
Silicon- and Boron-Induced Physio-Biochemical Alteration and Organic Acid Regulation Mitigates Aluminum Phytotoxicity in Date Palm Seedlings

 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Growth and Experimental Design
2.2. Chlorophyll a and Chlorophyll b
2.3. Quantification of Malondialdehyde (MDA)
2.4. Determination of Superoxide (O2•−)
2.5. Antioxidant Enzyme Assay
2.6. Micronutrient Quantification
2.7. Determination of Organic Acids
2.8. Abscisic Acid and Salicylic Acid Extraction and Quantification
2.9. Gene Expression Analysis
2.10. Statistical Analysis
3. Results
3.1. Effects of Si, B, and Their Combination on Date Palm Growth Parameters under Al3+ Phytotoxicity
3.2. Accumulation and Translocation of Si, B, and Al3+ in Date Palm
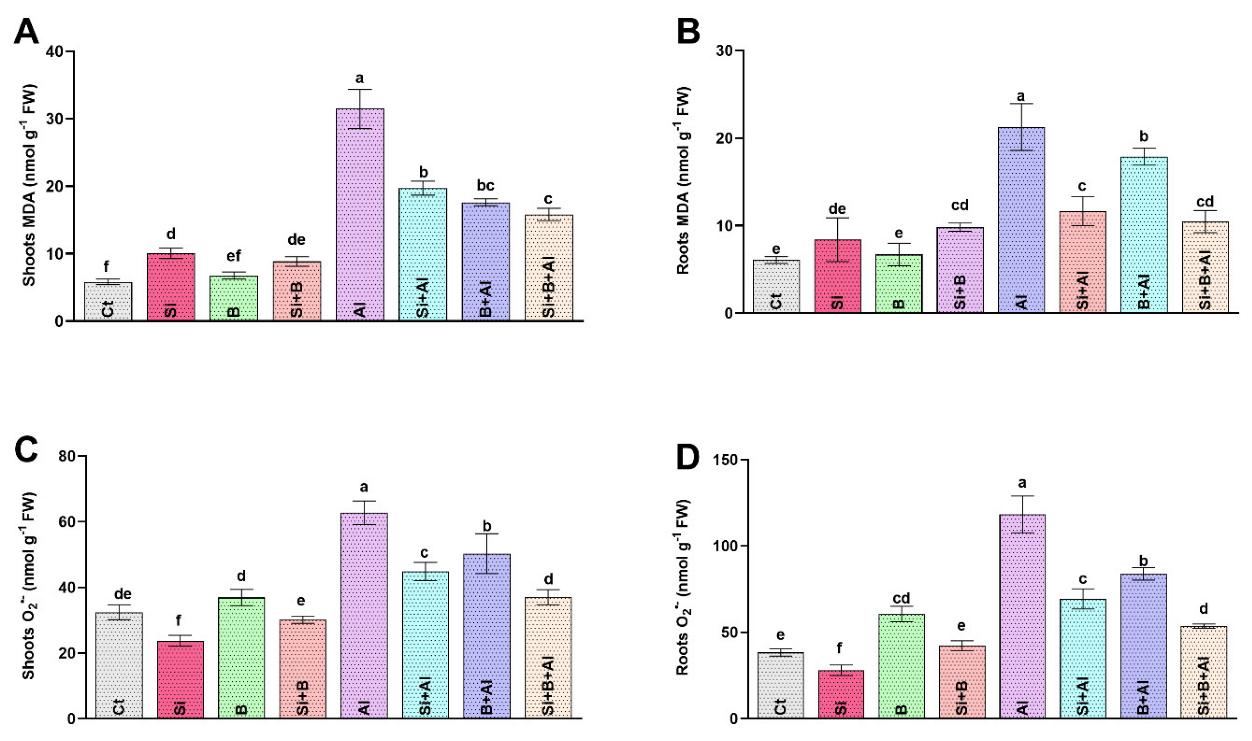
3.3. Effect of Exogenous Si, B, and Their Interaction on Lipid Peroxidation and Superoxide Anion Accumulation under Al3+ Stress
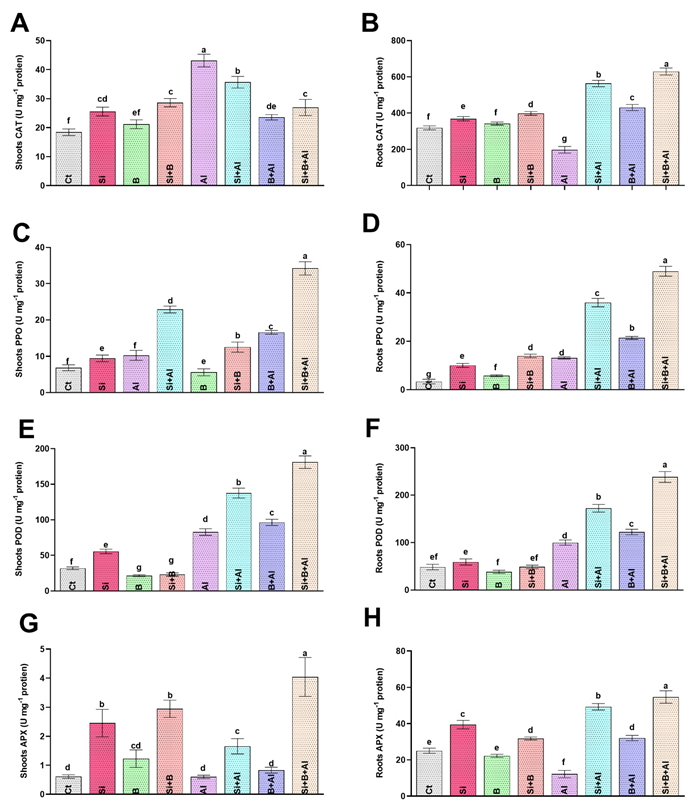
3.4. Modulation of Antioxidant Activities by Si, B, and Their Interaction
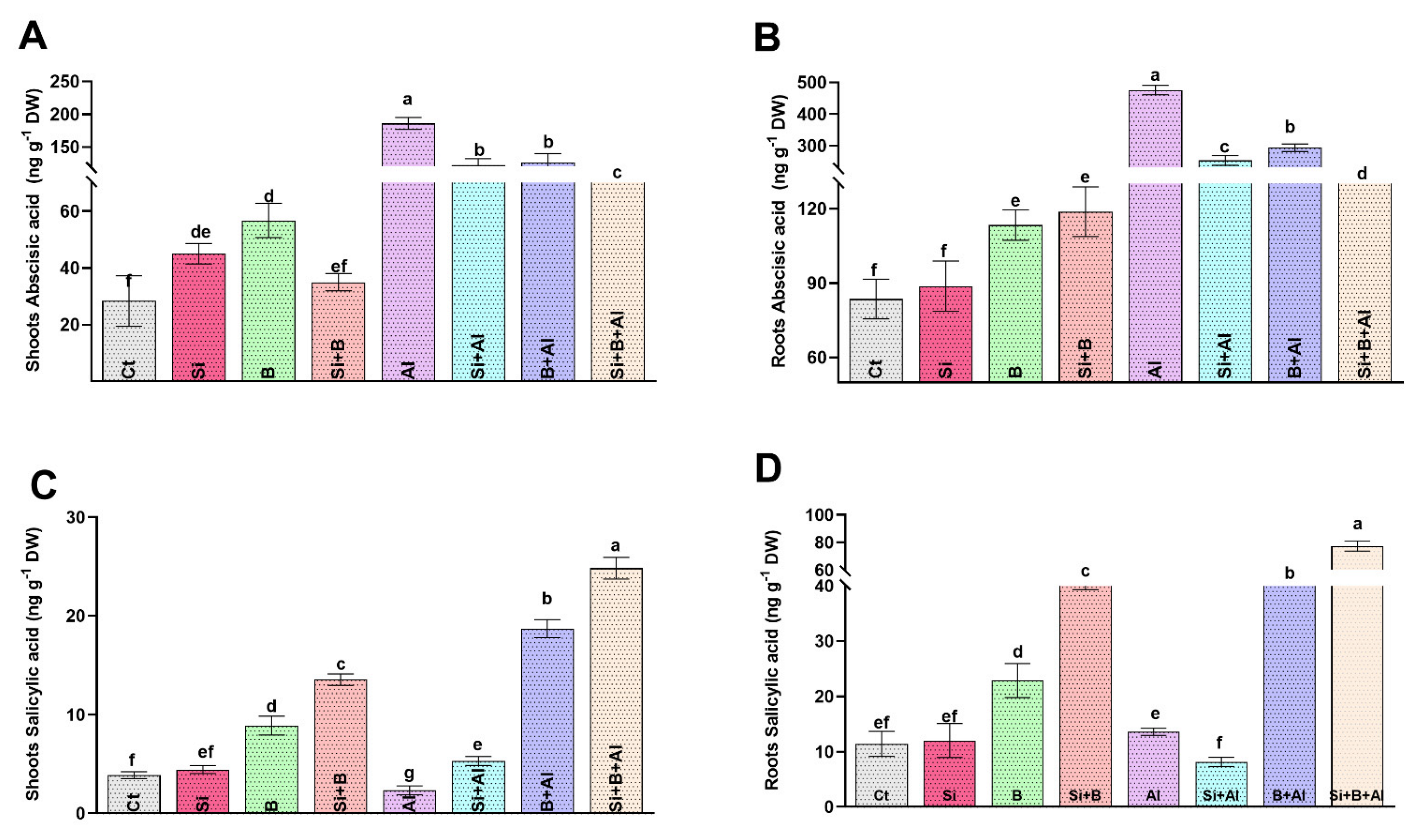
3.5. Effects of Si, B, and Si + B on the Endogenous ABA and SA Levels in Date Palm Seedlings under Normal and Al3+ Stress Conditions
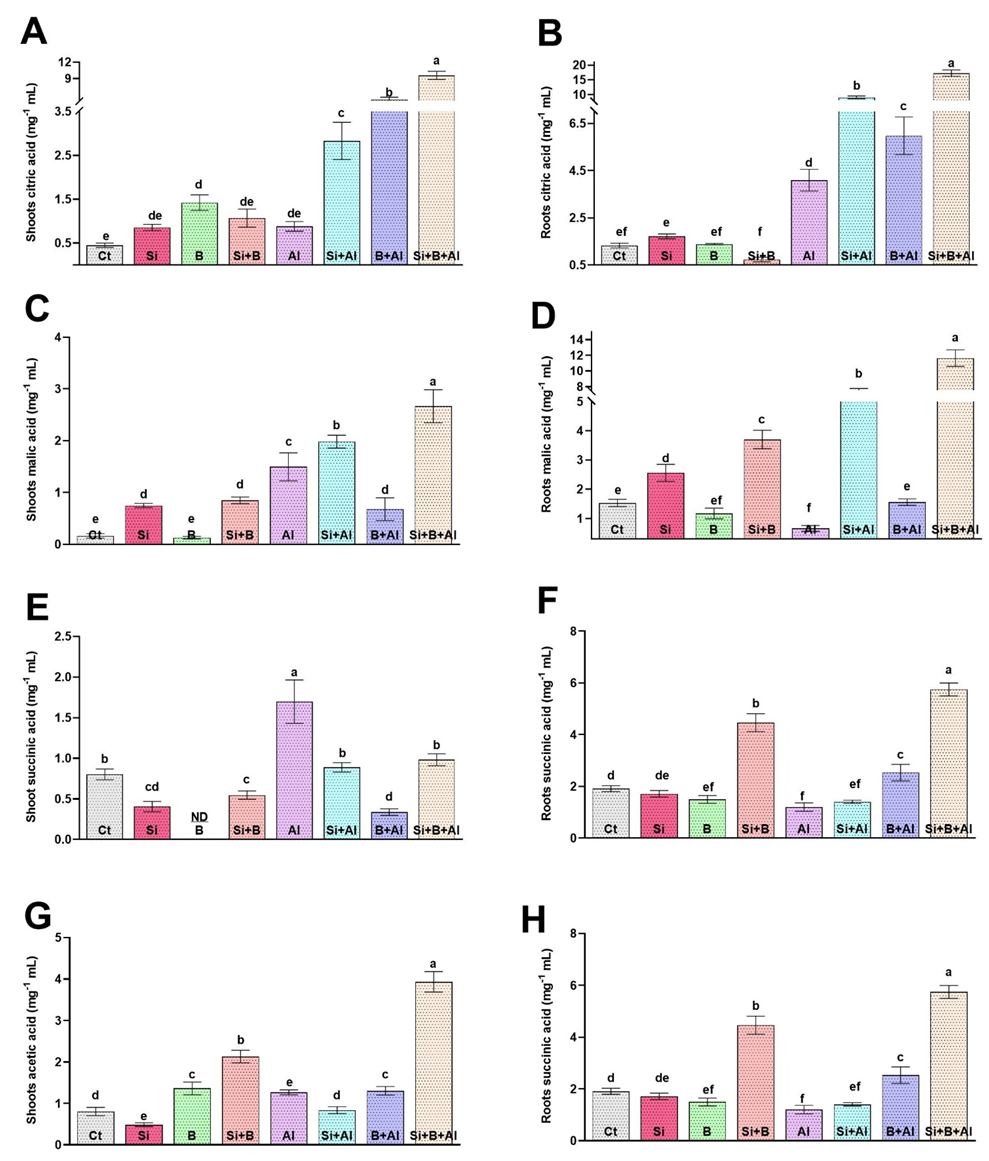
3.6. Effect of Exogenous Si and B on the Regulation of Citric Acid, Malic Acid, Succinic Acid, and Acetic Acid under Al3+ Stress Conditions
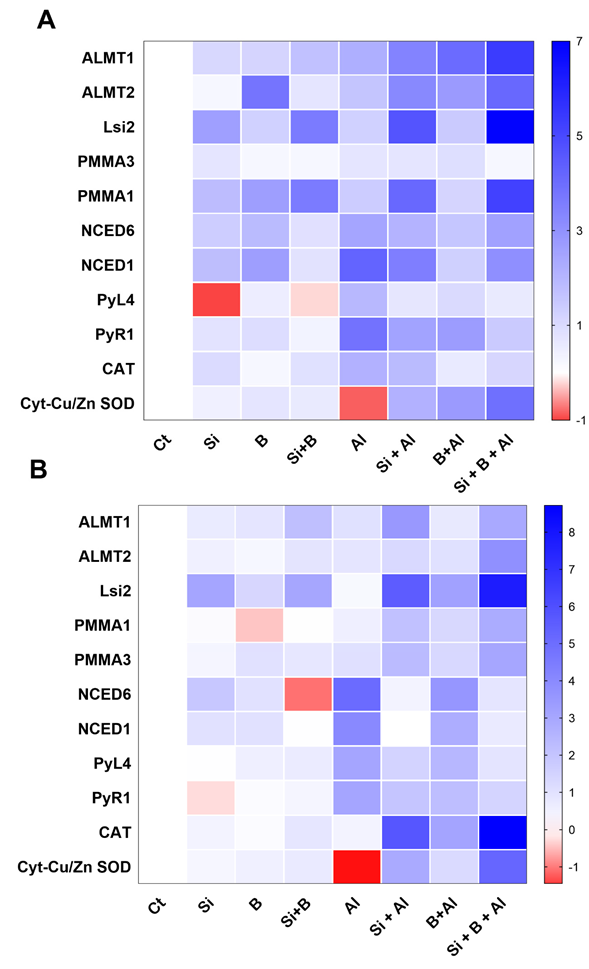
3.7. Expression of Genes Related to Si and Al3+ Transport, ATPase H+ Pump, and Biosynthesis of ABA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, S.; Ren, X.; Huang, B.; Wang, G.; Zhou, P.; An, Y. Aluminium-induced reduction of plant growth in alfalfa (Medicago sativa) is mediated by interrupting auxin transport and accumulation in roots. Sci. Rep. 2016, 6, 30079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, S.K.; Baluška, F.; Matsumoto, H. Aluminum stress signaling in plants. Plant Signal. Behav. 2009, 4, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miotto, A.; Tiecher, T.; Kaminski, J.; Brunetto, G.; De Conti, L.; Tiecher, T.L.; Martins, A.P.; Rheinheimer dos Santos, D. Soil acidity and aluminum speciation affected by liming in the conversion of a natural pasture from the Brazilian Campos Biome into no-tillage system for grain production. Arch. Agron. Soil Sci. 2020, 66, 138–151. [Google Scholar] [CrossRef]
- Alarcón-Poblete, E.; Inostroza-Blancheteau, C.; Alberdi, M.; Rengel, Z.; Reyes-Díaz, M. Molecular regulation of aluminum resistance and sulfur nutrition during root growth. Planta 2018, 247, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Y.; Mai, J.; Tao, L.; Qu, M.; Liu, J.; Shen, R.; Xu, G.; Feng, Y.; Xiao, H. Boron alleviates aluminum toxicity by promoting root alkalization in transition zone via polar auxin transport. Plant Physiol. 2018, 177, 1254–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribera-Fonseca, A.; Rumpel, C.; de la Luz Mora, M.; Nikolic, M.; Cartes, P. Sodium silicate and calcium silicate differentially affect silicon and aluminium uptake, antioxidant performance and phenolics metabolism of ryegrass in an acid Andisol. Crop Pasture Sci. 2018, 69, 205–215. [Google Scholar] [CrossRef]
