
Onion and Apple Functional Ingredients Intake Improves Antioxidant and Inflammatory Status and Vascular Injury in Obese Zucker Rats
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Onion and Apple Powder Preparation
2.2. Animals and Experimental Design
2.3. Urine, Faeces, Blood and Organs Sampling
2.4. Plasma Lipid Concentrations and Biochemical Parameters
2.5. Plasma Antioxidant Activity
2.6. Antioxidant Enzyme Activities
2.7. Oxidative Stress Biomarkers
2.8. Plasma Vascular Injury Biomarkers and Metabolic Hormone Concentration
2.9. Vascular Reactivity in Mesenteric Arteries
2.10. Detection of Superoxide Anion Generation
2.11. RNA Isolation and Quantitative Real Time PCR (RT-PCR) Assay
2.12. Statistical Analysis
3. Results and Discussion
3.1. Weight Gain and Feed Intake
3.2. Plasma Lipid Concentrations and Biochemical Parameters
3.3. Antioxidant, Inflammatory and Vascular Injury Biomarkers and Metabolic Hormones
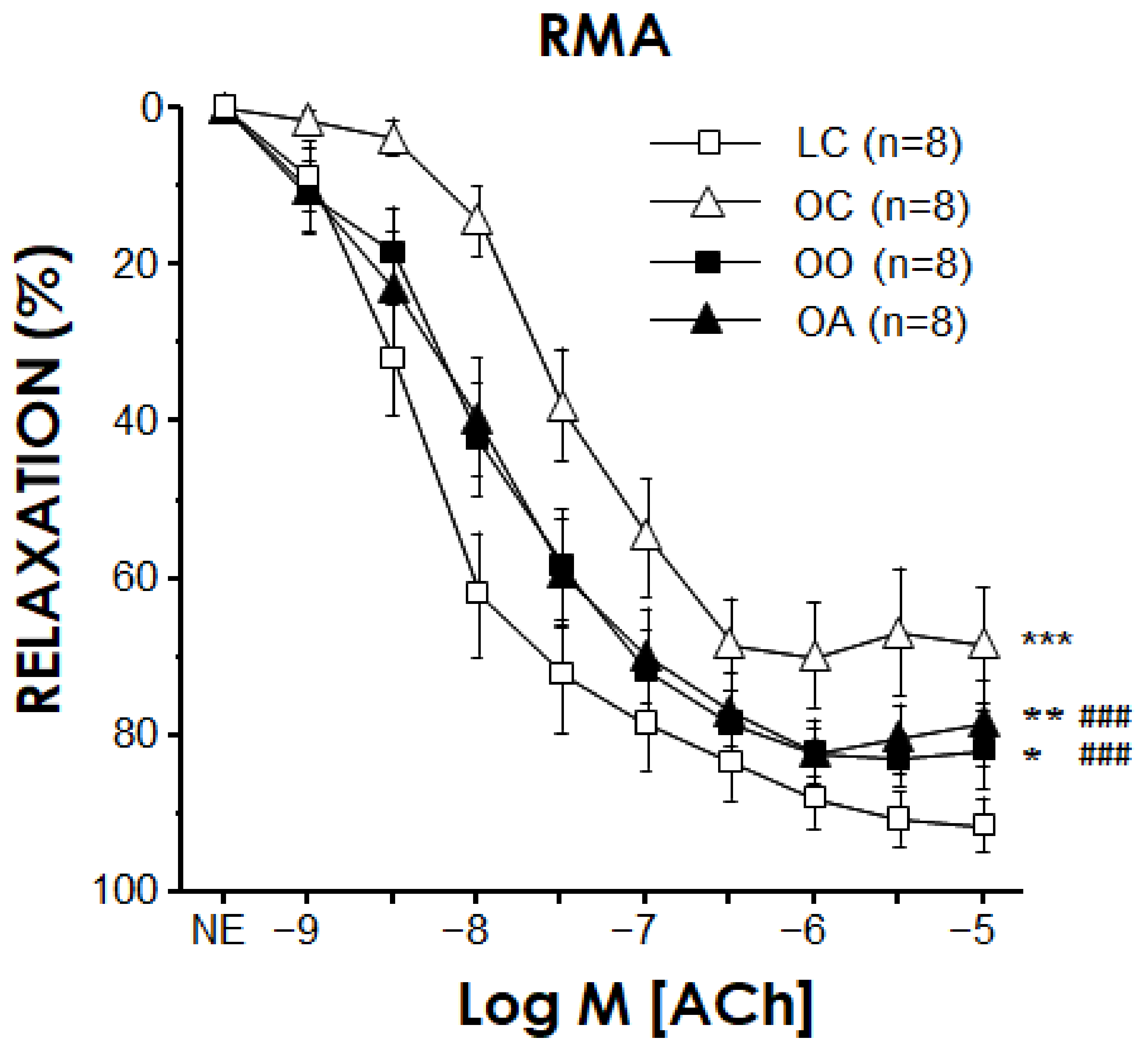
3.4. Vascular Reactivity in Mesenteric Arteries
