
Controlled Atmosphere Improves the Quality, Antioxidant Activity and Phenolic Content of Yellow Peach during the Shelf Life

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Controlled Atmosphere (CA) Treatment
2.2. Assay of Weight Loss Rate (WLR)
2.3. Assay of Browning Index (BI)
2.4. Assay of Firmness
2.5. Assay of the Decay Rate (DR)
2.6. Assay of Color
2.7. Assay of DPPH, ABTS and FRAP Scavenging Activity
2.8. Assay of Phenylalanine-Catalase Activity (PAL)
2.9. Assay of Polyphenol Oxidase Activity (PPO)
2.10. Assay of Peroxidase Activity (POD)
2.11. Assay of Total Flavonoid Content (TFC)
2.12. Assay of Total Phenolic Content (TPC)
2.13. Assay of Phenolic Acids Content
2.14. Statistical Analysis
3. Results
3.1. Color
3.2. Browning Index (BI)
3.3. Firmness
3.4. The Decay Rate (DR)
3.5. The Weight Loss Rate (WLR)
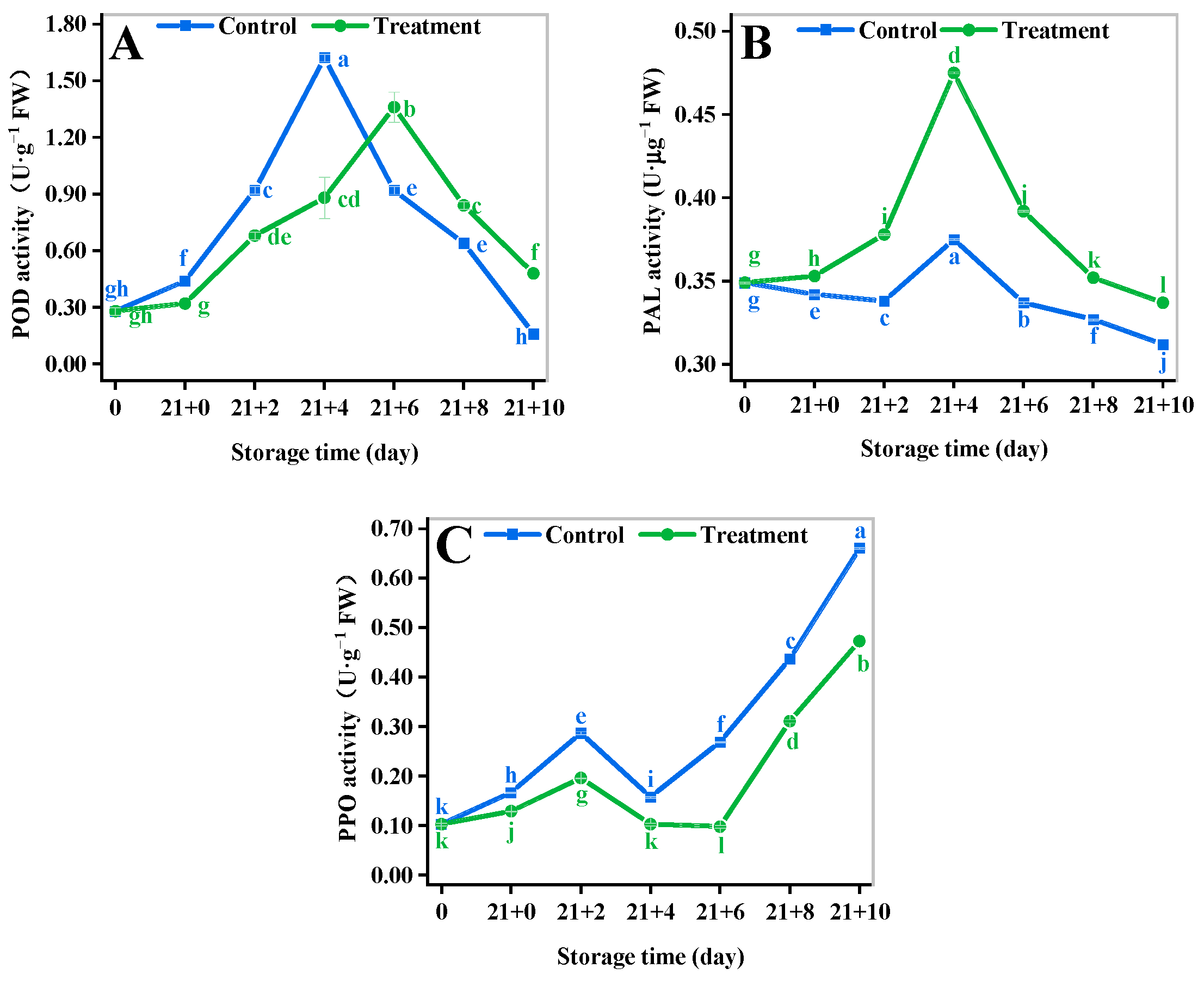
3.6. Browning-Related Enzymes
3.7. Total Phenolic Content (TPC)
3.8. Total Flavonoid Content (TFC)
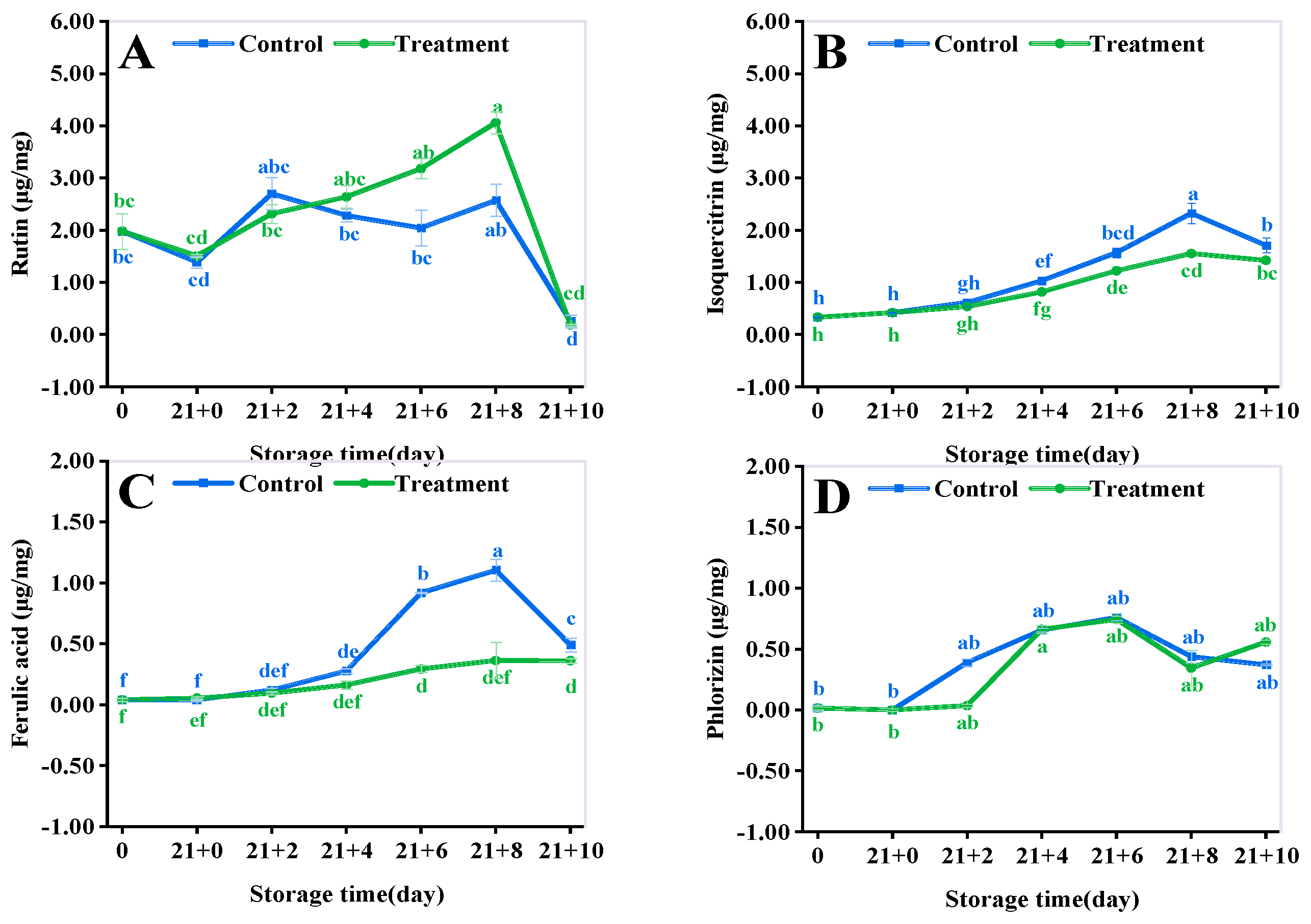
3.9. Phenolic Acid Content
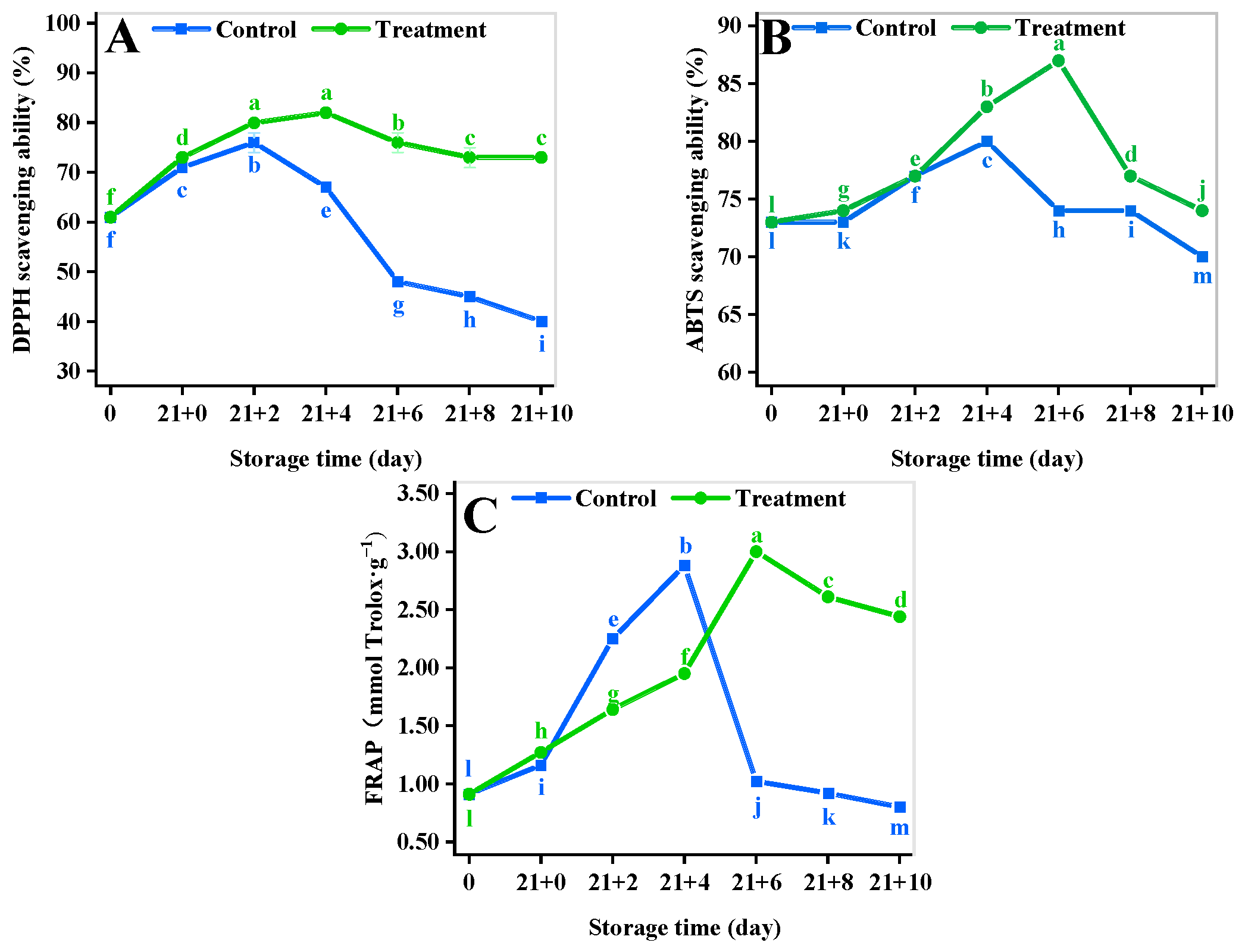
3.10. Antioxidant Activity Assay
3.11. Correlation Analysis
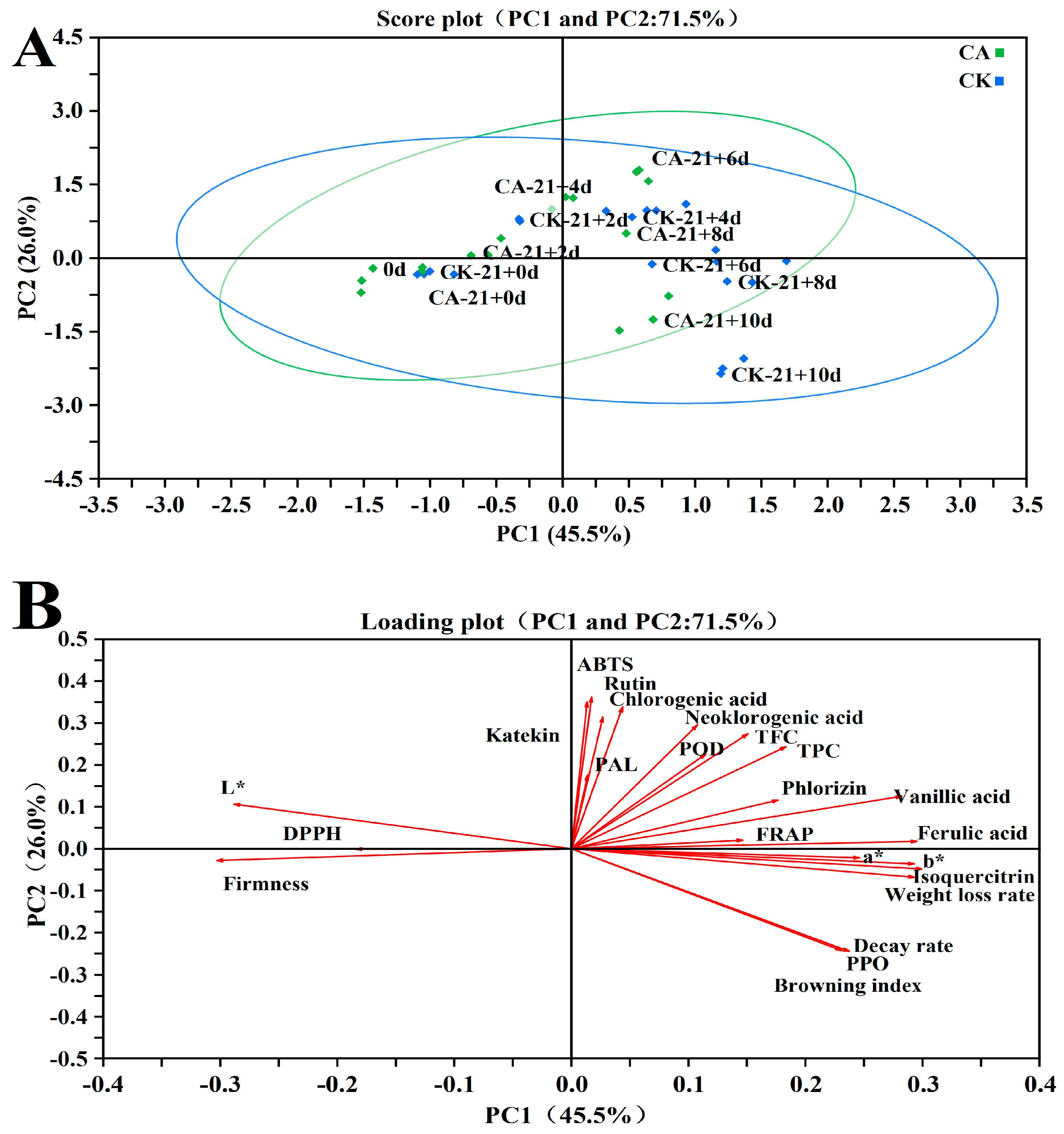
3.12. Principal Component Analysis (PCA)
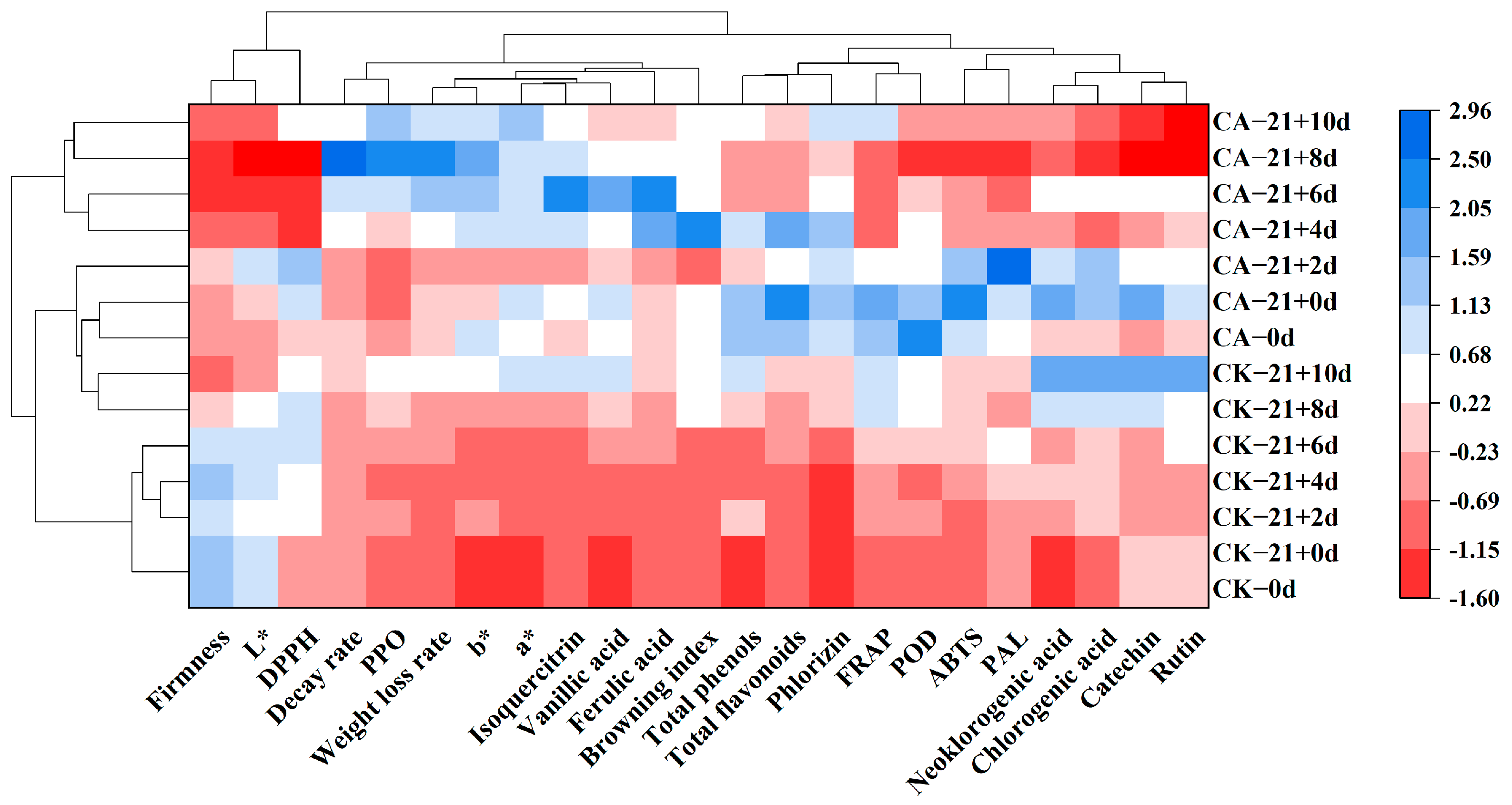
3.13. Hierarchical Clustering Analysis (HCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, L.; Chen, S.; Shao, J.; Zhang, C.; Mei, L.; Wang, K.; Jin, P.; Zheng, Y. Hydrogen sulfide alleviates chilling injury in peach fruit by maintaining cell structure integrity via regulating endogenous H2S, antioxidant and cell wall metabolisms. Food Chem. 2022, 391, 133283. [Google Scholar] [CrossRef] [PubMed]
- Byrne, D.; Vizzotto, M.; Cisneros-Zevallos, L.; Ramming, D.; Okie, W. Antioxidant Content of Peach and Plum Genotypes. Hortscience 2004, 39, 2327–9834. [Google Scholar] [CrossRef]
- Fan, X.; Argenta, L.; Mattheis, J.P. Interactive Effects of 1-MCP and Temperature on ‘Elberta’ Peach Quality. Hortscience 2002, 37, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, L.; Wang, Q.; Zuo, J. Low temperature conditioning combined with methyl jasmonate can reduce chilling injury in bell pepper. Sci. Hortic. 2019, 243, 434–439. [Google Scholar] [CrossRef]
- Besada, C.; Novillo, P.; Navarro, P.; Salvador, A. Effect of a low oxygen atmosphere combined with 1-MCP pretreatment on preserving the quality of ‘Rojo Brillante’ and ‘Triumph’ persimmon during cold storage. Sci. Hortic. 2014, 179, 51–58. [Google Scholar] [CrossRef]
