
A Computational Study of the Role of Secondary Metabolites for Mitigation of Acid Soil Stress in Cereals Using Dehydroascorbate and Mono-Dehydroascorbate Reductases

 ,
,  ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Target Enzymes
2.2. Ligands (Metabolites and Ions)
2.3. Molecular Docking
2.4. Visualization of the Interactions
2.5. Statistical Analysis
3. Results
3.1. Docking Scores against DHAR
3.2. Docking Scores against MDHAR
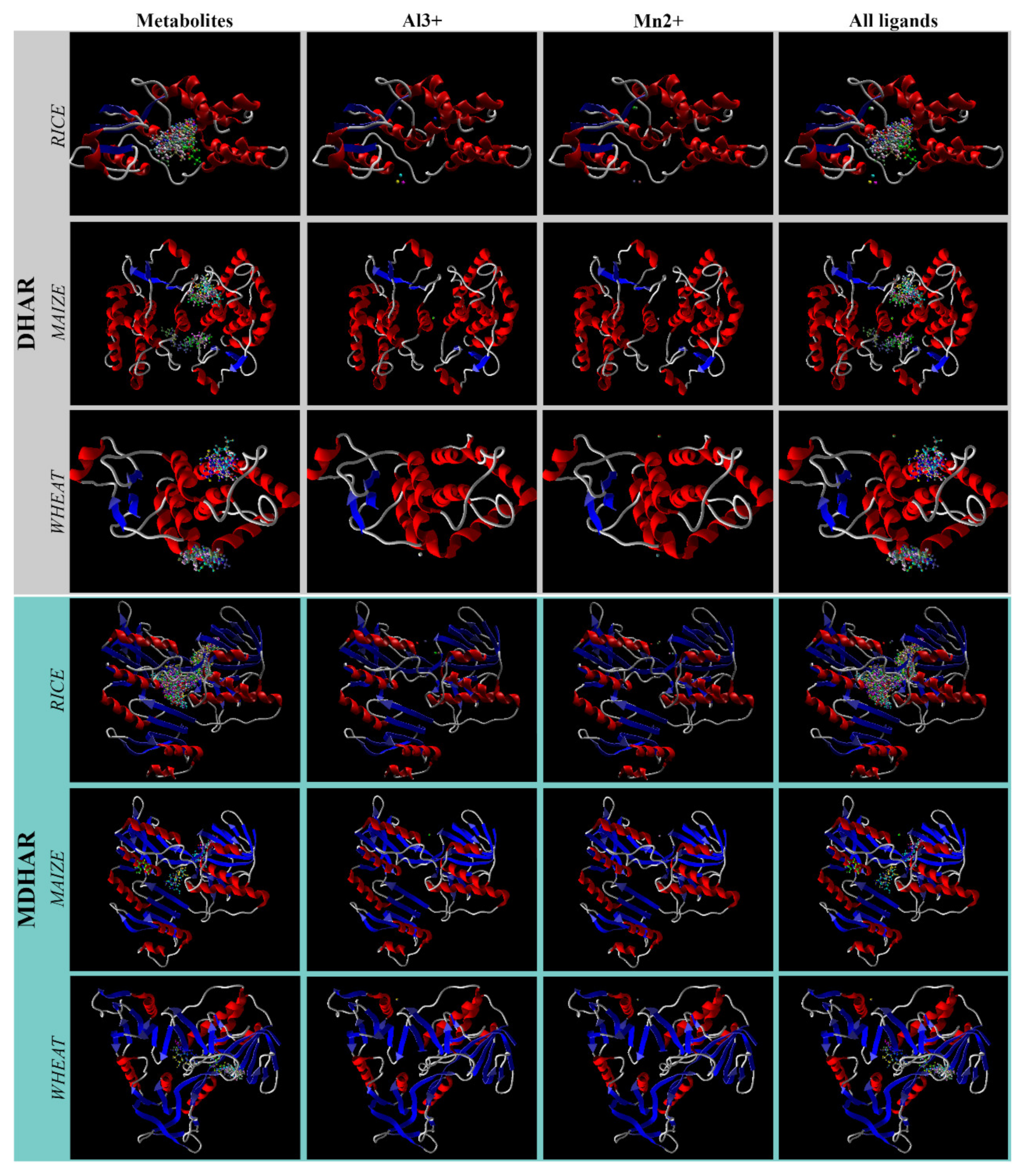
3.3. Docking Orientation and Overlapping Surfaces
3.4. Energy Map
3.5. Ligand Properties Affecting the Interactions
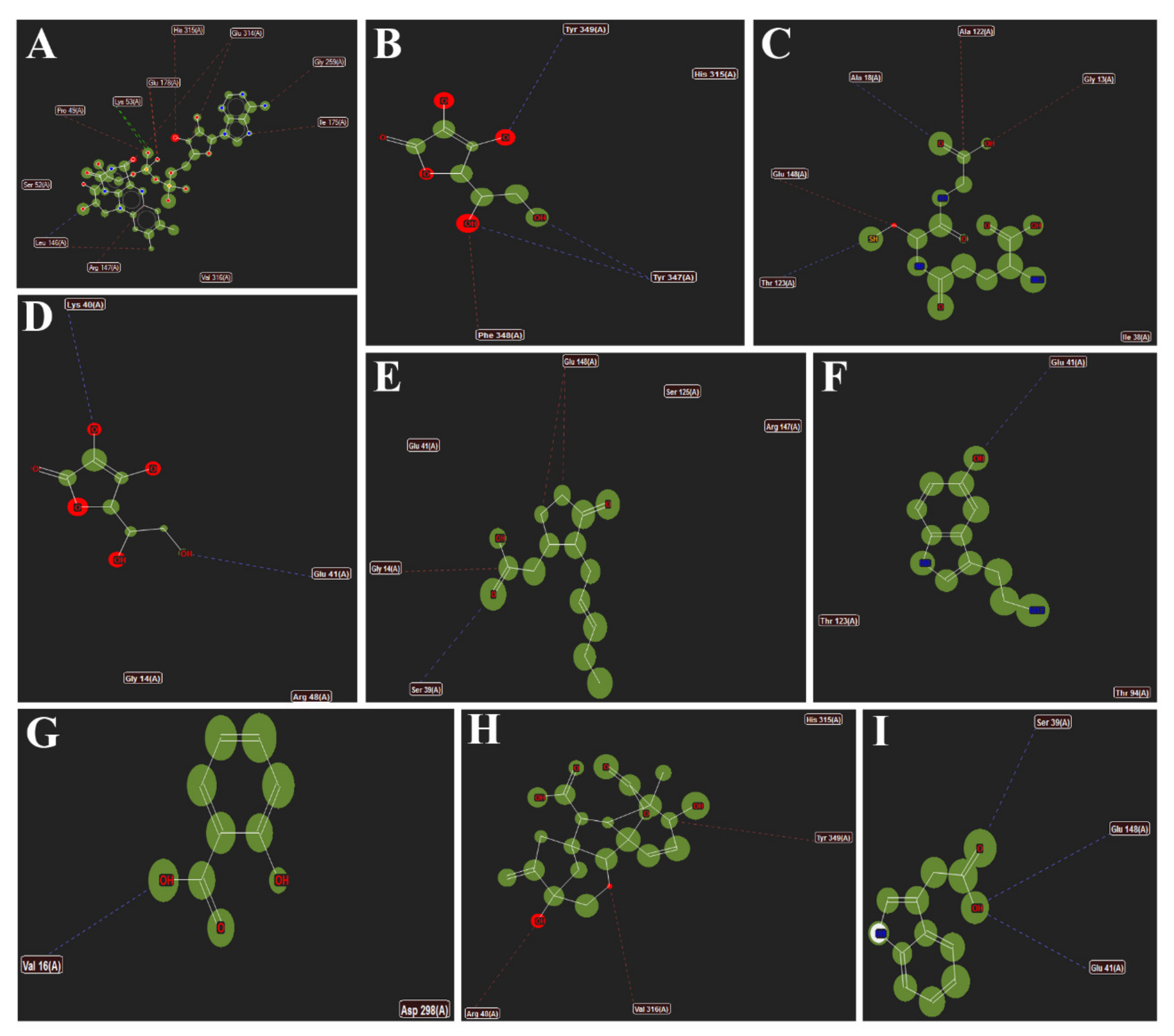
3.6. Interactions of Different Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foy, C.D. Physiological effects of hydrogen, Al and manganese toxicities in acid soil. In Soil Acidity and Liming; Pearson, R.W., Adams, F., Eds.; American Society of Agronomy: Madison, WI, USA, 1984; pp. 57–97. [Google Scholar]
- Vaughan, D.; Ord, B.G. Influence of phenolics acids on the sodium, calcium and chloride contents of Pisum sativum under axenic conditions. Soil Biol. Biochem. 1991, 23, 1191–1193. [Google Scholar] [CrossRef]
- Hafner, H.; Ndunguru, B.J.; Bationo, A.; Marschner, H. Effect of nitrogen, phosphorus and molybdenum application on growth and symbiotic N2-fixation of groundnut in an acid sandy soil in Niger. Fertz. Res. 1992, 31, 69–77. [Google Scholar] [CrossRef]
- Baziramakenga, R.; Leroux, G.D.; Simard, R.R. Effects of benzoic and cinnamic acids on membrane permeability of soybean roots. J. Chem. Ecol. 1995, 21, 1271–1285. [Google Scholar] [CrossRef]
- Barceló, J.; Poschenrieder, C.; Vázquez, M.D.; Gunsé, B. Aluminum phytotoxicity: A challenge for plant scientists. Fertz. Res. 1996, 43, 217–223. [Google Scholar] [CrossRef]
- Kidd, P.S.; Proctor, J. Why plant grows poorly on very acid soil: Are ecologists missing the obvious? J. Exp. Bot. 2001, 52, 791–799. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Yamaji, N.; Ma, J.F. Plasma membrane-localized transporter for aluminium in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 18381–18385. [Google Scholar] [CrossRef] [Green Version]
- Kochian, L.V.; Piñeros, M.A.; Hoekenga, O.A. The physiology, genetics and molecular biology of plant aluminium resistance and toxicity. Plant Soil 2005, 274, 175–195. [Google Scholar] [CrossRef]
- Delhaize, E.; Craig, S.; Beatson, C.D.; Bennet, R.J.; Jagadish, V.C.; Randall, P.J. Aluminium tolerance in wheat (Triticum aestivum L.) (I. Uptake and distribution of aluminium in root apices). Plant Physiol. 1993, 103, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Ryan, P.R.; Kochian, L.V. Interaction between aluminium toxicity and calcium uptake at root apex in near—Isogenic lines of wheat (Triticum aestivum L.) differing in aluminium tolerance. Plant Physiol. 1993, 102, 975–982. [Google Scholar] [CrossRef]
- Sivaguru, M.; Horst, W.J. The distal part of the transition zone is the most aluminium sensitive apical root zone of maize. Plant Physiol. 1998, 116, 155–163. [Google Scholar] [CrossRef] [Green Version]
- Kochian, L.V. Cellular mechanism of aluminium toxicity and resistance in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Ma, J.F. Syndrome of aluminium toxicity and diversity of aluminum resistance in higher plants. Int. Rev. Cytol. 2007, 264, 225–252. [Google Scholar] [PubMed]
