
Imaging of Lipid Peroxidation-Associated Chemiluminescence in Plants: Spectral Features, Regulation and Origin of the Signal in Leaves and Roots


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Oxidation of Lipids In Vitro
2.3. Luminescence Imaging
2.4. Hydroxy Fatty Acid Measurements
2.5. SOSG-EP Fluorescence Imaging and Spectroscopy
2.6. Hydroxy-Plastochromanol Measurements
3. Results
3.1. Spectral Characteristics of Plant Leaf SCL
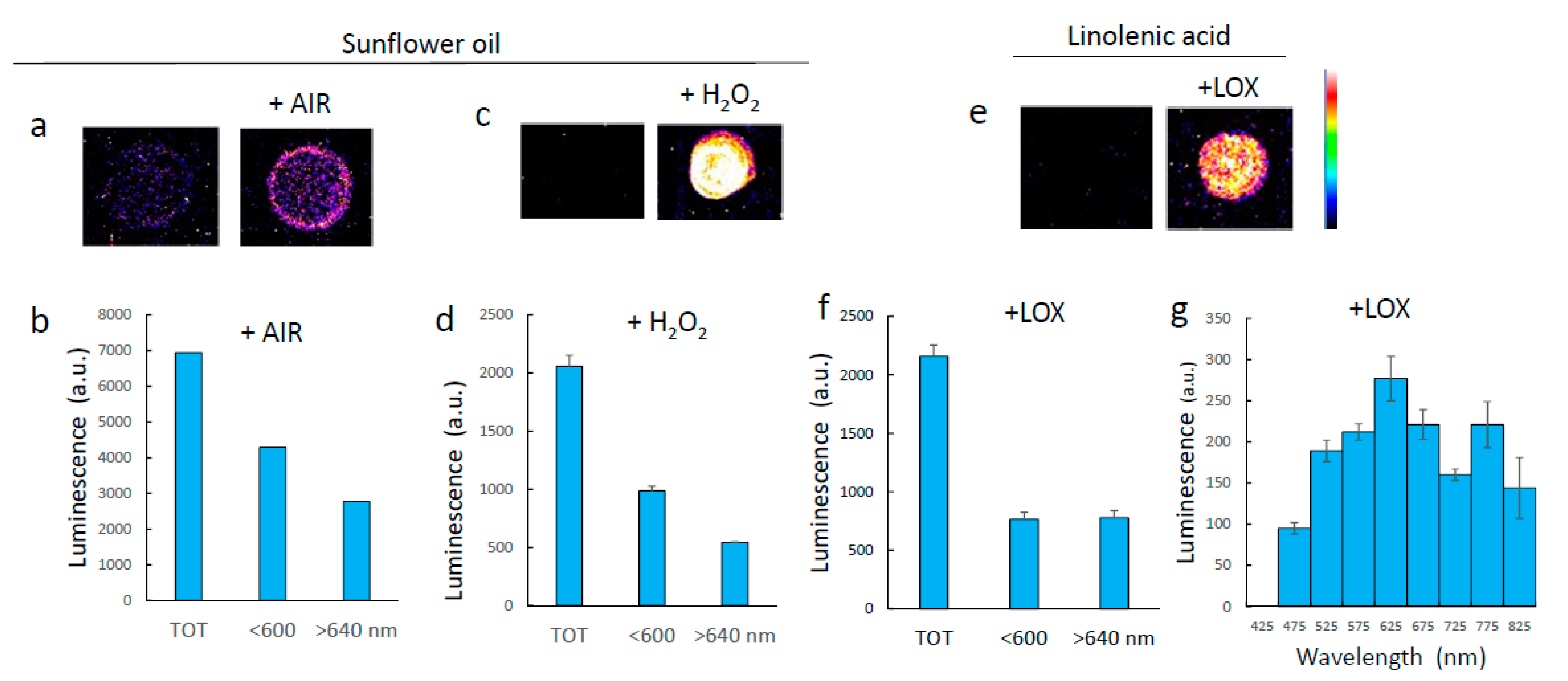
3.2. Luminescence of Lipids Oxidized In Vitro
3.3. Non-Chlorophyllous Material
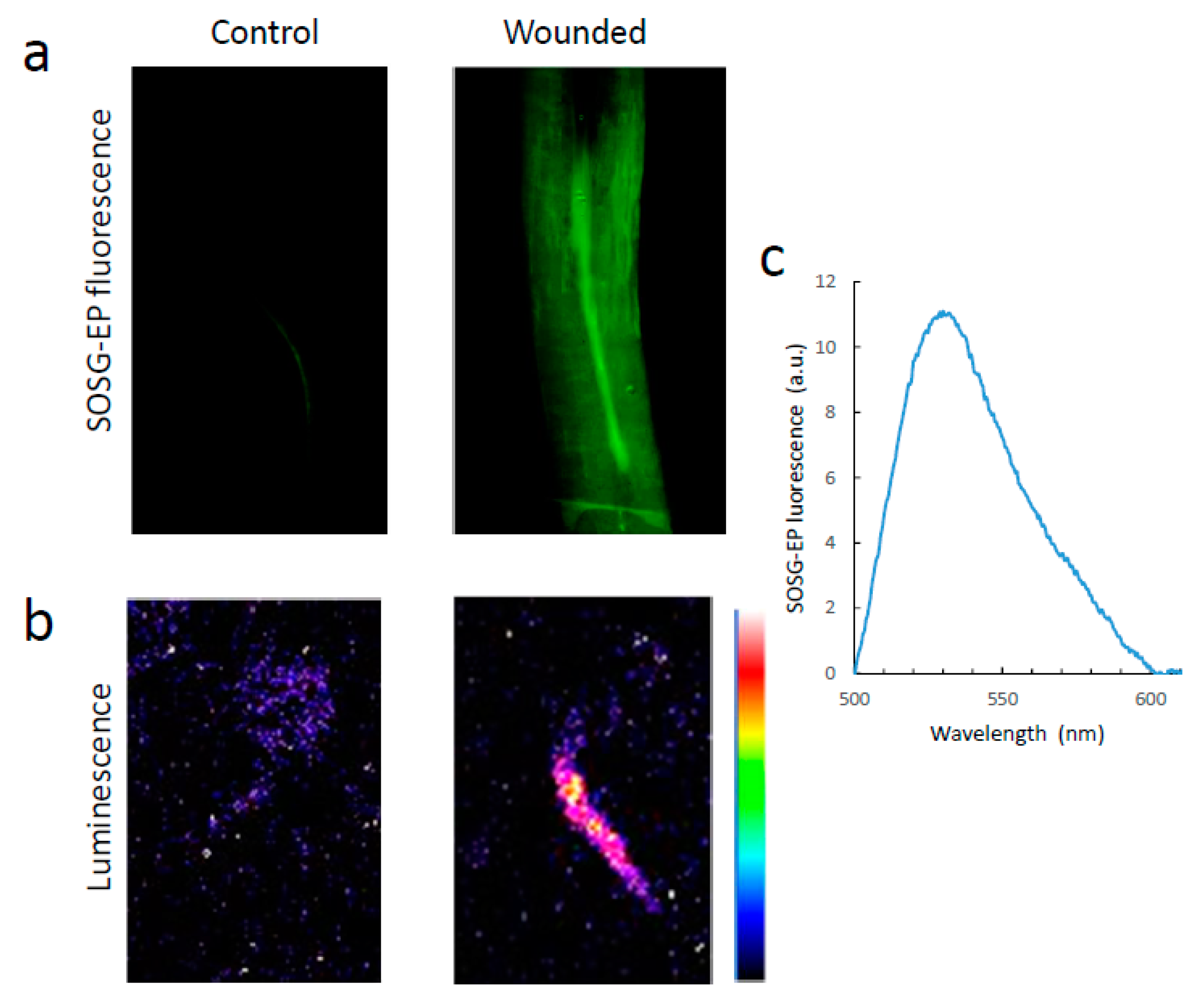
3.4. Singlet Oxygen
3.5. Detoxification of Lipid Peroxidation Products
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cadenas, E. Biological chemiluminescence. Photochem. Photobiol. 1984, 40, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M.; Triantaphylidès, C.; Genty, B. Autoluminescence imaging: A non-invasive tool for mapping oxidative stress. Trends Plant Sci. 2006, 11, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Cifra, M.; Pospisil, P. Ultra-weak photon emission from biological samples: Definition, mechanisms, properties, detection and applications. J. Photochem. Photobiol. B Biol. 2014, 139, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Naumova, E.V.; Vladimirovb, Y.A.; Beloussovb, L.V.; Tuchinc, V.V.; Volodyaevb, I.V. Methods of studying ultraweak photon emission from biological objects: I. History, types and properties, fundamental and application significance. Biophysics 2021, 66, 764–778. [Google Scholar] [CrossRef]
- Wilson, T.; Hastings, J.W. Bioluminescence. Annu. Rev. Cell Dev. Biol. 1998, 14, 197–230. [Google Scholar] [CrossRef]
- Birtic, S.; Ksas, B.; Genty, B.; Mueller, M.J.; Triantaphylidès, C.; Havaux, M. Using spontaneous photon emission to image lipid oxidation patterns in plant tissues. Plant J. 2011, 67, 1103–1115. [Google Scholar] [CrossRef]
- Miquel, M.; Browse, J. Arabidopsis mutants deficient in polyunsaturated fatty acid synthesis. Biochemical and genetic characterization of a plant oleoyl-phosphatidylcholine desaturase. J. Biol. Chem. 1992, 267, 1502–1509. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Acyl-lipid metabolism. Arab. Book 2013, 11, e0161. [Google Scholar] [CrossRef] [Green Version]
- Bour, A.; Kruglik, S.G.; Chabanon, M.; Rangamani, P.; Puff, N.; Bonneau, S. Lipid unsaturation properties govern the sensitivity of membranes to photoinduced oxidative stress. Biophys. J. 2019, 116, 910–920. [Google Scholar] [CrossRef] [Green Version]
