

Garlic-Derived Metabolites Exert Antioxidant Activity, Modulate Gut Microbiota Composition and Limit Citrobacter rodentium Infection in Mice
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Garlic Metabolites
2.2. Cell Culture
2.3. Reactive Oxygen Species Assay
2.4. Nitric Oxide Assay
2.5. Western Blot
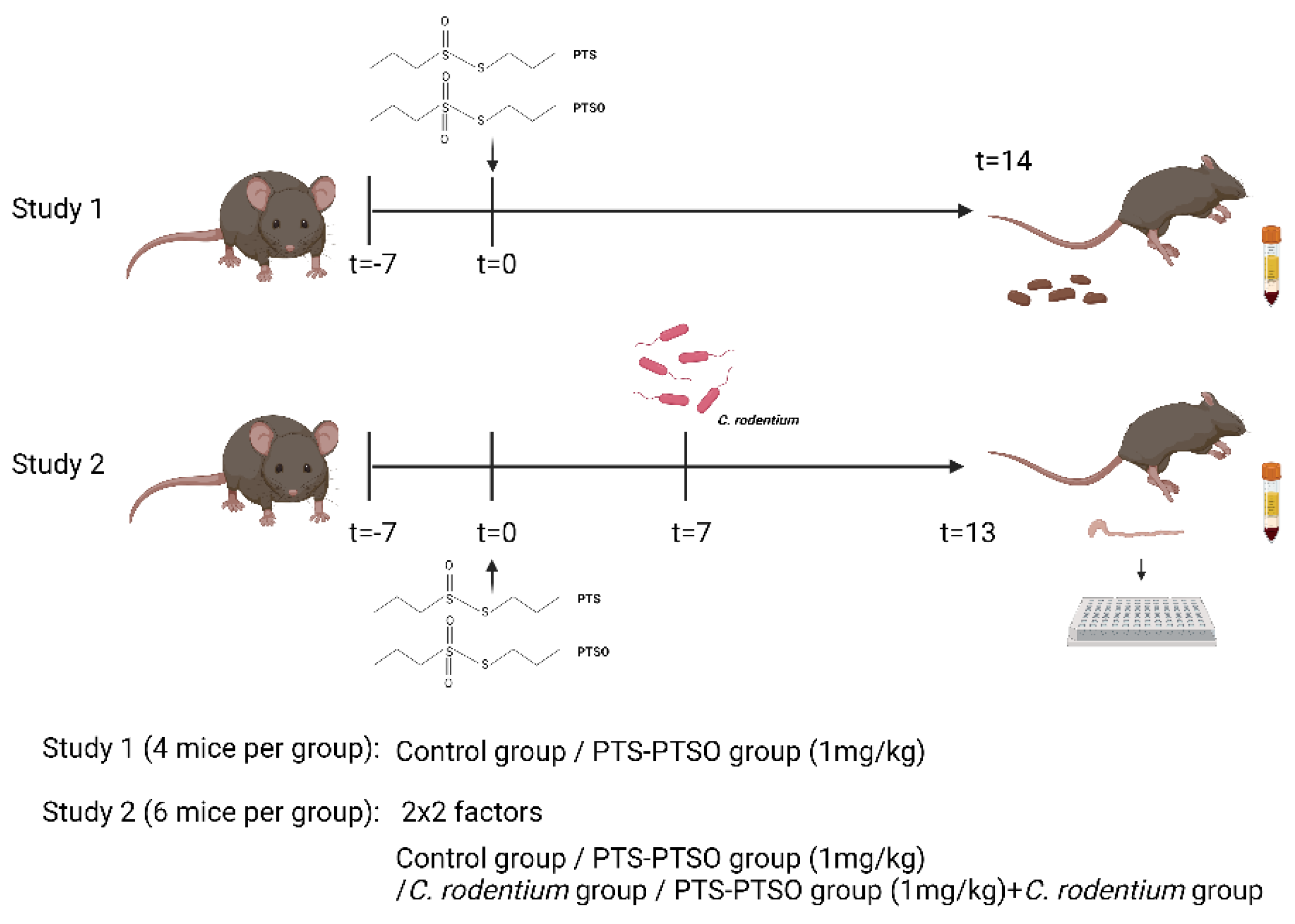
2.6. Animal Experiments
2.7. Faecal Microbiota Analysis
2.8. Total Glutathione Assessment
2.9. Quantitative Real-Time PCR
2.10. Statistical Analysis
3. Results
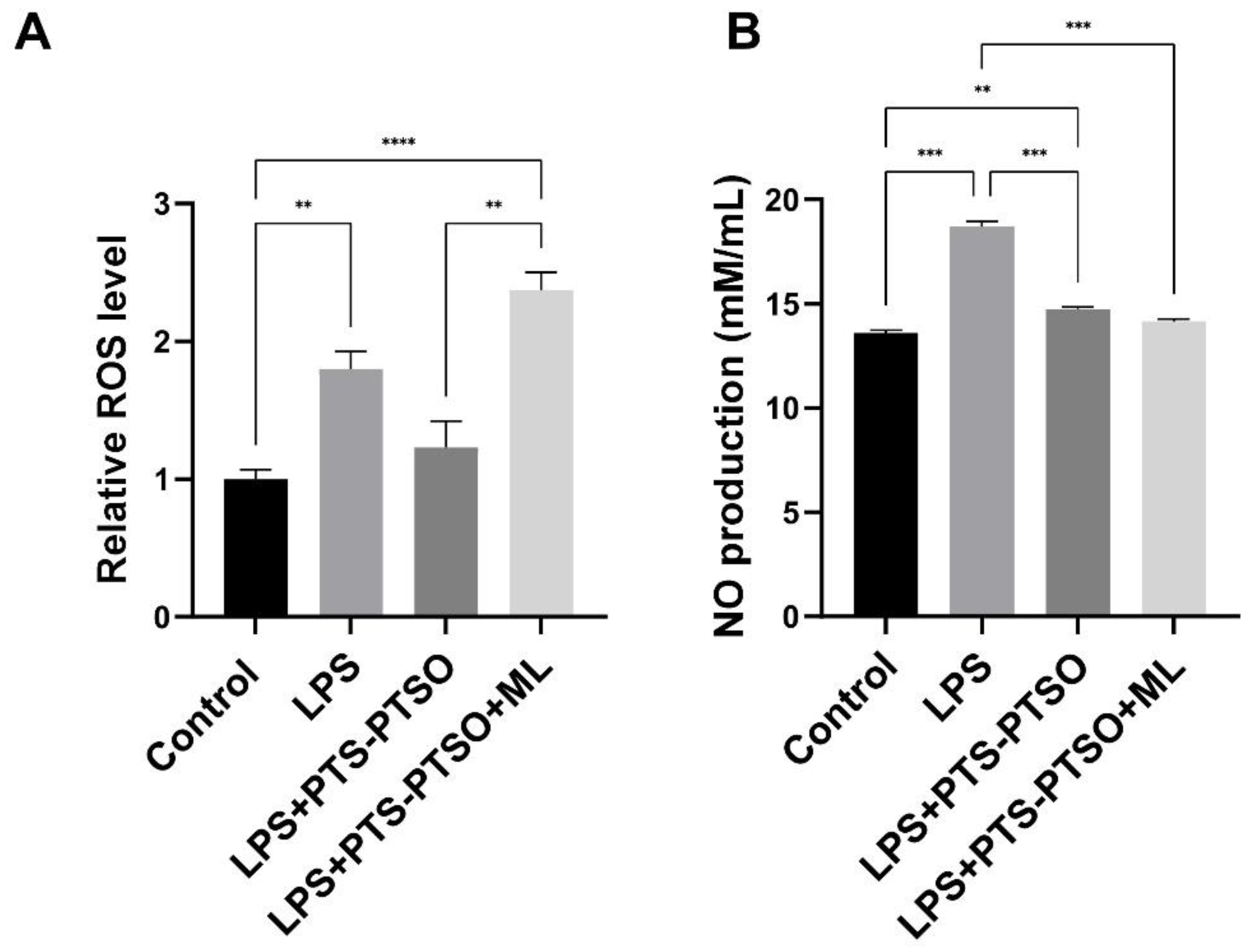
3.1. PTS-PTSO Decreases Reactive Oxygen Species and Nitric Oxide Production in Macrophages
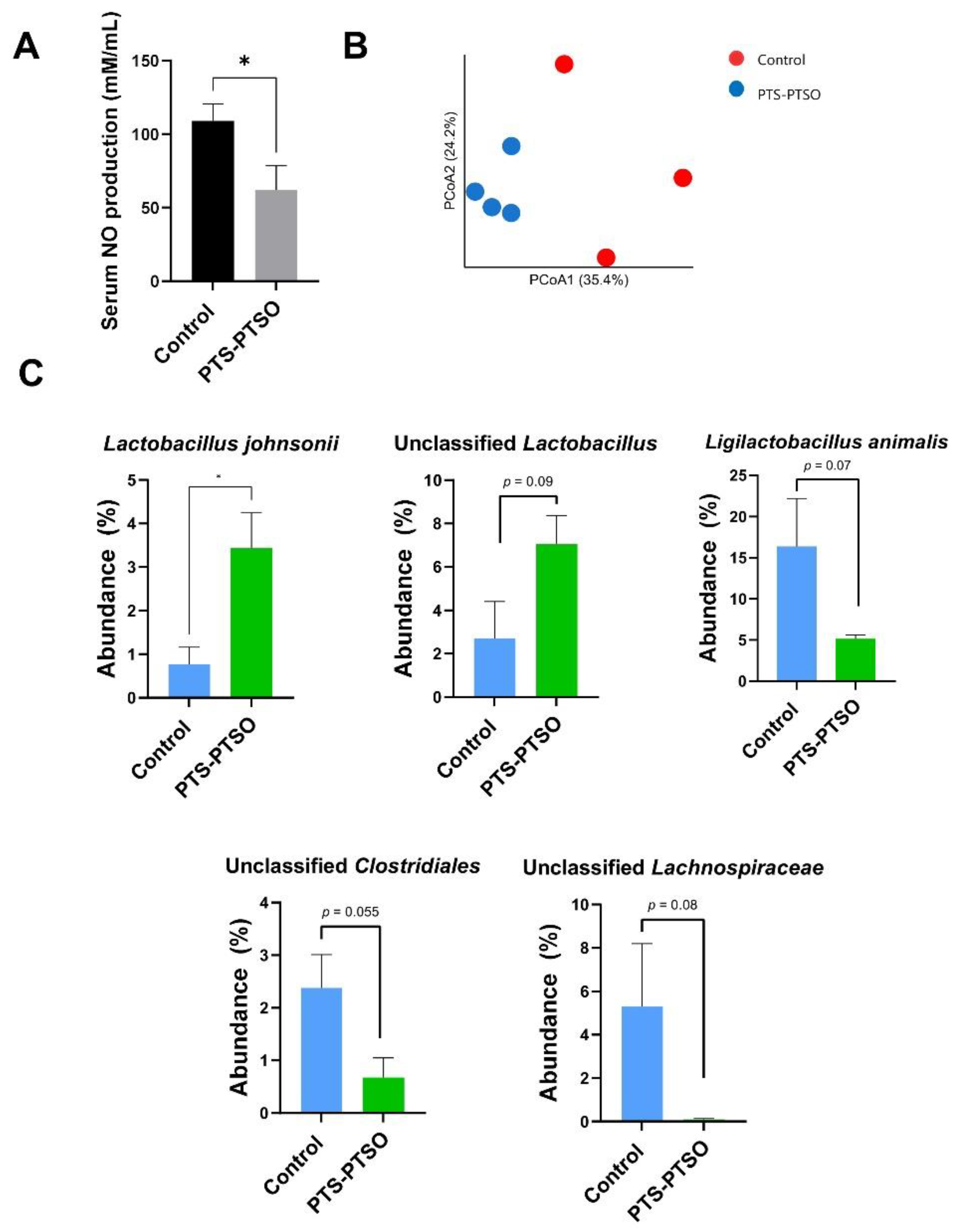
3.2. PTS-PTSO Modulates Serum Nitric Oxide and the Faecal Microbiota Composition in Mice
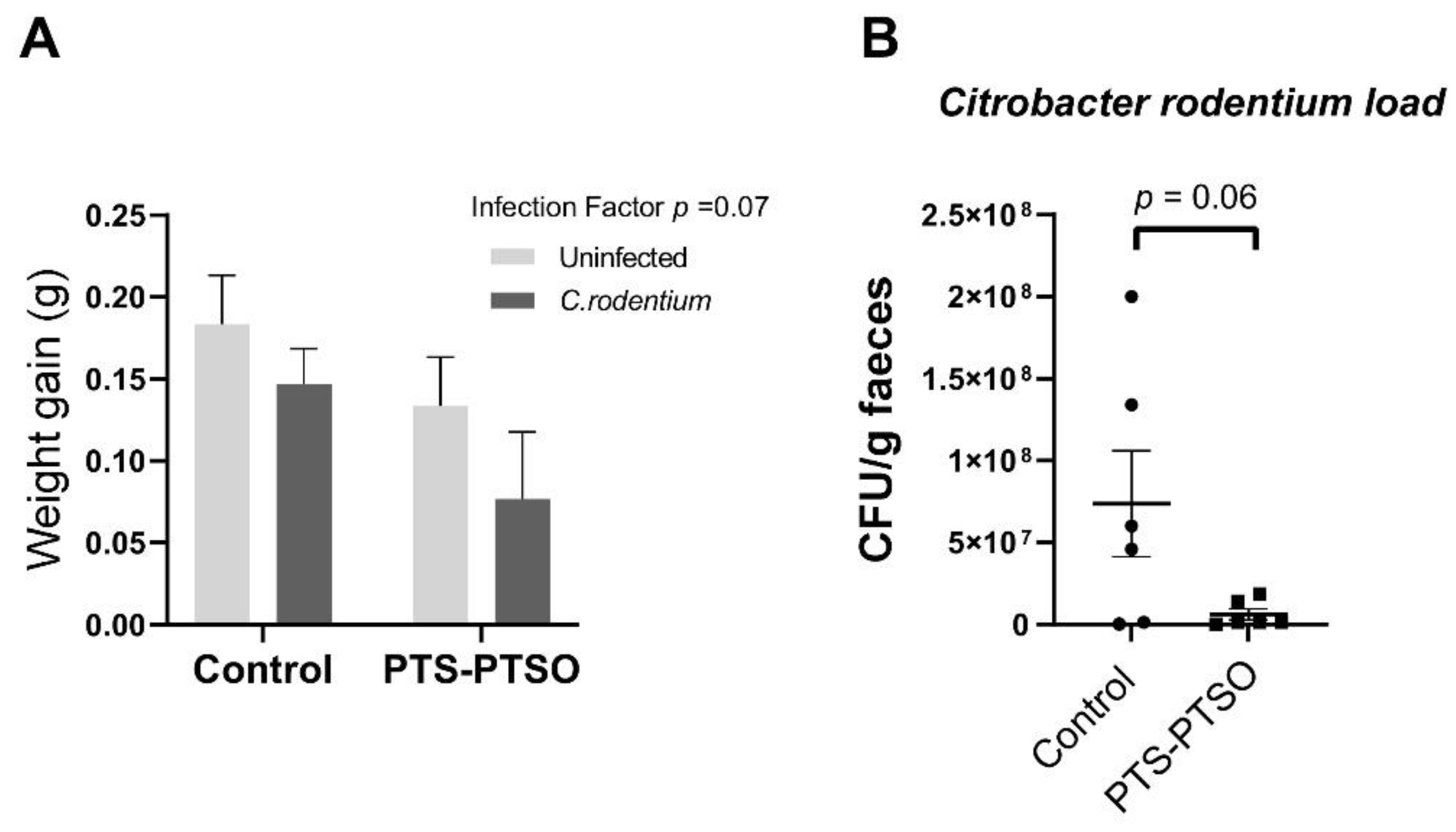
3.3. PTS-PTSO Reduces Faecal Excretion of Citrobacter Rodentium
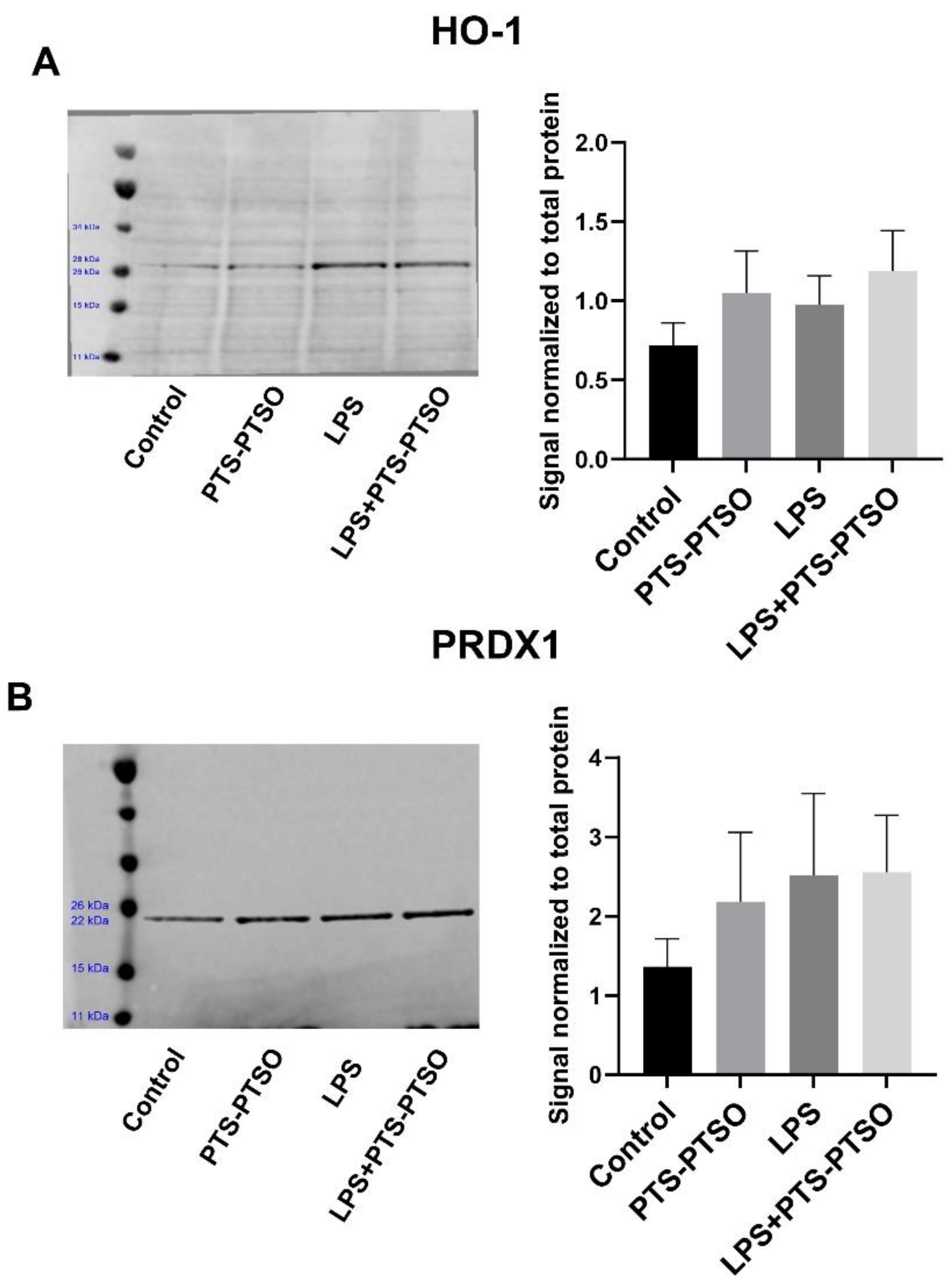
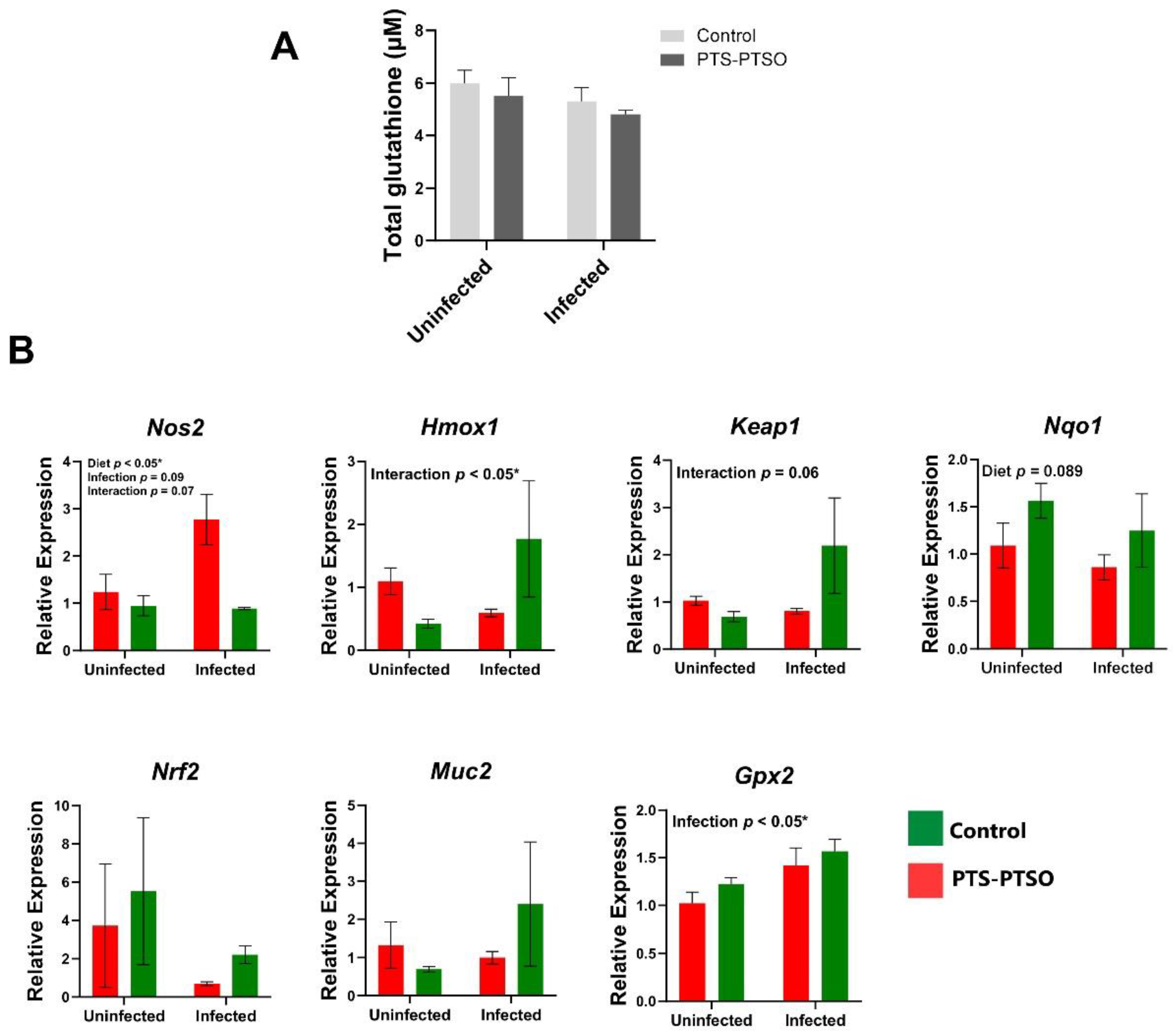
3.4. PTS-PTSO Increases Nrf2-Related Gene Expression in Mouse Colonic Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Francino, M.P. Antibiotics and the Human Gut Microbiome: Dysbioses and Accumulation of Resistances. Front. Microbiol. 2016, 6, 1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silberger, D.J.; Zindl, C.L.; Weaver, C.T. Citrobacter Rodentium: A Model Enteropathogen for Understanding the Interplay of Innate and Adaptive Components of Type 3 Immunity. Mucosal Immunol. 2017, 10, 1108–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, J.W.; Keeney, K.M.; Crepin, V.F.; Rathinam, V.A.K.; Fitzgerald, K.A.; Finlay, B.B.; Frankel, G. Citrobacter Rodentium: Infection, Inflammation and the Microbiota. Nat. Rev. Microbiol. 2014, 12, 612–623. [Google Scholar] [CrossRef]
