

Do Volatiles Affect Bacteria and Plants in the Same Way? Growth and Biochemical Response of Non-Stressed and Cd-Stressed Arabidopsis thaliana and Rhizobium E20-8


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals
2.2. Bacterial Strain
2.3. Bacterial Experiment Set-Up
2.4. Plant Experiment Set-Up
2.5. Biochemical Analyses
2.5.1. Extraction
2.5.2. Protein Content
2.5.3. Oxidative Damage
2.5.4. Antioxidant Enzymes
2.6. Statistical Analyses
3. Results
3.1. Cd and Volatiles Influence on Growth
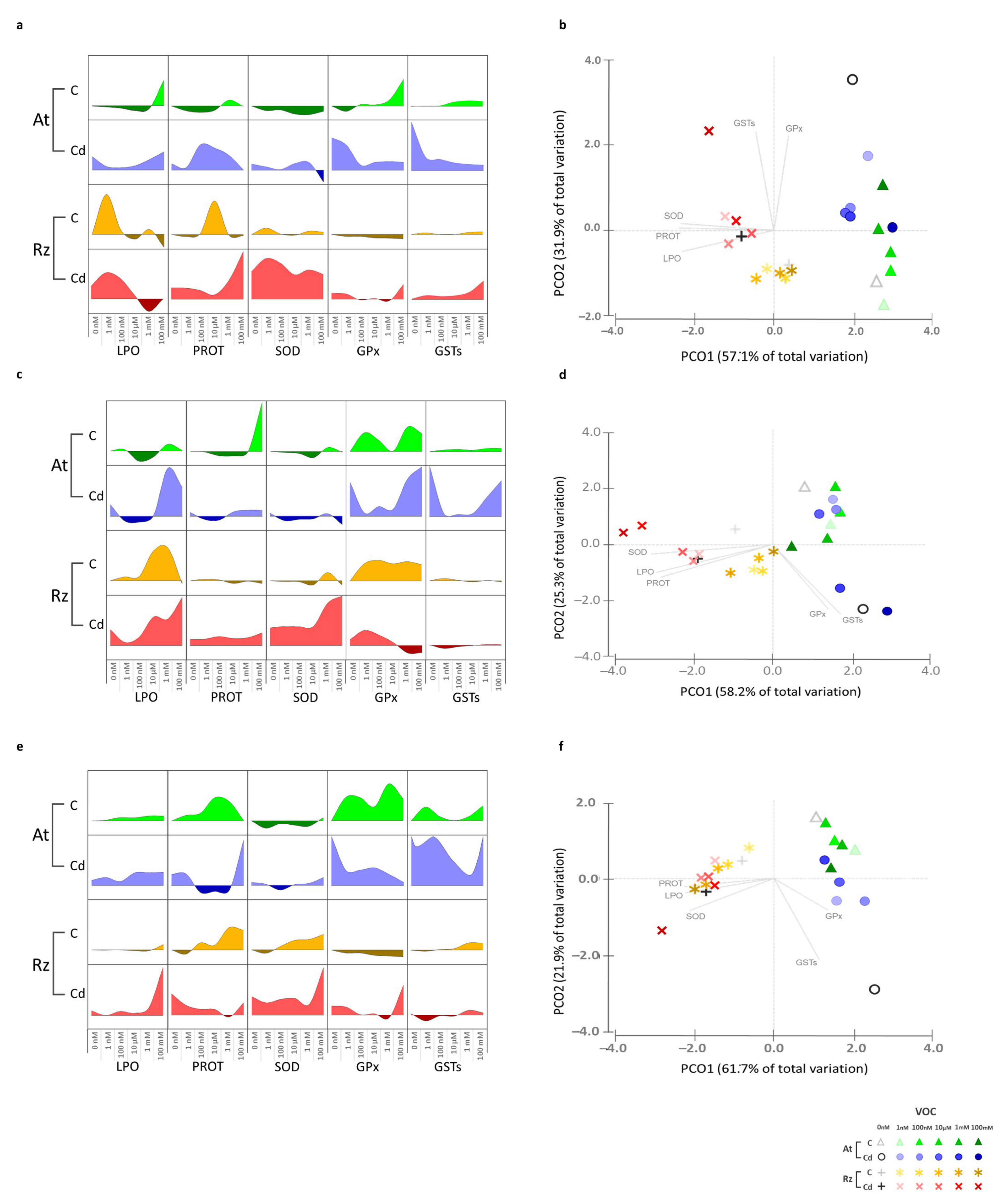
3.2. Cd and Volatile Influence on Plant and Bacterium Biochemistry
3.2.1. Cadmium
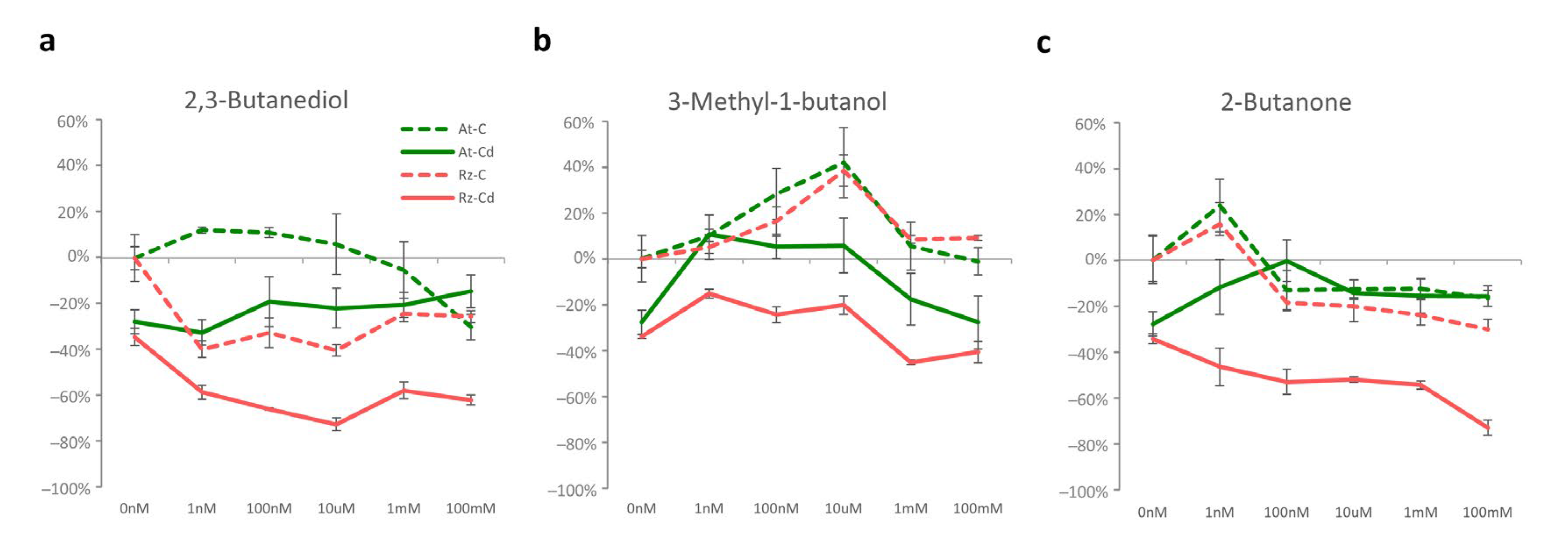
3.2.2. 2,3-Butanediol (Alone and in Combination with Cd)
3.2.3. 3-Methyl-1-Butanol (Alone and in Combination with Cd)
3.2.4. 2-Butanone (Alone and in Combination with Cd)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harman, G.; Khadka, R.; Doni, F.; Uphoff, N. Benefits to Plant Health and Productivity from Enhancing Plant Microbial Symbionts. Front. Plant Sci. 2021, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- Roman-Reyna, V.; Pinili, D.; Borja, F.N.; Quibod, I.L.; Groen, S.C.; Mulyaningsih, E.S.; Rachmat, A.; Slamet-Loedin, I.H.; Alexandrov, N.; Mauleon, R.; et al. The Rice Leaf Microbiome Has a Conserved Community Structure Controlled by Complex Host-Microbe Interactions. bioRxiv 2019. [Google Scholar] [CrossRef]
- Berg, G. Plant-Microbe Interactions Promoting Plant Growth and Health: Perspectives for Controlled Use of Microorganisms in Agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of Phosphorus and Nitrogen in the Rhizosphere and Plant Growth Promotion by Microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Van Loon, L.C. Plant Responses to Plant Growth-Promoting Rhizobacteria. Eur. J. Plant Pathol. 2007, 119, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Helepciuc, F.E.; Mitoi, M.E.; Manole-Pǎunescu, A.; Aldea, F.; Brezeanu, A.; Cornea, C.P. Induction of Plant Antioxidant System by Interaction with Beneficial and/or Pathogenic Microorganisms. Rom. Biotechnol. Lett. 2014, 19, 9366–9375. [Google Scholar]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Pare, P.W.; Kloepper, J.W. Bacterial Volatiles Promote Growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [Green Version]
- De Moraes, C.M.; Mescher, M.C.; Tumlinson, J.H. Caterpillar-Induced Nocturnal Plant Volatiles Repel Conspecific Females. Nature 2001, 410, 577–580. [Google Scholar] [CrossRef]
- Dicke, M.; De Boer, J.G.; Ho, M.; Rocha-granados, M.C. Mixed Blends of Herbivore-Induced Plant Volatiles and Foraging Success of Carnivorous Arthropods. Oikos 2003, 1, 38–48. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-Kingdom Interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef] [Green Version]
