mohamed, H.M.; El-Mansy, M.A.M.; Abd-Alrazk, M.M.A. FT-IR spectroscopic analyses of 4-hydroxy-1-methyl-3-2-nitro-2-oxoacetyl-2 (1H) quinolinone (HMNOQ). Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 113, 191195. [Google Scholar] [CrossRef] [PubMed]
- Pinelli, P.; Ieri, F.; Vignolini, P.; Bacci, L.; Baronti, S.; Romani, A. Extraction and HPLC analysis of phenolic compounds in leaves, stalks, and textile fibers of Urtica dioica L. J. Agric. Food Chem. 2008, 56, 9127–9132. [Google Scholar] [CrossRef]
- Miguel, F.G.; Cavalheiro, A.H.; Spinola, N.F.; Ribeiro, D.L.; Barcelos, G.R.M.; Antunes, L.M.G.; Hori, J.I.; Marquele-Oliveira, F.; Rocha, B.A.; Berretta, A.A. Validation of an RP-HPLC-DAD method for chamomile (Matricaria recutita) preparations and assessment of the marker, apigenin-7-glucoside, safety, and anti-inflammatory effect. Evid. Based Complement. Altern. Med. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pandit, S.; Kumar, M.; Ponnusankar, S.; Pal, B.C.; Mukherjee, P.K. RP-HPLC-DAD for simultaneous estimation of mahanine and mahanimbine in Murraya koenigii. Biomed. Chromatogr. 2011, 25, 959–962. [Google Scholar] [CrossRef]
- Singh, M.P.; Reddy, M.M.; Mathur, N.; Saxena, D.K.; Chowdhuri, D.K. Induction of hsp70, hsp60, hsp83 and hsp26 and oxidative stress markers in benzene, toluene and xylene exposed Drosophila melanogaster: Role of ROS generation. Toxicol. Appl. Pharmacol. 2009, 235, 226–243. [Google Scholar] [CrossRef]
- Hosamani, R.; Muralidhara. Neuroprotective efficacy of Bacopa monnieri against rotenone induced oxidative stress and neurotoxicity in Drosophila melanogaster. NeuroToxicology 2009, 30, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Akinade, T.C.; Babatunde, O.O.; Adedara, A.O.; Adeyemi, O.E.; Otenaike, T.A.; Ashaolu, O.P.; Johnson, T.O.; Terriente-Felix, A.; Whitworth, A.J.; Abolaji, A.O. Protective capacity of carotenoid trans-astaxanthin in rotenone-induced toxicity in Drosophila melanogaster. Sci. Rep. 2022, 12, 4594. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.P.; Bawani, S.S.; Anandhi, D.U.; Prashanth, K. Rotenone mediated developmental toxicity in Drosophila melanogaster. Environ. Toxicol. Pharmacol. 2022, 93, 103892. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Ram, K.R.; Mishra, M.; Shrivastava, M.; Saxena, D.K.; Chowdhuri, D.K. Effects of coexposure of benzene, toluene, and xylene to Drosophila melanogaster: Alteration in hsp70, hsp60, hsp83, hsp26, ROS generation and oxidative stress markers. Chemosphere 2010, 79, 577–587. [Google Scholar] [CrossRef]
- Singh, M.P.; Mishra, M.; Sharma, A.; Shukla, A.K.; Mudiam, M.K.; Patel, D.K.; Ram, K.R.; Chowdhuri, D.K. Genotoxicity and apoptosis in Drosophila melanogaster exposed to benzene, toluene and xylene: Attenuation by quercetin and curcumin. Toxicol. Appl. Pharmacol. 2011, 253, 14–30. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Feather-stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Nitta, Y.; Sugie, A. Studies of neurodegenerative diseases using Drosophila and the development of novel approaches for their analysis. Fly 2022, 16, 275–298. [Google Scholar] [CrossRef]