- Huyskens-Keil, S.; Prono-Widayat, H.; Lüdders, P.; Schreiner, M. Postharvest quality of pepino (Solanum muricatum Ait.) fruit in controlled atmosphere storage. J. Food Eng. 2006, 77, 628–634. [Google Scholar] [CrossRef]
- Park, M.H.; Sangwanangkul, P.; Choi, J.W. Reduced chilling injury and delayed fruit ripening in tomatoes with modified atmosphere and humidity packaging. Sci. Hortic. 2018, 231, 66–72. [Google Scholar] [CrossRef]
- Ferreiro, C.B.; Martínez, M.L.; Borrás, P.N.; Hernando, M.H.; Salvador, A. Short-term high CO2 treatment alleviates chilling injury of persimmon cv. Fuyu by preserving the parenchyma structure. Food Control. 2015, 51, 163–170. [Google Scholar]
- Giraldo, E.; Díaz, A.; Corral, J.M.; García, A. Applicability of 2-DE to assess differences in the protein profile between cold storage and not cold storage in nectarine fruits. J. Proteom. 2012, 75, 5774–5782. [Google Scholar] [CrossRef]
- Moradinezhad, F.; Khayyat, M.; Saeb, H. Combination effects of postharvest treatments and modified atmosphere packaging on shelf life and quality of Iranian pomegranate fruit cv. Sheshi-kab. Int. J. Postharvest Biol. Technol. Innov. 2013, 3, 244–256. [Google Scholar] [CrossRef]
- Dziedzic, E.; Baszczyk, J.; Bieniasz, M.; Dziadek, K.; Kope, A. Effect of modified (MAP) and controlled atmosphere (CA) storage on the quality and bioactive compounds of blue honeysuckle fruits (Lonicera caerulea L.). Sci. Hortic. 2020, 265, 109226. [Google Scholar] [CrossRef]
- Wang, Y.; Sugar, D. Internal browning disorder and fruit quality in modified atmosphere packaged ‘Bartlett’ pears during storage and transit. Postharvest Biol. Technol. 2013, 83, 72–82. [Google Scholar] [CrossRef]
- Burdon, J.; Billing, D.; Pidakala, P. Avoiding Chilling Damage in ‘Hass’ Avocado Fruit by Controlled Atmosphere Storage at Higher Temperature. Hortscience 2017, 52, 2327–9834. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, E.; Barrios, S.; Lema, P. Ready-to-eat cherry tomatoes: Passive modified atmosphere packaging conditions for shelf life extension. Food Packag. Shelf. 2019, 22, 100407. [Google Scholar] [CrossRef]
- Fernández-León, M.; Fernández-León, A.; Lozano, M.; Ayuso, M.C.; González-Gómez, D. Different postharvest strategies to preserve broccoli quality during storage and shelf life: Controlled atmosphere and 1-MCP. Food Chem. 2013, 138, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Youssef, S.E.; Mitcham, E.J. 272 Control of Brown Rot of Peaches and Apricots with Hot Water and Controlled-Atmosphere Storage. Hortscience 1994, 29, 2327–9834. [Google Scholar] [CrossRef] [Green Version]