- Kappers, I.F. Genetic Engineering of Terpenoid Metabolism Attracts Bodyguards to Arabidopsis. Science 2005, 309, 2070–2072. [Google Scholar] [CrossRef] [PubMed]
- Schnee, C.; Kollner, T.G.; Held, M.; Turlings, T.C.J.; Gershenzon, J.; Degenhardt, J. The Products of a Single Maize Sesquiterpene Synthase Form a Volatile Defense Signal That Attracts Natural Enemies of Maize Herbivores. Proc. Natl. Acad. Sci. USA 2006, 103, 1129–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidorova, D.E.; Plyuta, V.A.; Padiy, D.A.; Kupriyanova, E.V.; Roshina, N.V.; Koksharova, O.A.; Khmel, I.A. The Effect of Volatile Organic Compounds on Different Organisms: Agrobacteria, Plants and Insects. Microorganisms 2021, 10, 69. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Tang, M.; Tang, X.; Sun, W.; Gong, J.; Yi, Y. Bacillus subtilis—Arabidopsis thaliana: A Model Interaction System for Studying the Role of Volatile Organic Compounds in the Interchange between Plants and Bacteria. Botany 2019, 97, 661–669. [Google Scholar] [CrossRef]
- Park, K.Y.; Seo, S.Y.; Oh, B.R.; Seo, J.W.; Kim, Y.J. 2,3-Butanediol Induces Systemic Acquired Resistance in the Plant Immune Response. J. Plant Biol. 2018, 61, 424–434. [Google Scholar] [CrossRef]
- Ameye, M.; Audenaert, K.; De Zutter, N.; Steppe, K.; Van Meulebroek, L.; Vanhaecke, L.; De Vleesschauwer, D.; Haesaert, G.; Smagghe, G. Priming of Wheat with the Green Leaf Volatile Z-3-Hexenyl Acetate Enhances Defense against Fusarium Graminearum but Boosts Deoxynivalenol Production. Plant Physiol. 2015, 167, 1671–1684. [Google Scholar] [CrossRef] [Green Version]
- Cofer, T.M.; Engelberth, M.; Engelberth, J. Green Leaf Volatiles Protect Maize (Zea mays) Seedlings against Damage from Cold Stress. Plant Cell Environ. 2018, 41, 1673–1682. [Google Scholar] [CrossRef]
- Vespermann, A.; Kai, M.; Piechulla, B. Rhizobacterial Volatiles Affect the Growth of Fungi and Arabidopsis thaliana. Appl. Environ. Microbiol. 2007, 73, 5639–5641. [Google Scholar] [CrossRef] [Green Version]
- Fincheira, P.; Quiroz, A.; Tortella, G.; Diez, M.C.; Rubilar, O. Current Advances in Plant-Microbe Communication via Volatile Organic Compounds as an Innovative Strategy to Improve Plant Growth. Microbiol. Res. 2021, 247, 126726. [Google Scholar] [CrossRef]
- Santoro, M.V.; Zygadlo, J.; Giordano, W.; Banchio, E. Volatile Organic Compounds from Rhizobacteria Increase Biosynthesis of Essential Oils and Growth Parameters in Peppermint (Mentha piperita). Plant Physiol. Biochem. 2011, 49, 1177–1182. [Google Scholar] [CrossRef]