- Sharma, A.; Mishra, M.; Shukla, A.K.; Kumar, R.; Abdin, M.Z.; Chowdhuri, D.K. Organochlorine pesticide, endosulfan induced cellular and organismal response in Drosophila melanogaster. J. Hazard. Mater. 2012, 221–222, 275–287. [Google Scholar] [CrossRef]
- Chaitanya, M.V.N.L.; Ali, H.S.; Usamo, F.B. Regulatory considerations of herbal biomolecules. In Herbal Biomolecules in Healthcare Applications; Academic Press: Cambridge, MA, USA, 2022; pp. 669–676. [Google Scholar] [CrossRef]
- Mitra, E.; Ghosh, A.K.; Ghosh, D.; Mukherjee, D.; Chattopadhyay, A.; Dutta, S.; Bandyopadhyay, D. Protective effect of aqueous Curry leaf (Murraya koenigii) extracts against cadmium-induced oxidative stress in rat heart. Food Chem. Toxicol. 2012, 50, 1340–1353. [Google Scholar] [CrossRef]
- Biswas, A.K.; Chatli, M.K.; Sahoo, J. Antioxidant potential of curry (Murraya koenigii L.) and mint (Mentha spicata) leaf extracts and their effect on color and oxidative stability of raw ground pork meat during refrigeration storage. Food Chem. 2012, 133, 467–472. [Google Scholar] [CrossRef]
- Stalikas, C.D. Extraction, separation, and detection methods for phenolic acids and flavonoids. J. Sep. Sci. 2007, 30, 3268–3295. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, M.; Mewis, I.; Huyskens-Keil, S.; Jansen, M.A.K.; Zrenner, R.; Winkler, J.B.; Krumbein, A. UV-B-induced secondary plant metabolites-potential benefits for plant and human health. Crit. Rev. Plant Sci. 2012, 31, 229–240. [Google Scholar] [CrossRef]
- Jain, P.K.; Soni, A.; Jain, P.; Bhawsar, J. Phytochemical analysis of Mentha spicata plant extract using UV-VIS, FTIR, and GC/MS technique. J. Chem. Pharm. Res. 2016, 8, 16. [Google Scholar] [CrossRef]
- Wyrostek, J.; Kowalski, R.; Pankiewicz, U.; Solarska, E. Estimation of the content of selected active substances in primary and secondary herbal brews by UV-VIS and GC-MS spectroscopic analyses. J. anal. Met. Chem. 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Maobe, M.A.; Nyarango, R.M.; Box, P.O. Fourier transformer infrared spectrophotometer analysis of Urtica dioica medicinal herb used for the treatment of diabetes, malaria, and pneumonia in Kisii region, Southwest Kenya. World Appl. Sci. J. 2013, 21, 1128–1135. [Google Scholar] [CrossRef]
- Qais, F.A.; Shafiq, A.; Khan, H.M.; Husain, F.M.; Khan, R.A.; Alenazi, B.; Ahmad, I. Antibacterial effect of silver nanoparticles synthesized using Murraya koenigii (L.) against multidrug-resistant pathogens. Bioinorg. Chem. Appl. 2019, 2019, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Li, S.; Xiang, G.; Li, Q.; Fan, L.; He, L.; Gu, K. Determination of the acid values of edible oils via FTIR spectroscopy based on the OH stretching band. Food Chem. 2016, 212, 585–589. [Google Scholar] [CrossRef]
- Tsivelika, N.; Irakli, M.; Mavromatis, A.; Chatzopoulou, P.; Karioti, A. Phenolic Profile by HPLC-PDA-MS of Greek Chamomile Populations and Commercial Varieties and Their Antioxidant Activity. Foods 2021, 10, 2345. [Google Scholar] [CrossRef]