- Hopkins, E.G.D.; Roumeliotis, T.I.; Mullineaux-Sanders, C.; Choudhary, J.S.; Frankel, G. Intestinal Epithelial Cells and the Microbiome Undergo Swift Reprogramming at the Inception of Colonic Citrobacter Rodentium Infection. MBio 2019, 10, e00062-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.; Piao, M.; Song, Y. Dietary Quercetin Increases Colonic Microbial Diversity and Attenuates Colitis Severity in Citrobacter Rodentium-Infected Mice. Front. Microbiol. 2019, 10, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.; Wall, E.H.; Bravo, D.M.; Hristov, A.N. Host-Mediated Effects of Phytonutrients in Ruminants: A Review. J. Dairy Sci. 2017, 100, 5974–5983. [Google Scholar] [CrossRef] [Green Version]
- Borek, C. Antioxidant Health Effects of Aged Garlic Extract. J. Nutr. 2001, 131, 1010S–1015S. [Google Scholar] [CrossRef] [Green Version]
- Bakri, I.M.; Douglas, C.W.I. Inhibitory Effect of Garlic Extract on Oral Bacteria. Arch. Oral Biol. 2005, 50, 645–651. [Google Scholar] [CrossRef]
- Goncagul, G.; Ayaz, E. Antimicrobial Effect of Garlic (Allium sativum) and Traditional Medicine. J. Anim. Vet. Adv. 2010, 9, 1–4. [Google Scholar] [CrossRef]
- Fufa, B.K. Anti-Bacterial and Anti-Fungal Properties of Garlic Extract (Allium sativum): A Review. Microbiol. Res. J. Int. 2019, 28, 1–5. [Google Scholar] [CrossRef]
- Abad, P.; Arroyo-Manzanares, N.; Rivas-Montoya, E.; Ochando-Pulido, J.M.; Guillamon, E.; García-Campaña, A.M.; Martinez-Ferez, A. Effects of Different Vehiculization Strategies for the Allium Derivative Propyl Propane Thiosulfonate during Dynamic Simulation of the Pig Gastrointestinal Tract. Can. J. Anim. Sci. 2018, 99, 244–253. [Google Scholar] [CrossRef]