- Park, Y.S.; Dutta, S.; Ann, M.; Raaijmakers, J.M.; Park, K. Promotion of Plant Growth by Pseudomonas Fluorescens Strain SS101 via Novel Volatile Organic Compounds. Biochem. Biophys. Res. Commun. 2015, 461, 361–365. [Google Scholar] [CrossRef]
- Ursínyová, M.; Hladíková, V. Cadmium in the Environment of Central Europe. In Trace Elements—Their Distribution and Effects in the Environment; Markert, B., Friese, K., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2000; Volume 4, pp. 87–107. [Google Scholar]
- Khan, M.A.; Khan, S.; Khan, A.; Alam, M. Soil Contamination with Cadmium, Consequences and Remediation Using Organic Amendments. Sci. Total Environ. 2017, 601–602, 1591–1605. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, L.; Ma, Z.; Wang, J. Bacillus amyloliquefaciens SAY09 Increases Cadmium Resistance in Plants by Activation of Auxin-Mediated Signaling Pathways. Genes 2017, 8, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, A.I.G.; Corticeiro, S.C.; de Almeida Paula Figueira, E.M. Glutathione-Mediated Cadmium Sequestration in Rhizobium leguminosarum. Enzyme Microb. Technol. 2006, 39, 763–769. [Google Scholar] [CrossRef]
- Figueira, E.M.D.A.P.; Gusmão Lima, A.I.; Pereira, S.I.A. Cadmium Tolerance Plasticity in Rhizobium leguminosarum Bv. v Iciae: Glutathione as a Detoxifying Agent. Can. J. Microbiol. 2005, 51, 7–14. [Google Scholar] [CrossRef] [PubMed]
- ATSDR-Agency for Toxic Substances and Disease Registry, 2019. Substance Priority List. 2019. Available online: https://www.atsdr.cdc.gov/spl/index.html (accessed on 20 September 2022).
- Navarro-León, E.; Manuel, J.; Albacete, A.; Blasco, B. Ecotoxicology and Environmental Safety Tolerance to Cadmium Toxicity and Phytoremediation Potential of Three Brassica Rapa CAX1a TILLING Mutants. Ecotoxicol. Environ. Saf. 2019, 189, 109961. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xue, B.; Li, N.; Song, G. Effects of Cutting Frequency of Tall Fescue on Cadmium Form in Soil and Cadmium Enrichment in the Plant. Agron. J. 2020, 112, 4693–4704. [Google Scholar] [CrossRef]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A. The Effects of Cadmium Toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef]
- Wang, Q.-R.; Cui, Y.-S.; Liu, X.-M.; Dong, Y.-T.; Christie, P. Soil Contamination and Plant Uptake of Heavy Metals at Polluted Sites in China. J. Environ. Sci. Health Part A 2003, 38, 823–838. [Google Scholar] [CrossRef] [Green Version]
- Naeem, M.A.; Shabbir, A.; Amjad, M.; Abbas, G.; Imran, M.; Murtaza, B.; Tahir, M.; Ahmad, A. Acid Treated Biochar Enhances Cadmium Tolerance by Restricting Its Uptake and Improving Physio-Chemical Attributes in Quinoa (Chenopodium quinoa Willd.). Ecotoxicol. Environ. Saf. 2020, 191, 110218. [Google Scholar] [CrossRef]