- Anastasia, K.; Fabrizio, C.; Claudiu, S. Phenols and Polyphenols as Carbonic Anhydrase Inhibitors. Molecules 2016, 21, 121649. [Google Scholar]
- Zhang, X.; Zhou, Y.; Li, J.; Gu, X.; Zhao, L.; Li, B.; Wang, K.; Yang, Q.; Zhang, H. Pichia caribbica improves disease resistance of cherry tomatoes by regulating ROS metabolism. Biol. Control. 2022, 169, 104870. [Google Scholar] [CrossRef]
- Mokrani, A.; Krisa, S.; Cluzet, S.; Da Costa, G.; Temsamani, H.; Renouf, E.; Mérillon, J.; Madani, K.; Mesnil, M.; Monvoisin, A.; et al. Phenolic contents and bioactive potential of peach fruit extracts. Food Chem. 2016, 202, 212–220. [Google Scholar] [CrossRef]
- Ali, S.; Khan, A.S.; Malik, A.U.; Shahid, M. Effect of controlled atmosphere storage on pericarp browning, bioactive compounds and antioxidant enzymes of litchi fruits. Food Chem. 2016, 206, 18–29. [Google Scholar] [CrossRef]
- Brackmann, A.; Bortoluzzi, G. Influence of temperature, CA conditions and relative humidity on physiological disorders of ‘fuji’ apples. Acta Hortic. 1998, 464, 369–373. [Google Scholar] [CrossRef]
- Selcuk, N.; Erkan, M. The effects of modified and palliflex controlled atmosphere storage on postharvest quality and composition of ‘Istanbul’ medlar fruit. Postharvest Biol. Technol. 2015, 99, 9–19. [Google Scholar] [CrossRef]
- Li, L. Elevated CO2 Enhanced the Antioxidant Activity and Downregulated Cell Wall Metabolism of Wolfberry (Lycium barbarum L.). Antioxidants 2022, 11, 11010016. [Google Scholar]
- Foucher, L.; Barroca, M.J.; Dulyanska, Y.; Correia, P.M.R.; Guiné, R.P.F. Development of Innovative Candied Chestnuts from Three Chestnut Cultivars Grown in Portugal. Foods 2022, 11, 11070917. [Google Scholar] [CrossRef]
- Gull, A.; Bhat, N.; Wani, S.M.; Masoodi, F.A.; Ganai, S.A. Shelf life extension of apricot fruit by application of nanochitosan emulsion coatings containing pomegranate peel extract. Food Chem. 2021, 349, 129149. [Google Scholar] [CrossRef]
- Vegara, S.; Marti, N.; Lorente, J.; Coll, L.; Streitenberger, S.; Valero, M.; Saura, D. Chemical guide parameters for Punica granatum cv. ‘Mollar’ fruit juices processed at industrial scale. Food Chem. 2014, 147, 203–208. [Google Scholar] [CrossRef]