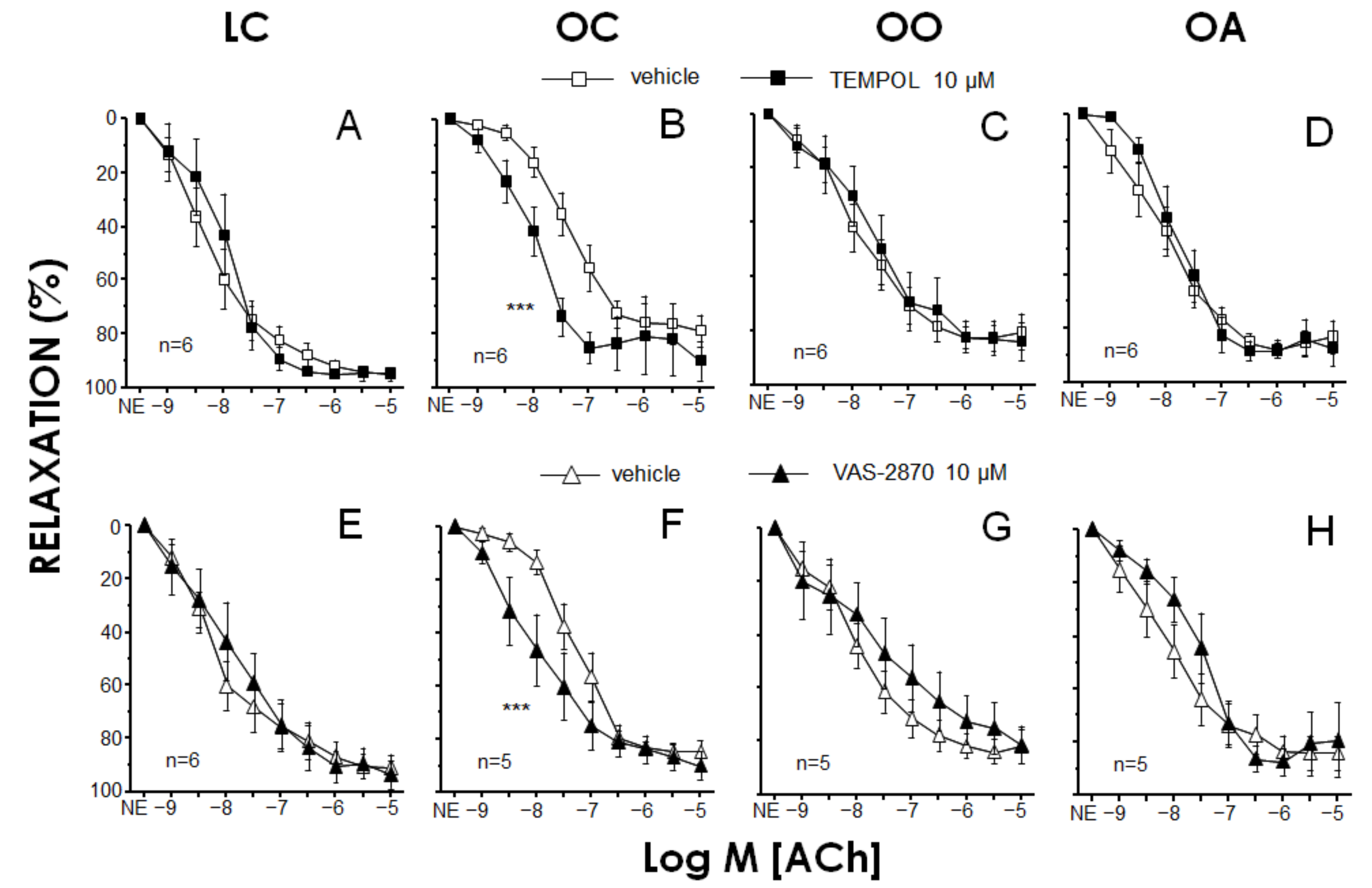
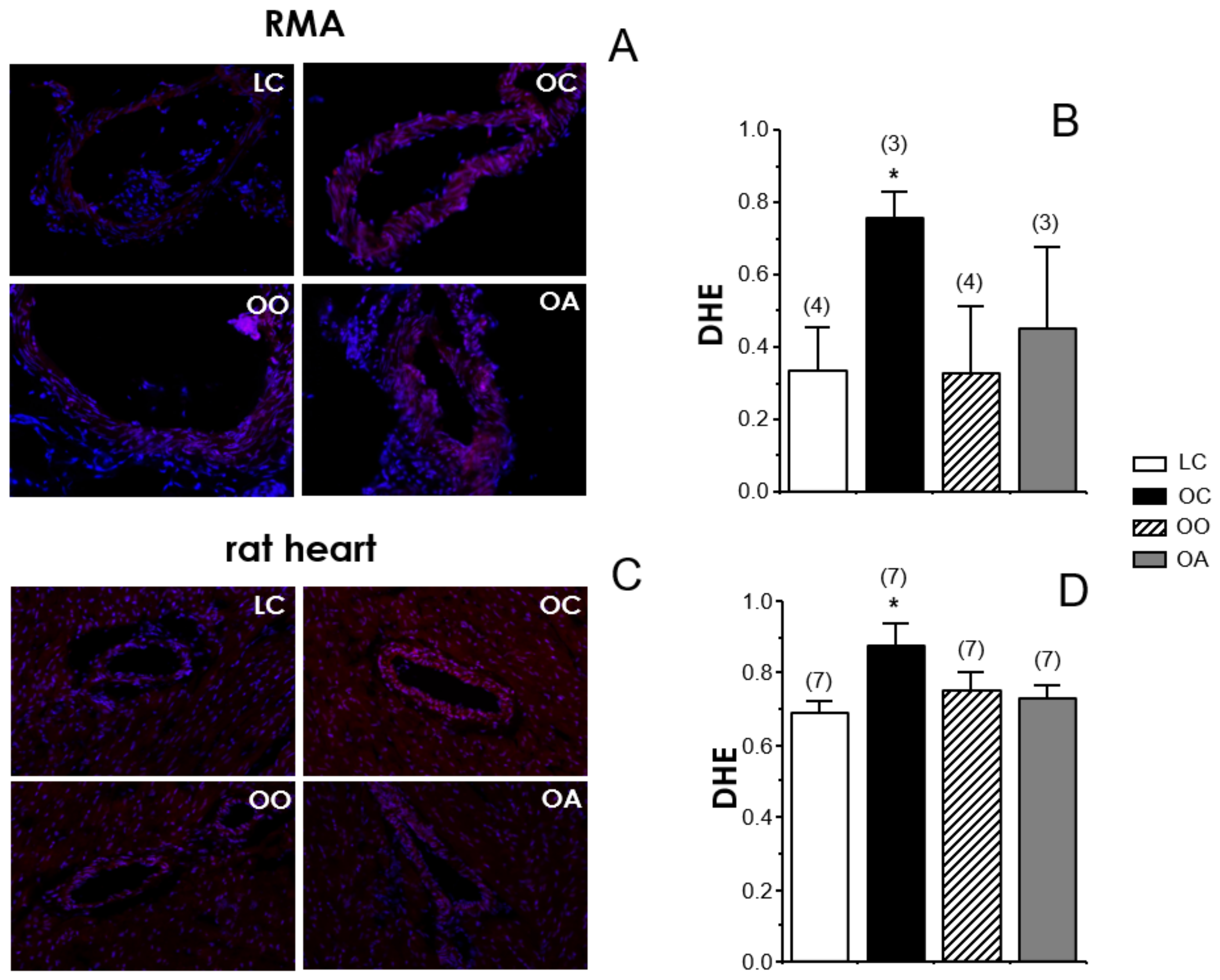
3.5. Detection of Superoxide Anion Generation
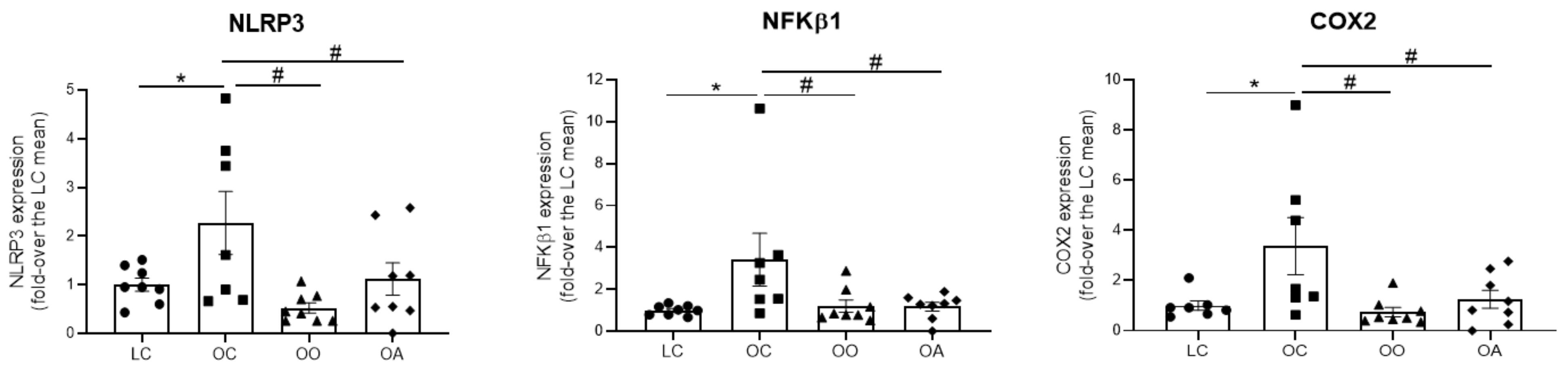
3.6. RNA Isolation and Quantitative Real Time PCR (RT-PCR) Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- WHO. World Health Organization. Available online: https://www.who.int/news-room/facts-in-pictures/detail/6-facts-on-obesity (accessed on 30 May 2022).
- Karasawa, M.G.; Chakravarthi, M. Fruits as prospective reserves of bioactive compounds: A review. Nat. Prod. Bioprospect. 2018, 8, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Thirumurugan, K. Modulatory functions of bioactive fruits, vegetables and spices in adipogenesis and angiogenesis. J. Funct. Food. 2019, 53, 318–336. [Google Scholar] [CrossRef]
- Arruda, H.S.; Neri-Numa, I.A.; Akemi Kido, L.; Marostica Junior, M.R.; Pastore, G.M. Recent advances and possibilities for the use of plant phenolic compounds to manage ageing-related diseases. J. Funct. Food 2020, 75, 104203. [Google Scholar] [CrossRef]
- Ávila-Gálvez, M.A.; Giménez-Bastida, J.A.; Espín, J.C.; González-Sarrías, A. Dietary phenolics against breast cancer. A critical evidence-based review and future perspectives. Int. J. Mol. Sci. 2020, 21, 5718. [Google Scholar] [CrossRef] [PubMed]
- Veiga, M.; Costa, E.M.; Silva, S.; Pintado, M. Impact of plant extracts upon human health: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 873–886. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Poudyal, H.; Panchal, S.K. Functional foods as potential therapeutic options for metabolic syndrome. Obes. Rev. 2015, 16, 914–941. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, J.; Thottiam Vasudevan, R. Bioingredients: Functional properties and health impacts. Curr. Opin. Food Sci. 2018, 19, 120–128. [Google Scholar] [CrossRef]
- Functional Food Ingredients Market. Available online: https://www.marketsandmarkets.com/Market-Reports/functional-food-ingredients-market-9242020.html (accessed on 26 May 2022).