- Ono, K.; Yamamoto, Y.; Hachiya, A.; Matsumoto, H. Synergistic inhibition of growth by aluminium and iron of tobacco (Nicotiana tabacum L.) cells in suspension culture. Plant Cell Physiol. 1995, 36, 115–125. [Google Scholar]
- Yamamoto, Y.; Hachiya, A.; Matsumoto, H. Oxidative damage to membranes by a combination of aluminium and iron in suspension-cultured tobacco cells. Plant Cell Physiol. 1997, 38, 1333–1339. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Kobayashi, Y.; Matsumoto, H. Lipid peroxidation is an early symptom triggered by aluminium, but not the primary cause of elongation inhibition in pea roots. Plant Physiol. 2001, 12, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.; Yamamoto, Y.; Matsumoto, H. Cell death process initiated by combination of aluminium and iron in suspension cultured tobacco cells. Soil Sci. Plant Nutr. 1999, 45, 647–657. [Google Scholar] [CrossRef]
- Kawano, T.; Kadono, T.; Furuichi, T.; Muto, S.; Lapeyrie, F. Aluminium induced distortion in calcium signaling involving oxidative burst and channel regulation in tobacco BT-2 cells. Biochem. Biophys. Res. Commun. 2003, 308, 35–42. [Google Scholar] [CrossRef]
- Exley, C. The pro-oxidant activity of aluminium. Free Radic. Biol. Med. 2004, 236, 380–387. [Google Scholar] [CrossRef]
- Pereira, J.F.; Zhou, G.; Delhaize, E.; Richardson, T.; Zhou, M.; Ryan, P.R. Engineering greater aluminium resistance in wheat by over-expressing TaALMT1. Ann. Bot. 2010, 106, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, H.; Motoda, H. Aluminium toxicity recovery process in root apices. Possible association with oxidative stress. Plant Sci. 2012, 185–186, 1–8. [Google Scholar] [CrossRef]
- Choudhury, S.; Sharma, P. Aluminum stress inhibits root growth and alters physiological and metabolic responses in chickpea (Cicer arietinum L.). Plant Physiol. Biochem. 2014, 85, 63–70. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kobayashi, Y.; Devi, S.R.; Rikiishi, S.; Matsumoto, H. Aluminium toxicity is associated with mitochondrial dysfunction and the production of reactive oxygen species in plant cells. Plant Physiol. 2002, 128, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Moulick, D.; Samanta, S.; Sarkar, S.; Mukherjee, A.; Pattnaik, B.K.; Saha, S.; Awasthi, J.P.; Bhowmick, S.; Ghosh, D.; Samal, A.C.; et al. Arsenic contamination, impact and mitigation strategies in rice agro-environment: An inclusive insight. Sci. Total Environ. 2021, 800, 149477. [Google Scholar] [CrossRef] [PubMed]
- Moulick, D.; Chowardhara, B.; Panda, S.K. Agroecotoxicological aspect of arsenic (As) and cadmium (Cd) on Field crops and its mitigation: Current status and future prospect. In Plant-Metal Interactions; Springer: Cham, Switzerland, 2019; pp. 217–246. [Google Scholar]
- Hossain, A.; Pramanick, B.; Bhutia, K.L.; Ahmad, Z.; Moulick, D.; Maitra, S.; Ahmad, A.; Aftab, T. Emerging roles of osmoprotectant glycine betaine against salt-induced oxidative stress in plants: A major outlook of maize (Zea mays L.). In Frontiers in Plant-Soil Interaction; Academic Press: Cambridge, MA, USA, 2021; pp. 567–587. [Google Scholar]
