


Ameliorative Effect of Tocotrienols on Perimenopausal-Associated Osteoporosis—A Review

 ,
,
Abstract
:1. Introduction
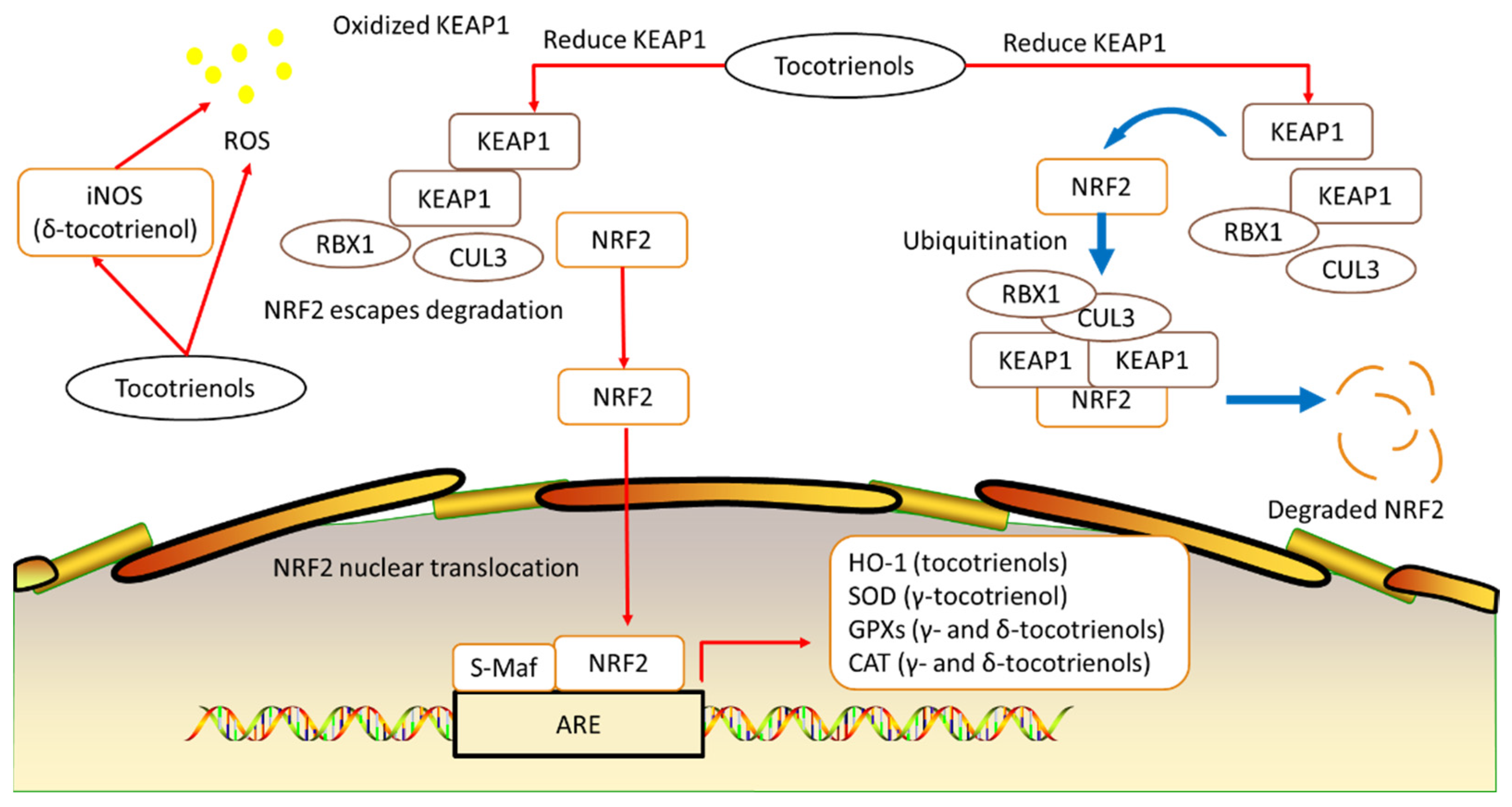
2. Tocotrienols’ Antioxidative Effects in Alleviating Osteoporosis
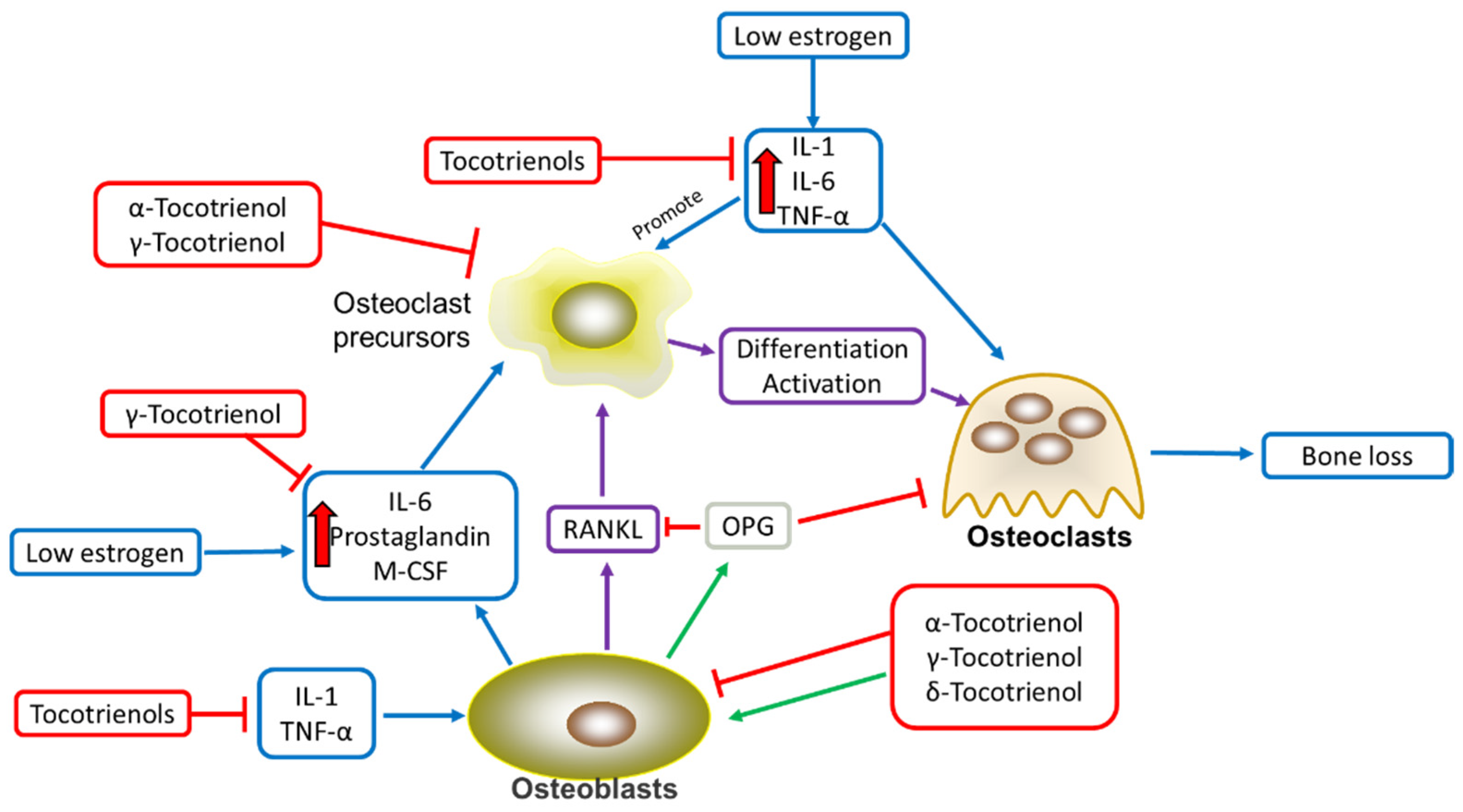
3. Tocotrienols’ Anti-Inflammatory Effects in Alleviating Osteoporosis
4. Inflammatory Mediators and Reactive Oxygen Species (ROS)
5. Improving Tocotrienols’ Bioavailability for the Future of Anti-Osteoporotic Use
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christiansen, C. Consensus development conference: Diagnosis, prophylaxis, and treatment of osteoporosis. Am. J. Med. 1993, 94, 646–650. [Google Scholar] [CrossRef]
- Monteleone, P.; Mascagni, G.; Giannini, A.; Genazzani, A.R.; Simoncini, T. Symptoms of menopause—Global prevalence, physiology and implications. Nat. Rev. Endocrinol. 2018, 14, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; Cooper, C.; Rizzoli, R.; Reginster, J.-Y. European Guidance for the Diagnosis and Management of Osteoporosis in Postmenopausal Women. Osteoporos. Int. 2019, 30, 3–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, J.C.; Burnett-Bowie, S.-A.M.; Finkelstein, J.S. Bone and the Perimenopause. Obstet. Gynecol. Clin. N. Am. 2011, 38, 503–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meganathan, P.; Fu, J.-Y. Biological Properties of Tocotrienols: Evidence in Human Studies. Int. J. Mol. Sci. 2016, 17, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranasinghe, R.; Mathai, M.; Zulli, A. Revisiting the therapeutic potential of tocotrienol. BioFactors 2022, 48, 813–856. [Google Scholar] [CrossRef]
- Wong, R.S.; Radhakrishnan, A.K. Tocotrienol research: Past into present. Nutr. Rev. 2012, 70, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Peh, H.Y.; Tan, W.D.; Liao, W.; Wong, W.F. Vitamin E therapy beyond cancer: Tocopherol versus tocotrienol. Pharmacol. Ther. 2016, 162, 152–169. [Google Scholar] [CrossRef]
- Jiang, Q.; Christen, S.; Shigenaga, M.K.; Ames, B.N. γ-Tocopherol, the major form of vitamin E in the US diet, deserves more attention. Am. J. Clin. Nutr. 2001, 74, 714–722. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.A.; Salser, W.A.; Parmar, R.; Emeson, E.E. Novel Tocotrienols of Rice Bran Inhibit Atherosclerotic Lesions in C57BL/6 ApoE-Deficient Mice. J. Nutr. 2001, 131, 2606–2618. [Google Scholar] [CrossRef]
- Khanna, S.; Roy, S.; Slivka, A.; Craft, T.K.S.; Chaki, S.; Rink, C.; Notestine, M.A.; Devries, A.C.; Parinandi, N.L.; Sen, C.K. Neuroprotective Properties of the Natural Vitamin E α-Tocotrienol. Stroke 2005, 36, 2258–2264. [Google Scholar] [CrossRef] [PubMed]
- Serbinova, E.; Kagan, V.; Han, D.; Packer, L. Free radical recycling and intramembrane mobility in the antioxidant properties of alpha-tocopherol and alpha-tocotrienol. Free Radic. Biol. Med. 1991, 10, 263–275. [Google Scholar] [CrossRef]