- Barba, F.J.; Terefe, N.S.; Buckow, R.; Knorr, D.; Orlien, V. New opportunities and perspectives of high-pressure treatment to improve health and safety attributes of foods. A review. Food Res. Int. 2015, 77, 725–742. [Google Scholar] [CrossRef]
- Santeramo, F.G.; Carlucci, D.; De Devitiis, B.; Seccia, A.; Stasi, A.; Viscecchia, R.; Nardone, G. Emerging trends in European food, diets and food industry. Food Res. Int. 2018, 104, 39–47. [Google Scholar] [CrossRef]
- Bhatta, S.; Stevanovic Janezic, T.; Ratti, C. Freeze-drying of plant-based foods. Foods 2020, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Harguindeguy, M.; Fissore, D. On the effects of freeze-drying processes on the nutritional properties of foodstuff: A review. Dry. Technol. 2020, 38, 846–868. [Google Scholar] [CrossRef]
- Waghmare, R.B.; Choudhary, P.; Moses, J.A.; Anandharamakrishnan, C.; Stapley, A.G.F. Trends in approaches to assist freeze-drying of food: A cohort study on innovations. Food Rev. Int. 2021. [Google Scholar] [CrossRef]
- Aprikian, O.; Busserolles, J.; Manach, C.; Mazur, A.; Morand, C.; Davicco, M.-J.; Besson, C.; Rayssiguier, Y.; Rémésy, C.; Demigné, C. Lyophilized apple counteracts the development of hypercholesterolemia, oxidative stress, and renal dysfunction in obese Zucker rats. J. Nutr. 2002, 132, 1969–1976. [Google Scholar] [CrossRef]
- Marrelli, M.; Amodeo, V.; Statti, G.; Conforti, F. Biological properties and bioactive components of Allium cepa L.: Focus on potential benefits in the treatment of obesity and related comorbidities. Molecules 2019, 24, 119. [Google Scholar] [CrossRef]
- Skinner, R.C.; Warren, D.C.; Naveed, M.; Agarwal, G.; Benedito, V.A.; Tou, J.C. Apple pomace improves liver and adipose inflammatory and antioxidant status in young female rats consuming a Western diet. J. Funct. Food. 2019, 61, 103471. [Google Scholar] [CrossRef]
- Feng, S.; Yi, J.; Li, X.; Wu, X.; Zhao, Y.; Ma, Y.; Bi, J. Systematic review of phenolic compounds in apple fruits: Compositions, distribution, absorption, metabolism, and processing stability. J. Agric. Food Chem. 2021, 69, 7–27. [Google Scholar] [CrossRef]
- Koutsos, A.; Tuohy, K.M.; Lovegrove, J.A. Apples and cardiovascular health—Is the gut microbiota a core consideration? Nutrients 2015, 7, 3959–3998. [Google Scholar] [CrossRef]
- Suleria, H.A.R.; Butt, M.S.; Anjum, F.M.; Saeed, F.; Khalid, N. Onion: Nature protection against physiological threats. Crit. Rev. Food Sci. Nutr. 2015, 55, 50–66. [Google Scholar] [CrossRef]
- Perussello, C.A.; Zhang, Z.; Marzocchella, A.; Tiwari, B.K. Valorization of apple pomace by extraction of valuable compounds. Compr. Rev. Food Sci. Food Saf. 2017, 16, 776–796. [Google Scholar] [CrossRef]
- Teshika, J.D.; Zakariyyah, A.M.; Zaynab, T.; Zengin, G.; Rengasamy, K.R.R.; Pandian, S.K.; Fawzi, M.M. Traditional and modern uses of onion bulb (Allium cepa L.): A systematic Review. Crit. Rev. Food Sci. Nutr. 2019, 59, S39–S70. [Google Scholar] [CrossRef]
- Zhang, S.; Hu, C.; Guo, Y.; Wang, X.; Meng, Y. Polyphenols in fermented apple juice: Beneficial effects on human health. J. Funct. Food. 2021, 76, 104294. [Google Scholar] [CrossRef]
- Colina-Coca, C.; González-Peña, D.; De Ancos, B.; Sánchez-Moreno, C. Dietary onion ameliorates antioxidant defence, inflammatory response, and cardiovascular risk biomarkers in hypercholesterolemic Wistar rats. J. Funct. Food 2017, 36, 300–309. [Google Scholar] [CrossRef]
- Jensen, E.N.; Buch-Andersen, T.; Ravn-Haren, G.; Dragsted, L.O. Mini-review: The effects of apples on plasma cholesterol levels and cardiovascular risk—A review of the evidence. J. Horticult. Sci. Biotechnol. 2009, 84, 34–41. [Google Scholar] [CrossRef]
- Yuste, S.; Ludwig, I.A.; Romero, M.P.; Piñol-Felis, C.; Catalán, U.; Pedret, A.; Valls, R.M.; Fernández-Castillejo, S.; Motilva, M.J.; Macià, A.; et al. Metabolic fate and cardiometabolic effects of phenolic compounds from red-fleshed apple in hypercholesterolemic rats: A comparative study with common white-fleshed apple. The AppleCOR study. Mol. Nutr. Food Res. 2021, 65, 2001225. [Google Scholar] [CrossRef]
- El Assar, M.; Angulo, J.; Rodríguez-Mañas, L. Diabetes and ageing-induced vascular inflammation. J. Physiol. 2016, 594, 2125–2146. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, H.; Xia, N. The interplay between adipose tissue and vasculature: Role of oxidative stress in obesity. Front. Cardiovasc. Med. 2021, 8, 650214. [Google Scholar] [CrossRef]
- Nyawo, T.A.; Pheiffer, C.; Mazibuko-Mbeje, S.E.; Mthembu, S.X.H.; Nyambuya, T.M.; Nkambule, B.B.; Sadie-Van Gijsen, H.; Strijdom, H.; Tiano, L.; Dludla, P.V. Physical exercise potentially targets epicardial adipose tissue to reduce cardiovascular disease risk in patients with metabolic diseases: Oxidative stress and inflammation emerge as major therapeutic targets. Antioxidants 2021, 10, 1758. [Google Scholar] [CrossRef]
- Silveira Rossi, J.L.; Barbalho, S.M.; Reverete de Araujo, R.; Bechara, M.D.; Sloan, K.P.; Sloan, L.A. Metabolic syndrome and cardiovascular diseases: Going beyond traditional risk factors. Diabetes Metab. Res. Rev. 2022, 38, e3502. [Google Scholar] [CrossRef]