- Ksas, B.; Légeret, B.; Ferretti, U.; Chevalier, A.; Pospíšil, P.; Alric, J.; Havaux, M. The plastoquinone pool outside the thylakoid membrane serves in plant photoprotection as a reservoir of singlet oxygen scavengers. Plant Cell Environ. 2018, 41, 2277–2287. [Google Scholar] [CrossRef]
- Boca, S.; Koestler, F.; Ksas, B.; Chevalier, A.; Leymarie, J.; Fekete, A.; Mueller, M.J.; Havaux, M. Arabidopsis lipocalins AtCHL and AtTIL have distinct but overlapping functions essential for lipid protection and seed longevity. Plant Cell Environ. 2014, 37, 368–381. [Google Scholar] [CrossRef] [PubMed]
- Malnoë, A.; Schultink, A.; Shahrasbi, S.; Rumeau, D.; Havaux, M.; Niyogi, K.K. The plastid lipocalin LCNP is required for sustained photoprotective energy dissipation in Arabidopsis. Plant Cell 2018, 30, 196–208. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.P.; Havaux, M.; Triantaphylidès, C.; Ksas, B.; Pascal, A.A.; Robert, B.; Davison, P.A.; Ruban, A.V.; Horton, P. Elevated zeaxanthin bound to oligomeric LHCII enhances the resistance of Arabidopsis to photooxidative stress by a lipid-protective, antioxidant mechanism. J. Biol. Chem. 2007, 282, 22605–22618. [Google Scholar] [CrossRef] [Green Version]
- Triantaphylides, C.; Krischke, M.; Hoeberichts, F.A.; Ksas, B.; Gresser, G.; Havaux, M.; Van Breusegem, F.; Mueller, M.J. Singlet oxygen is the major reactive oxygen species involved in photooxidative damage to plants. Plant Physiol. 2008, 148, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duran, N.; Cadenas, E. The role of singlet oxygen and triplet carbonyls in biological systems. Rev. Chem. Interact. 1987, 8, 147–187. [Google Scholar] [CrossRef]
- Wang, C.; Sullivan, R.W.; Kight, A.; Henry, R.L.; Huang, J.; Jomes, A.M.; Korth, K.L. Deletion of the chloroplast-localized Thylakoid formation 1 gene product in Arabidopsis leads to deficient thylakoid formation and variegated leaves. Plant Physiol. 2004, 136, 3594–3604. [Google Scholar] [CrossRef] [Green Version]
- Glauser, G.; Dubugnon, L.; Mousavi, S.A.; Rudaz, S.; Wolfender, J.L.; Farmer, E.E. Velocity estimates for signal propagation leading to systemic jasmonic acid accumulation in wounded Arabidopsis. J. Biol. Chem. 2009, 284, 34506–34513. [Google Scholar] [CrossRef] [Green Version]
