
Differential Radiomodulating Action of Olea europaea L. cv. Caiazzana Leaf Extract on Human Normal and Cancer Cells: A Joint Chemical and Radiobiological Approach

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
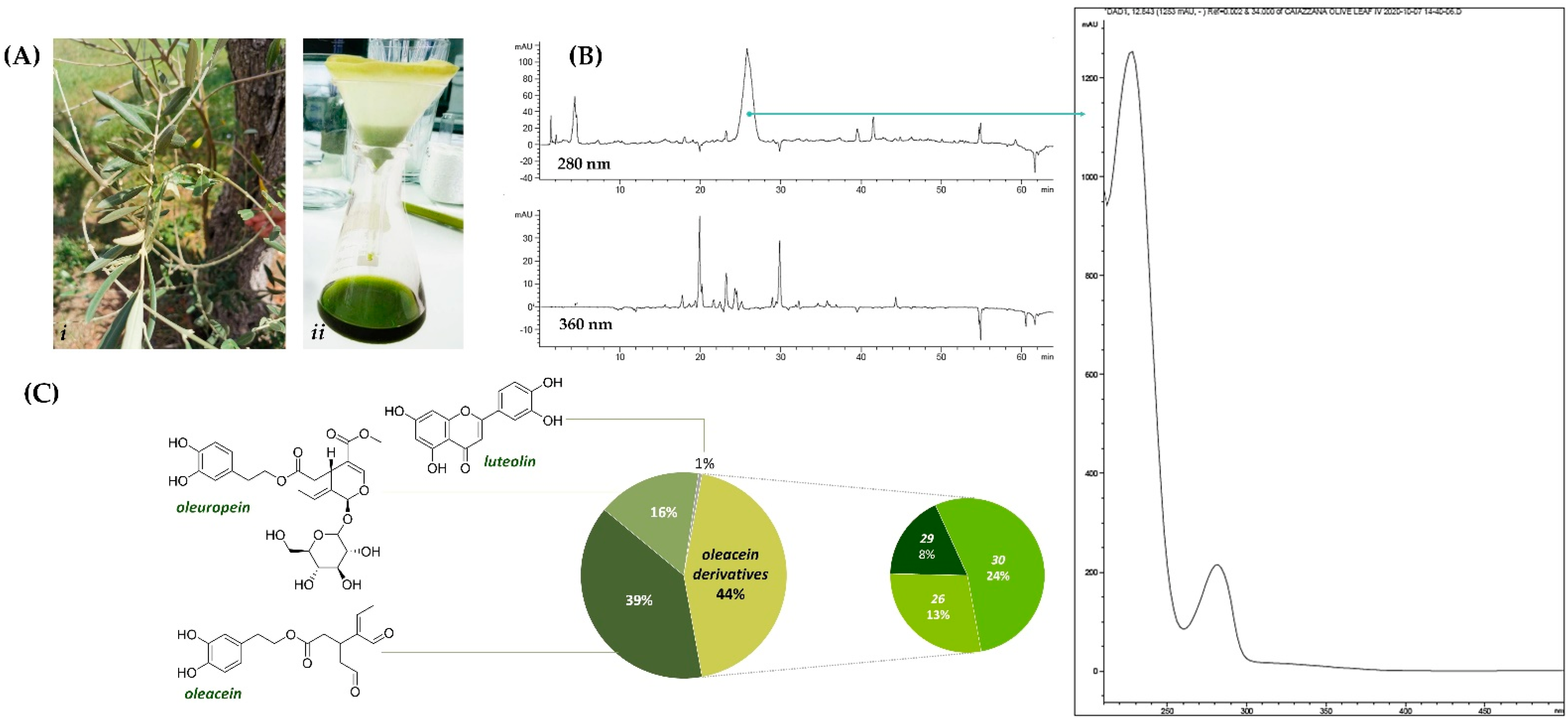
2.1. Olive Leaf Collection and Extraction
2.2. Chemical Characterization: UV-Vis, HPLC-UV-DAD and UHPLC-ESI-TOF/MS
2.3. Radical Scavenging Capacity: DPPH and ABTS Tests
2.4. Fe (III) Reducing Power
2.5. Cell Cultures: Maintenance and Preparation
2.6. Cell Irradiation
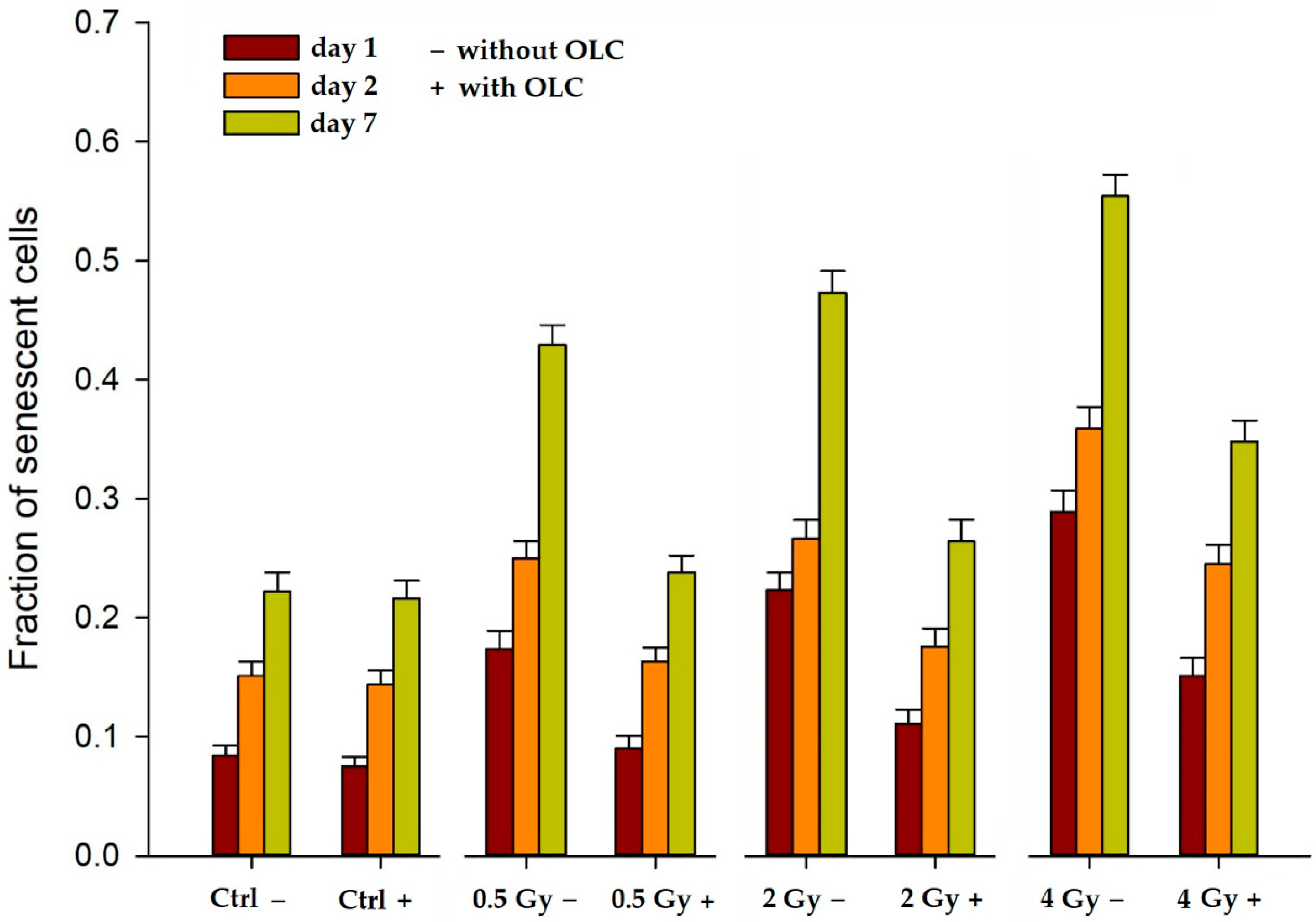
2.7. β-Galactosidase Assay for Quantification of Cellular Senescence in HUVECs
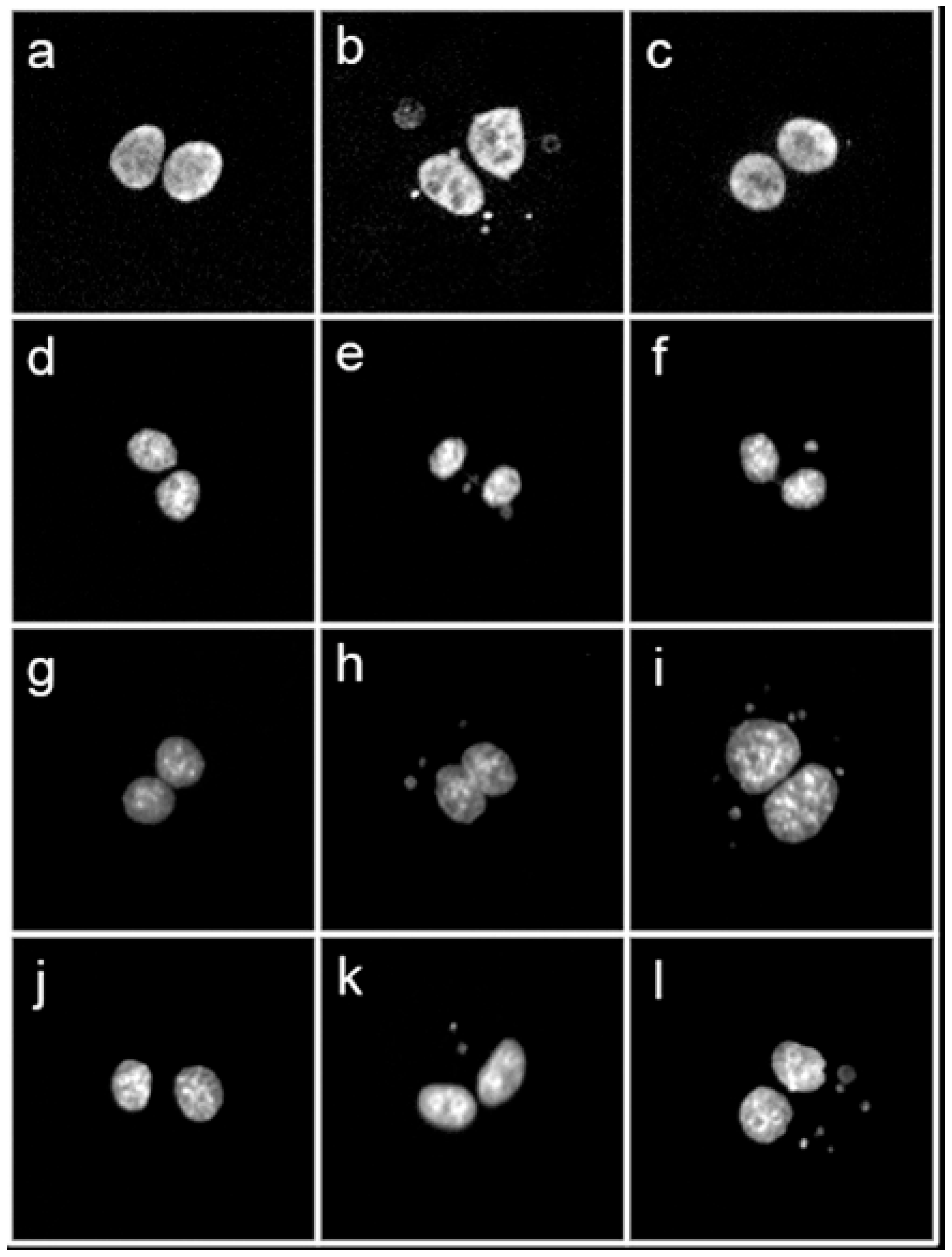
2.8. Determination of Radiation-Induced DNA Damage
3. Results
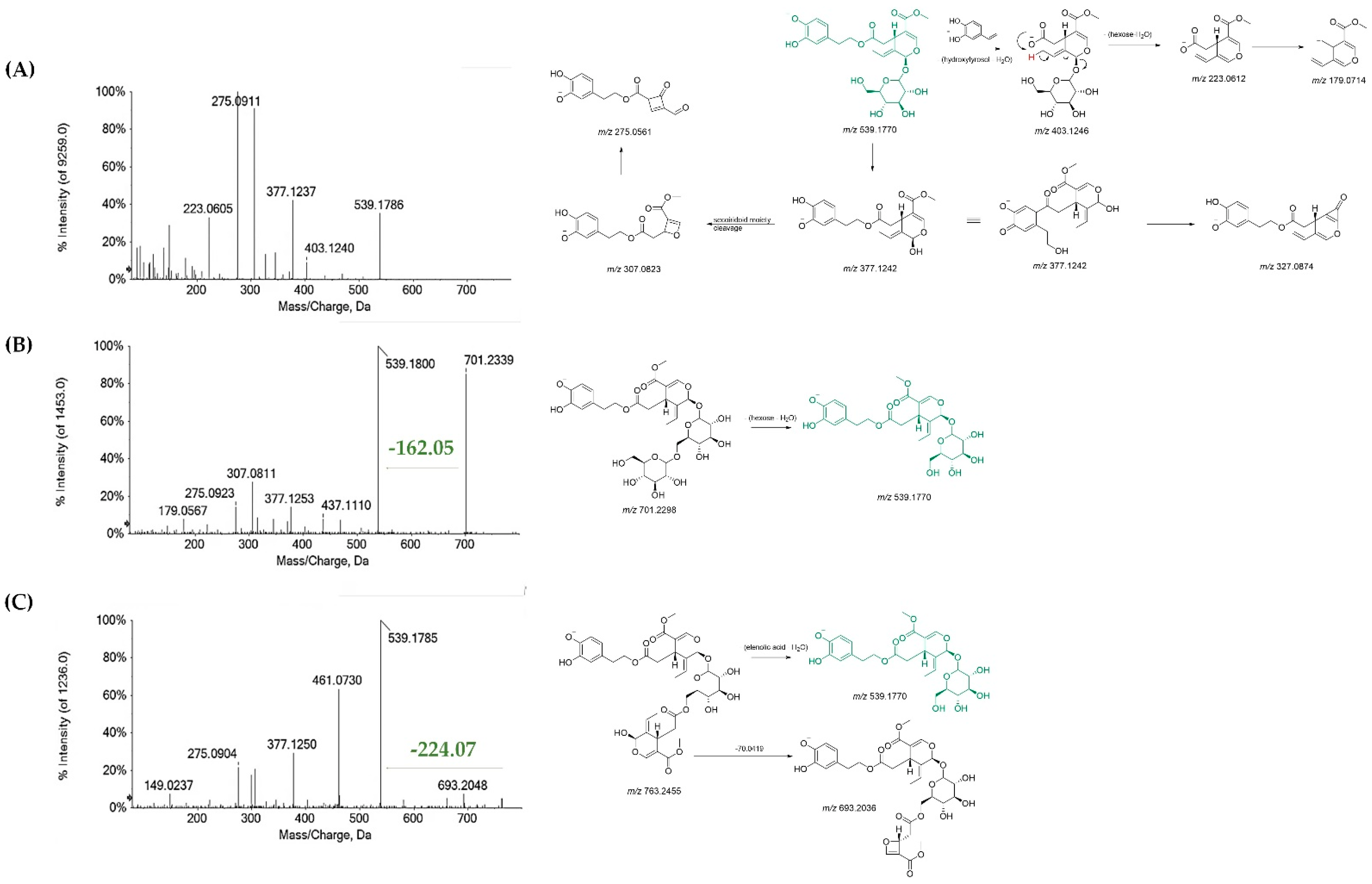
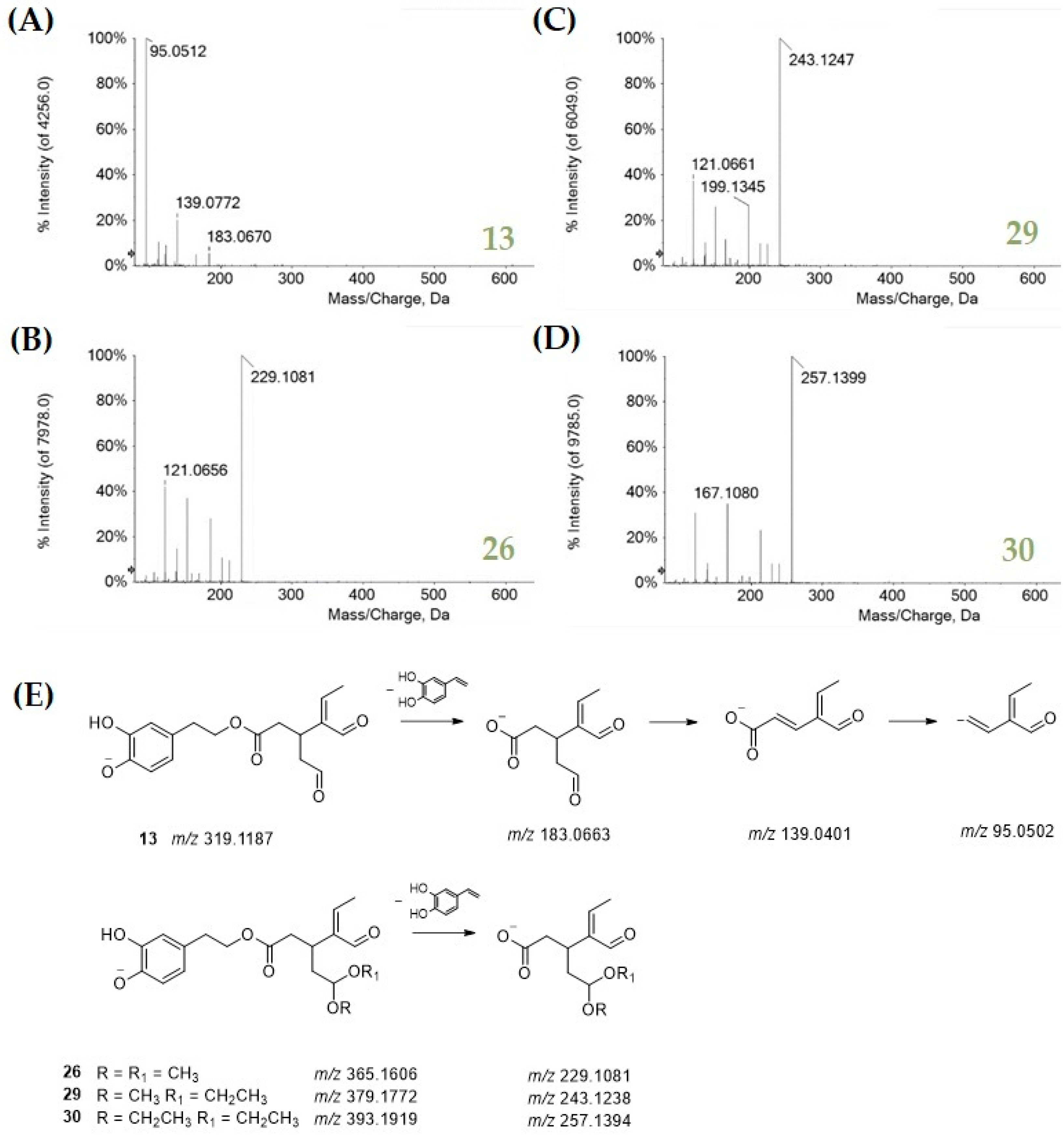
3.1. Chemical Profiling of Olive Tree Leaf Polyphenols
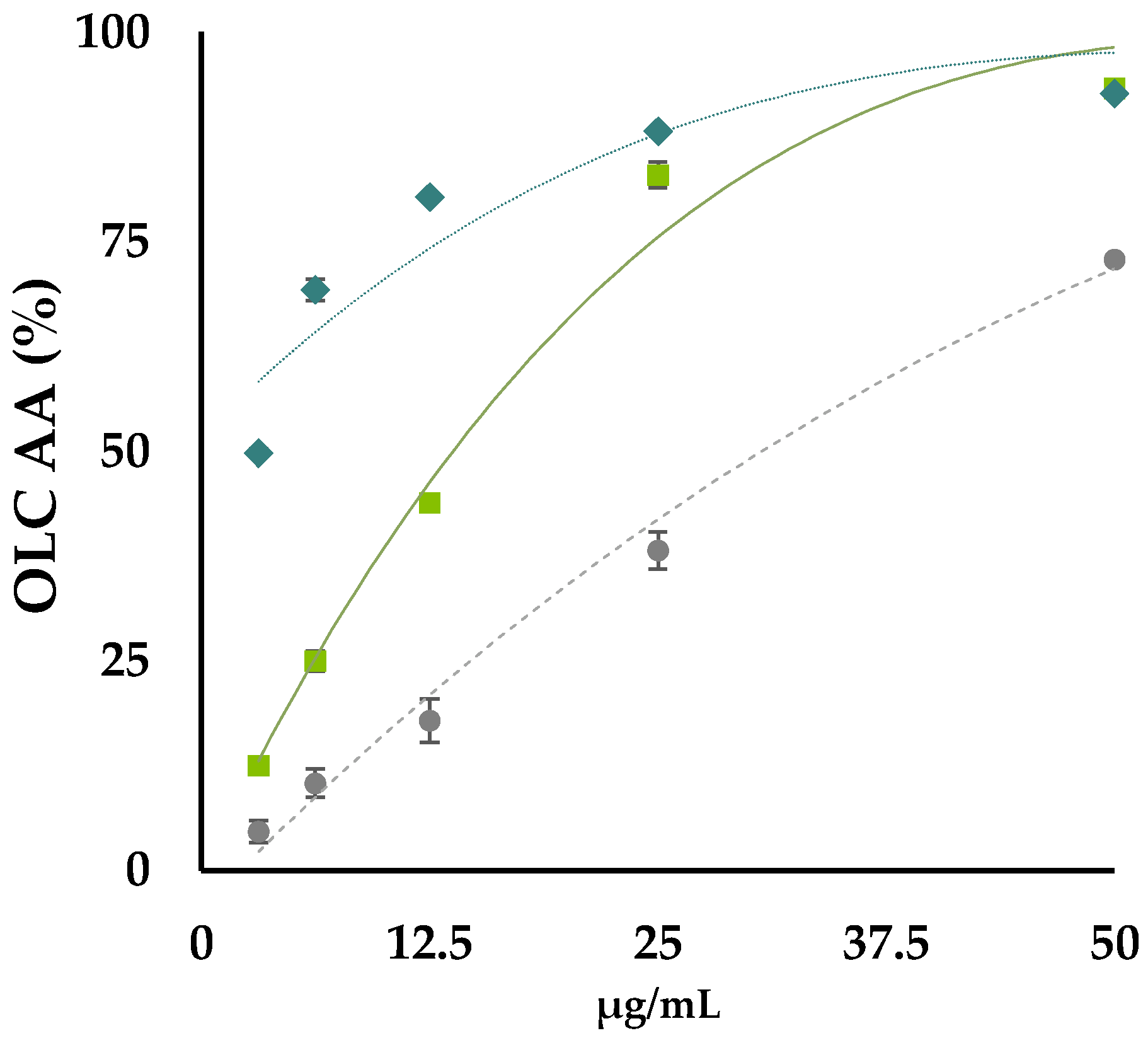
3.2. OLC Antiradical and Reducing Activities
3.3. Selective OLC-Mediated Radiomodulating Effects in Normal and Cancer Cells
3.3.1. OLC Decreases Radiation-Induced Premature Senescence (PS) in HUVECs
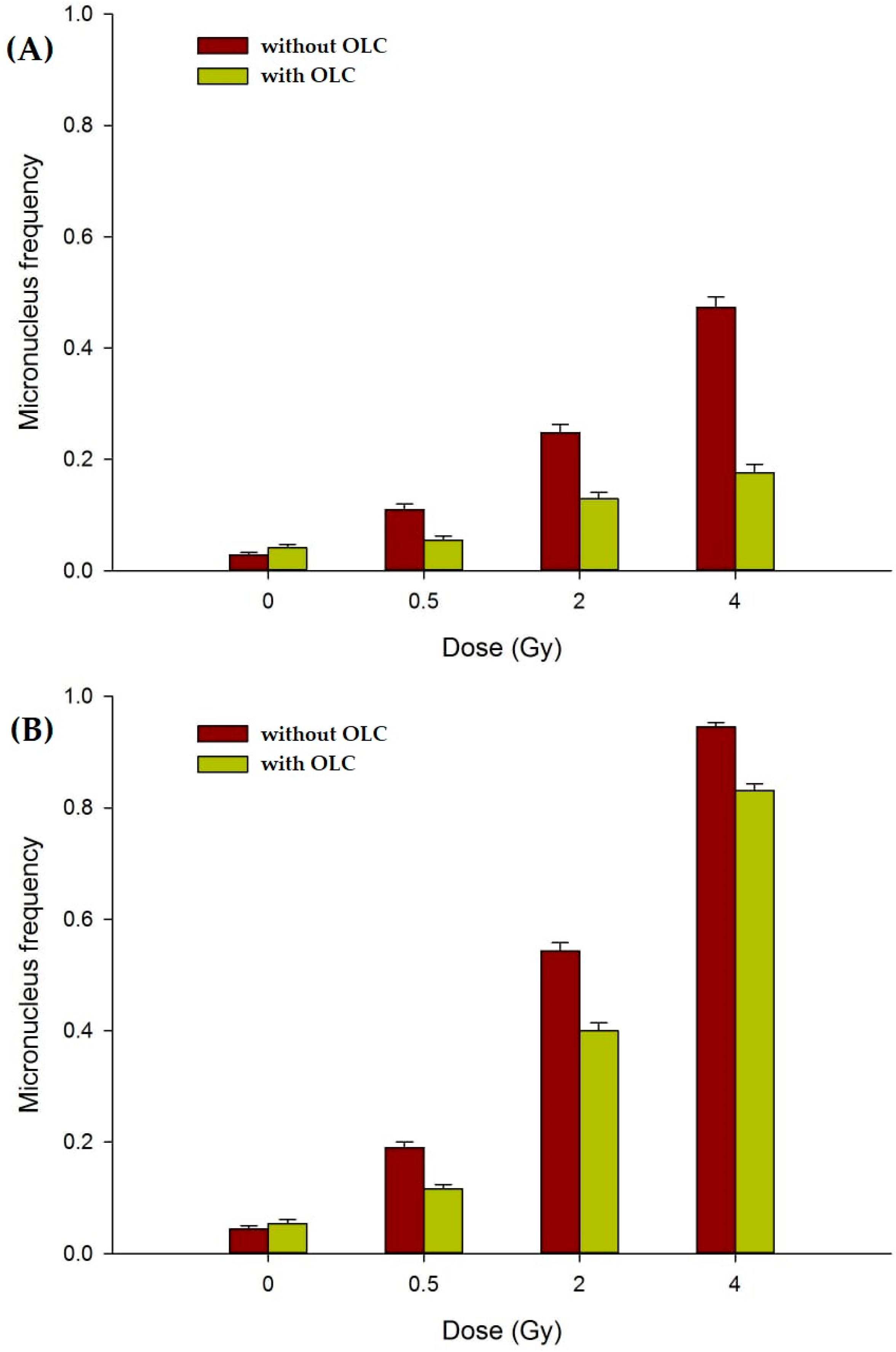
3.3.2. OLC Mitigates Radiation-Induced DNA Damage in Normal Cell Lines
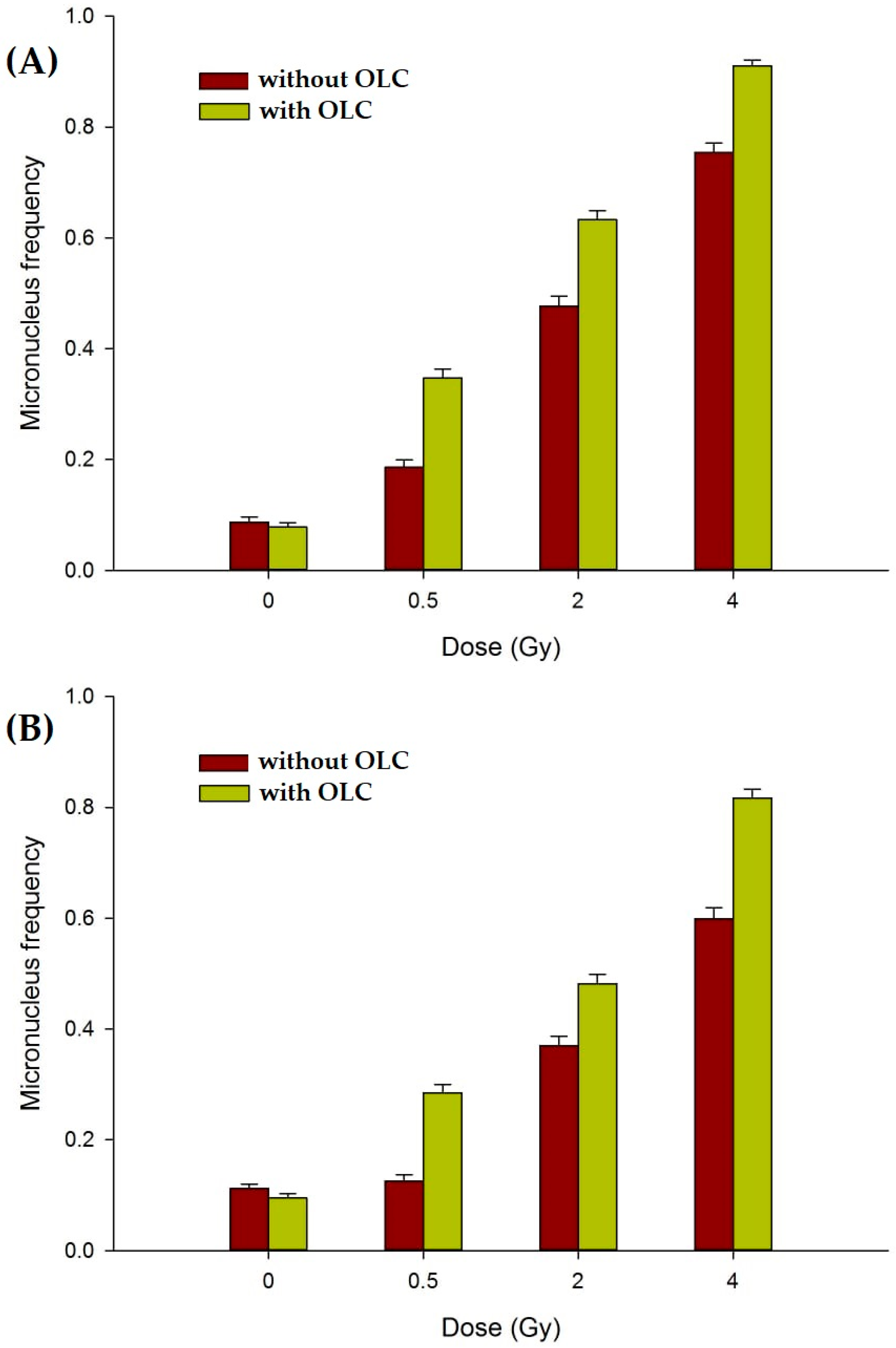
3.3.3. The OLC Presence Exacerbates Radiation-Induced DNA Damage in Cancer Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allali, S.; Kirova, Y. Radiodermatitis and Fibrosis in the Context of Breast Radiation Therapy: A Critical Review. Cancers 2021, 13, 5928. [Google Scholar] [CrossRef] [PubMed]