- Lutz, T.A. An overview of rodent models of obesity and type 2 diabetes. In Animal Models of Diabetes. Methods in Molecular Biology; King, A., Ed.; Humana: New York, NY, USA, 2020; Volume 2128, pp. 11–24. [Google Scholar] [CrossRef]
- González-Peña, D.; Colina-Coca, C.; Char, C.D.; Cano, M.P.; De Ancos, B.; Sánchez-Moreno, C. Hyaluronidase inhibiting activity and radical scavenging potential of flavonols in processed onion. J. Agric. Food Chem. 2013, 61, 4862–4872. [Google Scholar] [CrossRef]
- Colina-Coca, C.; González-Peña, D.; Vega, E.; De Ancos, B.; Sánchez-Moreno, C. Novel approach for the determination of volatile compounds in processed onion by headspace gas chromatography–mass spectrometry (HS GC-MS). Talanta 2013, 103, 137–144. [Google Scholar] [CrossRef]
- Colina-Coca, C.; De Ancos, B.; Sánchez-Moreno, C. Nutritional composition of processed onion: S-alk(en)yl-L-cysteine sulfoxides, organic acids, sugars, minerals, and vitamin C. Food Bioprocess Technol. 2014, 7, 289–298. [Google Scholar] [CrossRef]
- Reeves, P.G. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr. 1997, 127, 838S–841S. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef]
- Roldán-Marín, E.; Krath, B.N.; Poulsen, M.; Binderup, M.L.; Nielsen, T.H.; Hansen, M.; Langkilde, S.; Cano, M.P.; Sánchez-Moreno, C.; Dragsted, L.O. Effects of an onion by-product on bioactivity and safety markers in healthy rats. Br. J. Nutr. 2009, 102, 1574–1582. [Google Scholar] [CrossRef]
- Roldán-Marín, E.; Jensen, R.I.; Krath, B.N.; Kristensen, M.; Poulsen, M.; Cano, M.P.; Sánchez-Moreno, C.; Dragsted, L.O. An onion byproduct affects plasma lipids in healthy rats. J. Agric. Food Chem. 2010, 58, 5308–5314. [Google Scholar] [CrossRef]
- González-Peña, D.; Angulo, J.; Vallejo, S.; Colina-Coca, C.; De Ancos, B.; Sánchez-Ferrer, C.F.; Peiró, C.; Sánchez-Moreno, C. High-cholesterol diet enriched with onion affects endothelium-dependent relaxation and NADPH oxidase activity in mesenteric microvessels from Wistar rats. Nutr. Metab. 2014, 11, 57. [Google Scholar] [CrossRef]
- Angulo, J.; El Assar, M.; Sevilleja-Ortiz, A.; Fernández, A.; Sánchez-Ferrer, A.; Romero-Otero, J.; Martínez-Salamanca, J.I.; La Fuente, J.M.; Rodríguez-Mañas, L. Short-term pharmacological activation of Nrf2 ameliorates vascular dysfunction in aged rats and in pathological human vasculature. A potential target for therapeutic intervention. Redox Biol. 2019, 26, 101271. [Google Scholar] [CrossRef]
- Romacho, T.; Valencia, I.; Ramos-González, M.; Vallejo, S.; López-Esteban, M.; Lorenzo, O.; Cannata, P.; Romero, A.; San Hipólito-Luengo, A.; Gómez-Cerezo, J.F.; et al. Visfatin/eNampt induces endothelial dysfunction in vivo: A role for Toll-Like Receptor 4 and NLRP3 inflammasome. Sci. Rep. 2020, 10, 5386. [Google Scholar] [CrossRef]
- Alsoodeeri, F.N.; Alqabbani, H.M.; Aldossari, N.M. Effects of cinnamon (Cinnamomum cassia) consumption on serum lipid profiles in albino rats. J. Lipids 2020, 2020, 8469830. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.K.; Lee, J.-J.; Kim, Y.-K.; Lee, Y.; Ha, J.-H. Stachys sieboldii Miq. root attenuates weight gain and dyslipidemia in rats on a high-fat and high-cholesterol diet. Nutrients 2020, 12, 2063. [Google Scholar] [CrossRef]
- Shaban, E.E.; Elbakry, H.F.H.; Ibrahim, K.S.; El Sayed, E.M.; Salama, D.M.; Farrag, A.-R.H. The effect of white kidney bean fertilized with nano-zinc on nutritional and biochemical aspects in rats. Biotechnol. Rep. 2019, 23, e00357. [Google Scholar] [CrossRef]
- Yoshinari, O.; Shiojima, Y.; Igarashi, K. Anti-obesity effects of onion extract in Zucker diabetic fatty rats. Nutrients 2012, 4, 1518–1526. [Google Scholar] [CrossRef]
- Cho, K.-D.; Han, C.-K.; Lee, B.-H. Loss of body weight and fat and improved lipid profiles in obese rats fed apple pomace or apple juice concentrate. J. Med. Food 2013, 16, 823–830. [Google Scholar] [CrossRef]
- Li, W.; Yang, C.; Mei, X.; Huang, R.; Zhang, S.; Tang, Y.; Dong, Q.; Zhou, C. Effect of the polyphenol-rich extract from Allium cepa on hyperlipidemic Sprague-Dawley rats. J. Food Biochem. 2021, 45, e13565. [Google Scholar] [CrossRef]
- Cavalot, F.; Pagliarino, A.; Valle, M.; Di Martino, L.; Bonomo, K.; Massucco, P.; Anfossi, G.; Trovati, M. Postprandial blood glucose predicts cardiovascular events and all-cause mortality in type 2 diabetes in a 14-year follow-up: Lessons from the San Luigi Gonzaga Diabetes study. Diabetes Care 2011, 34, 2237–2243. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Tan, Y.; Zhou, J.; Xiao, L.; Johnson, M.; Qu, X. Green tea polyphenols ameliorate metabolic abnormalities and insulin resistance by enhancing insulin signalling in skeletal muscle of Zucker fatty rats. Clin. Sci. 2020, 134, 1167–1180. [Google Scholar] [CrossRef]