- Sorlozano-Puerto, A.; Albertuz-Crespo, M.; Lopez-Machado, I.; Ariza-Romero, J.J.; Baños-Arjona, A.; Exposito-Ruiz, M.; Gutierrez-Fernandez, J. In Vitro Antibacterial Activity of Propyl-Propane-Thiosulfinate and Propyl-Propane-Thiosulfonate Derived from Allium Spp. against Gram-Negative and Gram-Positive Multidrug-Resistant Bacteria Isolated from Human Samples. Biomed Res. Int. 2018, 2018, 7861207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezza, T.; Garrido-Mesa, J.; Diez-Echave, P.; Hidalgo-García, L.; Ruiz-Malagón, A.J.; García, F.; Sánchez, M.; Toral, M.; Romero, M.; Duarte, J. Allium-Derived Compound Propyl Propane Thiosulfonate (PTSO) Attenuates Metabolic Alterations in Mice Fed a High-Fat Diet through Its Anti-Inflammatory and Prebiotic Properties. Nutrients 2021, 13, 2595. [Google Scholar] [CrossRef] [PubMed]
- Vezza, T.; Algieri, F.; Garrido-Mesa, J.; Utrilla, M.P.; Rodríguez-Cabezas, M.E.; Baños, A.; Guillamón, E.; García, F.; Rodríguez-Nogales, A.; Gálvez, J. The Immunomodulatory Properties of Propyl-Propane Thiosulfonate Contribute to Its Intestinal Anti-Inflammatory Effect in Experimental Colitis. Mol. Nutr. Food Res. 2019, 63, 1800653. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Myhill, L.J.; Andersen-Civil, A.I.S.; Thamsborg, S.M.; Blanchard, A.; Williams, A.R. Garlic-Derived Organosulfur Compounds Regulate Metabolic and Immune Pathways in Macrophages and Attenuate Intestinal Inflammation in Mice. Mol. Nutr. Food Res. 2022, 66, e2101004. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Andersen-Civil, A.I.S.; Myhill, L.J.; Thamsborg, S.M.; Kot, W.; Krych, L.; Nielsen, D.S.; Blanchard, A.; Williams, A.R. The Phytonutrient Cinnamaldehyde Limits Intestinal Inflammation and Enteric Parasite Infection. J. Nutr. Biochem. 2022, 100, 108887. [Google Scholar] [CrossRef]