- van der Laan, H.P.; Van den Bosch, L.; Schuit, E.; Steenbakkers, R.J.H.M.; van der Schaaf, A.; Langendijk, J.A. Impact of radiation-induced toxicities on quality of life of patients treated for head and neck cancer. Radiother. Oncol. 2021, 160, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Suit, H.; Goldberg, S.; Niemierko, A.; Ancukiewicz, M.; Hall, E.; Goitein, M.; Wong, W.; Paganetti, H. Secondary Carcinogenesis in Patients Treated with Radiation: A Review of Data on Radiation-Induced Cancers in Human, Non-human Primate, Canine and Rodent Subjects. Radiat. Res. 2007, 167, 12–42. [Google Scholar] [CrossRef] [PubMed]
- Kamran, S.C.; Berrington de Gonzalez, A.; Ng, A.; Haas-Kogan, D.; Viswanathan, A.N. Therapeutic radiation and the potential risk of second malignancies. Cancer 2016, 122, 1809–1821. [Google Scholar] [CrossRef]
- Barnett, G.C.; West, C.M.; Dunning, A.M.; Elliott, R.M.; Coles, C.E.; Pharoah, P.D.; Burnet, N.G. Normal tissue reactions to radiotherapy: Towards tailoring treatment dose by genotype. Nat. Rev. Cancer. 2009, 9, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Shimura, T.; Kakuda, S.; Ochiai, Y.; Nakagawa, H.; Kuwahara, Y.; Takai, Y.; Kobayashi, J.; Komatsu, K.; Fukumoto, M. Acquired radioresistance of human tumor cells by DNA-PK/AKT/GSK3beta-mediated cyclin D1 overexpression. Oncogene 2010, 29, 4826–4837. [Google Scholar] [CrossRef]