- Packer, L.; Weber, S.U.; Rimbach, G. Molecular Aspects of α-Tocotrienol Antioxidant Action and Cell Signalling. J. Nutr. 2001, 131, 369S–373S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, T.C.; Elangovan, S.; Wu, J.M. Differential suppression of proliferation in MCF-7 and MDA-MB-231 breast cancer cells exposed to alpha-, gamma- and delta-tocotrienols is accompanied by altered expression of oxidative stress modulatory enzymes. Anticancer Res. 2010, 30, 4169–4176. [Google Scholar] [PubMed]
- Ibrahim, N.; Noor, H.M.; Shuid, A.N.; Mohamad, S.; Malik, M.M.A.; Jayusman, P.A.; Mohamed, I.N. Osteoprotective Effects in Postmenopausal Osteoporosis Rat Model: Oral Tocotrienol vs. Intraosseous Injection of Tocotrienol-Poly Lactic-Co-Glycolic Acid Combination. Front. Pharmacol. 2021, 12, 706747. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.-L.; Klein, A.; Chin, K.-Y.; Mo, H.; Tsai, P.; Yang, R.-S.; Chyu, M.-C.; Ima-Nirwana, S. Tocotrienols for bone health: A translational approach. Ann. N. Y. Acad. Sci. 2017, 1401, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.K.; Kamisah, Y.; Mohamed, N.; Muhammad, N.; Masbah, N.; Fahami, N.A.M.; Mohamed, I.N.; Shuid, A.N.; Saad, Q.M.; Abdullah, A.; et al. Potential Role of Tocotrienols on Non-Communicable Diseases: A Review of Current Evidence. Nutrients 2020, 12, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervellati, C.; Romani, A.; Cremonini, E.; Bergamini, C.M.; Fila, E.; Squerzanti, M.; Greco, P.; Massari, L.; Bonaccorsi, G. Higher Urinary Levels of 8-Hydroxy-2′-deoxyguanosine Are Associated with a Worse RANKL/OPG Ratio in Postmenopausal Women with Osteopenia. Oxid. Med. Cell. Longev. 2016, 2016, 6038798. [Google Scholar] [CrossRef] [Green Version]
- Manan, N.A.; Mohamed, N.; Shuid, A.N. Effects of Low-Dose versus High-Dose γ-Tocotrienol on the Bone Cells Exposed to the Hydrogen Peroxide-Induced Oxidative Stress and Apoptosis. Evid.-Based Complement. Altern. Med. 2012, 2012, 680834. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liang, M.; Li, H.; Cai, L.; Yang, L. Rice Protein Exerts Anti-Inflammatory Effect in Growing and Adult Rats via Suppressing NF-κB Pathway. Int. J. Mol. Sci. 2019, 20, 6164. [Google Scholar] [CrossRef]
- Arioz, B.I.; Taştan, B.; Tarakcioglu, E.; Tufekci, K.U.; Olcum, M.; Ersoy, N.; Bagriyanik, A.; Genc, K.; Genc, S. Melatonin Attenuates LPS-Induced Acute Depressive-Like Behaviors and Microglial NLRP3 Inflammasome Activation Through the SIRT1/Nrf2 Pathway. Front. Immunol. 2019, 10, 1511. [Google Scholar] [CrossRef] [PubMed]
- Ke, K.; Sul, O.-J.; Rajasekaran, M.; Choi, H.-S. MicroRNA-183 increases osteoclastogenesis by repressing heme oxygenase-1. Bone 2015, 81, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, A.A.; Reis, J.C.; Papasian, C.J.; Morrison, D.C.; Qureshi, N. Tocotrienols inhibit lipopolysaccharide-induced pro-inflammatory cytokines in macrophages of female mice. Lipids Health Dis. 2010, 9, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theriault, A.; Chao, J.-T.; Gapor, A. Tocotrienol is the most effective vitamin E for reducing endothelial expression of adhesion molecules and adhesion to monocytes. Atherosclerosis 2002, 160, 21–30. [Google Scholar] [CrossRef]