- Gupta, N.; Gaurav, S.S.; Kumar, A. Molecular basis of aluminium toxicity in plants: A review. Am. J. Plant Sci. 2013, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Tripathi, D.K.; Singh, S.; Sharma, S.; Dubey, N.K.; Chauhan, D.K.; Vaculík, M. Toxicity of aluminium on various levels of plant cells and organism: A review. Environ. Exp. Bot. 2017, 137, 177–193. [Google Scholar] [CrossRef]
- Pontigo, S.; Godoy, K.; Jiménez, H.; Gutiérrez-Moraga, A.; Mora, M.D.L.L.; Cartes, P. Silicon-mediated alleviation of aluminum toxicity by modulation of Al/Si uptake and antioxidant performance in ryegrass plants. Front. Plant Sci. 2017, 8, 642. [Google Scholar] [CrossRef] [Green Version]
- Ashenef, A. Essential and toxic metals in tea (Camellia sinensis) imported and produced in Ethiopia. Food Addit. Contam. Part B 2014, 7, 30–36. [Google Scholar] [CrossRef]
- Bojórquez-Quintal, E.; Escalante-Magaña, C.; Echevarría-Machado, I.; Martínez-Estévez, M. Aluminum, a friend or foe of higher plants in acid soils. Front. Plant Sci. 2017, 8, 1767. [Google Scholar] [CrossRef] [PubMed]
- Bilal, S.; Shahzad, R.; Khan, A.L.; Kang, S.-M.; Imran, Q.M.; Al-Harrasi, A.; Yun, B.-W.; Lee, I.-J. Endophytic microbial consortia of phytohormones-producing fungus Paecilomyces formosus LHL10 and bacteria Sphingomonas sp. LK11 to Glycine max L. regulates physio-hormonal changes to attenuate aluminum and zinc stresses. Front. Plant Sci. 2018, 9, 1273. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, N.; Kar, D.; Deepak Mahajan, B.; Nanda, S.; Rahiman, R.; Panchakshari, N.; Bhagavatula, L.; Datta, S. The multitasking abilities of MATE transporters in plants. J. Exp. Bot. 2019, 70, 4643–4656. [Google Scholar] [CrossRef]
- Zhang, J.; Li, D.; Wei, J.; Ma, W.; Kong, X.; Rengel, Z.; Chen, Q. Melatonin alleviates aluminum-induced root growth inhibition by interfering with nitric oxide production in Arabidopsis. Environ. Exp. Bot. 2019, 161, 157–165. [Google Scholar] [CrossRef]
- Jones, D.; Blancaflor, E.; Kochian, L.; Gilroy, S. Spatial coordination of aluminium uptake, production of reactive oxygen species, callose production and wall rigidification in maize roots. Plant Cell Environ. 2006, 29, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Borsani, O.; Valpuesta, V.; Botella, M.A. Evidence for a Role of Salicylic Acid in the Oxidative Damage Generated by NaCl and Osmotic Stress in Arabidopsis Seedlings. Plant Physiol. 2001, 126, 1024–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartes, P.; McManus, M.; Wulff-Zottele, C.; Leung, S.; Gutiérrez-Moraga, A.; de la Luz Mora, M. Differential superoxide dismutase expression in ryegrass cultivars in response to short term aluminium stress. Plant Soil 2012, 350, 353–363. [Google Scholar] [CrossRef]
- Riaz, M.; Yan, L.; Wu, X.; Hussain, S.; Aziz, O.; Wang, Y.; Imran, M.; Jiang, C. Boron alleviates the aluminum toxicity in trifoliate orange by regulating antioxidant defense system and reducing root cell injury. J. Environ. Manag. 2018, 208, 149–158. [Google Scholar] [CrossRef]
- Ribeiro, M.A.Q.; Almeida, A.-A.F.d.; Mielke, M.S.; Gomes, F.P.; Pires, M.V.; Baligar, V.C. Aluminum effects on growth, photosynthesis, and mineral nutrition of cacao genotypes. J. Plant Nutr. 2013, 36, 1161–1179. [Google Scholar] [CrossRef]
- Montpetit, J.; Vivancos, J.; Mitani-Ueno, N.; Yamaji, N.; Rémus-Borel, W.; Belzile, F.; Ma, J.F.; Bélanger, R.R. Cloning, functional characterization and heterologous expression of TaLsi1, a wheat silicon transporter gene. Plant Mol. Biol. 2012, 79, 35–46. [Google Scholar] [CrossRef]