- Kubola, J.; Siriamornpun, S. Phenolic contents and antioxidant activities of bitter gourd (Momordica charantia L.) leaf, stem and fruit fraction extracts in vitro. Food Chem. 2008, 110, 881–890. [Google Scholar] [CrossRef]
- Maroga, G.M.; Soundy, P.; Sivakumar, D. Different Postharvest Responses of Fresh-Cut Sweet Peppers Related to Quality and Antioxidant and Phenylalanine Ammonia Lyase Activities during Exposure to Light-Emitting Diode Treatments. Foods 2019, 8, 8090359. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Zhang, J.; Chen, S.; Li, Y.; Lei, X.; Qiao, H.; Zhu, Q.; Hu, B.; Zhou, Q.; Jiao, J. Conversion of Fibroblasts to Parvalbumin Neurons by One Transcription Factor, Ascl1, and the Chemical Compound Forskolin. J. Biol. Chem. 2016, 291, 13560–13570. [Google Scholar] [CrossRef] [Green Version]
- Kou, L.; Yang, T.; Luo, Y.; Liu, X.; Huang, L.; Coding, E. Pre-harvest calcium application increases biomass and delays senescence of broccoli microgreens. Postharvest Biol. Technol. 2014, 87, 70–78. [Google Scholar] [CrossRef]
- Xie, Y.; Zheng, Y.; Da, I.X.; Wang, Q.; Cao, J.; Xiao, J. Seasonal dynamics of total flavonoid contents and antioxidant activity of Dryopteris erythrosora. Food Chem. 2015, 186, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Radunic, M.; Spika, M.J.; Ban, S.G.; Gadze, J.; Diaz-Perez, J.C.; Dan, M.L. Physical and chemical properties of pomegranate fruit accessions from Croatia. Food Chem. 2015, 177, 53–60. [Google Scholar] [CrossRef]
- Tounsi, M.S.; Wannes, W.A.; Ouerghemmi, I.; Jegham, S.; Njima, Y.B.; Hamdaoui, G.; Zemni, H.; Marzouk, B. Juice components and antioxidant capacity of four Tunisian Citrus varieties. J. Sci. Food Agric. 2011, 91, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T. Effect of alginate coating on physicochemical and sensory qualities of button mushrooms (Agaricus bisporus) under a high oxygen modified atmosphere. Postharvest Biol. Technol. 2013, 76, 91–97. [Google Scholar] [CrossRef]
- Grace, M.H.; Xiong, J.; Esposito, D.; Ehlenfeldt, M. LMA Simultaneous LC-MS quantification of anthocyanins and non-anthocyanin phenolics from blueberries with widely divergent profiles and biological activities. Food Chem. 2019, 3, S0046. [Google Scholar]