- Naito, Y.; Shimozawa, M.; Kuroda, M.; Nakabe, N.; Manabe, H.; Katada, K.; Kokura, S.; Ichikawa, H.; Yoshida, N.; Noguchi, N.; et al. Tocotrienols reduce 25-hydroxycholesterol-induced monocyte–endothelial cell interaction by inhibiting the surface expression of adhesion molecules. Atherosclerosis 2005, 180, 19–25. [Google Scholar] [CrossRef]
- Noguchi, N.; Hanyu, R.; Nonaka, A.; Okimoto, Y.; Kodama, T. Inhibition of THP-1 cell adhesion to endothelial cells by α-tocopherol and α-tocotrienol is dependent on intracellular concentration of the antioxidants. Free Radic. Biol. Med. 2003, 34, 1614–1620. [Google Scholar] [CrossRef]
- Brooks, R.; Kalia, P.; Ireland, D.C.; Beeton, C.; Rushton, N. Direct Inhibition of Osteoclast Formation and Activity by the Vitamin E Isomer gamma-Tocotrienol. Int. J. Vitam. Nutr. Res. 2011, 81, 358–367. [Google Scholar] [CrossRef]
- Sen, C.K.; Khanna, S.; Rink, C.; Roy, S. Tocotrienols: The Emerging Face of Natural Vitamin E. Vitam. Horm. 2007, 76, 203–261. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, J.; Horowitz, M.; Choi, Y. Osteoimmunology: Interactions of the Bone and Immune System. Endocr. Rev. 2008, 29, 403–440. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Kitaura, H.; Zhou, P.; Ross, F.P.; Teitelbaum, S. IL-1 mediates TNF-induced osteoclastogenesis. J. Clin. Investig. 2005, 115, 282–290. [Google Scholar] [CrossRef]
- Cannon, J.G.; Cortez-Cooper, M.; Meaders, E.; Stallings, J.; Haddow, S.; Kraj, B.; Sloan, G.; Mulloy, A. Follicle-stimulating hormone, interleukin-1, and bone density in adult women. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R790–R798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salamone, L.M.; Whiteside, T.; Friberg, D.; Epstein, R.S.; Kuller, L.H.; Cauley, J.A. Cytokine Production and Bone Mineral Density at the Lumbar Spine and Femoral Neck in Premenopausal Women. Calcif. Tissue Res. 1998, 63, 466–470. [Google Scholar] [CrossRef]
- Hunter, I.; Nixon, G.F. Spatial Compartmentalization of Tumor Necrosis Factor (TNF) Receptor 1-dependent Signaling Pathways in Human Airway Smooth Muscle Cells. Lipid rafts are essential for TNF-alpha-mediated activation of RhoA but dispensable for the activation of the NF-kappaB and MAPK pathways. J. Biol. Chem. 2006, 281, 34705–34715. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Xia, Q.; Wu, Y.; Zhang, X.; Wen, F.; Chen, X.; Zhang, S.; Heng, B.C.; He, Y.; Ouyang, H.-W. 3D-Printed Atsttrin-Incorporated Alginate/Hydroxyapatite Scaffold Promotes Bone Defect Regeneration with TNF/TNFR Signaling Involvement. Adv. Health Mater. 2015, 4, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Massaad, C.A.; Klann, E. Reactive Oxygen Species in the Regulation of Synaptic Plasticity and Memory. Antioxid. Redox Signal. 2011, 14, 2013–2054. [Google Scholar] [CrossRef] [Green Version]