- Shahnaz, G.; Shekoofeh, E.; Kourosh, D.; Moohamadbagher, B. Interactive effects of silicon and aluminum on the malondialdehyde (MDA), proline, protein and phenolic compounds in Borago officinalis L. J. Med. Plants Res. 2011, 5, 5818–5827. [Google Scholar]
- Tripathi, D.K.; Singh, V.P.; Ahmad, P.; Chauhan, D.K.; Prasad, S.M. Silicon in Plants: Advances and Future Prospects; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Kopittke, P.M.; Gianoncelli, A.; Kourousias, G.; Green, K.; McKenna, B.A. Alleviation of Al Toxicity by Si Is Associated with the Formation of Al–Si Complexes in Root Tissues of Sorghum. Front. Plant Sci. 2017, 8, 2189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirisuntornlak, N.; Ullah, H.; Sonjaroon, W.; Anusontpornperm, S.; Arirob, W.; Datta, A. Interactive effects of silicon and soil pH on growth, yield and nutrient uptake of maize. Silicon 2021, 13, 289–299. [Google Scholar] [CrossRef]
- Pavlovic, J.; Kostic, L.; Bosnic, P.; Kirkby, E.A.; Nikolic, M. Interactions of silicon with essential and beneficial elements in plants. Front. Plant Sci. 2021, 12, 1224. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-H.; Khan, A.L.; Lee, I.-J. Silicon: A duo synergy for regulating crop growth and hormonal signaling under abiotic stress conditions. Crit. Rev. Biotechnol. 2016, 36, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-H.; Khan, A.L.; Waqas, M.; Lee, I.-J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Bashri, G.; Shweta, S.; Ahmad, P.; Singh, V. Efficacy of silicon against aluminum toxicity in plants: An overview. Silicon Plants: Adv. Future Prospect. 2017, 1, 355–366. [Google Scholar]
- Zhou, X.-X.; Yang, L.-T.; Qi, Y.-P.; Guo, P.; Chen, L.-S. Mechanisms on boron-induced alleviation of aluminum-toxicity in Citrus grandis seedlings at a transcriptional level revealed by cDNA-AFLP analysis. PLoS ONE 2015, 10, e0115485. [Google Scholar] [CrossRef] [Green Version]
- Li, X.W.; Liu, J.Y.; Fang, J.; Tao, L.; Shen, R.F.; Li, Y.L.; Xiao, H.D.; Feng, Y.M.; Wen, H.X.; Guan, J.H. Boron supply enhances aluminum tolerance in root border cells of pea (Pisum sativum) by interacting with cell wall pectins. Front. Plant Sci. 2017, 8, 742. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.; Yan, L.; Wu, X.; Hussain, S.; Aziz, O.; Imran, M.; Rana, M.S.; Jiang, C. Boron reduces aluminum-induced growth inhibition, oxidative damage and alterations in the cell wall components in the roots of trifoliate orange. Ecotoxicol. Environ. Saf. 2018, 153, 107–115. [Google Scholar] [CrossRef]
- Riaz, M.; Yan, L.; Wu, X.; Hussain, S.; Aziz, O.; Jiang, C. Boron deprivation induced inhibition of root elongation is provoked by oxidative damage, root injuries and changes in cell wall structure. Environ. Exp. Bot. 2018, 156, 74–85. [Google Scholar] [CrossRef]