- Yuan, Y.; Xu, X.; Jing, C.; Zou, P.; Zhang, C.; Li, Y. Microwave assisted hydrothermal extraction of polysaccharides from Ulva prolifera: Functional properties and bioactivities. Carbohydr. Polym. 2018, 181, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lin, S.; Jia, Y.; Rehman, F.; Wang, Y. Anthocyanin and spermidine derivative hexoses coordinately increase in the ripening fruit of Lycium ruthenicum. Food Chem. 2019, 311, 125874. [Google Scholar] [CrossRef]
- Tareen, M.J.; Singh, Z.; Khan, A.S.; Abbasi, N.A.; Naveed, M. Combined Applications of Aminoethoxyvinylglycine with Salicylic Acid or Nitric Oxide Reduce Oxidative Stress in Peach During Ripening and Cold Storage. J. Plant Growth Regul. 2017, 36, 83–994. [Google Scholar] [CrossRef]
- Smirnoff, N. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 2010, 125, 27–58. [Google Scholar] [CrossRef]
- Sun, J.; Lin, H.; Zhang, S.; Lin, Y.; Wang, H.; Lin, M.; Hung, Y.C.; Chen, Y. The roles of ROS production-scavenging system in Lasiodiplodia theobromae (Pat.) Griff. & Maubl.-induced pericarp browning and disease development of harvested longan fruit. Food Chem. 2018, 247, 16–22. [Google Scholar]
- Bolanos, E.N.A.; Velazquez, R.C.; Cardenaz, A.V.; Santamaria, I.R.; Vera, N.G.; Fuentes, A.D.H.; Silva, E.M. Effect of Stroage Temperatur and Time on Quality in Minimally Processed Litchi Fruit (Litchi Chinensis Sonn.). J. Food Qual. 2010, 33, 299–311. [Google Scholar] [CrossRef]
- Palmaorozco, G.; Sampedro, J.G.; Ortizmoreno, A.; Najera, H. Inactivation of Polyphenol Oxidase in Mamey Fruit. J. Food Sci. 2012, 77, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.; Wood, R. Pectic Enzymes and Phenolic Substances in Apples Rotted by Fungi. Ann. Bot. 1961, 25, 435–452. [Google Scholar] [CrossRef]
- Babbar, N.; Oberoi, H.S.; Uppal, D.S.; Patil, R.T. Total phenolic content and antioxidant capacity of extracts obtained from six important fruit residues. Food Res. Int. 2011, 44, 391–396. [Google Scholar] [CrossRef]
- Alothman, M.; Bhat, R.; Karim, A.A. Antioxidant capacity and phenolic content of selected tropical fruits from Malaysia, extracted with different solvents. Food Chem. 2009, 115, 785–788. [Google Scholar] [CrossRef]
- Silva, R. Enzyme technology in food preservation: A promising and sustainable strategy for biocontrol of post-harvest fungal pathogens. Food Chem. 2018, 277, 531–532. [Google Scholar] [CrossRef]
- Li, D.; Zhang, X.; Qu, H.; Li, L.; Mao, B.; Xu, Y.; Lin, X.; Luo, Z. Delaying the biosynthesis of aromatic secondary metabolites in postharvest strawberry fruit exposed to elevated CO2 atmosphere-ScienceDirect. Food Chem. 2020, 306, 125611. [Google Scholar] [CrossRef]