- Nagappan, T.; Ramasamy, P.; Wahid, M.E.A.; Segaran, T.C.; Vairappan, C.S. Biological activity of carbazole alkaloids and essential oil of Murraya koenigii against antibiotic-resistant microbes and cancer cell lines. Molecules 2011, 16, 9651–9664. [Google Scholar] [CrossRef] [Green Version]
- Ningappa, M.B.; Srinivas, L. Purification and characterization of∼ 35 kDa antioxidant protein from curry leaves (Murraya koenigii L.). Toxicol. Vitr. 2008, 22, 699–709. [Google Scholar] [CrossRef]
- Javed, H.; Meeran, M.; Azimullah, S.; Bader Eddin, L.; Dwivedi, V.D.; Jha, N.K.; Ojha, S. α-Bisabolol, a Dietary Bioactive Phytochemical Attenuates Dopaminergic Neurodegeneration through Modulation of Oxidative Stress, Neuroinflammation, and Apoptosis in Rotenone-Induced Rat Model of Parkinson’s disease. Biomolecules 2020, 10, 1421. [Google Scholar] [CrossRef] [PubMed]
- Pandareesh, M.D.; Shrivash, M.K.; Naveen Kumar, H.N.; Misra, K.; Srinivas Bharath, M.M. Curcumin Monoglucoside Shows Improved Bioavailability and Mitigates Rotenone Induced Neurotoxicity in Cell and Drosophila Models of Parkinson’s Disease. Neurochem. Res. 2016, 41, 3113–3128. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Siddique, H.R.; Mathur, N.; Mishra, R.K.; Mitra, K.; Saxena, D.K.; Chowdhuri, D.K. Adverse effect of organophosphate compounds, dichlorvos and chlorpyrifos in the reproductive tissues of transgenic Drosophila melanogaster: 70 kDa heat shock protein as a marker of cellular damage. Toxicology 2007, 238, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.V.; Yenisetti, S.C.; Rajini, P.S. Evidence of neuroprotective effects of saffron and crocin in a Drosophila model of parkinsonism. Neurotoxicology 2016, 52, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Krishna, G.; Muralidhara. Aqueous extract of tomato seeds attenuates rotenone-induced oxidative stress and neurotoxicity in Drosophila melanogaster. J. Sci. Food Agric. 2016, 96, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Riemensperger, T.; Issa, A.R.; Pech, U.; Coulom, H.; Nguyễn, M.V.; Cassar, M.; Jacquet, M.; Fiala, A.; Birman, S. A single dopamine pathway underlies progressive locomotor deficits in a Drosophila model of Parkinson disease. Cell Rep. 2013, 5, 952–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casanova, Y.; Negro, S.; Barcia, E. Application of neurotoxin-and pesticide-induced animal models of Parkinson’s disease in the evaluation of new drug delivery systems. Acta Pharm. 2022, 72, 35–58. [Google Scholar] [CrossRef]
- Zheng, L.; Yu, P.; Zhang, Y.; Wang, P.; Yan, W.; Guo, B.; Huang, C.; Jiang, Q. Evaluating the bio-application of biomacromolecule of lignin-carbohydrate complexes (LCC) from wheat straw in bone metabolism via ROS scavenging. Int. J. Biol. Macromol. 2021, 176, 13–25. [Google Scholar] [CrossRef]
- Dong, H.; Zheng, L.; Yu, P.; Jiang, Q.; Wu, Y.; Huang, C.; Yin, B. Characterization and application of lignin–carbohydrate complexes from lignocellulosic materials as antioxidants for scavenging in vitro and in vivo reactive oxygen species. ACS Sustain. Chem. Eng. 2019, 8, 256–266. [Google Scholar] [CrossRef]