- Mariappan, N.; Elks, C.M.; Fink, B.; Francis, J. TNF-induced mitochondrial damage: A link between mitochondrial complex I activity and left ventricular dysfunction. Free Radic. Biol. Med. 2009, 46, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-κB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol. 2006, 72, 1493–1505. [Google Scholar] [CrossRef]
- Grinberg, Y.Y.; Dibbern, M.E.; Levasseur, V.A.; Kraig, R.P. Insulin-like growth factor-1 abrogates microglial oxidative stress and TNF-α responses to spreading depression. J. Neurochem. 2013, 126, 662–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, D.; Kovacs, P.; Eszlari, N.; Gonda, X.; Juhasz, G. Psychological side effects of immune therapies: Symptoms and pathomechanism. Curr. Opin. Pharmacol. 2016, 29, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Han, L.; Martin-Millan, M.; Plotkin, L.I.; Stewart, S.A.; Roberson, P.K.; Kousteni, S.; O’Brien, C.A.; Bellido, T.; Parfitt, A.M.; et al. Skeletal Involution by Age-associated Oxidative Stress and Its Acceleration by Loss of Sex Steroids. J. Biol. Chem. 2007, 282, 27285–27297. [Google Scholar] [CrossRef]
- Wauquier, F.; Leotoing, L.; Coxam, V.; Guicheux, J.; Wittrant, Y. Oxidative stress in bone remodelling and disease. Trends Mol. Med. 2009, 15, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.-C.; Lu, D.; Bai, J.; Zheng, H.; Ke, Z.-Y.; Li, X.-M.; Luo, S.-Q. Oxidative stress inhibits osteoblastic differentiation of bone cells by ERK and NF-κB. Biochem. Biophys. Res. Commun. 2004, 314, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.S.; Neri, S.; Sciacchitano, S.; Di Pino, L.; Costa, M.P.; Marchese, G.; Celotta, G.; Cassibba, N.; Pennisi, G.; Caschetto, S. Behaviour of some indicators of oxidative stress in postmenopausal and fertile women. Maturitas 2006, 53, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Bonaccorsi, G.; Cremonini, E.; Romani, A.; Fila, E.; Castaldini, M.C.; Ferrazzini, S.; Giganti, M.; Massari, L. Oxidative Stress and Bone Resorption Interplay as a Possible Trigger for Postmenopausal Osteoporosis. BioMed Res. Int. 2014, 2014, 569563. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.A.F.; Tirapeli, K.G.; Chaves-Neto, A.H.; Brasilino, M.D.S.; da Rocha, C.Q.; Belló-Klein, A.; Llesuy, S.F.; Dornelles, R.C.M.; Nakamune, A.C.D.M.S. Ilex paraguariensis supplementation may be an effective nutritional approach to modulate oxidative stress during perimenopause. Exp. Gerontol. 2017, 90, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Gao, J.; Zheng, Y.; Wang, X.; Chen, C.; Cao, K.; Xu, J.; Li, Y.; Lu, W.; Liu, J.; et al. Zeaxanthin induces Nrf2-mediated phase II enzymes in protection of cell death. Cell Death Dis. 2014, 5, e1218. [Google Scholar] [CrossRef] [Green Version]