- Myhill, L.J.; Stolzenbach, S.; Mejer, H.; Jakobsen, S.R.; Hansen, T.V.A.; Andersen, D.; Brix, S.; Hansen, L.H.; Krych, L.; Nielsen, D.S. Fermentable Dietary Fiber Promotes Helminth Infection and Exacerbates Host Inflammatory Responses. J. Immunol. 2020, 204, 3042–3055. [Google Scholar] [CrossRef]
- Pardo-Hernández, M.; López-Delacalle, M.; Rivero, R.M. ROS and NO Regulation by Melatonin under Abiotic Stress in Plants. Antioxidants 2020, 9, 1078. [Google Scholar] [CrossRef]
- Blokhina, O.; Fagerstedt, K. V Oxidative Metabolism, ROS and NO under Oxygen Deprivation. Plant Physiol. Biochem. 2010, 48, 359–373. [Google Scholar] [CrossRef]
- Qi, S.; Xin, Y.; Guo, Y.; Diao, Y.; Kou, X.; Luo, L.; Yin, Z. Ampelopsin Reduces Endotoxic Inflammation via Repressing ROS-Mediated Activation of PI3K/Akt/NF-ΚB Signaling Pathways. Int. Immunopharmacol. 2012, 12, 278–287. [Google Scholar] [CrossRef]
- Neumann, C.A.; Krause, D.S.; Carman, C.V.; Das, S.; Dubey, D.P.; Abraham, J.L.; Bronson, R.T.; Fujiwara, Y.; Orkin, S.H.; Van Etten, R.A. Essential Role for the Peroxiredoxin Prdx1 in Erythrocyte Antioxidant Defence and Tumour Suppression. Nature 2003, 424, 561–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, K.; Lee, J.T.; Joe, E.; Kwon, T.K. An IκBα Phosphorylation Inhibitor Induces Heme Oxygenase-1(HO-1) Expression through the Activation of Reactive Oxygen Species (ROS)–Nrf2–ARE Signaling and ROS–PI3K/Akt Signaling in an NF-ΚB-Independent Mechanism. Cell. Signal. 