- Hossain, A.; Ahmad, Z.; Moulick, D.; Maitra, S.; Bhadra, P.; Ahmad, A.; Garai, S.; Mondal, M.; Roy, A.; Sabagh, A.E.; et al. Jasmonates and Salicylates: Mechanisms, Transport and Signalling During Abiotic Stress in Plants. In Jasmonates and Salicylates Signaling in Plants; Springer: Cham, Switzerland, 2021; pp. 1–29. [Google Scholar]
- Mazumder, M.K.; Moulick, D.; Choudhury, S. Iron (Fe 3+)-mediated redox responses and amelioration of oxidative stress in cadmium (Cd 2+) stressed mung bean seedlings: A biochemical and computational analysis. J. Plant Biochem. Biotechnol. 2021, 31, 49–60. [Google Scholar] [CrossRef]
- Mazumder, M.K.; Sharma, P.; Moulick, D.; Tata, S.K.; Choudhury, S. Salicylic acid ameliorates zinc and chromium-induced stress responses in wheat seedlings: A biochemical and computational analysis. Cereal Res. Commun. 2021, 1–12. [Google Scholar] [CrossRef]
- Moulick, D.; Ghosh, D.; Santra, S.C. Evaluation of effectiveness of seed priming with selenium in rice during germination under arsenic stress. Plant Physiol. Biochem. 2016, 109, 571–578. [Google Scholar] [CrossRef]
- Moulick, D.; Santra, S.C.; Ghosh, D. Seed priming with Se alleviate As induced phytotoxicity during germination and seedling growth by restricting As translocation in rice (Oryza sativa L cv IET-4094). Ecotoxicol. Environ. Saf. 2017, 145, 449–456. [Google Scholar] [CrossRef]
- Moulick, D.; Santra, S.C.; Ghosh, D. Seed priming with Se mitigates As-induced phytotoxicity in rice seedlings by enhancing essential micronutrient uptake and translocation and reducing As translocation. Environ. Sci. Pollut. Res. 2018, 25, 26978–26991. [Google Scholar] [CrossRef]
- Chowardhara, B.; Borgohain, P.; Saha, B.; Awasthi, J.P.; Moulick, D.; Panda, S.K. Phytotoxicity of Cd and Zn on three popular Indian mustard varieties during germination and early seedling growth. Biocatal. Agric. Biotechnol. 2019, 21, 101349. [Google Scholar] [CrossRef]
- Saha, B.; Chowardhara, B.; Kar, S.; Devi, S.S.; Awasthi, J.P.; Moulick, D.; Tanti, B.; Panda, S.K. Advances in heavy metal-induced stress alleviation with respect to exogenous amendments in crop plants. In Priming and Pretreatment of Seeds and Seedlings; Springer: Singapore, 2019; pp. 313–332. [Google Scholar]
- Moulick, D.; Samanta, S.; Saha, B.; Mazumder, M.K.; Dogra, S.; Panigrahi, K.C.; Banerjee, S.; Ghosh, D.; Santra, S.C. Salinity Stress Responses in Three Popular Field Crops Belonging to Fabaceae Family: Current Status and Future Prospect. In The Plant Family Fabaceae; Springer: Singapore, 2020; pp. 519–541. [Google Scholar]
- Meriga, B.; Reddy, B.K.; Rao, K.R.; Raddy, L.A.; Kavi Kishor, P.B. Aluminium-induced production of oxygen radicals, lipid peroxidation and DNA damage in seedlings of rice (Oryza sativa). J. Plant Physiol. 2004, 161, 63–68. [Google Scholar] [CrossRef]
- Dipierro, N.; Mondelli, D.; Paciolla, C.; Brunetti, G.; Dipierro, S. Changes in ascorbate system in the response of pumpkin (Cucarbita pepo L.) roots to aluminium stress. J. Plant Physiol. 2005, 162, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Schuch, M.W.; Cellini, A.; Masia, A.; Marino, G. Aluminium induced effects on growth, morphogenesis and oxidative stress reactions in in vitro cultures of quince. Sci. Hortic. 2010, 125, 151–158. [Google Scholar] [CrossRef]