- Shellie, K.C. Ultra-low oxygen refrigerated storage of ‘rio red’ grapefruit: Fungistatic activity and fruit quality. Postharvest Biol. Technol. 2002, 25, 73–85. [Google Scholar] [CrossRef]
- Kim, Y.; Brecht, J.K.; Talcott, S.T. Antioxidant phytochemical and fruit quality changes in mango (Mangifera indica L.) following hot water immersion and controlled atmosphere storage. Food Chem. 2007, 105, 1327–1334. [Google Scholar] [CrossRef]
- Dlmanal, T. Controlled atmosphere and shelf life performance of a new late-maturing Japanese pear, Pyrus pyrifolia (Burm. F.) Nakai, cultivar ‘Atago’. Not. Bot. Horti Agrobot. Cluj Napoca 2020, 48, 177–188. [Google Scholar]
- Gonzalez, M.E.; Barrett, D.M. Thermal, High Pressure, and Electric Field Processing Effects on Plant Cell Membrane Integrity and Relevance to Fruit and Vegetable Quality. J. Food Sci. 2010, 75, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagpala, E.; Noferini, M.; Farneti, B.; Piccinini, L.; Costa, G. Cherry-Meter: An innovative non-destructive (vis/NIR) device for cherry fruit ripening and quality assessment. Acta Hortic. 2017, 1161, 491–496. [Google Scholar] [CrossRef]
- Mattheis, J.P. Controlled Atmosphere pO2 Alters Ripening Dynamics of 1-MCP treated ‘d’Anjou’ Pear (Pyrus communis L.) Fruit. Hortscience 2017, 52, 2327–9834. [Google Scholar] [CrossRef]
- Nakano, K.; Maezawa, S. Combined effect of heat treatment and modified atmosphere packaging on the color development of cherry tomato. Postharvest Biol. Technol. 2004, 34, 113–116. [Google Scholar]
- Lv, P.; Na, L.; Liu, H.; Gu, H.; Zhao, W.E. Changes in carotenoid profiles and in the expression pattern of the genes in carotenoid metabolisms during fruit development and ripening in four watermelon cultivars. Food Chem. 2015, 174, 52–59. [Google Scholar] [CrossRef]
- Lufu, R.; Ambaw, A.; Opara, U.L. Water loss of fresh fruit: Influencing pre-harvest, harvest and postharvest factors. Sci. Hortic. 2020, 272, 109519. [Google Scholar] [CrossRef]
- Jiang, X.; Lin, H.; Shi, J.; Neethirajan, S.; Lin, Y.; Chen, Y.; Wang, H.; Lin, Y. Effects of a novel chitosan formulation treatment on quality attributes and storage behavior of harvested litchi fruit. Food Chem. 2018, 252, 134–141. [Google Scholar] [CrossRef]
- Li, S.; Jiang, H.; Wang, Y.; Liang, L.; Bi, Y. Effect of benzothiadiazole treatment on improving the mitochondrial energy metabolism involved in induced resistance of apple fruit during postharvest storage. Food Chem. 2019, 302, 125288. [Google Scholar] [CrossRef]