2011, 23, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Botero, S.; Shea-Donohue, T.; Urban, J.F., Jr. The Pathogenicity of an Enteric Citrobacter Rodentium Infection Is Enhanced by Deficiencies in the Antioxidants Selenium and Vitamin E. Infect. Immun. 2011, 79, 1471–1478. [Google Scholar] [CrossRef] [Green Version]
- Fields, J.A.; Thompson, S.A. Campylobacter Jejuni CsrA Mediates Oxidative Stress Responses, Biofilm Formation, and Host Cell Invasion. J. Bacteriol. 2008, 190, 3411–3416. [Google Scholar] [CrossRef] [Green Version]
- Dinkova-Kostova, A.T.; Talalay, P. NAD(P)H:quinone acceptor oxidoreductase 1 (NQO1), a multifunctional antioxidant enzyme and exceptionally versatile cytoprotector. Arch. Biochem. Biophys. 2010, 501, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, M.P.; Steele, D.; Chaignat, C.-L.; Kieny, M.P. A Review of Vaccine Research and Development: Human Enteric Infections. Vaccine 2006, 24, 2732–2750. [Google Scholar] [CrossRef]
- Hill Gaston, J.S.; Lillicrap, M.S. Arthritis Associated with Enteric Infection. Best Pract. Res. Clin. Rheumatol. 2003, 17, 219–239. [Google Scholar] [CrossRef]
- Wlodarska, M.; Willing, B.P.; Bravo, D.M.; Finlay, B.B. Phytonutrient Diet Supplementation Promotes Beneficial Clostridia Species and Intestinal Mucus Secretion Resulting in Protection against Enteric Infection. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinado, M.J.; Ruiz, R.; Echávarri, A.; Rubio, L.A. Garlic Derivative Propyl Propane Thiosulfonate Is Effective against Broiler Enteropathogens In Vivo. Poult. Sci. 2012, 91, 2148–2157. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Mokhosoev, I.M.; Feldman, N.B.; Lutsenko, S. V ROS and RNS Signalling: Adaptive Redox Switches through Oxidative/Nitrosative Protein Modifications. Free Radic. Res. 2018, 52, 507–543. [Google Scholar] [CrossRef]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive Oxygen Species (ROS) in Macrophage Activation and Function in Diabetes. Immunobiology 2019, 224, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Kovac, S.; Angelova, P.R.; Holmström, K.M.; Zhang, Y.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 Regulates ROS Production by Mitochondria and NADPH Oxidase. Biochim. Biophys. Acta-Gen. Subj. 2015, 1850, 794–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogdan, C. Nitric Oxide and the Immune Response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Ahn, H.; Kim, J.Y.; Kim, Y. Inhibitory Activity of Plant Extracts on Nitric Oxide Synthesis in LPS-activated Macrophages. Phyther. Res. 2003, 17, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Dam, B.; Misra, A.; Banerjee, S. Role of Gut Microbiota in Combating Oxidative Stress. In Oxidative Stress in Microbial Diseases; Springer: Singapore, 2019; pp. 43–82. [Google Scholar]
- Tomasello, G.; Mazzola, M.; Leone, A.; Sinagra, E.; Zummo, G.; Farina, F.; Damiani, P.; Cappello, F.; Gerges Geagea, A.; Jurjus, A. Nutrition, Oxidative Stress and Intestinal Dysbiosis: Influence of Diet on Gut Microbiota in Inflammatory Bowel Diseases. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech Repub. 2016, 160, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Olejar, K.J.; On, S.L.W.; Chelikani, V. The Potential of Lactobacillus spp. for Modulating Oxidative Stress in the Gastrointestinal Tract. Antioxidants 2020, 9, 610. [Google Scholar] [CrossRef]