- Wang, L.J.; Li, S.H. Salicylic acid induced heat or cold tolerance in relation to Ca2+ homeostasis and antioxidant system in young grape plants. Plant Sci. 2006, 170, 685–694. [Google Scholar] [CrossRef]
- Horst, W.J. The physiology manganese toxicity. In Manganese in Soil and Plants; Graham, R.D., Hannam, R.J., Uren, N.C., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1998; pp. 175–188. [Google Scholar]
- Doncheva, S.; Poschenrieder, C.; Stoyanova, Z.; Georgieva, K.; Velichkova, M.; Bareceló, J. Silicon amelioration of manganese toxicity in Mn sensitive and Mn tolerant maize varieties. Environ. Exp. Bot. 2009, 65, 189–197. [Google Scholar] [CrossRef]
- Sharma, A. Manganese in cell metabolism of higher plants. Bot. Rev. 1991, 57, 117–149. [Google Scholar]
- Gonzalez, A.; Steffen, K.L.; Lynch, J.P. Light and excess manganese. Plant Physiol. 1998, 118, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Zhao, Y.; Xu, Z.; Zhang, W.; Jiang, K. Physiological responses of Broussonetia papyrifera to manganese stress, a candidate plant for Phytoremediation. Ecotoxicol. Environ. Saf. 2019, 181, 18–25. [Google Scholar] [CrossRef]
- Braun, H.P. Effect of manganese toxicity on the proteome of the leaf apoplast in cowpea. Plant Physiol. 2003, 133, 1935–1946. [Google Scholar]
- Delhaize, E.; Gruber, B.D.; Pittman, J.K.; White, R.G.; Leung, H.; Miao, Y.; Jiang, L.; Ryan, P.R.; Richardson, A.E. A role for AtMTP11 gene of Arabidopsis in manganese transport and tolerance. Plant J. 2007, 51, 198–210. [Google Scholar] [CrossRef]
- Huang, H.; Zhao, Y.; Xu, Z.; Ding, Y.; Zhang, W.; Wu, L. Biosorption characteristics of a highly Mn(II)-resistant Ralstonia pickettii strain isolated from Mn ore. PLoS ONE 2018, 13, e0203285. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, A.; Hernandez, J.A.; del Rio, L.A.; Sevilla, F. Evidence for the presence of the ascorbate-glutathione cycle in mitochondria and peroxisomes of pea leaves. Plant Physiol. 1997, 114, 275–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, A.J.; Brach, T.; Marty, L.; Kreye, S.; Rouhier, N.; Jacquot, J.P.; Hell, R. Redox sensitive GFP in Arabidopsis thaliana is a quantitative biosensor for the redox potential of the cellular glutathione redox buffer. Plant J. 2007, 52, 973–986. [Google Scholar] [CrossRef] [PubMed]
- Bari, R.; Jones, J.D.G. Role of plant hormones in plant defense responses. Plant Mol Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, D.; Wei, J.; Ma, W.; Kong, X.; Rengel, Z.; Xiao, L.; Xia, G. Melatonin alleviates aluminium induced root growth inhibition by interfering with nitric oxide production in Arabidopsis. Environ. Exp. Bot. 2018, 161, 157–165. [Google Scholar] [CrossRef]
- Zhu, C.Q.; Zhang, J.H.; Sun, L.M.; Zhu, L.F.; Abliz, B.; Hu, W.J.; Zhong, C.; Bai, Z.G.; Sajid, H.; Cao, X.C.; et al. Hydrogen sulfide alleviates aluminium toxicity via decreasing apoplast and symplast Al contents in rice. Front. Plant Sci. 2018, 9, 294. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.; Panda, S.K. Role of salicylic acid in regulating cadmium (Cd) induced oxidative stress in Oryza sativa L. roots. Bulg. J. Plant Physiol. 2004, 30, 95–110. [Google Scholar]