- McCann, E.; O’Sullivan, J.; Marcone, S. Targeting cancer-cell mitochondria and metabolism to improve radiotherapy response. Transl. Oncol. 2021, 14, 100905. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, H.; Van De Gucht, M.; De Ridder, M. Hypoxic Radioresistance: Can ROS Be the Key to Overcome It? Cancers 2019, 11, 112. [Google Scholar] [CrossRef]
- Arnold, C.R.; Mangesius, J.; Skvortsova, I.-I.; Ganswindt, U. The Role of Cancer Stem Cells in Radiation Resistance. Front. Oncol. 2020, 10, 164. [Google Scholar] [CrossRef]
- De Ruysscher, D.; Niedermann, G.; Burnet, N.G.; Siva, S.; Lee, A.W.M.; Hegi-Johnson, F. Radiotherapy toxicity. Nat. Rev. Dis. Primers 2019, 5, 13. [Google Scholar] [CrossRef]
- Mun, G.I.; Kim, S.; Choi, E.; Kim, C.S.; Lee, Y.S. Pharmacology of natural radioprotectors. Arch. Pharm. Res. 2018, 41, 1033–1050. [Google Scholar] [CrossRef] [PubMed]
- Faramarzi, S.; Piccolella, S.; Manti, L.; Pacifico, S. Could polyphenols really be a good radioprotective strategy? Molecules 2021, 26, 4969. [Google Scholar] [CrossRef] [PubMed]
- Zdrowowicz, M.; Spisz, P.; Hać, A.; Herman-Antosiewicz, A.; Rak, J. Influence of Hypoxia on Radiosensitization of Cancer Cells by 5-Bromo-20-deoxyuridine. Int. J. Mol. Sci. 2022, 23, 1429. [Google Scholar] [CrossRef] [PubMed]
- Hazra, B.; Ghosh, S.; Kumar, A.; Pandey, B.N. The prospective role of plant products in radiotherapy of cancer: A current overview. Front. Pharmacol. 2012, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Sultana, M.; Qazi, A.; Qazi, M.H.; Parveen, G.; Waquar, S.; Ashraf, A.B.; Rasool, M. Role of Natural Radiosensitizers and Cancer Cell Radioresistance: An Update. Anal. Cell. Pathol. 2016, 2016, 6146595. [Google Scholar] [CrossRef] [PubMed]