- Riaz, M.; Yan, L.; Wu, X.; Hussain, S.; Aziz, O.; Jiang, C. Boron increases root elongation by reducing aluminum induced disorganized distribution of HG epitopes and alterations in subcellular cell wall structure of trifoliate orange roots. Ecotoxicol. Environ. Saf. 2018, 165, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Riaz, M.; Liu, Y.; Zeng, Y.; Jiang, C. Aluminum toxicity could be mitigated with boron by altering the metabolic patterns of amino acids and carbohydrates rather than organic acids in trifoliate orange. Tree Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Q.; Cao, X.C.; Zhu, L.F.; Hu, W.J.; Hu, A.Y.; Abliz, B.; Bai, Z.G.; Huang, J.; Liang, Q.D.; Sajid, H. Boron reduces cell wall aluminum content in rice (Oryza sativa) roots by decreasing H2O2 accumulation. Plant Physiol. Biochem. 2019, 138, 80–90. [Google Scholar] [CrossRef]
- Al-Yahyai, R.; Khan, M.M. Date palm status and perspective in Oman. In Date Palm Genetic Resources and Utilization; Springer: Berlin/Heidelberg, Germany, 2015; pp. 207–240. [Google Scholar]
- Yaish, M.W.; Kumar, P.P. Salt tolerance research in date palm tree (Phoenix dactylifera L.), past, present, and future perspectives. Front. Plant Sci. 2015, 6, 348. [Google Scholar] [CrossRef] [Green Version]
- Alansi, S.; Al-Qurainy, F.; Nadeem, M.; Khan, S.; Tarroum, M.; Alshameri, A.; Gaafar, A.-R.Z. Cryopreservation: A tool to conserve date palm in Saudi Arabia. Saudi J. Biol. Sci. 2019, 26, 1896–1902. [Google Scholar] [CrossRef]
- Chaâbene, Z.; Rorat, A.; Hakim, I.R.; Bernard, F.; Douglas, G.C.; Elleuch, A.; Vandenbulcke, F.; Mejdoub, H. Insight into the expression variation of metal-responsive genes in the seedling of date palm (Phoenix dactylifera). Chemosphere 2018, 197, 123–134. [Google Scholar] [CrossRef]
- Awad, K.M.; Salih, A.M.; Khalaf, Y.; Suhim, A.A.; Abass, M.H. Phytotoxic and genotoxic effect of Aluminum to date palm (Phoenix dactylifera L.) in vitro cultures. J. Genet. Eng. Biotechnol. 2019, 17, 1–8. [Google Scholar] [CrossRef]
- Khan, A.; Bilal, S.; Khan, A.L.; Imran, M.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.-J. Silicon-mediated alleviation of combined salinity and cadmium stress in date palm (Phoenix dactylifera L.) by regulating physio-hormonal alteration. Ecotoxicol. Environ. Saf. 2020, 188, 109885. [Google Scholar] [CrossRef]
- Khan, A.; Kamran, M.; Imran, M.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Amri, I.; Lee, I.-J.; Khan, A.L. Silicon and salicylic acid confer high-pH stress tolerance in tomato seedlings. Sci. Rep. 2019, 9, 1–16. [Google Scholar]
- Chen, D.; Chen, D.; Xue, R.; Long, J.; Lin, X.; Lin, Y.; Jia, L.; Zeng, R.; Song, Y. Effects of boron, silicon and their interactions on cadmium accumulation and toxicity in rice plants. J. Hazard. Mater. 2019, 367, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Sumanta, N.; Haque, C.I.; Nishika, J.; Suprakash, R. Spectrophotometric analysis of chlorophylls and carotenoids from commonly grown fern species by using various extracting solvents. Res. J. Chem. Sci. ISSN 2014, 2231, 606X. [Google Scholar]
- Bilal, S.; Shahzad, R.; Khan, A.L.; Al-Harrasi, A.; Kim, C.K.; Lee, I.-J. Phytohormones enabled endophytic Penicillium funiculosum LHL06 protects Glycine max L. from synergistic toxicity of heavy metals by hormonal and stress-responsive proteins modulation. J. Hazard. Mater. 2019, 379, 120824. [Google Scholar] [CrossRef]
- Khan, A.; Bilal, S.; Khan, A.L.; Imran, M.; Shahzad, R.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Azhri, M.; Mohanta, T.K.; Lee, I.-J. Silicon and gibberellins: Synergistic function in harnessing ABA signaling and heat stress tolerance in date palm (Phoenix dactylifera L.). Plants 2020, 9, 620. [Google Scholar] [CrossRef] [PubMed]