- Bereswill, S.; Ekmekciu, I.; Escher, U.; Fiebiger, U.; Stingl, K.; Heimesaat, M.M. Lactobacillus Johnsonii Ameliorates Intestinal, Extra-Intestinal and Systemic pro-Inflammatory Immune Responses Following Murine Campylobacter Jejuni Infection. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Valladares, R.; Sankar, D.; Li, N.; Williams, E.; Lai, K.-K.; Abdelgeliel, A.S.; Gonzalez, C.F.; Wasserfall, C.H.; Larkin, J., III; Schatz, D. Lactobacillus johnsonii N6. 2 Mitigates the Development of Type 1 Diabetes in BB-DP Rats. PLoS ONE 2010, 5, e10507. [Google Scholar] [CrossRef] [Green Version]
- Gould, R.L.; Pazdro, R. Impact of Supplementary Amino Acids, Micronutrients, and Overall Diet on Glutathione Homeostasis. Nutrients 2019, 11, 1056. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Han, N.; Zhao, K.; Li, Y.; Chi, Y.; Wang, B. Protective Effects of Pyrroloquinoline Quinone against Oxidative Stress-Induced Cellular Senescence and Inflammation in Human Renal Tubular Epithelial Cells via Keap1/Nrf2 Signaling Pathway. Int. Immunopharmacol. 2019, 72, 445–453. [Google Scholar] [CrossRef]
- Cascajosa-Lira, A.; Pichardo, S.; Baños, A.; Guillamón, E.; Molina-Hernández, V.; Moyano, R.; Jos, Á.; Cameán, A.M. Acute and Subchronic 90-Days Toxicity Assessment of Propyl-Propane-Thiosulfinate (PTS) in Rats. Food Chem. Toxicol. 2022, 161, 112827. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Lillehoj, H.S.; Lee, S.H.; Lillehoj, E.P. Bravo D. Improved resistance to Eimeria acervulina infection in chickens due to dietary supplementation with garlic metabolites. Br. J. Nutrition 2013, 109, 76–88. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | GeneBank Accession | Primer Sequence (5-3 Forward/Reverse) |
|---|---|---|