- Akter, R.; Najda, A.; Rahman, M.H.; Shah, M.; Wesołowska, S.; Hassan, S.S.u.; Mubin, S.; Bibi, P.; Saeeda, S. Potential Role of Natural Products to Combat Radiotherapy and Their Future Perspectives. Molecules 2021, 26, 5997. [Google Scholar] [CrossRef]
- Piccolella, S.; Crescente, P.; Nocera, P.; Pacifico, F.; Manti, L.; Pacifico, S. Ultrasound-assisted aqueous extraction, LC-MS/MS analysis and radiomodulating capability of autochthonous Italian sweet cherry fruits. Food Funct. 2018, 9, 1840–1849. [Google Scholar] [CrossRef]
- Calvaruso, M.; Pucci, G.; Musso, R.; Bravatà, V.; Cammarata, F.P.; Russo, G.; Forte, G.I.; Minafra, L. Nutraceutical Compounds as Sensitizers for Cancer Treatment in Radiation Therapy. Int. J. Mol. Sci. 2019, 20, 5267. [Google Scholar] [CrossRef]
- Wang, J.; Yu, M.; Xiao, L.; Xu, S.; Yi, Q.; Jin, W. Radiosensitizing effect of oleanolic acid on tumor cells through the inhibition of GSH synthesis in vitro. Oncol. Rep. 2013, 30, 917–924. [Google Scholar] [CrossRef]
- Sebastià, N.; Montoro, A.; Hervás, D.; Pantelias, G.; Hatzi, V.I.; Soriano, J.M.; Villaescusa, J.I.; Terzoudi, G.I. Curcumin and trans-resveratrol exert cell cycle-dependent radioprotective or radiosensitizing effects as elucidated by the PCC and G2-assay. Mutat. Res. 2014, 766–767, 49–55. [Google Scholar] [CrossRef]