- Rac, M.; Shumbe, L.; Oger, C.; Guy, A.; Vigor, C.; Ksas, B.; Durand, T.; Havaux, M. Luminescence imaging of leaf damage induced by lipid peroxidation products and its modulation by β-cyclocitral. Physiol. Plant. 2021, 171, 246–259. [Google Scholar] [CrossRef]
- Fatima, K.; Masood, N.; Luqman, S. Quenching of singlet oxygen by natural and synthetic antioxidants and assessment of electronic UV/visible absorption spectra for alleviating or enhancing the efficacy of photodynamic therapy. Biomed. Res. Ther. 2016, 3, 514–527. [Google Scholar] [CrossRef]
- Ogilby, P.R.; Foote, C.S. Chemistry of singlet oxygen. 42. Effect of solvent, solvent isotopic substitution, and temperature on the lifetime of singlet oxygen molecular oxygen (1∆g). J. Am. Chem. Soc. 1983, 105, 3423–3430. [Google Scholar] [CrossRef]
- Montillet, J.-L.; Cacas, J.-L.; Garnier, L.; Montané, M.-H.; Douki, T.; Bessoule, J.-J.; Polkowska-Kowalczyk, L.; Maciejewska, U.; Agnel, J.-P.; Vial, A.; et al. The upstream oxylipin profile of Arabidopsis thaliana: A tool to scan for oxidative stresses: Lipid peroxidation in Arabidopsis. Plant J. 2004, 40, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Ksas, B.; Havaux, M. Determination of ROS-induced lipid peroxidation by HPLC-based quantification of hydroxy polyunsaturated fatty acids. Methods Mol. Biol. 2022, 2526. in press. [Google Scholar]
- Ramel, F.; Ksas, B.; Akkari, E.; Mialoundama, A.S.; Monnet, F.; Krieger-Liszkay, A.; Ravanat, J.L.; Mueller, M.J.; Bouvier, F.; Havaux, M. Light-induced acclimation of the Arabidopsis chlorina1 mutant to singlet oxygen. Plant Cell 2013, 25, 1445–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, E.; Creelman, R.A.; Mullet, J.E. A chlorophyll lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1995, 92, 8675–8679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reymond, P.; Weber, H.; Damond, M.; Farmer, E.E. Differential gene expression in response to mechanical wounding and insect feeding in Arabidopsis. Plant Cell 2000, 12, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.L.; Xing, D.; Tan, S.C.; Tang, Y.H.; He, Y.H. Imaging of ultraweak bio-chemiluminescence and singlet oxygen generation in germinating soybean in response to wounding. Luminescence 2003, 18, 37–41. [Google Scholar] [CrossRef]