- Gajewska, E.; Skłodowska, M. Differential biochemical responses of wheat shoots and roots to nickel stress: Antioxidative reactions and proline accumulation. Plant Growth Regul. 2008, 54, 179–188. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Ali, A.; Bilal, S.; Khan, A.L.; Mabood, F.; Al-Harrasi, A.; Lee, I.-J. Endophytic Aureobasidium pullulans BSS6 assisted developments in phytoremediation potentials of Cucumis sativus under Cd and Pb stress. J. Plant Interact. 2019, 14, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Bilal, S.; Khan, A.L.; Waqas, M.; Shahzad, R.; Kim, I.-D.; Lee, I.-J.; Shin, D.-H. Biochemical constituents and in vitro antioxidant and anticholinesterase potential of seeds from native Korean persimmon genotypes. Molecules 2016, 21, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahzad, R.; Khan, A.L.; Waqas, M.; Ullah, I.; Bilal, S.; Kim, Y.-H.; Asaf, S.; Kang, S.-M.; Lee, I.-J. Metabolic and proteomic alteration in phytohormone-producing endophytic Bacillus amyloliquefaciens RWL-1 during methanol utilization. Metabolomics 2019, 15, 16. [Google Scholar] [CrossRef]
- Seskar, M.; Shulaev, V.; Raskin, I. Endogenous methyl salicylate in pathogen-inoculated tobacco plants. Plant Physiol. 1998, 116, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Bilal, S.; Khan, A.L.; Shahzad, R.; Kim, Y.-H.; Imran, M.; Khan, M.J.; Al-Harrasi, A.; Kim, T.H.; Lee, I.-J. Mechanisms of Cr (VI) resistance by endophytic Sphingomonas sp. LK11 and its Cr (VI) phytotoxic mitigating effects in soybean (Glycine max L.). Ecotoxicol. Environ. Saf. 2018, 164, 648–658. [Google Scholar] [CrossRef]
- Shetty, R.; Vidya, C.S.-N.; Prakash, N.B.; Lux, A.; Vaculík, M. Aluminum toxicity in plants and its possible mitigation in acid soils by biochar: A review. Sci. Total Environ. 2021, 765, 142744. [Google Scholar] [CrossRef] [PubMed]
- Hodson, M.J.; Evans, D.E. Aluminium–silicon interactions in higher plants: An update. J. Exp. Bot. 2020, 71, 6719–6729. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Awan, S.A.; Rizwan, M.; Ali, S.; Hassan, M.J.; Brestic, M.; Zhang, X.; Huang, L. Effects of silicon on heavy metal uptake at the soil-plant interphase: A review. Ecotoxicol. Environ. Saf. 2021, 222, 112510. [Google Scholar] [CrossRef] [PubMed]
- Shehzad, M.A.; Maqsood, M.; Nawaz, F.; Abbas, T.; Yasin, S. Boron-induced improvement in physiological, biochemical and growth attributes in sunflower (Helianthus annuus L.) exposed to terminal drought stress. J. Plant Nutr. 2018, 41, 943–955. [Google Scholar] [CrossRef]
- Abdel-Motagally, F.; El-Zohri, M. Improvement of wheat yield grown under drought stress by boron foliar application at different growth stages. J. Saudi Soc. Agric. Sci. 2018, 17, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Azeem, M.; Shoujun, Y.; Qasim, M.; Abbasi, M.W.; Ahmed, N.; Hanif, T.; Adnan, M.Y.; Ahmad, R.; Dong, R. Foliar enrichment of potassium and boron overcomes salinity barriers to improve growth and yield potential of cotton (Gossypium hirsutum L.). J. Plant Nutr. 2021, 44, 438–454. [Google Scholar] [CrossRef]