- Kefayat, A.; Ghahremani, F.; Safavi, A.; Hajiaghababa, A.; Moshtaghian, J. C-phycocyanin: A natural product with radiosensitizing property for enhancement of colon cancer radiation therapy efficacy through inhibition of COX-2 expression. Sci. Rep. 2019, 9, 19161. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, V.; Portaccio, M.; Piccolella, S.; Manti, L.; Pacifico, S.; Lepore, M. Study of SH-SY5Y Cancer Cell Response to Treatment with Polyphenol Extracts Using FT-IR Spectroscopy. Biosensors 2017, 7, 57. [Google Scholar] [CrossRef]
- Castillo, J.J.; Alcaraz, M.; Benavente-garcía, O. Antioxidant and radioprotective effects of olive leaf extract. In Olives and Olive Oil in Health and Disease Prevention, 1st ed.; Preedy, V., Watson, R., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2010; Chapter 102; pp. 951–958. [Google Scholar]
- Xu, T.; Xiao, D. Oleuropein enhances radiation sensitivity of nasopharyngeal carcinoma by downregulating PDRG1 through HIF1α-repressed microRNA-519d. J. Exp. Clin. Cancer Res. 2017, 36, 3. [Google Scholar] [CrossRef] [PubMed]
- Scognamiglio, M.; D’Abrosca, B.; Pacifico, S.; Fiumano, V.; De Luca, P.; Monaco, P.; Fiorentino, A. Polyphenol characterization and antioxidant evaluation of Olea europaea varieties cultivated in Cilento National Park (Italy). Food Res. Int. 2012, 46, 294–303. [Google Scholar] [CrossRef]
- Il Germoplasma Dell’olivo in Campania. Available online: http://agricoltura.regione.campania.it/pubblicazioni/pdf/germoplasma_olivo.pdf (accessed on 2 November 2021).