- Bellanti, F.; Matteo, M.; Rollo, T.; De Rosario, F.; Greco, P.; Vendemiale, G.; Serviddio, G. Sex hormones modulate circulating antioxidant enzymes: Impact of estrogen therapy. Redox Biol. 2013, 1, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Simkova, A.; Bulas, J.; Balogová, S.; Reptova, A.; Kisa, B.; Luha, J.; Kinova, S. Metabolic syndrome and its effect on aortic stiffness in premenopausal women. Bratisl. Lek. List. 2013, 114, 279–282. [Google Scholar] [CrossRef] [Green Version]
- Ogunro, P.S.; Bolarinde, A.A.; Owa, O.O.; Salawu, A.A.; Oshodi, A.A. Antioxidant status and reproductive hormones in women during reproductive, perimenopausal and postmenopausal phase of life. Afr. J. Med. Med. Sci. 2014, 43, 49–57. [Google Scholar]
- Wdowiak, A. Comparing antioxidant enzyme levels in follicular fluid in ICSI-treated patients. Gynecol. Obstet. Fertil. 2015, 43, 515–521. [Google Scholar] [CrossRef]
- Vural, P.; Akgül, C.; Canbaz, M. Effects of menopause and tibolone on antioxidants in postmenopausal women. Ann. Clin. Biochem. 2005, 42 Pt 3, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Bednarek-Tupikowska, G.; Tworowska, U.; Jedrychowska, I.; Radomska, B.; Tupikowski, K.; Bidzinska-Speichert, B.; Milewicz, A. Effects of oestradiol and oestroprogestin on erythrocyte antioxidative enzyme system activity in postmenopausal women. Clin. Endocrinol. 2006, 64, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.-Y.; Che, H.-L.; Tan, D.M.-Y.; Teng, K.-T. Bioavailability of tocotrienols: Evidence in human studies. Nutr. Metab. 2014, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, N.; Khamis, M.F.; Yunoh, M.F.M.; Abdullah, S.; Mohamed, N.; Shuid, A.N. Targeted Delivery of Lovastatin and Tocotrienol to Fracture Site Promotes Fracture Healing in Osteoporosis Model: Micro-Computed Tomography and Biomechanical Evaluation. PLoS ONE 2014, 9, e115595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamad, N.-V.; Ima-Nirwana, S.; Chin, K.-Y. Therapeutic potential of annatto tocotrienol with self-emulsifying drug delivery system in a rat model of postmenopausal bone loss. Biomed. Pharmacother. 2021, 137, 111368. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.-V.; Ima-Nirwana, S.; Chin, K.-Y. Self-emulsified annatto tocotrienol improves bone histomorphometric parameters in a rat model of oestrogen deficiency through suppression of skeletal sclerostin level and RANKL/OPG ratio. Int. J. Med. Sci. 2021, 18, 3665–3673. [Google Scholar] [CrossRef]
- Zainal, Z.; Khaza’Ai, H.; Radhakrishnan, A.K.; Chang, S.K. Therapeutic potential of palm oil vitamin E-derived tocotrienols in inflammation and chronic diseases: Evidence from preclinical and clinical studies. Food Res. Int. 2022, 156, 111175. [Google Scholar] [CrossRef]