- Gu, J.; Guo, M.; Zheng, L.; Yin, X.; Zhou, L.; Fan, D.; Shi, L.; Huang, C.; Ji, G. Protective Effects of Lignin-Carbohydrate Complexes from Wheat Stalk against Bisphenol a Neurotoxicity in Zebrafish via Oxidative Stress. Antioxidants 2021, 10, 1640. [Google Scholar] [CrossRef]
- Vasile, D.; Enescu, C.M.; Dincă, L. Which Are the Main Medicinal Plants That Could Be Harvested from Eastern Romania? Sci. Papers. Ser. Manag. Econ. Eng. Agric. Rural Dev. 2018, 18, 523–528. Available online: https://managementjournal.usamv.ro/index.php/scientific-papers/1623-which-are-the-main-medicinal-plants-that-could-be-harvested-from-eastern-romania-1623#spucontentCitation67 (accessed on 12 August 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Plant Species | Code | Plant Part Used | Solvents (%) Yield (w/w) | |
|---|---|---|---|---|---|
| Aqueous Extracts | Ethanolic Extracts | ||||
| 1. | U. dioica | UD | Leaves | 14.34 | 10.45 |
| 2. | M. chamomilla | MC | Flower | 8.56 | 14.68 |
| 3. | M. koenigii | MK | Leaves | 21.62 | 15.27 |
| Assays | Plant Species | Plant Part Used | EC50 (mg/mL) of Aqueous Extracts | EC50 (mg/mL) of Ethanolic Extracts |
|---|---|---|---|---|
| DPPH | UD | Leaves | 0.42 | 0.16 |
| MC | Flowers | 1.00 | 0.12 | |
| MK | Leaves | 0.33 | 0.10 | |
| ABTS | UD | Leaves | 0.55 | 0.10 |
| MC | Flowers | 1.04 | 0.16 | |
| MK | Leaves | 0.51 | 0.07 |
| Plant Species | Plant Part Used | TPC mg (GAE)/g (Aqueous) | TPC mg (GAE)/g (Ethanolic) | TFC mg (QE)/g (Aqueous) | TFC mg (QE)/g (Ethanolic) |
|---|---|---|---|---|---|
| U. dioica | Leaves | 26.08 ± 2.02 | 42.16 ± 2.06 | 5.458 ± 2.3 | 12.48 ± 1.04 |
| M. chamomilla | Flowers | 24.01 ± 1.50 | 40.5 ± 4.04 | 5.465 ± 1.06 | 12.64 ± 2.3 |
| M. koenigii | Leaves | 35.14 ± 3.0 | 48.93 ± 2.03 | 9.641 ± 2.5 | 22.88 ± 1.05 |
| S. No. | Phytoconstituents | Tests | Positive Results | UD | MC | MK |
|---|---|---|---|---|---|---|
| 1. | Phenols | Ferric chloride test | Bluish-green | + | + | + |
| 2. | Flavonoids | Alkaline reagent test | Orange-red | + | + | + |
| 3. | Alkaloids | Wagner’s test | Red precipitate | + | + | + |
| 4. | Tannins | FeCl3 test | Black blue | + | + | + |
| 5. | Carbohydrates | Molisch’s test | Red or dull violet | + | − | + |
| 6. | Saponins | Foam test | White precipitate | + | + | + |
| 7. | Terpenoids | Salkowski test | Change from pink to violet | + | + | + |
| 8. | Steroids | Liebermann’s test | violet to blue or green color | + | + | + |
| 9. | Glycosides | Keller-Killiani test | Brick red | − | + | + |
| S. No. | Frequency Range (cm−1) | Functional Groups | Phytocompounds Identified | ||
|---|---|---|---|---|---|
| UD | MC | MK | |||
| 1. | 3358.28 | 3327.40 | 3307.88 | H-bonded, OH stretching | Hydroxyl compounds |
| 2. | 2923.25 | 2921.90 | 2974.04 | Asymmetric stretching –CH(CH2) vibration | Saturated aliphatic Compounds (Lipids) |
| 3. | 2853.54 | 2802.86 | 2804.74 | Symmetric stretching –CH2(CH2) vibration | Proteins, lipids |
| 4. | 1698.70, 1622.77 | 1601.79 | 1611.44 | C=O stretching vibration | Ketone compound |
| 5. | 1455.85 | 1404.99 | - | C=C-C aromatic ring stretching | Aromatic compound |
| 6. | 1399.89 | - | 1396.51 | O-H, alcoholic group | Phenol or tertiary alcohol |