- Formato, M.; Piccolella, S.; Zidorn, C.; Vastolo, A.; Calabrò, S.; Cutrignelli, M.I.; Pacifico, S. UHPLC-ESI-QqTOF Analysis and In Vitro Rumen Fermentation for Exploiting Fagus sylvatica Leaf in Ruminant Diet. Molecules 2022, 27, 2217. [Google Scholar] [CrossRef]
- Longo, E.; Morozova, K.; Scampicchio, M. Effect of light irradiation on the antioxidant stability of oleuropein. Food Chem. 2017, 237, 91–97. [Google Scholar] [CrossRef]
- Piccolella, S.; Crescente, G.; Volpe, M.G.; Paolucci, M.; Pacifico, S. UHPLC-HR-MS/MS-guided recovery of bioactive flavonol compounds from Greco di Tufo vine leaves. Molecules 2019, 24, 3630. [Google Scholar] [CrossRef]
- Dichio, B.; Margiotta, G.; Xiloyannis, C.; Bufo, S.A.; Sofo, A.; Cataldi, T.R.I. Changes in water status and osmolyte contents in leaves and roots of olive plants (Olea europaea L.) subjected to water deficit. Trees 2009, 23, 247–256. [Google Scholar] [CrossRef]
- Asteggiano, A.; Franceschi, P.; Zorzi, M.; Aigotti, R.; Dal Bello, F.; Baldassarre, F.; Lops, F.; Carlucci, A.; Medana, C.; Ciccarella, G. HPLC-HRMS Global Metabolomics Approach for the Diagnosis of “Olive Quick Decline Syndrome” Markers in Olive Trees Leaves. Metabolites 2021, 11, 40. [Google Scholar] [CrossRef]
- Quirantes-Piné, R.; Lozano-Sánchez, J.; Herrero, M.; Ibáñez, E.; Segura-Carretero, A.; Fernández-Gutiérrez, A. HPLC–ESI–QTOF–MS as a Powerful Analytical Tool for Characterising Phenolic Compounds in Olive-leaf Extracts. Phytochem. Anal. 2013, 24, 213–223. [Google Scholar] [CrossRef]
- Alañón, M.E.; Ivanović, M.; Gómez-Caravaca, A.M.; Arráez-Román, D.; Segura-Carretero, A. Choline chloride derivative-based deep eutectic liquids as novel green alternative solvents for extraction of phenolic compounds from olive leaf. Arab. J. Chem. 2020, 13, 1685–1701. [Google Scholar] [CrossRef]
- Nediani, C.; Ruzzolini, J.; Romani, A.; Calorini, L. Oleuropein, a Bioactive Compound from Olea europaea L., as a Potential Preventive and Therapeutic Agent in Non-Communicable Diseases. Antioxidants 2019, 8, 578. [Google Scholar] [CrossRef]
- Wang, X.F.; Li, C.; Shi, Y.P.; Di, D.L. Two new secoiridoid glycosides from the leaves of Olea europaea L. J. Asian Nat. Prod. Res. 2009, 11, 940–944. [Google Scholar] [CrossRef]
- He, Z.D.; But, P.P.H.; Chan, T.W.D.; Dong, H.; Xu, H.X.; Lau, C.P.; Sun, H.D. Antioxidative Glucosides from the Fruits of Ligustrum Lucidum. Chem Pahrm. Bull. 2001, 49, 780–784. [Google Scholar] [CrossRef]
- Naruszewicz, M.; Czerwiska, M.E.; Kiss, A.K. Oleacein. Translation from Mediterranean Diet to Potential Antiatherosclerotic Drug. Curr. Pharm. Des. 2015, 21, 1205–1212. [Google Scholar] [CrossRef]
- Sanchez de Medina, V.; Luque de Castro, M.D.; Miho, H.; Melliou, E.; Magiatis, P.; Priego-Capote, F. Quantitative method for determination of oleocanthal and oleacein in virgin olive oils by liquid chromatography-tandem mass spectrometry. Talanta 2017, 162, 24–31. [Google Scholar] [CrossRef]
- Celano, R.; Piccinelli, A.L.; Pugliese, A.; Carabetta, S.; di Sanzo, R.; Rastrelli, L.; Russo, M. Insights into the Analysis of Phenolic Secoiridoids in Extra Virgin Olive Oil. J. Agric. Food Chem. 2018, 66, 6053–6063. [Google Scholar] [CrossRef]
- Cádiz-Gurrea, M.d.l.L.; Pinto, D.; Delerue-Matos, C.; Rodrigues, F. Olive Fruit and Leaf Wastes as Bioactive Ingredients for Cosmetics—A Preliminary Study. Antioxidants 2021, 10, 245. [Google Scholar] [CrossRef]
- Qi, R.; Jin, W.; Wang, J.; Yi, Q.; Yu, M.; Xu, S.; Jin, W. Oleanolic acid enhances the radiosensitivity of tumor cells under mimetic hypoxia through the reduction in intracellular GSH content and HIF-1α expression. Oncol. Rep. 2014, 31, 2399–2406. [Google Scholar] [CrossRef]
- Lins, P.G.; Marina Piccoli Pugine, S.; Scatolini, A.M.; de Melo, M.P. In vitro antioxidant activity of olive leaf extract (Olea europaea L.) and its protective effect on oxidative damage in human erythrocytes. Heliyon 2018, 4, e00805. [Google Scholar] [CrossRef]
- Jimenez Trinidad, F.R.; Arrieta Ruiz, M.; Solanes Batlló, N.; Vea Badenes, À.; Bobi Gibert, J.; Valera Cañellas, A.; Roqué Moreno, M.; Freixa Rofastes, X.; Sabaté Tenas, M.; Dantas, A.P.; et al. Linking In Vitro Models of Endothelial Dysfunction with Cell Senescence. Life 2021, 11, 1323. [Google Scholar] [CrossRef]
- Coppé, J.P.; Desprez, P.Y.; Krtolica, A.; Campisi, J. The senescence-associated secretory phenotype: The dark side of tumor suppression. Annu. Rev. Pathol. 2010, 5, 99–118. [Google Scholar] [CrossRef]
- Nguyen, H.Q.; To, N.H.; Zadigue, P.; Kerbrat, S.; De La Taille, A.; Le Gouvello, S.; Belkacemi, Y. Ionizing radiation-induced cellular senescence promotes tissue fibrosis after radiotherapy. A review. Crit. Rev. Oncol. Hematol. 2018, 129, 13–26. [Google Scholar] [CrossRef]
- Banerjee, P.; Kotla, S.; Reddy Velatooru, L.; Abe, R.J.; Davis, E.A.; Cooke, J.P.; Schadler, K.; Deswal, A.; Herrmann, J.; Lin, S.H.; et al. Senescence-Associated Secretory Phenotype as a Hinge Between Cardiovascular Diseases and Cancer. Front. Cardiovasc. Med. 2021, 8, 763930. [Google Scholar] [CrossRef]
- Itahana, K.; Campisi, J.; Dimri, G.P. Methods to detect biomarkers of cellular senescence: The senescence-associated beta-galactosidase assay. Methods Mol. Biol. 2007, 371, 21–31. [Google Scholar] [CrossRef]
- Wang, Y.; Boerma, M.; Zhou, D. Ionizing Radiation-Induced Endothelial Cell Senescence and Cardiovascular Diseases. Radiat. Res. 2016, 186, 153–261. [Google Scholar] [CrossRef]
- Manti, L.; Perozziello, F.M.; Borghesi, M.; Candiano, G.; Chaudhary, P.; Cirrone, G.A.P.; Doria, D.; Gwynne, D.; Leanza, R.; Prise, K.M.; et al. The radiobiology of laser-driven particle beams: Focus on sub-lethal responses of normal human cells. J. Instrum. 2017, 12, C03084. [Google Scholar] [CrossRef]
- Fenech, M. The in vitro micronucleus technique. Mutat. Res. 2000, 455, 81–95. [Google Scholar] [CrossRef]
- Manti, L.; Durante, M.; Cirrone, G.A.P.; Grossi, G.; Lattuada, M.; Pugliese, M.; Sabini, M.G.; Scampoli, P.; Valastro, L.; Gialanella, G. Modelled microgravity does not modify the yield of chromosome aberrations induced by high-energy protons in human lymphocytes. Int. J. Radiat. Biol. 2005, 81, 147–155. [Google Scholar] [CrossRef]
- Manti, L.; Durante, M.; Grossi, G.; Ortenzia, O.; Pugliese, M.; Scampoli, P.; Gialanella, G. Measurements of metaphase and interphase chromosome aberrations transmitted through early cell replication rounds in human lymphocytes exposed to low-LET protons and high-LET 12C ions. Mutat. Res. 2006, 596, 151–165. [Google Scholar] [CrossRef]