- Nazrun, A.S.; Norazlina, M.; Norliza, M.; Nirwana, S.I. The Anti-Inflammatory Role of Vitamin E in Prevention of Osteoporosis. Adv. Pharmacol. Sci. 2012, 2012, 142702. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
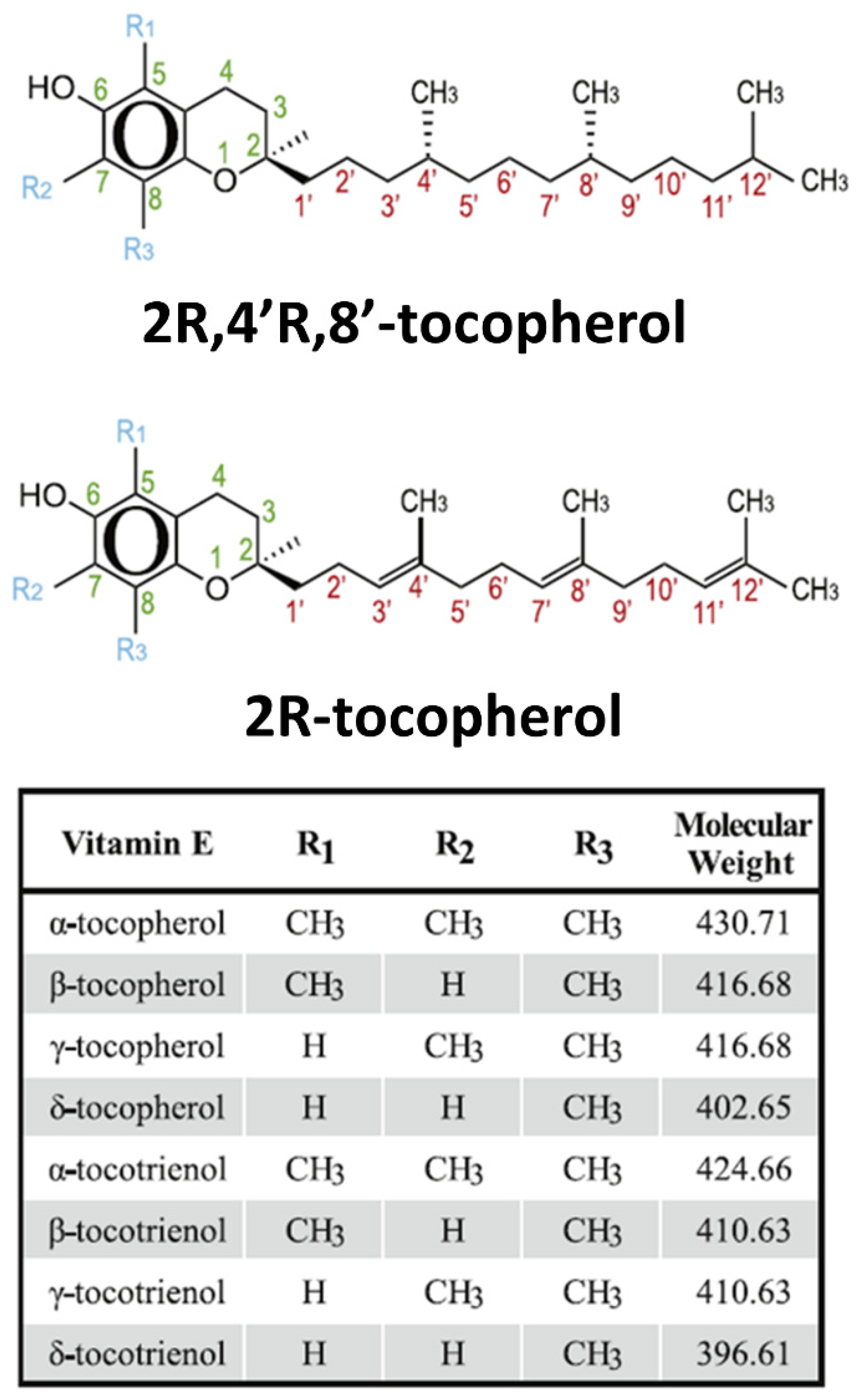
| Property | Disease/Model | Tocotrienol | Combine with | Effect(s) |
|---|---|---|---|---|
| Anti-cancer | Bladder and pancreas cancer | Gamma- and delta-tocotrienols | Gemcitabine | Induced p21 and p27 Suppressed MMP-9, COX-1, Ki-67, Cyclin-D1, Bax, Bcl-1, Bcl-2, Bcl-xL, and Mcl-1 Activated caspase-3, PARP cleavage Downregulated STAT3 and NF-κB/65 pathways |
| Liver cancer | Paclitaxel Doxorubicin | Inhibited NF-κB, JakStat-3/6. STAT-3, 5-LOX-induced eicosanoids, PGE2, and LTB4 | ||
| Brain cancer | Jerantinine A | Induced apoptosis, Fas and p53, and G0/G1 cell cycle arrest | ||
| Gastric and colorectal cancer | Capecitabine | Downregulated survivin, cIAP-1, cIAL-2, Cyclin D1, c-myc, MMP-p, VEGF, ICAM-1, and CXCR4 by inhibiting NFκB/p65 pathway | ||
| Prostate cancer | Docetaxel | Modulated pro-survival Id-1, Id-3, EGF-R, JNK, NF-κB, sphingolipids, eicosanoids, and Ang-1/Tie-2 pathway 2 | ||
| Tocotrienols | Inhibited cell growth in PC3 and LNCaP prostate cancer cells | |||
| Gamma-tocotrienol | Suppressed NF-κB, EGFR, and Id family proteins (Id1 and Id3) | |||
| Oral cancer | Gamma- and delta-tocotrienols | Inhibited docetaxel-induced NF-κB/p65 pathway | ||
| Breast cancer | Activated caspases, EGF-R, and Id-1 and suppressed NF-κB | |||
| Breast cancer | Tamoxifen | Activated estrogen-responsive genes, caspase-3, and apoptosis | ||
| TRF–palm oil | Prevented chemically induced mammary tumorigenesis | |||
| Human breast cancer cells | Tocotrienols | Effective apoptotic inducers | ||
| Mammary cancer cells | Tocotrienols | Induced apoptosis by activating caspase-8 signaling pathway and disrupted mitochondrial function | ||
| Breast cancer cells | Tocotrienols | Induced cell death on proteins with inhibitory effects on cell growth and differentiation MM-1 and interferon-inducible protein 9–27 | ||
| Breast cancer cells | Gamma-tocotrienol | Induced cell death by disrupting Id1, a key cancer-promoting protein | ||
| Cervical cancer | Alpha- and gamma-tocotrienols | Inhibited HeLa cell proliferation by upregulating IL-6 and downregulating cyclin D3, p16, and CDK6 expression in the cell-cycle signaling pathway | ||
| Mesothelioma | Gamma- and delta-tocotrienols | Cisplatin | Suppressed PI3K/Akt pathway | |
| Melanoma | Statin | Inhibited mevalonate pathway Inactivated Ras | ||
| Antioxidant | Primary hippocampal neurons | Gamma-tocotrienol | Provided neuroprotection by upregulating the Bcl-xL family of proteins, which induces anti-apoptosis and maintain optimum neurotransmission | |
| Alpha-tocotrienol | Provided neuroprotection to glutamate-induced hippocampal neurons by preventing the loss of mitochondrial membrane potential | |||