- Chakraborty, S.; Mishra, A.; Verma, E.; Tiwari, B.; Mishra, A.K.; Singh, S.S. Physiological mechanisms of aluminum (Al) toxicity tolerance in nitrogen-fixing aquatic macrophyte Azolla microphylla Kaulf: Phytoremediation, metabolic rearrangements, and antioxidative enzyme responses. Environ. Sci. Pollut. Res. 2019, 26, 9041–9054. [Google Scholar] [CrossRef]
- Silva, S.; Pinto, G.; Dias, M.C.; Correia, C.M.; Moutinho-Pereira, J.; Pinto-Carnide, O.; Santos, C. Aluminium long-term stress differently affects photosynthesis in rye genotypes. Plant Physiol. Biochem. 2012, 54, 105–112. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Xie, Y.; Sangari, S. Silicon mechanisms to ameliorate heavy metal stress in plants. BioMed Res. Int. 2018, 2018, 8492898. [Google Scholar] [CrossRef] [Green Version]
- Ikegawa, H.; Yamamoto, Y.; Matsumoto, H. Responses to aluminium of suspension-cultured tobacco cells in a simple calcium solution. Soil Sci. Plant Nutr. 2000, 46, 503–514. [Google Scholar]
- Chauhan, D.K.; Yadav, V.; Vaculík, M.; Gassmann, W.; Pike, S.; Arif, N.; Singh, V.P.; Deshmukh, R.; Sahi, S.; Tripathi, D.K. Aluminum toxicity and aluminum stress-induced physiological tolerance responses in higher plants. Crit. Rev. Biotechnol. 2021, 41, 715–730. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-L.; Fan, W.; Zheng, S.-J. Mechanisms and regulation of aluminum-induced secretion of organic acid anions from plant roots. J. Zhejiang Univ.-Sci. B 2019, 20, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Long, Y.; Huang, J.; Xia, J. Molecular mechanisms for coping with Al toxicity in plants. Int. J. Mol. Sci. 2019, 20, 1551. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Ryan, P.R.; Delhaize, E. Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 2001, 6, 273–278. [Google Scholar] [CrossRef]
- Dawood, M.; Cao, F.; Jahangir, M.M.; Zhang, G.; Wu, F. Alleviation of aluminum toxicity by hydrogen sulfide is related to elevated ATPase, and suppressed aluminum uptake and oxidative stress in barley. J. Hazard. Mater. 2012, 209, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.A.; Hippler, F.W.; Prado, L.A.d.S.; Rima, J.A.; Boaretto, R.M.; Quaggio, J.A.; Façanha, A.R.; Mattos-Jr, D. Boron modulates the plasma membrane H+-ATPase activity affecting nutrient uptake of Citrus trees. Ann. Appl. Biol. 2021, 178, 293–303. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Lee, I.-J. Synergistic interaction of fungal endophytes, Paecilomyces formosus LHL10 and Penicillium funiculosum LHL06, in alleviating multi-metal toxicity stress in Glycine max L. Environ. Sci. Pollut. Res. 2021, 28, 67429–67444. [Google Scholar] [CrossRef]
- Xu, J.; Yang, J.; Duan, X.; Jiang, Y.; Zhang, P. Increased expression of native cytosolic Cu/Zn superoxide dismutase and ascorbate peroxidase improves tolerance to oxidative and chilling stresses in cassava (Manihot esculenta Crantz). BMC Plant Biol. 2014, 14, 1–14. [Google Scholar] [CrossRef]
- Tahjib-Ul-Arif, M.; Zahan, M.; Karim, M.; Imran, S.; Hunter, C.T.; Islam, M.; Mia, M.; Hannan, M.; Rhaman, M.S.; Hossain, M. Citric acid-mediated abiotic stress tolerance in plants. Int. J. Mol. Sci. 2021, 22, 7235. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, J.; Liu, X.; Chen, X.; Liu, L.; Niu, Y.; Wang, R. The relief effects of organic acids on Scirpus triqueter L. under pyrene–lead stress. Environ. Sci. Pollut. Res. 2019, 26, 15828–15837. [Google Scholar] [CrossRef]