- Manti, L.; Jamali, M.; Prise, K.M.; Michael, B.D.; Trott, K.R. Genomic instability in Chinese hamster cells after exposure to X rays or alpha particles of different mean linear energy transfer. Radiat. Res. 1997, 147, 22–28. [Google Scholar] [CrossRef]
- Sannino, A.; Zeni, O.; Romeo, S.; Massa, R.; Gialanella, G.; Grossi, G.; Manti, L.; Vijayalaxmi Scarfì, M.R. Adaptive response in human blood lymphocytes exposed to non-ionizing radiofrequency fields: Resistance to ionizing radiation-induced damage. J. Radiat. Res. 2014, 55, 210–217. [Google Scholar] [CrossRef]
- Seshacharyulu, P.; Baine, M.J.; Souchek, J.J.; Menning, M.; Kaur, S.; Yan, Y.; Ouellette, M.M.; Jain, M.; Lin, C.; Batra, S.K. Biological determinants of radioresistance and their remediation in pancreatic cancer. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 69–92. [Google Scholar] [CrossRef]
- Sabetta, W.; Mascio, I.; Squeo, G.; Gadaleta, S.; Flamminii, F.; Conte, P.; Di Mattia, C.D.; Piga, A.; Caponio, F.; Montemurro, C. Bioactive Potential of Minor Italian Olive Genotypes from Apulia, Sardinia and Abruzzo. Foods 2021, 10, 1371. [Google Scholar] [CrossRef]
- Ryan, D.; Antolovich, M.; Herlt, T.; Prenzler, P.D.; Lavee, S.; Robards, K. Identification of phenolic compounds in tissues of the novel olive cultivar hardy’s mammoth. J. Agric. Food Chem. 2002, 50, 6716–6724. [Google Scholar] [CrossRef]
- Bláha, P.; Feoli, C.; Agosteo, S.; Calvaruso, M.; Cammarata, F.P.; Catalano, R.; Ciocca, M.; Cirrone, G.A.P.; Conte, V.; Cuttone, G.; et al. The Proton-Boron Reaction Increases the Radiobiological Effectiveness of Clinical Low- and High-Energy Proton Beams: Novel Experimental Evidence and Perspectives. Front. Oncol. 2021, 11, 682647. [Google Scholar] [CrossRef]
- Schillaci, F.; Anzalone, A.; Cirrone, G.A.P.; Carpinelli, M.; Cuttone, G.; Cutroneo, M.; De Martinis, C.; Giove, D.; Korn, G.; Maggiore, M.; et al. ELIMED, MEDical and multidisciplinary applications at ELI-Beamlines. J. Phys. Conf. Ser. 2014, 508, 012010. [Google Scholar] [CrossRef]
- Cárdeno, A.; Sánchez-Hidalgo, M.; Rosillo, M.A.; Alarcón de la Lastra, C. Oleuropein, a secoiridoid derived from olive tree, inhibits the proliferation of human colorectal cancer cell through downregulation of HIF-1α. Nutr. Cancer 2013, 65, 147–156. [Google Scholar] [CrossRef]
- Benot-Dominguez, R.; Tupone, M.G.; Castelli, V.; d’Angelo, M.; Benedetti, E.; Quintiliani, M.; Cinque, B.; Forte, I.M.; Cifone, M.G.; Ippoliti, R.; et al. Olive leaf extract impairs mitochondria by pro-oxidant activity in MDA-MB-231 and OVCAR-3 cancer cells. Biomed. Pharmacother. 2021, 134, 111139. [Google Scholar] [CrossRef]
- Hahn, D.; Shin, S.H.; Bae, J.S. Natural Antioxidant and Anti-Inflammatory Compounds in Foodstuff or Medicinal Herbs Inducing Heme Oxygenase-1 Expression. Antioxidants 2020, 9, 1191. [Google Scholar] [CrossRef]
- Olivares-Vicente, M.; Barrajon-Catalan, E.; Herranz-Lopez, M.; Segura-Carretero, A.; Joven, J.; Encinar, J.A.; Micol, V. Plant-Derived Polyphenols in Human Health: Biological Activity, Metabolites and Putative Molecular Targets. Curr. Drug Metab. 2018, 19, 351–369. [Google Scholar] [CrossRef]
- Nabiałek-Trojanowska, I.; Lewicka, E.; Wrona, A.; Kaleta, A.M.; Lewicka-Potocka, Z.; Raczak, G.; Dziadziuszko, R. Cardiovascular complications after radiotherapy. Cardiol. J. 2020, 27, 836–847. [Google Scholar] [CrossRef]
- Bian, Y.; Wei, J.; Zhao, C.; Li, G. Natural Polyphenols Targeting Senescence: A Novel Prevention and Therapy Strategy for Cancer. Int. J. Mol. Sci. 2020, 21, 684. [Google Scholar] [CrossRef]
- Csekes, E.; Račková, L. Skin Aging, Cellular Senescence and Natural Polyphenols. Int. J. Mol. Sci. 2021, 22, 12641. [Google Scholar] [CrossRef]
- Menicacci, B.; Cipriani, C.; Margheri, F.; Mocali, A.; Giovannelli, L. Modulation of the Senescence-Associated Inflammatory Phenotype in Human Fibroblasts by Olive Phenols. Int. J. Mol. Sci. 2017, 18, 2275. [Google Scholar] [CrossRef]
- Gurău, F.; Baldoni, S.; Prattichizzo, F.; Espinosa, E.; Amenta, F.; Procopio, A.D.; Albertini, M.C.; Bonafè, M.; Olivieri, F. Anti-senescence compounds: A potential nutraceutical approach to healthy aging. Ageing Res. Rev. 2018, 46, 14–31. [Google Scholar] [CrossRef]
- Frediani, E.; Scavone, F.; Laurenzana, A.; Chillà, A.; Tortora, K.; Cimmino, I.; Leri, M.; Bucciantini, M.; Mangoni, M.; Fibbi, G.; et al. Olive phenols preserve lamin B1 expression reducing cGAS/STING/NFκB-mediated SASP in ionizing radiation-induced senescence. J. Cell. Mol. Med. 2022, 26, 2337–2350. [Google Scholar] [CrossRef]
- Greenrod, W.; Fenech, M. The principal phenolic and alcoholic components of wine protect human lymphocytes against hydrogen peroxide- and ionizing radiation-induced DNA damage in vitro. Mutagenesis 2003, 18, 119–126. [Google Scholar] [CrossRef]
- Alcaraz, M.; Olivares, A.; Achel, D.G.; García-Gamuz, J.A.; Castillo, J.; Alcaraz-Saura, M. Genoprotective Effect of Some Flavonoids against Genotoxic Damage Induced by X-rays In Vivo: Relationship between Structure and Activity. Antioxidants 2021, 11, 94. [Google Scholar] [CrossRef]
- Amani, F.; Allahbakhshian Farsani, M.; Gholami, M.; Aghamiri, S.M.R.; Bakhshandeh, M.; Hossein Mohammadi, M. The protective effect of oleuropein against radiation-induced cytotoxicity, apoptosis, and genetic damage in cultured human lymphocytes. Int. J. Radiat. Biol. 2021, 97, 179–193. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, Z.; Yang, X.; Liu, L.; Ahn, K.S. An updated review on the potential antineoplastic actions of oleuropein. Phytother. Res. 2022, 36, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Cui, D.; Wang, S.; Wang, P.; Xing, X.; Li, H. Oleuropein represses the radiation resistance of ovarian cancer by inhibiting hypoxia and microRNA-299-targetted heparanase expression. Food Funct. 2017, 8, 2857–2864. [Google Scholar] [CrossRef] [PubMed]
- Acquaviva, R.; Di Giacomo, C.; Sorrenti, V.; Galvano, F.; Santangelo, R.; Cardile, V.; Gangia, S.; D’Orazio, N.; Abraham, N.G.; Vanella, L. Antiproliferative effect of oleuropein in prostate cell lines. Int. J. Oncol. 2012, 41, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Bektay, M.Y.; Güler, E.M.; Gökçe, M.; Kızıltaş, M.V. Investigation of the Genotoxic, Cytotoxic, Apoptotic, and Oxidant Effects of Olive Leaf Extracts on Liver Cancer Cell Lines. Turk. J. Pharm. Sci. 2021, 18, 781–789. [Google Scholar] [CrossRef]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and prooxidant behavior of flavonoids: Structure-activity relationships. Free Radic. Biol. Med. 1997, 22, 749–760. [Google Scholar] [CrossRef]
- Theofylaktou, D.; Takan, I.; Karakülah, G.; Biz, G.M.; Zanni, V.; Pavlopoulou, A.; Georgakilas, A.G. Mining Natural Products with Anticancer Biological Activity through a Systems Biology Approach. Oxid. Med. Cell Longev. 2021, 2021, 9993518. [Google Scholar] [CrossRef]