- Ma, Q. Moderation of respiratory cascades and energy metabolism of fresh-cut pear fruit in response to high CO2 controlled atmosphere. Postharvest Biol. Technol. 2021, 172, 111379. [Google Scholar]
- Saquet, A.A.; Streif, J. Respiration rate and ethylene metabolism of ‘Jonagold’ apple and ‘Conference’ pear under regular air and controlled atmosphere. Bragantla 2017, 76, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.A.; Verboven, P.; Schenk, A.; Nicola, B.M. Prediction of water loss from pears (Pyrus communis cv. Conference) during controlled atmosphere storage as affected by relative humidity. J. Food Eng. 2007, 83, 149–155. [Google Scholar] [CrossRef]
- Mcdonald, B.; Harman, J.E. Controlled-atmosphere storage of kiwifruit. I. Effect on fruit firmness and storage life. Sci. Hortic. 1982, 17, 113–123. [Google Scholar] [CrossRef]
- Luo, Z.; Xiang, W.; Yan, X.; Chen, C. Alleviation of chilling injury and browning of postharvest bamboo shoot by salicylic acid treatment. Food Chem. 2012, 131, 456–461. [Google Scholar] [CrossRef]
- Luna, M.C.; Tudela, J.A.; Tomás-Barberán, F.; Gil, M.I. Modified atmosphere (MA) prevents browning of fresh-cut romaine lettuce through multi-target effects related to phenolic metabolism. Postharvest Biol. Technol. 2016, 119, 84–93. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, B.; Zhang, W.; Cao, J.; Jiang, W. Enhancement of quality and antioxidant metabolism of sweet cherry fruit by near-freezing temperature storage. Postharvest Biol. Technol. 2019, 147, 113–122. [Google Scholar] [CrossRef]
- Kaewsuksaeng, S.; Uthairatanakij, A.; Kanlayanarat, S. High O2 effects on peel browning and phenolics changes in longan (Dimocarpus longan Lour.) fruit. Acta Hortic. 2010, 857, 389–394. [Google Scholar] [CrossRef]
- Guo, C.; Bi, J.; Li, X.; Jian, L.; Hu, J. Investigation on the phenolic composition, related oxidation and antioxidant activity of thinned peach dried by different methods. LWT Food Sci. Tech. Braz. 2021, 147, 111573. [Google Scholar] [CrossRef]
- Ma, Y.; Ban, Q.; Shi, J.; Dong, T.; Jiang, C.Z.; Wang, Q. 1-Methylcyclopropene (1-MCP), storage time, and shelf life and temperature affect phenolic compounds and antioxidant activity of ‘Jonagold’ apple. Postharvest Biol. Technol. 2019, 150, 71–79. [Google Scholar] [CrossRef]