- Budak, N.H.; Kumbul Doguc, D.; Savas, C.M.; Seydim, A.C.; Kok Tas, T.; Ciris, M.I.; Guzel-Seydim, Z.B. Effects of apple cider vinegars produced with different techniques on blood lipids in high-cholesterol-fed rats. J. Agric. Food Chem. 2011, 59, 6638–6644. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.A.; Boby, N.; Lee, E.-B.; Hong, J.-H.; Park, S.-C. Anti-obesity effects of Ecklonia cava extract in high-fat diet-induced obese rats. Antioxidants 2022, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Codoñer-Franch, P.; Betoret, E.; López-Jaén, A.B.; Betoret, N.; Fito, P.; Valls-Bellés, V. Dried apple enriched with mandarin juice counteracts tamoxifen-induced oxidative stress in rats. Int. J. Food Sci. Nutr. 2013, 64, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Del Pino-García, R.; Rivero-Pérez, M.D.; González-SanJosé, M.L.; Croft, K.D.; Muñiz, P. Bioavailability of phenolic compounds and antioxidant effects of wine pomace seasoning after oral administration in rats. J. Funct. Food 2016, 25, 486–496. [Google Scholar] [CrossRef]
- Del Pino-García, R.; Rivero-Pérez, M.D.; González-SanJosé, M.L.; Castilla-Camina, P.; Croft, K.D.; Muñiz, P. Attenuation of oxidative stress in type 1 diabetic rats supplemented with a seasoning obtained from winemaking by-products and its effect on endothelial function. Food Funct. 2016, 7, 4410. [Google Scholar] [CrossRef]
- Del Pino-García, R.; Rivero-Pérez, M.D.; González-SanJosé, M.L.; Croft, K.D.; Muñiz, P. Antihypertensive and antioxidant effects of supplementation with red wine pomace in spontaneously hypertensive rats. Food Funct. 2017, 8, 2444. [Google Scholar] [CrossRef]
- Carmiel-Haggai, M.; Cederbaum, A.I.; Nieto, N. A high-fat diet leads to the progression of non-alcoholic fatty liver disease in obese rats. FASEB J. 2005, 19, 136–138. [Google Scholar] [CrossRef]
- Oana, F.; Takeda, H.; Hayakawa, K.; Matsuzawa, A.; Akahane, S.; Isaji, M.; Akahane, M. Physiological difference between obese (fa/fa) Zucker rats and lean Zucker rats concerning adiponectin. Metab. Clin. Exp. 2005, 54, 995–1001. [Google Scholar] [CrossRef]
- Deka, B.; Ramrao Barge, S.; Bharadwaj, S.; Kashyap, B.; Manna, P.; Borah, J.C.; Talukdar, N.C. Beneficial effect of the methanolic leaf extract of Allium hookeri on stimulating glutathione biosynthesis and preventing impaired glucose metabolism in type 2 diabetes. Arch. Biochem. Biophys. 2021, 708, 108961. [Google Scholar] [CrossRef]
- Reznick, A.Z.; Packer, L. Oxidative damage to proteins: Spectrophotometric method for carbonyl assay. Methods Enzymol. 1994, 233, 357–363. [Google Scholar] [CrossRef]
- De Souza, M.O.; Silva, M.; Silva, M.E.; Oliveira, R.P.; Pedrosa, M.L. Diet supplementation with acai (Euterpe oleracea Mart.) pulp improves biomarkers of oxidative stress and the serum lipid profile in rats. Nutrition 2010, 26, 804–810. [Google Scholar] [CrossRef]
- Kujawska, M.; Ignatowicz, E.; Ewertowska, M.; Markowski, J.; Jodynis-Liebert, J. Cloudy apple juice protects against chemical-induced oxidative stress in rat. Eur. J. Nutr. 2011, 50, 53–60. [Google Scholar] [CrossRef]
- Noratto, G.D.; Lage, N.N.; Chew, B.P.; Mertens-Talcott, S.U.; Talcott, S.T.; Pedrosa, M.L. Non-anthocyanin phenolics in cherry (Prunus avium L.) modulate IL-6, liver lipids and expression of PPARδ and LXRs in obese diabetic (db/db) mice. Food Chem. 2018, 266, 405–414. [Google Scholar] [CrossRef]
- Chiorcea-Paquim, A.M. 8-oxoguanine and 8-oxodeoxyguanosine biomarkers of oxidative DNA damage: A review on HPLC–ECD determination. Molecules 2022, 27, 1620. [Google Scholar] [CrossRef]
- Azuma, K.; Ippoushi, K.; Terao, J. Evaluation of tolerable levels of dietary quercetin for exerting its antioxidative effect in high cholesterol-fed rats. Food Chem. Toxicol. 2010, 48, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Pradeep, S.R.; Srinivasan, K. Alleviation of oxidative stress-mediated nephropathy by dietary fenugreek (Trigonella foenum-graecum) seeds and onion (Allium cepa) in streptozotocin-induced diabetic rats. Food Funct. 2018, 9, 134. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.; Margaritis, M.; Channon, K.M.; Antoniades, C. Evaluating oxidative stress in human cardiovascular disease: Methodological aspects and considerations. Curr. Med. Chem. 2012, 19, 2504–2520. [Google Scholar] [CrossRef] [PubMed]
- Molinett, S.; Nuñez, F.; Moya-León, M.A.; Zúñiga-Hernández, J. Chilean strawberry consumption protects against LPS-induced liver injury by anti-inflammatory and antioxidant capability in Sprague-Dawley rats. Evid.-Based Complement. Altern. Med. 2015, 2015, 320136. [Google Scholar] [CrossRef]
- Waltz, P.; Escobar, D.; Botero, A.M.; Zuckerbraun, B.S. Nitrate/Nitrite as critical mediators to limit oxidative injury and inflammation. Antioxid. Redox Signal. 2015, 23, 328–339. [Google Scholar] [CrossRef]
- DeMartino, A.W.; Kim-Shapiro, D.B.; Patel, R.P.; Gladwin, M.T. Nitrite and nitrate chemical biology and signalling Br. J. Pharmacol. 2019, 176, 228–245. [Google Scholar] [CrossRef]
- Maneesai, P.; Iampanichakul, M.; Chaihongsa, N.; Poasakate, A.; Potue, P.; Rattanakanokchai, S.; Bunbupha, S.; Chiangsaen, P.; Pakdeechote, P. Butterfly pea flower (Clitoria ternatea Linn.) extract ameliorates cardiovascular dysfunction and oxidative stress in nitric oxide-deficient hypertensive rats. Antioxidants 2021, 10, 523. [Google Scholar] [CrossRef]
- Poasakate, A.; Maneesai, P.; Rattanakanokchai, S.; Bunbupha, S.; Tong-Un, T.; Pakdeechote, P. Genistein prevents nitric oxide deficiency-induced cardiac dysfunction and remodeling in rats. Antioxidants 2021, 10, 237. [Google Scholar] [CrossRef]