- Cao, Y.; Zhang, Z.W.; Xue, L.W.; Du, J.B.; Shang, J.; Xu, F.; Yuan, S.; Lin, H.-H. Lack of salicylic acid in Arabidopsis protects plants against moderate salt stress. Z. Nat. C J. Biosci. 2009, 64, 231–238. [Google Scholar] [CrossRef]
- Nazar, R.; Umar, S.; Khan, N.A. Exogenous salicylic acid improves photosynthesis and growth through increase in ascorbate-glutathione metabolism and S assimilation in mustard under salt stress. Plant Signal. Behav. 2015, 10, e1003751. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Zheng, J.; Zhang, X.; Hu, Q.; Qian, R. Salicylic acid alleviates the adverse effects of salt stress on Dianthus superbus (Carophyllaceae) by activating photosynthesis, protecting morphological structure and enhancing the antioxidant system. Front. Plant Sci. 2017, 8, 600. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, C.; Li, L.P.; Sun, Z.Y. Effects of exogenous salicylic acid on growth and H2O2—Metabolizing enzymes in rice seedlings under lead stress. J. Environ. Sci. 2007, 19, 44–49. [Google Scholar] [CrossRef]
- Choudhury, S.; Moulick, D.; Mazumder, M.K. Secondary metabolites protect against metal and metalloid stress in rice: An in silico investigation using dehydroascorbate reductase. Acta Physiol. Plant 2020, 43, 3. [Google Scholar] [CrossRef]
- Zengin, F. Exogenous treatment with salicylic acid alleviating copper toxicity in bean seedlings. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2014, 84, 749–755. [Google Scholar] [CrossRef]
- Yoon, J.Y.; Hamayun, M.; Lee, S.K.; Lee, I.J. Methyl jasmonate alleviated salinity stress in soybean. J. Crop Sci. Biotechnol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Clarke, S.M.; Cristescu, S.M.; Miersch, O.; Harren, F.J.; Wastenack, C.; Mur, L.A. Jasmonates act with salicylic acid to confer basal thermotolerance in Arabidopsis thaliana. New Phytol. 2009, 182, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Zheng, B. Molecular responses during plant grafting and its regulation by auxins, cytokinins, and gibberellins. Biomolecules 2019, 9, 397. [Google Scholar] [CrossRef] [Green Version]
- Hare, P.D.; Cress, W.A.; Van, S.J. Dissecting the role of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Asada, K. The water—Water cycle in the chloroplast: Scavenging active oxygen species and dissipation of excess photons. Ann. Rev. Plant Physiol. Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Jimenez, A.; Mullineaux, P.M.; Sevilla, F. Tolerance of pea (Pisum sativum L.) to long term salt stress is associated with induction of antioxidant defenses. Plant Cell Environ. 2000, 23, 853–862. [Google Scholar] [CrossRef]
- Shao, H.B.; Jiang, S.Y.; Li, F.M.; Chu, L.Y.; Zhao, C.X.; Shao, M.A.; Zhao, X.; Li, F. Some advances in plant stress physiology and their implications in the systems biology era. Biointerfaces 2007, 54, 33–36. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Grimma, B.; Wobusa, U.; Wescheka, W. Differential responses of antioxidant compounds to salinity stress in salt-tolerant and salt-sensitive seedlings of foxtail millet (Setaria itilica). Physiol. Plant. 2000, 109, 435–442. [Google Scholar] [CrossRef]