- Shahzad, R.; Bilal, S.; Imran, M.; Khan, A.L.; Alosaimi, A.A.; Al-Shwyeh, H.A.; Almahasheer, H.; Rehman, S.; Lee, I.-J. Amelioration of heavy metal stress by endophytic Bacillus amyloliquefaciens RWL-1 in rice by regulating metabolic changes: Potential for bacterial bioremediation. Biochem. J. 2019, 476, 3385–3400. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, A.L.; Imran, M.; Asaf, S.; Kim, Y.-H.; Bilal, S.; Numan, M.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.-J. Silicon-induced thermotolerance in Solanum lycopersicum L. via activation of antioxidant system, heat shock proteins, and endogenous phytohormones. BMC Plant Biol. 2020, 20, 248. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Khan, A.L.; Muneer, S.; Kim, Y.-H.; Al-Rawahi, A.; Al-Harrasi, A. Silicon and salinity: Crosstalk in crop-mediated stress tolerance mechanisms. Front. Plant Sci. 2019, 10, 1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjan, A.; Sinha, R.; Sharma, T.R.; Pattanayak, A.; Singh, A.K. Alleviating aluminum toxicity in plants: Implications of reactive oxygen species signaling and crosstalk with other signaling pathways. Physiol. Plant. 2021, 173, 1765–1784. [Google Scholar] [CrossRef]
- Shukla, P.S.; Shotton, K.; Norman, E.; Neily, W.; Critchley, A.T.; Prithiviraj, B. Seaweed extract improve drought tolerance of soybean by regulating stress-response genes. AoB Plants 2018, 10, plx051. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Imran, M.; Jan, R.; Kim, K.M.; Lee, I.-J. Synergistic association of endophytic fungi enhances Glycine max L. resilience to combined abiotic stresses: Heavy metals, high temperature and drought stress. Ind. Crops Prod. 2020, 143, 111931. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Vishwakarma, K.; Singh, V.P.; Prakash, V.; Sharma, S.; Muneer, S.; Nikolic, M.; Deshmukh, R.; Vaculík, M.; Corpas, F.J. Silicon crosstalk with reactive oxygen species, phytohormones and other signaling molecules. J. Hazard. Mater. 2020, 408, 124820. [Google Scholar] [CrossRef]
- Jogawat, A.; Yadav, B.; Lakra, N.; Singh, A.K.; Narayan, O.P. Crosstalk between phytohormones and secondary metabolites in the drought stress tolerance of crop plants: A review. Physiol. Plant. 2021, 172, 1106–1132. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Ashfaque, F.; Chhillar, H.; Irfan, M.; Khan, N.A. The intricacy of silicon, plant growth regulators and other signaling molecules for abiotic stress tolerance: An entrancing crosstalk between stress alleviators. Plant Physiol. Biochem. 2021, 162, 36–47. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Khan, A.L.; Kim, D.-H.; Lee, S.-Y.; Kim, K.-M.; Waqas, M.; Jung, H.-Y.; Shin, J.-H.; Kim, J.-G.; Lee, I.-J. Silicon mitigates heavy metal stress by regulating P-type heavy metal ATPases, Oryza sativa low silicon genes, and endogenous phytohormones. BMC Plant Biol. 2014, 14, 13. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.-M.; Wang, J.; Wang, S.-H.; Xu, L.-L. Salicylic acid-induced aluminum tolerance by modulation of citrate efflux from roots of Cassia tora L. Planta 2003, 217, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Yıldırım, K. Transcriptomic and hormonal control of boron uptake, accumulation and toxicity tolerance in poplar. Environ. Exp. Bot. 2017, 141, 60–73. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Shoot Length (cm) | Root Length (cm) | Shoot Dry Weight (mg) | Root dry Weight (mg) | Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Carot (mg g−1 FW) |