- Itoh, S.; Yumura, Y. Studies on the Contamination of Vegetable Crops by Excessive Absorption of Heavy Metals. Bull. Veg. Ornam. Crop. Res. Stn. 1979, 123, 145–166. [Google Scholar]
- Andresen, E.; Ku, H. Cadmium Toxicity in Plants; Springer: Berlin/Heidelberg, Germany, 2013; pp. 395–413. [Google Scholar] [CrossRef]
- Lu, M.; Xu, K.; Chen, J. Effect of Pyrene and Cadmium on Microbial Activity and Community Structure in Soil. Chemosphere 2013, 91, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Liu, Q.; Liu, Y.J.; Jia, F.A.; He, X.H. Responses of Soil Microbial Activity to Cadmium Pollution and Elevated CO2. Sci. Rep. 2014, 4, 4287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Nie, S.; Liang, J.; Zeng, G.; Wu, H.; Hua, S.; Liu, J.; Yuan, Y.; Xiao, H.; Deng, L.; et al. Effects of Heavy Metals and Soil Physicochemical Properties on Wetland Soil Microbial Biomass and Bacterial Community Structure. Sci. Total Environ. 2016, 557–558, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Workentine, M.L.; Weljie, A.M.; Vogel, H.J.; Ceri, H.; Viti, C.; Tatti, E.; Zhang, P.; Hynes, A.P.; Turner, R.J.; et al. Metabolomic Investigation of the Bacterial Response to a Metal Challenge. Appl. Environ. Microbiol. 2009, 75, 719–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, B.E.; Halbfeld, C.; Blank, L.M. Exploration and Exploitation of the Yeast Volatilome. Curr. Metab. 2016, 5, 102–118. [Google Scholar] [CrossRef]
- Cardoso, P.; Santos, M.; Freitas, R.; Rocha, S.M.; Figueira, E. Response of Rhizobium to Cd Exposure: A Volatile Perspective. Environ. Pollut. 2017, 231, 802–811. [Google Scholar] [CrossRef]
- Garbeva, P.; Hordijk, C.; Gerards, S.; de Boer, W. Volatiles Produced by the Mycophagous Soil Bacterium Collimonas. FEMS Microbiol. Ecol. 2014, 87, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, P.; Corticeiro, S.; Freitas, R.; Figueira, E. Different Efficiencies of the Same Mechanisms Result in Distinct Cd Tolerance within Rhizobium. Ecotoxicol. Environ. Saf. 2018, 150, 260–269. [Google Scholar] [CrossRef]
- Matos, D.; Sá, C.; Cardoso, P.; Pires, A.; Figueira, E. Rhizobium Sensing of Airborne Saturated Aldehydes of Different Sizes Modulates the Response to Cd Exposure. J. Hazard. Mater. 2020, 395, 122629. [Google Scholar] [CrossRef]
- Sá, C.; Matos, D.; Pires, A.; Cardoso, P.; Figueira, E. Effects of Volatile Sulfur Compounds on Growth and Oxidative Stress of Rhizobium Leguminosarum E20-8 Exposed to Cadmium. Sci. Total Environ. 2021, 800, 149478. [Google Scholar] [CrossRef] [PubMed]