- Mukai, Y.; Sato, S. Polyphenol-containing azuki bean (Vigna angularis) extract attenuates blood pressure elevation and modulates nitric oxide synthase and caveolin-1 expressions in rats with hypertension. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 491–497. [Google Scholar] [CrossRef]
- Barbu, E.; Popescu, M.-R.; Popescu, A.-C.; Balanescu, S.-M. Inflammation as a precursor of atherothrombosis, diabetes and early vascular aging. Int. J. Mol. Sci. 2022, 23, 963. [Google Scholar] [CrossRef]
- Belobrajdic, D.P.; Lam, Y.Y.; Mano, M.; Wittert, G.A.; Bird, A.R. Cereal based diets modulate some markers of oxidative stress and inflammation in lean and obese Zucker rats. Nutr. Metab. 2011, 8, 27. [Google Scholar] [CrossRef]
- Alessi, M.C.; Bastelica, D.; Mavri, A.; Morange, P.; Berthet, B.; Grino, M.; Juhan-Vague, I. Plasma PAI-1 levels are more strongly related to liver steatosis than to adipose tissue accumulation. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1262–1268. [Google Scholar] [CrossRef]
- Martins, S.V.; Lopes, P.A.; Alfaia, C.M.; Rodrigues, P.O.; Alves, S.P.; Pinto, R.M.A.; Castro, M.F.; Bessa, R.J.B.; Prates, J.A.M. Serum adipokine profile and fatty acid composition of adipose tissues are affected by conjugated linoleic acid and saturated fat diets in obese Zucker rats. Br. J. Nutr. 2010, 103, 869–878. [Google Scholar] [CrossRef]
- Sundstrom, J.; Evans, J.C.; Benjamin, E.J.; Levy, D.; Larson, M.G.; Sawyer, D.B.; Siwik, D.A.; Colucci, W.S.; Wilson, P.W.F.; Vasan, R.S. Relations of plasma total TIMP-1 levels to cardiovascular risk factors and echocardiographic measures: The Framingham heart study. Eur. Heart J. 2004, 25, 1509–1516. [Google Scholar] [CrossRef]
- Lieb, W.; Song, R.J.; Xanthakis, V.; Vasan, R.S. Association of circulating tissue inhibitor of metalloproteinases-1 and procollagen type III aminoterminal peptide levels with incident heart failure and chronic kidney disease. J. Am. Heart Assoc. 2019, 8, e011426. [Google Scholar] [CrossRef]
- Jiang, Y.C.; Han, X.; Dou, J.Y.; Yuan, M.H.; Zhou, M.J.; Cui, Z.Y.; Lian, L.H.; Nan, J.; Zhang, X.; Wu, Y.L. Protective role of Siberian onions against toxin induced liver dysfunction: An insight into health-promoting effects. Food Funct. 2022, 13, 4678. [Google Scholar] [CrossRef]
- Cho, J.W.; Cho, S.Y.; Lee, S.R.; Lee, K.S. Onion extract and quercetin induce matrix metalloproteinase-1 in vitro and in vivo. Int. J. Mol. Med. 2010, 25, 347–352. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Khotina, V.A.; Omelchenko, A.V.; Kalmykov, V.A.; Orekhov, A.N. The role of the VEGF family in atherosclerosis development and its potential as treatment targets. Int. J. Mol. Sci. 2022, 23, 931. [Google Scholar] [CrossRef]
- Marino, M.; Bo, C.D.; Tucci, M.; Klimis-Zacas, D.; Riso, P.; Porrini, M. Modulation of adhesion process, E-selectin and VEGF production by anthocyanins and their metabolites in an in vitro model of atherosclerosis. Nutrients 2020, 12, 655. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Moral, N.; Needs, P.W.; Moyle, C.W.A.; Kroon, P.A. Hydrophobic interactions drive binding between vascular endothelial growth factor-A (VEGFA) and polyphenolic inhibitors. Molecules 2019, 24, 2785. [Google Scholar] [CrossRef]
- Roy, S.; Mondru, A.K.; Chakraborty, T.; Das, A.; Dasgupta, S. Apple polyphenol phloretin complexed with ruthenium is capable of reprogramming the breast cancer microenvironment through modulation of PI3K/Akt/mTOR/VEGF pathways. Toxicol. Appl. Pharmacol. 2022, 434, 115822. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, I.; Tomassoni, D.; Moruzzi, M.; Roy, P.; Cifani, C.; Amenta, F.; Tayebati, S.K. Cardiovascular changes related to metabolic syndrome: Evidence in obese Zucker rats. Int. J. Mol. Sci. 2020, 21, 2035. [Google Scholar] [CrossRef] [PubMed]
- Brüll, V.; Burak, C.; Stoffel-Wagner, B.; Wolffram, S.; Nickenig, G.; Müller, C.; Langguth, P.; Alteheld, B.; Fimmers, R.; Naaf, S.; et al. Effects of a quercetin-rich onion skin extract on 24 h ambulatory blood pressure and endothelial function in overweight-to-obese patients with (pre-)hypertension: A randomised double-blinded placebo-controlled cross-over trial. Br. J. Nutr. 2015, 114, 1263–1277. [Google Scholar] [CrossRef] [PubMed]
- Mueller, D.; Triebel, S.; Rudakovski, O.; Richling, E. Influence of triterpenoids present in apple peel on inflammatory gene expression associated with inflammatory bowel disease (IBD). Food Chem. 2013, 139, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Gao, L.; Guo, Y.; Xu, Y. Short-term phlorizin treatment attenuates adipose tissue inflammation without alerting obesity in high-fat diet fed mice. J. Food Biochem. 2017, 41, e12407. [Google Scholar] [CrossRef]
- Alsanea, S.; Gao, M.; Liu, D. Phloretin prevents high-fat diet-induced obesity and improves metabolic homeostasis. AAPS J. 2017, 19, 797–805. [Google Scholar] [CrossRef]
- Brüll, V.; Burak, C.; Stoffel-Wagner, B.; Wolffram, S.; Nickenig, G.; Müller, C.; Langguth, P.; Alteheld, B.; Fimmers, R.; Stehle, P.; et al. No effects of quercetin from onion skin extract on serum leptin and adiponectin concentrations in overweight-to-obese patients with (pre-)hypertension: A randomized double-blinded, placebo-controlled crossover trial. Eur. J. Nutr. 2017, 56, 2265–2275. [Google Scholar] [CrossRef]