|---|---|---|---|---|---|---|---|
| Ct | 20.0 ± 1.51 e | 6.6 ± 0.69 b | 1371.7 ± 71.69 d | 1144.0 ±108.75 b | 61.6 ± 5.8 a | 113.6 ± 11.4 e | 109.5 ± 8.2 f |
| Si | 28.0 ± 1.15 b | 8.4 ± 0.69 a | 2090.8 ±107.78 b | 1390.6 ± 104.47 a | 65.1 ± 4.6 a | 174.4 ± 12.6 b | 151.2 ± 6.6 d |
| B | 26.6 ± 1.03 c | 8.7 ± 1.05 a | 1936.6 ±103.94 b | 1309.6 ± 55.23 a | 66.8 ± 1.6 a | 183.1 ± 11.9 b | 168.2 ± 6.8 c |
| Si + B | 31.1 ± 1.75 a | 9.6 ± 0.84 a | 2442.2 ±116.21 a | 1438.8 ± 126.56 a | 65.5 ± 6.3 a | 216.0 ± 18.1 a | 191.0 ± 6.1 b |
| Al3+ | 15.4 ± 1.95 f | 4.9 ± 1.19 c | 980.0 ± 72.94 e | 784.00 ± 62.44 c | 33.1 ± 5.2 b | 57.2 ± 10.4 f | 83.6 ± 6.5 g |
| Si + Al3+ | 24.1 ± 1.07 cd | 7.7 ± 1.15 a | 1597.8 ±123.05 c | 1094.8 ± 48.59 b | 62.0 ± 2.2 a | 149.8 ± 14.3 cd | 139.0 ± 5.8 e |
| B + Al3+ | 22.5 ± 1.14 d | 8.4 ± 1.26 a | 1627.0 ±105.03 c | 968.20 ± 34.13 b | 60.0 ± 5.8 a | 137.0 ± 6.97 d | 155.8 ± 4.9 d |
| Si + B + Al3+ | 26.0 ± 1.53 bc | 9.2 ± 1.13 a | 2003.6 ± 110.59 b | 1086.2 ± 124.86 b | 59.5 ± 11.0 a | 156.6 ± 11.9 cb | 204.6 ± 8.7 a |
| Treatment | B µg g−1 | Si µg g−1 | Al µg g−1 | |||
|---|---|---|---|---|---|---|
| Leaves | Root | Leaves | Root | Leaves | Root | |
| Ct | 45.04 ± 3.64 e | 28.63 ± 3.83 cd | 381.89 ± 10.31 e | 306.02 ± 11.3 f | 18.44 ± 2.64 c | 127.52 ± 7.1 ef |
| Si | 41.23 ± 4.30 ef | 25.06 ± 1.81 d | 673.08 ± 21.32 a | 509.36 ± 21.2 a | 22.91 ± 2.52 c | 114.49 ± 6.23 fg |
| B | 75.57 ± 4.57 b | 50.36 ± 2.96 a | 433.53 ± 18.12 d | 296.52 ± 14.5 f | 42.38 ± 3.29 c | 98.32 ± 6.62 g |
| Si + B | 36.25 ± 3.14 f | 39.27 ± 3.04 b | 513.36 ± 25.34 c | 456.72 ± 17.2 b | 31.45 ± 2.72 b | 144.89 ± 8.33 e |
| Al | 47.94 ± 3.04 e | 23.64 ± 2.71 d | 346.57 ± 14.55 f | 229.91 ± 21.7 g | 116.86 ± 8.31 a | 541.29 ± 29.1 a |
| Si + Al | 58.61 ± 3.51 d | 32.91 ± 3.31 c | 572.89 ± 22.32 b | 424.52 ± 12.5 c | 68.23 ± 3.556 b | 323.74 ± 23.02 b |
| B + Al | 89.23 ± 5.06 a | 49.43 ± 3.79 a | 403.01 ± 19.33 e | 331.97 ± 11.4 e | 83.95 ± 4.63 b | 276.71 ± 16.74 c |
| Si + B + Al | 68.16 ± 4.63 c | 49.95 ± 5.32 a | 522.90 ± 16.52 c | 379.67 ± 18.1 d | 50.60 ± 3.12 b | 218.05 ± 11.72 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilal, S.; Khan, A.; Imran, M.; Khan, A.L.; Asaf, S.; Al-Rawahi, A.; Al-Azri, M.S.A.; Al-Harrasi, A.; Lee, I.-J. Silicon- and Boron-Induced Physio-Biochemical Alteration and Organic Acid Regulation Mitigates Aluminum Phytotoxicity in Date Palm Seedlings. Antioxidants 2022, 11, 1063. https://doi.org/10.3390/antiox11061063
Bilal S, Khan A, Imran M, Khan AL, Asaf S, Al-Rawahi A, Al-Azri MSA, Al-Harrasi A, Lee I-J. Silicon- and Boron-Induced Physio-Biochemical Alteration and Organic Acid Regulation Mitigates Aluminum Phytotoxicity in Date Palm Seedlings. Antioxidants. 2022; 11(6):1063. https://doi.org/10.3390/antiox11061063
Chicago/Turabian StyleBilal, Saqib, Adil Khan, Muhammad Imran, Abdul Latif Khan, Sajjad Asaf, Ahmed Al-Rawahi, Masoud Sulaiman Abood Al-Azri, Ahmed Al-Harrasi, and In-Jung Lee. 2022. "Silicon- and Boron-Induced Physio-Biochemical Alteration and Organic Acid Regulation Mitigates Aluminum Phytotoxicity in Date Palm Seedlings" Antioxidants 11, no. 6: 1063. https://doi.org/10.3390/antiox11061063