- Mukherjee, S. Novel perspective of molecular crosstalk mechanisms of serotonin and melatonin in plants. Plant Physiol. Biochem. 2018, 132, 33–45. [Google Scholar] [CrossRef]
- Ke, Q.; Ye, J.; Wang, B.; Ren, J.; Yin, L.; Deng, X.; Wang, S. Melatonin mitigates salt stress in wheat seedlings by modulating polyamine metabolism. Front. Plant Sci. 2018, 9, 914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository—New features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazumder, M.K.; Borah, A.; Choudhury, S. Inhibitory potential of plant secondary metabolites on anti-Parkinsonian drug targets: Relevance to pathophysiology, and motor and non-motor behavioural abnormalities. Med. Hypotheses 2020, 137, 109544. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, M.K.; Choudhury, S.; Borah, A. An in silico investigation on the inhibitory potential of the constituents of Pomegranate juice on antioxidant defense mechanism: Relevance to neurodegenerative diseases. IBRO Rep. 2019, 6, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high-accuracy molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef]
- Ahmad, P.; Abass Ahanger, M.; Nasser Alyemeni, M.; Wijaya, L.; Alam, P.; Ashraf, M. Mitigation of sodium chloride toxicity in Solanum lycopersicum L. by supplementation of jasmonic acid and nitric oxide. J. Plant Interact. 2018, 13, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Alyemeni, M.N.; Ahanger, M.A.; Egamberdieva, D.; Wijaya, L.; Alam, P. Salicylic acid (SA) induced alterations in growth, biochemical attributes and antioxidant enzyme activity in faba bean (Vicia faba L.) seedlings under NaCl toxicity. Russ. J. Plant Physiol. 2018, 65, 104–114. [Google Scholar] [CrossRef]
- Ahmad, P.; Alyemeni, M.N.; Vijaya, L.; Alam, P.; Ahanger, M.A.; Alamri, S.A. Jasmonic acid alleviates negative impacts of cadmium stress by modifying osmolytes and antioxidants in faba bean (Vicia faba L.). Arch. Agron. Soil Sci. 2017, 63, 1889–1899. [Google Scholar] [CrossRef]
- Ahmad, P.; Nabi, G.; Ashraf, M. Cadmium-induced oxidative damage in mustard [Brassica juncea (L.) Czern. & Coss.] plants can be alleviated by salicylic acid. S. Afr. J. Bot. 2011, 77, 36–44. [Google Scholar]
- Ahmad, P.; Sarwat, M.; Sharma, S. Reactive oxygen species, antioxidants and signaling in plants. J. Plant Biol. 2008, 51, 167–173. [Google Scholar] [CrossRef]
- Mukherjee, S.; David, A.; Yadav, S.; Baluška, F.; Bhatla, S.C. Salt stress-induced seedling growth inhibition coincides with differential distribution of serotonin and melatonin in sunflower seedling roots and cotyledons. Physiol. Plant 2014, 152, 714–728. [Google Scholar] [CrossRef] [PubMed]
- Viehweger, K. How plants cope with heavy metals. Bot. Stud. 2014, 55, 35. [Google Scholar] [CrossRef] [Green Version]
- Byeon, Y.; Choi, G.H.; Lee, H.Y.; Back, K. Melatonin biosynthesis requires N-acetylserotonin methyltransferase activity of caffeic acid O-methyltransferase in rice. J. Exp. Bot. 2015, 66, 6917–6925. [Google Scholar] [CrossRef] [Green Version]
- Bali, S.; Kaur, P.; Sharma, A.; Ohri, P.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Jasmonic acid-induced tolerance to root-knot nematodes in tomato plants through altered photosynthetic and antioxidative defense mechanisms. Protoplasma 2017, 255, 471–484. [Google Scholar] [CrossRef]