- Flor-Henry, M.; McCabe, T.C.; de Bruxelles, G.L.; Roberts, M.R. Use of a highly sensitive two-dimensional luminescence imaging system to monitor endogenous bioluminescence in plant leaves. BMC Plant Biol. 2004, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Cendrero-Mateo, M.P.; Moran, M.S.; Papuga, S.A.; Thorp, K.R.; Alonso, L.; Moreno, J.; Ponce-Campos, G.; Rascher, U.; Wang, G. Plant chlorophyll fluorescence: Active and passive measurements at canopy and leaf scales with different nitrogen treatments. J. Exp. Bot. 2016, 67, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Hideg, E.; Kobayashi, M.; Inaba, H. Utraweak photoemission from dark-adapted leaves and isolated chloroplast. FEBS Lett. 1990, 275, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Hideg, E. On the spontaneous ultraweak light emission of plants. J. Photochem. Photobiol. 1993, 18, 239–244. [Google Scholar] [CrossRef]
- Brunetti, I.; Cilento, G.; Nassi, L. Energy transfer from enzymatically-gerenerated triplet species to acceptors in micelles. Photochem. Photobiol. 1983, 38, 511–519. [Google Scholar] [CrossRef]
- Bohne, C.; Campa, A.; Cilento, G.; Nassi, L.; Villablanca, M. Chlorophyll: An efficient detector of electronically excited species in biochemical systems. Anal. Biochem. 1986, 155, 1–9. [Google Scholar] [CrossRef]
- Salin, M.L.; Bridges, S.M. Chemiluminescence in wounded root tissue. Plant Physiol. 1981, 67, 43–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasad, A.; Sedlářová, M.; Balukova, A.; Rác, M.; Pospíšil, P. Reactive Oxygen Species as a Response to Wounding: In Vivo Imaging in Arabidopsis thaliana. Front. Plant Sci. 2020, 10, 1660. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Sedlářová, M.; Pospíšil, P. Singlet oxygen imaging using fluorescent probe Singlet Oxygen Sensor Green in photosynthetic organisms. Sci. Rep. 2018, 8, 13685. [Google Scholar] [CrossRef]
- Szymanska, R.; Nowicka, B.; Kruk, J. Hydroxy-plastochromanol and plastoquinone-C as singlet oxygen products during photo-oxidative stress in Arabidopsis. Plant Cell Environ. 2014, 37, 1464–1473. [Google Scholar] [CrossRef]
- Frankel, E.N. Secondary products of lipid oxidation. Chem. Phys. Lipids 1987, 44, 73–85. [Google Scholar] [CrossRef]
- Mano, J. Reactive carbonyl species: Their production from lipid peroxides, action in environmental stress, and the detoxification mechanism. Plant Physiol. Biochem. 2012, 59, 90–97. [Google Scholar] [CrossRef]
- Mano, J.; Biswas, M.S.; Sugimoto, K. Reactive carbonyl species: A missing link in ROS signaling. Plants 2019, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Youn, B.; Kim, S.-J.; Moinuddin, S.G.A.; Lee, C.; Bedgar, D.L.; Harper, A.R.; Davin, L.B.; Lewis, N.G.; Kang, C. Mechanistic and structural studies of apoform, binary, and ternary complexes of the Arabidopsis alkenal double bond reductase At5g16970. J. Biol. Chem. 2006, 281, 40076–40088. [Google Scholar] [CrossRef] [Green Version]