| Nrf2 | AH006764.2 | F: CGAGATATACGCAGGAGAGGTAAGA R: GCTCGACAATGTTCTCCAGCTT |
| Keap1 | AB020063.1 | F: CAACTTCGCGGAGCAGATCG R: AGCTGGCAGTGTGACAGGTT |
| Nqo1 | NM_008706.5 | F: CATCCTGCGTTTCTGTGGCT R: TCTCCTCCCAGACGGTTTCC |
| Gpx2 | U62658.1 | F: CAAGTATGTCCGACCTGGGG R: GGGTAGGGCAGCTTGTCTTT |
| Hmox1 | NM_010442.2 | F: GAACCCAGTCTATGCCCCAC R: GCGTGCAAGGGATGATTTCC |
| Muc2 | NM_023566.4 | F: GTCCTGACCAAGAGCGAACA R: TTGAAGGCCACCACGTTCTT |
| Nos2 | NM_010927.4 | F: GGTGAAGGGACTGAGCTGTT R: TGCACTTCTGCTCCAAATCCA |
| Gapdh | BC023196.2 | F: TATGTCGTGGAGTCTACTGGT R: GAGTTGTCATATTTCTCGTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, L.; Andersen-Civil, A.I.S.; Castro-Meija, J.L.; Nielsen, D.S.; Blanchard, A.; Olsen, J.E.; Thamsborg, S.M.; Williams, A.R. Garlic-Derived Metabolites Exert Antioxidant Activity, Modulate Gut Microbiota Composition and Limit Citrobacter rodentium Infection in Mice. Antioxidants 2022, 11, 2033. https://doi.org/10.3390/antiox11102033
Zhu L, Andersen-Civil AIS, Castro-Meija JL, Nielsen DS, Blanchard A, Olsen JE, Thamsborg SM, Williams AR. Garlic-Derived Metabolites Exert Antioxidant Activity, Modulate Gut Microbiota Composition and Limit Citrobacter rodentium Infection in Mice. Antioxidants. 2022; 11(10):2033. https://doi.org/10.3390/antiox11102033
Chicago/Turabian StyleZhu, Ling, Audrey I. S. Andersen-Civil, Josue L. Castro-Meija, Dennis S. Nielsen, Alexandra Blanchard, John E. Olsen, Stig M. Thamsborg, and Andrew R. Williams. 2022. "Garlic-Derived Metabolites Exert Antioxidant Activity, Modulate Gut Microbiota Composition and Limit Citrobacter rodentium Infection in Mice" Antioxidants 11, no. 10: 2033. https://doi.org/10.3390/antiox11102033