- Bali, S.; Kaur, P.; Kohli, S.K.; Ohri, P.; Thukral, A.K.; Bhardwaj, R.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Jasmonic acid induced changes in physio-biochemical attributes and ascorbate-glutathione pathway in Lycopersicon esculentum under lead stress at different growth stages. Sci. Total Environ. 2018, 645, 1344–1360. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Combined effect of 24-epibrassinolide and salicylic acid mitigates lead (Pb) toxicity by modulating various metabolites in Brassica juncea L. seedlings. Protoplasma 2018, 255, 11–24. [Google Scholar] [CrossRef]
- Kohli, S.K.; Handa, N.; Sharma, A.; Gautam, V.; Arora, S.; Bhardwaj, R.; Alyemeni, M.N.; Ahmad, P. Interaction of 24-epibrassinolide and salicylic acid regulates pigment contents, antioxidative defense responses, and gene expression in Brassica juncea L. seedlings under Pb stress. Environ. Sci. Pollut. Res. 2018, 25, 15159–15173. [Google Scholar] [CrossRef]
- Mir, M.A.; John, R.; Alyemeni, M.N.; Alam, P.; Ahmad, P. Jasmonic acid ameliorates alkaline stress by improving growth performance, ascorbate glutathione cycle and glyoxylase system in maize seedlings. Sci. Rep. 2018, 8, 2831. [Google Scholar] [CrossRef]
- Mir, M.A.; Sirhindi, G.; Alyemeni, M.N.; Alam, P.; Ahmad, P. Jasmonic acid improves growth performance of soybean under nickel toxicity by regulating nickel uptake, redox balance, and oxidative stress metabolism. J. Plant Growth Regul. 2018, 37, 1195–1209. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | PubChem ID | Mol. Formula | Mol. Wt (g/mol) | XlogP3 | HBD | HBA | TPSA (Å2) |
|---|---|---|---|---|---|---|---|
| Salicylic acid | 338 | C7H6O3 | 138.12 | 2.3 | 2 | 3 | 57.5 |
| IAA | 802 | C10H9NO2 | 175.18 | 1.4 | 2 | 2 | 53.1 |
| Serotonin | 5202 | C10H12N2O | 176.21 | 0.2 | 3 | 2 | 62 |
| GA3 | 6466 | C19H22O6 | 346.4 | 0.2 | 3 | 6 | 104 |
| GSH | 124886 | C10H17N3O6S | 307.33 | −4.5 | 6 | 8 | 160 |
| DHA | 440667 | C6H6O6 | 174.11 | −1 | 2 | 6 | 101 |
| Jasmonic acid | 5281166 | C12H18O3 | 210.27 | 1.6 | 1 | 3 | 54.4 |
| MDHA | 53262277 | C6H7O6 | 175.12 | −1 | 2 | 5 | 90.8 |
| Ascorbic acid | 54670067 | C6H8O6 | 176.12 | −1.6 | 4 | 6 | 107 |
| Aluminium ion | 104727 | Al3+ | 26.981538 | - | 0 | 0 | 0 |
| Manganese ion | 27854 | Mn2+ | 54.93804 | - | 0 | 0 | 0 |
| Ligands | OsDHAR | ZmDHAR | TaDHAR | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MolDock | Rerank | HBond | MolDock | Rerank | HBond | MolDock | Rerank | HBond | |
| SA | −53.61 | −42.70 | −4.22 | −65.25 | −57.66 | −8.19 | −58.23 | −52.20 | −6.82 |
| IAA | −87.50 | −69.73 | −5.15 | −84.28 | −60.29 | −4.08 | −80.75 | −63.68 | −4.90 |
| 5-HT | −72.01 | −58.53 | −4.99 | −92.70 | −50.79 | −8.91 | −79.74 | −64.88 | −2.50 |
| GA3 | −84.02 | −70.38 | −3.46 | −99.20 | −78.81 | −5.72 | −92.83 | −58.92 | −5.62 |
| GSH | −91.60 | −75.41 | −5.74 | −103.79 | −85.35 | −8.92 | −96.60 | −82.38 | −9.31 |
| JA | −93.23 | −72.72 | −4.60 | −96.34 | −18.03 | −5.00 | −84.62 | −65.49 | −4.62 |
| AsA | −62.38 | −56.41 | −8.61 | −72.86 | −67.44 | −9.82 | −70.58 | −63.03 | −13.44 |
| Al3+ | −25.81 | −30.59 | 0.00 | −41.36 | −34.43 | 0.00 | −22.05 | −21.93 | 0.00 |