- Mano, J.; Belles-Boix, E.; Babiychuk, E.; Inzé, D.; Torii, Y.; Hiraoka, E.; Takimoto, K.; Slooten, L.; Asada, K.; Kushnir, S. Protection against photooxidative injury of tobacco leaves by 2-alkenal reductase. Detoxication of lipid peroxide-derived reactive carbonyls. Plant Physiol. 2005, 139, 1773–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, Y.H.; Chang, H.S.; Gupta, R.; Wang, X.; Zhu, T.; Luan, S. Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress, and hormonal responses in Arabidopsis. Plant Physiol. 2002, 129, 661–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fode, B.; Siemsen, T.; Thurow, C.; Weigel, R.; Gatz, C. Thee Arabidopsis GRAS Protein SCL14 Interacts with Class II TGA Transcription Factors and Is Essential for the Activation of Stress-Inducible Promoters. Plant Cell 2009, 20, 3122–3135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Alessandro, S.; Ksas, B.; Havaux, M. Decoding β-cyclocitral-mediated retrograde signaling reveals the role of a detoxification response in plant tolerance to photooxidative stress. Plant Cell 2018, 30, 2495–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchida, K.; Kobayashi, M. Ultraviolet A irradiation induces ultraweak photon emission with characteristic spectral patterns from biomolecules present in human skin. Sci. Rep. 2020, 10, 21667. [Google Scholar] [CrossRef] [PubMed]
- Boveris, A.; Cadenas, E.; Reiter, R.; Filipkowski, M.; Nakase, Y.; Chance, B. Organ chemiluminescence: Noninvasive assay for oxidative radical reactions. Proc. Natl. Acad. Sci. USA 1980, 77, 347–351. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, S.; Martinez, G.R.; Medeiros, M.H.G.; Di Mascio, P. Singlet molecular oxygen generated by biological hydroperoxides. J. Photochem. Photobiol. B Biol. 2014, 139, 24–33. [Google Scholar] [CrossRef]
- Adam, W.; Kazakov, D.V.; Kazakov, V.P. Singlet-oxygen chemiluminescence in peroxide reactions. Chem. Rev. 2005, 105, 3371–3387. [Google Scholar] [CrossRef]
- Borrell, P.M.; Borrell, P.; Grant, K.R. A study of three dimol emission of singlet oxygen, 1O2 (1∆g), using a discharge flow stock tube. J. Chem. Soc. Faraday Trans. 2 1980, 76, 1442–1449. [Google Scholar] [CrossRef]