- Aponte, M.; Ungaro, F.; d’Angelo, I.; De Caro, C.; Russo, R.; Blaiotta, G.; Dal Piaz, F.; Calignano, A.; Miro, A. Improving in vivo conversion of oleuropein into hydroxytyrosol by oral granules containing probiotic Lactobacillus plantarum 299v and an Olea europaea standardized extract. Int. J. Pharm. 2018, 543, 73–82. [Google Scholar] [CrossRef]
- de Bock, M.; Thorstensen, E.B.; Derraik, J.G.; Henderson, H.V.; Hofman, P.L.; Cutfield, W.S. Human absorption and metabolism of oleuropein and hydroxytyrosol ingested as olive (Olea europaea L.) leaf extract. Mol. Nutr. Food Res. 2013, 57, 2079–2085. [Google Scholar] [CrossRef]
- López-Yerena, A.; Vallverdú-Queralt, A.; Lamuela-Raventós, R.M.; Escribano-Ferrer, E. LC-ESI-LTQ-Orbitrap-MS for profiling the distribution of oleacein and its metabolites in rat tissues. Antioxidants 2021, 10, 1083. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak n. | RT (min) | Tentative Assignment | Formula | [M-H]−calc. (m/z) | [M-H]− Found (m/z) | Error (ppm) | RDB | HR-MS/MS Fragment Ions (m/z) (Base Peaks in Bold) |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.316 | Hexitol | C6H14O6 | 181.0718 | 181.0719 | 0.8 | 0 | 181.0722; 163.0636; 136.8708; 119.0352; 113.0240; 101.0241; 89.0245 |
| 2 | 0.326 | Quinic acid | C7H12O6 | 191.0561 | 191.0565 | 2.0 | 2 | 191.0558; 173.0447; 127.0403; 109.0293; 93.0348 |
| 3 | 1.651 | Hydroxytyrosol | C8H10O3 | 153.0557 | 153.0557 | 0.0 | 4 | 153.0561; 123.0454 |
| 4 | 1.835 | Hydroxytyrosol hexoside | C14H20O8 | 315.1085 | 315.1084 | −0.4 | 5 | 315.1081; 153.0556; 135.0454; 123.0453; 119.0352; 119.0352; 113.0251; 101.0252; 89.0254 |
| 5 | 3.375 | 12-hydroxyjasmonate sulfate | C12H18O7S | 305.0700 | 305.0702 | 0.5 | 4 | 305.0705; 225.1135; 174.9560; 147.0816; 130.9665; 96.9606 |
| 6 | 4.509 | Unknown | C16H26O10 | 377.1453 | 377.1454 | 0.2 | 4 | 377.1455; 197.0824; 153.0925 |
| 7 | 5.329 | Phenethyl primeveroside | C19H28O10 | 415.1610 | 415.1609 | −0.2 | 6 | 191.0557; 179.0566; 161.0455; 149.0458; 131.0359; 119.0356; 113.0245; 101.0252; 89.0250 |
| 8 | 6.377 | 2-(2-ethyl-3-hydroxy-6-propionylcyclohexyl) acetic acid glucoside | C19H32O9 | 403.1974 | 403.1977 | 0.9 | 4 | 403.1992; 241.1448; 223.1346; 161.0463; 119.0355; 113.0252; 101.0253; 89.0247 |
| 9 | 6.482 | Luteolin di-hexoside | C27H30O6 | 609.1461 | 609.1463 | 0.3 | 13 | 609.1486; 447.0949; 285.0403 |
| 10 | 7.789 | Rutin | C27H30O6 | 609.1461 | 609.1472 | 1.8 | 13 | 609.1482; 301.0347; 300.0271; 271.0229 |
| 11 | 8.058 | Luteolin hexoside 1 | C21H20O11 | 447.0933 | 447.0935 | 0.5 | 12 | 447.0945; 285.0405;284.0329 |
| 12 | 8.063 | Verbascoside | C29H36O15 | 623.1981 | 623.1983 | 0.2 | 12 | 623.1999; 461.1665; 161.0243 |
| 13 | 8.958 | Oleacein | C17H20O6 | 319.1187 | 319.1190 639.2455 [2M-H]− | 0.9 | 8 | 183.0670; 165.0561; 139.0772; 123.0454; 113.0251; 95.0512 |
| 14 | 9.194 | Quercetin deoxyhexoside | C21H20O11 | 447.0933 | 447.0931 | −0.4 | 12 | 447.0950; 301.0359; 300.0279; 271.0246; 255.0290; 178.9986; 151.0036 |
| 15 | 9.437 | Lipedoside A isomer 1 | C29H36O14 | 607.2032 | 607.2029 | −0.5 | 12 | 607.2027; 461.1684; 163.0400; 145.0296 |
| 16 | 9.450 | Apigenin hexosyldeoxyhexoside 1 | C27H30O14 | 577.1563 | 577.1563 | 0 | 13 | 577.1575; 269.0447; 268.0371 |
| 17 | 9.551 | Luteolin hexoside 2 | C21H20O11 | 447.0933 | 447.0940 | 1.6 | 12 | 285.0403 |
| 18 | 9.588 | Oleuropein hexoside | C31H42O18 | 701.2298 | 701.2287 | −1.6 | 11 | 701.2339; 539.1800; 469.1362; 437.1110; 377.1253; 307.0811; 275.0923; 179.0567 |
| 19 | 9.620 | Lucidumoside B | C25H34O13 | 541.1927 | 541.1931 | 0.9 | 9 | 361.1293; 225.0770; 193.0504; 181.0872; 149.0606; 121.0659; 89.0248 |
| 20 | 9.873 | Apigenin hexosyldeoxyhexoside 2 | C27H30O14 | 577.1563 | 577.1577 | 2.5 | 13 | 577.1577; 269.0450 |
| 21 | 9.865 | Lipedoside A isomer 2 | C29H36O14 | 607.2032 | 607.2044 | 1.9 | 12 | 607.2049; 461.1679; 163.0404; 145.0295 |
| 22 | 10.183 | Oleuropein | C25H32O13 | 539.1770 | 539.1784 | 2.6 | 10 | 539.1786; 403.1240; 377.1237; 345.0975; 327.0874; 307.0823; 275.0911; 223.0605; 179.0564; 149.0247; 95.0509 |
| 23 | 10.343 | Luteolin hexoside 3 | C21H20O11 | 447.0933 | 447.0930 | −0.6 | 12 | 447.0929; 285.0406 |
| 24 | 10.539 | Diosmin | C28H32O15 | 607.1668 | 607.1679 | 1.7 | 13 | 607.1689; 299.0552; 284.0311 |
| 25 | 12.136 | Luteolin | C15H10O6 | 285.0405 | 285.0406 | 0.5 | 11 | 285.0413; 217.0507; 199.0402; 175.0403; 151.0038; 133.0300; 107.0140 |
| 26 | 13.213 | Oleacein dimethyl acetal | C19H26O7 | 365.1606 | 365.1603 | −0.8 | 7 | 229.1086; 211.0976; 201.1133; 185.1185; 169.0868; 153.0922; 138.0687; 121.0665 |
| 27 | 14.273 | Apigenin | C15H10O5 | 269.0455 | 269.0453 | −0.9 | 11 | 269.0453; 225.0550; 201.0554; 183.0445; 181.0664; 159.0453; 151.0037; 149.0241; 117.0347; 107.0141 |
| 28 | 14.999 | Diosmetin | C16H12O6 | 299.0561 | 299.0561 | 0.0 | 11 | 299.0559; 284.0321; 256.0374 |
| 29 | 15.313 | Oleacein ethylmethyl acetal | C20H28O7 | 379.1762 | 379.1769 | 1.8 | 7 | 243.1243; 225.1138; 215.1285; 199.1338; 167.1075; 153.0921; 138.0690; 121.0659 |
| 30 | 17.198 | Oleacein diethyl acetal | C21H30O7 | 393.1919 | 393.1925 | 1.6 | 7 | 257.1399; 239.1289; 229.1446; 213.1497; 167.1080; 139.0766; 121.0664 |
| 31 | 17.671 | Oleuropein derivative | C36H44O18 | 763.2455 | 763.2461 | 0.8 | 15 | 693.2048; 539.1785; 461.0730; 377.1250; 307.0829; 275.0904; 149.0237 |
| 32 | 25.740 | Oleanolic acid | C30H48O3 | 455.3531 | 455.3527 | −0.9 | 7 | 455.3531; 407.3318; 405.3162; 373.2506; 345.2192 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacifico, S.; Bláha, P.; Faramarzi, S.; Fede, F.; Michaličková, K.; Piccolella, S.; Ricciardi, V.; Manti, L. Differential Radiomodulating Action of Olea europaea L. cv. Caiazzana Leaf Extract on Human Normal and Cancer Cells: A Joint Chemical and Radiobiological Approach. Antioxidants 2022, 11, 1603. https://doi.org/10.3390/antiox11081603
Pacifico S, Bláha P, Faramarzi S, Fede F, Michaličková K, Piccolella S, Ricciardi V, Manti L. Differential Radiomodulating Action of Olea europaea L. cv. Caiazzana Leaf Extract on Human Normal and Cancer Cells: A Joint Chemical and Radiobiological Approach. Antioxidants. 2022; 11(8):1603. https://doi.org/10.3390/antiox11081603
Chicago/Turabian StylePacifico, Severina, Pavel Bláha, Shadab Faramarzi, Francesca Fede, Katarina Michaličková, Simona Piccolella, Valerio Ricciardi, and Lorenzo Manti. 2022. "Differential Radiomodulating Action of Olea europaea L. cv. Caiazzana Leaf Extract on Human Normal and Cancer Cells: A Joint Chemical and Radiobiological Approach" Antioxidants 11, no. 8: 1603. https://doi.org/10.3390/antiox11081603