| 7. | - | 1271.82 | 1275.04 | CN stretching | Aromatic primary amine |
| 8. | 1160.88 | 1174.09 | 1140.63 | Polymeric OH, C-O stretching | Cyclic ether |
| 9. | 1055.84 | 1074.42, 1043.61 | 1044.62 | Phosphate ion | Phosphate compound |
| 10. | - | - | 877.47 | p-O-C stretching | Aromatic phosphate |
| 11. | 720.06 | 620.82 | 659.68 | C-Cl stretching | Aliphatic chloro compound |
| S. No | Retention Time (Rt min.) | Compound | Molecular Formulae | Chemical Structure | Molecular Weight (g/mol) | Pharmacological Actions | ||
|---|---|---|---|---|---|---|---|---|
| UD | MC | MK | ||||||
| 1. | 1.931 | - | 1.645 | Fumaric acid | C4H4O4 |  | 116.07 | Reduces gallstone formation, used for the treatment of multiple sclerosis and psoriasis. |
| 2. | 4.240 | 4.235 | 4.276 | Gallic acid | C7H6O5 |  | 170.12 | Expectorant, cytotoxic steroid, memory enhancer, anti-inflammatory, anti-neoplastic, and antioxidant properties. |
| 3. | - | 7.523 | 7.473 | Protocatechuic acid | C7H6O4 |  | 154.12 | Neuroprotective, antioxidant, anticancer, antibacterial, anti-aging, and anti-asthma properties. |
| 4. | 9.211 | - | 9.865 | Catechins | C15H14O6 |  | 290.27 | Used to prevent and treat various diseases, high antioxidant activity, and used in cosmetics. |
| 5. | - | 11.387 | - | 4-O-Caffeoylquinic acid | C16H18O9 |  | 354.31 | Cytoprotective, neuroprotective, and hepatoprotective effects. |
| 6. | 15.982 | 14.683 | 15.554 | Caffeic acid derivative | C9H8O4 |  | 180.16 | Prevents DNA damage and oxidative stress induced by free radicals. |
| 7. | 16.683 | 16.299 | 16.693 | Epicatechin | C15H14O6 |  | 290.27 | Reduces blood glucose levels in diabetic patients and stimulates mitochondrial respiration. |
| 8. | 17.248 | 17.558 | 17.280 | Rutin | C27H30O16 |  | 610.5 | Hypolipidemic, anti-protozoal, vasoactive, cytoprotective, anti-allergic, anti-platelet, anti-hypertensive, and anti-spasmodic properties. |
| 9. | 17.515 | 17.428 | 17.625 | Syringic acid | C9H10O5 |  | 198.17 | Used in the prevention of CVDs, cancer, diabetes, and possesses antioxidant activities. |
| 10. | 18.094 | 18.763 | 18.651 | Isorhamnetin-3-O-glucoside | C22H22O12 |  | 478.4 | Anti-viral, antioxidant, anticancer, anti-tumor, anti-inflammatory, and antimicrobial properties. |
| 11. | - | 19.243 | - | Apigenin-7-O-glucoside | C21H20O10 |  | 432.4 | Prominent chemopreventive, anti-candidal effect, antifungal potential, and strengthens the failing heart. |
| 12. | 20.728 | 20.152 | 20.835 | p-coumaric acid | C9H8O3 |  | 164.16 | Anti-inflammatory, antimicrobial, anti-viral, and antibacterial properties. |
| 13. | - | 21.739 | - | 4,5-O-dicaffeoylquinic acid | C25H24O12 |  | 516.4 | In melanocytes, significantly reduces tyrosinase activity and melanin synthesis in a dose-dependent manner. |
| 14. | 22.564 | 22.571 | 22.677 | 2-O-Caffeoylmalic acid | C13H12O8 |  | 296.230 | Prevents ROS production and possesses high antioxidant activity. |
| 15. | 23.232 | 23.924 | 23.232 | Ferulic acid | C10H10O4 |  | 194.18 | Wide range of therapeutic uses against various diseases including cancer, arthritis, etc. |