- Ismael, M.A.; Elyamine, M.; Moussa, M.G.; Cai, M.; Hu, C. Interactions with Selenium Fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.M.; Lee, D.A.; Schroeder, J.I. Long-Distance Root-to-Shoot Transport of Phytochelatins and Cadmium in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 10118–10123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perfus-Barbeoch, L.; Leonhardt, N.; Vavasseu, A.; Forestier, C.; Cadarache, C.E.A.; Membranaires, E.; Cea-cnrs, U.M.R. Heavy Metal Toxicity: Cadmium Permeates through Calcium Channels and Disturbs the Plant Water Status. Plant J. 2002, 32, 539–548. [Google Scholar] [CrossRef]
- Clemens, S. Toxic Metal Accumulation, Responses to Exposure and Mechanisms of Tolerance in Plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef]
- Cappellari, L.D.; Chiappero, J.; Palermo, T.B.; Giordano, W.; Banchio, E. Volatile Organic Compounds from Rhizobacteria Increase the Biosynthesis of Secondary Metabolites and Improve the Antioxidant Status in Mentha piperita L. Grown under Salt Stress. Agronomy 2020, 10, 1094. [Google Scholar] [CrossRef]
- Corticeiro, S.; Freitas, R.; Figueira, E. The Role of GSTs in the Tolerance of Rhizobium leguminosarum to Cadmium. BioMetals 2013, 26, 879–886. [Google Scholar] [CrossRef]
- Somasegaran, P.; Hoben, H.J. Handbook for Rhizobia: Methods in Legume-Rhizobium Technology; Somasegaran, P., Hoben, H.J., Eds.; Springer: Berlin/Heidelberg, Germany, 1994; Volume 70, pp. 224–225. [Google Scholar] [CrossRef]
- Sá, C.; Matos, D.; Pires, A.; Cardoso, P.; Figueira, E. Airborne Exposure of Rhizobium leguminosarum Strain E20-8 to Volatile Monoterpenes: Effects on Cells Challenged by Cadmium. J. Hazard. Mater. 2019, 387, 121783. [Google Scholar] [CrossRef]
- Matos, D.; Sá, C.; Cardoso, P.; Pires, A.; Rocha, S.M.; Figueira, E. The Role of Volatiles in Rhizobium Tolerance to Cadmium: Effects of Aldehydes and Alcohols on Growth and Biochemical Endpoints. Ecotoxicol. Environ. Saf. 2019, 186, 109759. [Google Scholar] [CrossRef]
- Lopes, T.; Cruz, C.; Cardoso, P.; Pinto, R.; Marques, P.A.A.P.; Figueira, E. A Multifactorial Approach to Untangle Graphene Oxide (GO) Nanosheets Effects on Plants: Plant Growth-Promoting Bacteria Inoculation, Bacterial Survival, and Drought. Nanomaterials 2021, 11, 771. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal Lipid Peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, C.S.; Oliveira, R.; Bento, F.; Geraldo, D.; Rodrigues, J.V.; Marcos, J.C. Simplified 2,4-Dinitrophenylhydrazine Spectrophotometric Assay for Quantification of Carbonyls in Oxidized Proteins. Anal. Biochem. 2014, 458, 69–71. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Udechukwu, M.C.; Yiridoe, C.; Gibson, A.; Gong, M. Antioxidant Mechanism of Potato Protein Hydrolysates against in Vitro Oxidation of Reduced Glutathione. J. Funct. Foods 2016, 20, 195–203. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the Quantitative and Qualitative Characterization of Erythrocyte Glutathione Peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases. The First Enzymatic Step in Mercapturic Acid Formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.N.; Clarke, K. PERMANOVA+ for Primer: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Han, S.H.; Lee, S.J.; Moon, J.H.; Park, K.H.; Yang, K.Y.; Cho, B.H.; Kim, K.Y.; Kim, Y.W.; Lee, M.C.; Anderson, A.J.; et al. GacS-Dependent Production of 2R, 3R-Butanediol by Pseudomonas Chlororaphis O6 Is a Major Determinant for Eliciting Systemic Resistance against Erwinia carotovora but Not against Pseudomonas syringae pv. tabaci in Tobacco. Mol. Plant Microbe Interact. 2006, 19, 924–930. [Google Scholar] [CrossRef] [Green Version]
- Li, T.-Y.; Ye, C.; Zhang, Y.-J.; Zhang, J.-X.; Yang, M.; He, X.-H.; Mei, X.-Y.; Liu, Y.-X.; Zhu, Y.-Y.; Huang, H.-C.; et al. 2,3-Butanediol from the Leachates of Pine Needles Induces the Resistance of Panax Notoginseng to the Leaf Pathogen Alternaria Panax. Plant Divers. 2022, in press. [Google Scholar] [CrossRef]
- Park, A.R.; Kim, J.; Kim, B.; Ha, A.; Son, J.; Song, C.W.; Song, H.; Kim, J. Enhanced Abiotic Stress Tolerance of Tomato and Turfgrass under Drought or Chilling Stress. J. Microbiol. Biotechnol. 2022, 32, 582–593. [Google Scholar] [CrossRef]
- Lee, S.; Hung, R.; Yin, G.; Klich, M.A.; Grimm, C.; Bennett, J.W. Arabidopsis Thaliana as Bioindicator of Fungal VOCs in Indoor Air. Mycobiology 2016, 44, 162–170. [Google Scholar] [CrossRef]
- Lee, S.; Behringer, G.; Hung, R.; Bennett, J. Effects of Fungal Volatile Organic Compounds on Arabidopsis thaliana Growth and Gene Expression. Fungal Ecol. 2019, 37, 1–9. [Google Scholar] [CrossRef]
- Gamboa, R.; Damaris, B.; Jorge, D.; Torres, M.; Ramírez, E.; Kiel, A.L.; Gloria, M.; Randy, C.; Castro, O. Plant Growth—Promoting and Non-Promoting Rhizobacteria from Avocado Trees Differentially Emit Volatiles That Influence Growth of Arabidopsis thaliana. Protoplasma 2022, 259, 835–854. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.L.; Wang, Y.H.; Wu, X.Q. Enhanced Iron Uptake in Plants by Volatile Emissions of Rahnella aquatilis JZ-GX1. Front. Plant Sci. 2021, 12, 704000. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Ryu, C.-M. Two Volatile Organic Compounds Trigger Plant Self-Defense against a Bacterial Pathogen and a Sucking Insect in Cucumber under Open Field Conditions. Int. J. Mol. Sci. 2013, 14, 9803–9819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewari, R.K.; Yadav, N.; Gupta, R.; Kumar, P. Oxidative Stress Under Macronutrient Deficiency in Plants. J. Soil Sci. Plant Nutr. 2021, 21, 832–859. [Google Scholar] [CrossRef]