- Senter, S.D.; Callahan, A. Variability in the Quantities of Condensed Tannins and Other Major Phenols in Peach Fruit During Maturation. J. Food Sci. 2010, 55, 1585–1587. [Google Scholar] [CrossRef]
- Singh, B.K.; Sharma, S.R.; Singh, B. Antioxidant enzymes in cabbage: Variability and inheritance of superoxide dismutase, peroxidase and catalase. Sci. Hortic. 2010, 124, 9–13. [Google Scholar] [CrossRef]
- Yu, C.; Jian, S.; Xu, X.; Bao, Y.; Jiang, Y. Variations in contents of (−) -epicatechin and activities of phenylalanine ammonialyase and polyphenol oxidase of longan fruit during development. Sci. Hortic. 2010, 125, 230–232. [Google Scholar] [CrossRef]
- Wang, S.Y.; Bunce, J.A.; Maas, J.L. Elevated carbon dioxide increases contents of antioxidant compounds in field-grown strawberries. J. Agric. Food Chem. 2003, 51, 4315–4320. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.Q.; Zhang, M.; Xie, K.D.; Fan, Y.J.; Song, X.; Wang, R.; Wu, X.M.; Zhang, H.Y.; Guo, W. Polyploidy remodels fruit metabolism by modifying carbon source utilization and metabolic flux in Ponkan mandarin (Citrus reticulata Blanco). Plant Sci. 2019, 289, 110276. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Liu, Z.; Zhang, X.; Li, H.; Wang, L. Effect of pulsed controlled atmosphere with CO2 on the quality of watercored apple during storage. Sci. Hortic. 2020, 278, 109854. [Google Scholar] [CrossRef]
- Buccheri, M.; Picchi, V.; Grassi, M.; Bianchi, G.; Scalzo, R.L. Dynamic changes of antioxidants and fermentative metabolites in apple peel in relation to storage, controlled atmosphere, and initial low oxygen stress. Sci. Hortic. 2021, 288, 110312. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, X.; He, Y.; Yuan, C.; Cheng, X.; Li, G.; Shan, Y.; Zhu, X. Controlled Atmosphere Improves the Quality, Antioxidant Activity and Phenolic Content of Yellow Peach during the Shelf Life. Antioxidants 2022, 11, 2278. https://doi.org/10.3390/antiox11112278
Dong X, He Y, Yuan C, Cheng X, Li G, Shan Y, Zhu X. Controlled Atmosphere Improves the Quality, Antioxidant Activity and Phenolic Content of Yellow Peach during the Shelf Life. Antioxidants. 2022; 11(11):2278. https://doi.org/10.3390/antiox11112278
Chicago/Turabian StyleDong, Xinrui, Yi He, Chushan Yuan, Xiaomei Cheng, Gaoyang Li, Yang Shan, and Xiangrong Zhu. 2022. "Controlled Atmosphere Improves the Quality, Antioxidant Activity and Phenolic Content of Yellow Peach during the Shelf Life" Antioxidants 11, no. 11: 2278. https://doi.org/10.3390/antiox11112278