| Mn2+ | −26.98 | −30.41 | 0.00 | −41.37 | −34.42 | 0.00 | −23.44 | −21.66 | 0.00 |
| Ligands | OsMDHAR | ZmMDHAR | TaMDHAR | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MolDock | Rerank | HBond | MolDock | Rerank | HBond | MolDock | Rerank | HBond | |
| SA | −69.24 | −56.19 | −5.00 | −71.05 | −59.15 | −4.82 | −70.20 | −59.84 | −5.78 |
| IAA | −102.96 | −83.96 | −4.81 | −100.38 | −79.58 | −5.73 | −98.33 | −81.03 | −3.17 |
| 5-HT | −102.67 | −84.13 | −6.06 | −99.78 | −78.83 | −4.12 | −103.73 | −84.33 | −5.87 |
| GA3 | −116.56 | −78.70 | −6.36 | −117.03 | −60.76 | −7.58 | −119.83 | −95.04 | −5.81 |
| GSH | −126.49 | −78.36 | −12.95 | −125.39 | −91.09 | −15.93 | −141.58 | −122.03 | −18.32 |
| JA | −111.14 | −91.62 | −4.44 | −109.56 | −89.51 | −7.06 | −114.21 | −92.53 | −6.59 |
| AsA | −81.00 | −66.94 | −8.07 | −88.97 | −80.97 | −13.70 | −89.68 | −78.91 | −12.48 |
| Al3+ | −41.24 | −41.05 | 0.00 | −39.50 | −31.69 | 0.00 | −32.45 | −32.75 | 0.00 |
| Mn2+ | −41.24 | −41.05 | 0.00 | −39.51 | −31.70 | 0.00 | −32.45 | −32.75 | 0.00 |
| Ligand Property | OsDHAR | ZmDHAR | TaDHAR | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MolDock | Rerank | Hbond | MolDock | Rerank | Hbond | MolDock | Rerank | Hbond | |
| Mol. Wt | 0.61 | 0.64 | NC | NC | 0.51 | NC | NC | NC | NC |
| XlogP3 | NC | NC | −0.51 | NC | NC | −0.58 | NC | NC | −0.58 |
| HBD | NC | NC | 0.60 | NC | 0.72 | NC | NC | NC | NC |
| HBA | NC | NC | 0.73 | NC | NC | NC | NC | NC | 0.68 |
| TPSA | NC | NC | 0.74 | NC | 0.53 | NC | NC | NC | 0.58 |
| Ligand Property | OsMDHAR | ZmMDHAR | TaMDHAR | ||||||
|---|---|---|---|---|---|---|---|---|---|
| MolDock | Rerank | Hbond | MolDock | Rerank | Hbond | MolDock | Rerank | Hbond | |
| Mol. Wt | 0.57 | NC | NC | 0.54 | NC | NC | 0.59 | 0.54 | NC |
| XlogP3 | NC | NC | −0.95 | NC | NC | −0.87 | NC | NC | −0.93 |
| HBD | NC | NC | 0.73 | NC | NC | 0.79 | NC | NC | 0.79 |
| HBA | NC | NC | 0.83 | NC | NC | 0.85 | NC | NC | 0.83 |
| TPSA | NC | NC | 0.90 | NC | NC | 0.88 | NC | NC | 0.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choudhury, S.; Mazumder, M.K.; Moulick, D.; Sharma, P.; Tata, S.K.; Ghosh, D.; Ali, H.M.; Siddiqui, M.H.; Brestic, M.; Skalicky, M.; et al. A Computational Study of the Role of Secondary Metabolites for Mitigation of Acid Soil Stress in Cereals Using Dehydroascorbate and Mono-Dehydroascorbate Reductases. Antioxidants 2022, 11, 458. https://doi.org/10.3390/antiox11030458
Choudhury S, Mazumder MK, Moulick D, Sharma P, Tata SK, Ghosh D, Ali HM, Siddiqui MH, Brestic M, Skalicky M, et al. A Computational Study of the Role of Secondary Metabolites for Mitigation of Acid Soil Stress in Cereals Using Dehydroascorbate and Mono-Dehydroascorbate Reductases. Antioxidants. 2022; 11(3):458. https://doi.org/10.3390/antiox11030458
Chicago/Turabian StyleChoudhury, Shuvasish, Muhammed Khairujjaman Mazumder, Debojyoti Moulick, Parul Sharma, Sandeep Kumar Tata, Dibakar Ghosh, Hayssam M. Ali, Manzer H. Siddiqui, Marian Brestic, Milan Skalicky, and et al. 2022. "A Computational Study of the Role of Secondary Metabolites for Mitigation of Acid Soil Stress in Cereals Using Dehydroascorbate and Mono-Dehydroascorbate Reductases" Antioxidants 11, no. 3: 458. https://doi.org/10.3390/antiox11030458