| +SA mammary tumor epithelial cells | Gamma-tocotrienol | Provided anti-apoptotic effects by increasing PARP cleavage and activation-mediated protein kinase-like ER kinase/eukaryotic translational initiation factor/activating transcription factor (PERK/eIF2alpha/ATF-4) pathway, a marker of ER stress response | ||
| Osteoblasts exposed to hydrogen peroxide | Gamma-tocotrienol | Reduced production of malondialdehyde and free radical release and increased antioxidant status in cancerous patients in a dose-dependent manner Prevented the reduction of SOD, catalase, and Gpx activity at lower concentrations | ||
| Experimental diabetic rats | TRF | Increased SOD activity and vitamin C levels, reduced lipid peroxidation in the thoracic aorta homogenates, and reduced vascular smooth muscle cell proliferation and degeneration | ||
| UV-radiation-induced oxidative stress in murine skin | TRF | Inhibited oxidative stress | ||
| Diabetic animals | TRF | Improved wound-healing mechanisms | ||
| High-fat-diet- and streptozotocin-induced diabetes | TRF | Reduced skeletal muscle-related protein levels (4-hydroxynonenal, protein carbonyls, Nrf2, and HO-1) | ||
| Anti-inflammatory | LPS- and SFA-stimulated NLRP-3 inflammasome activation in BMDM in vitro | Gamma-tocotrienol | Repressed NLRP3 inflammasome activation by downregulating arachidonic acid metabolism, diacylglycerol, prostaglandin release, and COX-2 activation | |
| NLRP-3 inflammasome activation in macrophages in vitro | Annatto-seed-derived delta-tocotrienol | Inhibited the priming of the inflammasome by suppressing the NF-κB pathway and ROS production | ||
| Chronic inflammation mimetic type 2-diabetes (leptin receptor knockout mice) | Gamma-tocotrienol | Attenuated the NLRP3-inflammasome activity by downregulating A20-induced TNF-α interacting protein 3-activated NF-κB signaling cascade and suppressed caspase-1 cleavage by AMP-activated protein kinase autophagy | ||
| TRF | Inhibited iNOS and COX-2 production and NF-κB expression | |||
| Cardioprotective (i) Lipid-lowering effects | Delta- and gamma-tocotrienols | Reduced triglyceride synthesis and transport | ||
| Tocotrienols | Reduced synthesis and increased degradation of HMG-CoA reductase Influenced mevalonate pathway in mammalian cells by post-transcriptional suppression of HMG-CoA reductase | |||
| Tocotrienols in chick diet | Lowered hepatic cholesterogenesis, serum total cholesterol, and low-density lipoprotein cholesterol with concomitant increase in lipogenic activity | |||
| (ii) Antiatherogenic effects | Rice bran tocotrienols | Reduced size of atherosclerotic lesions in mice | ||
| Patients with high cholesterol | Tocotrienols | Reversal of arterial blockage of carotid artery | ||
| Atherosclerosis-prone mice | Palm oil tocotrienols | Reduced formation of lesion through antioxidant-dependent and -independent mechanisms | ||