- Chen, T.; Cohen, D.; Itkin, M.; Malitsky, S.; Fluhr, R. Lipoxygenase functions in 1O2 production during root responses to osmotic stress. Plant Physiol. 2021, 185, 1638–1651. [Google Scholar] [CrossRef]
- Triantaphylides, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Gruszka, J.; Pawlak, A.; Kruk, J. Tocochromanols, plastoquinol, and other biological prenyllipids as singlet oxygen quenchers—Determination of singlet oxygen quenching rate constants and oxidation products. Free Radic. Biol. Med. 2008, 45, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Sasaki, K.; Enomoto, M.; Ehara, Y. Highly sensitive determination of transient generation of biophotons during hypersensitive response to cucumber mosaic virus in cowpea. J. Exp. Bot. 2007, 58, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Iwasa, T.; Tada, M. Polychromatic spectral pattern analysis of ultra-weak photon emissions from a human body. J. Photochem. Photobiol. B 2016, 159, 186–190. [Google Scholar] [CrossRef]
- Hofmann, N.R. A structure for plant-specific transcription factors: The GRAS domain revealed. Plant Cell 2016, 28, 993–994. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Iyer, L.M.; Aravind, L. Bacterial GRAS domain proteins throw new light on gibberellic acid response mechanisms. Bioinformatics 2012, 28, 2407–2411. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.; Hilbert, B.; Dueckershoff, K.; Roitsch, T.; Krischke, M.; Mueller, M.J.; Berger, S. General detoxification and stress responses are mediated by oxidized lipids through TGA transcription factors in Arabidopsis. Plant Cell 2008, 20, 768–785. [Google Scholar] [CrossRef] [Green Version]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havaux, M.; Ksas, B. Imaging of Lipid Peroxidation-Associated Chemiluminescence in Plants: Spectral Features, Regulation and Origin of the Signal in Leaves and Roots. Antioxidants 2022, 11, 1333. https://doi.org/10.3390/antiox11071333
Havaux M, Ksas B. Imaging of Lipid Peroxidation-Associated Chemiluminescence in Plants: Spectral Features, Regulation and Origin of the Signal in Leaves and Roots. Antioxidants. 2022; 11(7):1333. https://doi.org/10.3390/antiox11071333
Chicago/Turabian StyleHavaux, Michel, and Brigitte Ksas. 2022. "Imaging of Lipid Peroxidation-Associated Chemiluminescence in Plants: Spectral Features, Regulation and Origin of the Signal in Leaves and Roots" Antioxidants 11, no. 7: 1333. https://doi.org/10.3390/antiox11071333