| 16. | - | 24.579 | - | Naringin | C27H32O14 |  | 580.5 | Anti-carcinogenic and acts as inhibitor of selected cytochrome P450 enzymes. |
| 17. | 25.247 | 25.460 | 25.814 | Quercetin (quercetin-3-O-rhamonoside) | C21H20O11 |  | 448.4 | Used in the treatment of inflammatory, allergic, and metabolic disorders and act as anti-protozoal. |
| 18. | 27.723 | 27.787 | 27.379 | Myricetin | C15H10O8 |  | 318.23 | Acts as an anti-epileptic, anti-amyloidogenic, anti-diabetic, antioxidant, antibacterial, anti-ulcer, antiviral, anticancer, and anti-inflammatory agent. |
| 19. | 28.231 | 28.789 | 29.031 | Quercetin | C21H20O11 |  | 448.4 | Decreases tumor necrosis factor (TNF-α) production in macrophages and LPS-driven IL-8 synthesis in lung A549 cells generated by lipopolysaccharide (LPS). |
| 20. | 32.156 | 32.956 | 31.456 | Kaempferol | C15H10O6 |  | 286.24 | Anxiolytic, anti-diabetic, anti-estrogenic, anti-osteoporotic, cardioprotective, and neuroprotective properties. |
| 21. | - | 34.320 | - | Luteolin | C15H10O6 |  | 286.24 | Exhibits anti-inflammatory properties due to ability to regulate transcription factors like NF-B, AP-1, and STAT3. |
| 22. | - | 34.745 | - | Cirsiliol | C17H14O7 |  | 330.29 | Act as inhibitor of arachidonate 5-lipoxygenase and has anticancer, hypnotic, sedative, and anti-inflammatory properties. |
| 23. | 35.339 | 35.445 | 36.021 | Isorhamnetin | C16H12O7 |  | 316.26 | Cerebrovascular and cardiovascular protective properties; in addition, has antioxidant, anti-tumor, anti-inflammatory, organ protection, and obesity prevention properties. |
| 24. | 37.277 | - | 37.552 | Kaempferol 3-O-glucoside | C21H14O11 |  | 448.38 | Lowers the risk of chronic diseases, particularly cancer, and boosts the body’s antioxidant defenses against free radicals. |
| 25. | 39.901 | 41.026 | 40.516 | Isorhamnetin 3-O-rutinoside | C28H32O16 |  | 624.5 | Inhibits membrane proteins and has anti-apoptosis, antioxidation, anti-tumor, anti-inflammation, antiviral, antibacterial, anti-amyloidogenic, and anti-diabetic properties. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shabir, S.; Yousuf, S.; Singh, S.K.; Vamanu, E.; Singh, M.P. Ethnopharmacological Effects of Urtica dioica, Matricaria chamomilla, and Murraya koenigii on Rotenone-Exposed D. melanogaster: An Attenuation of Cellular, Biochemical, and Organismal Markers. Antioxidants 2022, 11, 1623. https://doi.org/10.3390/antiox11081623
Shabir S, Yousuf S, Singh SK, Vamanu E, Singh MP. Ethnopharmacological Effects of Urtica dioica, Matricaria chamomilla, and Murraya koenigii on Rotenone-Exposed D. melanogaster: An Attenuation of Cellular, Biochemical, and Organismal Markers. Antioxidants. 2022; 11(8):1623. https://doi.org/10.3390/antiox11081623
Chicago/Turabian StyleShabir, Shabnam, Sumaira Yousuf, Sandeep Kumar Singh, Emanuel Vamanu, and Mahendra P. Singh. 2022. "Ethnopharmacological Effects of Urtica dioica, Matricaria chamomilla, and Murraya koenigii on Rotenone-Exposed D. melanogaster: An Attenuation of Cellular, Biochemical, and Organismal Markers" Antioxidants 11, no. 8: 1623. https://doi.org/10.3390/antiox11081623