| (iii) Others | Tocotrienols | Provided protection against endothelial dysfunction and platelet aggregation Lowered blood pressure Improved arterial compliance | ||
| Palm oil tocotrienols | Stabilized proteasomes | |||
| Neuroprotective | Palm oil alpha-tocotrienol | Attenuated both enzymatic and nonenzymatic mediators of arachidonic acid metabolism and neurodegeneration | ||
| Alpha-tocotrienol | Reduced size of cerebral infarcts in mice | |||
| Anti-diabetic | Diabetic rats | TRF | Improved blood glucose, dyslipidemia, and oxidative stress | |
| Type 2 diabetes | Beta-tocotrienol | Total vitamin E | Reduced risk of type 2 diabetes mellitus | |
| Streptozotocin-induced diabetes | Tocotrienols-rich diet | Decreased advanced glycosylation end products in nondiabetic rats Improved glycemic control in rats with streptozotocin-induced diabetes | ||
| Anti-osteoporosis | Alpha-tocotrienol | Reduced proinflammatory effect of TNF-α, expression of RANKL in osteoblasts Inhibited the differentiation of IL-1 induced-osteoclast Reduced osteoclast formation | ||
| Gamma-tocotrienol | Inhibited IL-1-mediated PGE2 and PGD2 production Inhibited COX-2 and 5-LOX from transforming arachidonic acid into an inflammatory mediator Increased transcription and expression of SOD, Gpx, and catalase Protected osteoblasts from harmful ROS damages Increased formation of collagen fibrils Increased mineralization of extracellular matrix Increased OPG and reduced RANKL and osteoclast formation | |||
| Delta-tocotrienol | Reduced the expression of TNF-α, IL-1β, and IL-6 Inhibited 5-LOX from transforming arachidonic acid into an inflammatory mediator Reduced the expression of iNOS Increased the expression and activities of Gpxs and catalase Increased formation of collagen fibrils Increased mineralization of extracellular matrix Increased OPG and reduced RANKL and osteoclast formation | |||
| Tocotrienols | Suppressed the synthesis of leukotrienes Reduced the levels of IL-1 and IL-6 Reduced KEAP1, promoting transcription of HO-1, Gpxs, and SOD Increased mRNA expression of osteocalcin, BMP-2, RUNX-2, and VEGF-α of tibia and many protective effects |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, G.; Kow, A.S.F.; Tham, C.L.; Ho, Y.-C.; Lee, M.T. Ameliorative Effect of Tocotrienols on Perimenopausal-Associated Osteoporosis—A Review. Antioxidants 2022, 11, 2179. https://doi.org/10.3390/antiox11112179
Liang G, Kow ASF, Tham CL, Ho Y-C, Lee MT. Ameliorative Effect of Tocotrienols on Perimenopausal-Associated Osteoporosis—A Review. Antioxidants. 2022; 11(11):2179. https://doi.org/10.3390/antiox11112179
Chicago/Turabian StyleLiang, Gengfan, Audrey Siew Foong Kow, Chau Ling Tham, Yu-Cheng Ho, and Ming Tatt Lee. 2022. "Ameliorative Effect of Tocotrienols on Perimenopausal-Associated Osteoporosis—A Review" Antioxidants 11, no. 11: 2179. https://doi.org/10.3390/antiox11112179