- Sytar, O.; Kumar, A.; Latowski, D.; Kuczynska, P.; Strzałka, K.; Prasad, M.N.V. Heavy Metal-Induced Oxidative Damage, Defense Reactions, and Detoxification Mechanisms in Plants. Acta Physiol. Plant. 2013, 35, 985–999. [Google Scholar] [CrossRef]
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A Unified Mechanism of Action for Volatile Isoprenoids in Plant Abiotic Stress. Nat. Chem. Biol. 2009, 5, 283–291. [Google Scholar] [CrossRef]
- Loreto, F.; Mannozzi, M.; Maris, C.; Nascetti, P.; Ferranti, F.; Pasqualini, S. Ozone Quenching Properties of Isoprene and Its Antioxidant Role in Leaves. Plant Physiol. 2001, 126, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Loreto, F.; Velikova, V. Isoprene Produced by Leaves Protects the Photosynthetic Apparatus against Ozone Damage, Quenches Ozone Products, and Reduces Lipid Peroxidation of Cellular Membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Vaishnav, A.; Kumari, S.; Jain, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. PGPR-Mediated Expression of Salt Tolerance Gene in Soybean through Volatiles under Sodium Nitroprusside. J. Basic Microbiol. 2016, 56, 1274–1288. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Ryu, C.-M.; Lee, S.; Park, H.B.; Han, K.S.; Lee, J.H.; Lee, K.; Chung, W.S.; Jeong, M.-J.; Kim, H.K.; et al. Proteome Analysis of Arabidopsis Seedlings Exposed to Bacterial Volatiles. Planta 2010, 232, 1355–1370. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sá, C.; Matos, D.; Cardoso, P.; Figueira, E. Do Volatiles Affect Bacteria and Plants in the Same Way? Growth and Biochemical Response of Non-Stressed and Cd-Stressed Arabidopsis thaliana and Rhizobium E20-8. Antioxidants 2022, 11, 2303. https://doi.org/10.3390/antiox11112303
Sá C, Matos D, Cardoso P, Figueira E. Do Volatiles Affect Bacteria and Plants in the Same Way? Growth and Biochemical Response of Non-Stressed and Cd-Stressed Arabidopsis thaliana and Rhizobium E20-8. Antioxidants. 2022; 11(11):2303. https://doi.org/10.3390/antiox11112303
Chicago/Turabian StyleSá, Carina, Diana Matos, Paulo Cardoso, and Etelvina Figueira. 2022. "Do Volatiles Affect Bacteria and Plants in the Same Way? Growth and Biochemical Response of Non-Stressed and Cd-Stressed Arabidopsis thaliana and Rhizobium E20-8" Antioxidants 11, no. 11: 2303. https://doi.org/10.3390/antiox11112303