- Kim, K.A.; Jung-Eun Yim, J.E. The effect of onion peel extract on inflammatory mediators in Korean overweight and obese women. Clin. Nutr. Res. 2016, 5, 261–269. [Google Scholar] [CrossRef]
- Eisner, A.; Ramachandran, P.; Cabalbag, C.; Metti, D.; Shamloufard, P.; Kern, M.; Hong, M.Y.; Hooshmand, S. Effects of dried apple consumption on body composition, serum lipid profile, glucose regulation, and inflammatory markers in overweight and obese children. J. Med. Food 2020, 23, 242–249. [Google Scholar] [CrossRef]
- Sánchez, D.; Quiñones, M.; Moulay, L.; Muguerza, B.; Miguel, M.; Aleixandre, A. Soluble fiber-enriched diets improve inflammation and oxidative stress biomarkers in Zucker fatty rats. Pharmacol. Res. 2011, 64, 31–35. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Yoon, T.; Kim, S.J.; Yang, W.K.; Kim, H.K. Anti-obesity activity of Allium fistulosum L. extract by down-regulation of the expression of lipogenic genes in high-fat diet-induced obese mice. Mol. Med. Rep. 2011, 4, 431–435. [Google Scholar] [CrossRef]
- Park, S.; Son, H.K.; Chang, H.C.; Lee, J.J. Effects of cabbage-apple juice fermented by Lactobacillus plantarum EM on lipid profile improvement and obesity amelioration in rats. Nutrients 2020, 12, 1135. [Google Scholar] [CrossRef]
- Halah, M.P.; Marangon, P.B.; Antunes-Rodrigues, J.; Elias, J.L.K. Neonatal nutritional programming impairs adiponectin effects on energy homeostasis in adult life of male rats. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E29–E37. [Google Scholar] [CrossRef]
- Phuwamongkolwiwat, P.; Hira, T.; Hara, H. A nondigestible saccharide, fructooligosaccharide, increases the promotive effect of a flavonoid, α-glucosyl-isoquercitrin, on glucagon-like peptide 1 (GLP-1) secretion in rat intestine and enteroendocrine cells. Mol. Nutr. Food Res. 2014, 58, 1581–1584. [Google Scholar] [CrossRef]
- Ronveaux, C.C.; Tomé, D.; Raybould, H.E. Glucagon-like peptide 1 interacts with ghrelin and leptin to regulate glucose metabolism and food intake through vagal afferent neuron signaling. J. Nutr. 2015, 145, 672–680. [Google Scholar] [CrossRef]
- Raasmaja, A.; Lecklin, A.; Li, X.M.; Zou, J.; Zhu, G.G.; Laakso, I.; Hiltunen, R. A water-alcohol extract of Citrus grandis whole fruits has beneficial metabolic effects in the obese Zucker rats fed with high fat/high cholesterol diet. Food Chem. 2013, 138, 1392–1399. [Google Scholar] [CrossRef]
- Liou, C.J.; Wu, S.J.; Shen, S.C.; Chen, L.C.; Chen, Y.L.; Huang, W.C. Phloretin ameliorates hepatic steatosis through regulation of lipogenesis and Sirt1/AMPK signaling. Cell Biosci. 2020, 10, 114. [Google Scholar] [CrossRef]
- Skrypnik, D.; Mostowska, A.; Jagodziński, P.P.; Bogdański, P. Association of rs699947 (−2578 C/A) and rs2010963 (−634 G/C) single nucleotide polymorphisms of the VEGF Gene, VEGF-A and leptin serum level, and cardiovascular risk in patients with excess body mass: A case-control study. J. Clin. Med. 2020, 9, 469. [Google Scholar] [CrossRef]
- Skrypnik, D.; Skrypnik, K.; Pelczyńska, M.; Sobieska, M.; Tinkov, A.A.; Suliburska, J.; Bogdański, P. The effect of Plantago major supplementation on leptin and VEGF-A serum levels, endothelial dysfunction and angiogenesis in obese women—A randomised trial. Food Funct. 2021, 12, 1708–1718. [Google Scholar] [CrossRef]
- González-Peña, D.; Dudzik, D.; García, A.; De Ancos, B.; Barbas, C.; Sánchez-Moreno, C. Metabolomic fingerprinting in the comprehensive study of liver changes associated with onion supplementation in hypercholesterolemic Wistar rats. Int. J. Mol. Sci. 2017, 18, 267. [Google Scholar] [CrossRef]
- González-Peña, D.; Checa, A.; De Ancos, B.; Wheelock, C.E.; Sánchez-Moreno, C. New insights into the effects of onion consumption on lipid mediators using a diet-induced model of hypercholesterolemia. Redox Biol. 2017, 11, 205–212. [Google Scholar] [CrossRef]
- Fernández-Jalao, I.; Balderas, C.; Sánchez-Moreno, C.; De Ancos, B. Impact of an in vitro dynamic gastrointestinal digestion on phenolic compounds and antioxidant capacity of apple treated by high-pressure processing. Innov. Food Sci. Emerg. Technol. 2020, 66, 102486. [Google Scholar] [CrossRef]
- Herranz, B.; Fernández-Jalao, I.; Álvarez, M.D.; Quiles, A.; Sánchez-Moreno, C.; Hernando, I.; De Ancos, B. Phenolic compounds, microstructure and viscosity of onion and apple products subjected to in vitro gastrointestinal digestion. Innov. Food Sci. Emerg. Technol. 2019, 51, 114–125. [Google Scholar] [CrossRef]
- Romanko, O.P.; Stepp, D.W. Reduced constrictor reactivity balances impaired vasodilation in the mesenteric circulation of the obese Zucker rat. Am. J. Physiol. Heart. Circ. Physiol. 2005, 289, H2097–H2102. [Google Scholar] [CrossRef]
- Agouni, A.; Lagrue-Lak-Hal, A.H.; Mostefai, H.A.; Tesse, A.; Mulder, P.; Rouet, P.; Desmoulin, F.; Heymes, C.; Martínez, M.C.; Andriantsitohaina, R. Red wine polyphenols prevent metabolic and cardiovascular alterations associated with obesity in Zucker fatty rats (Fa/Fa). PLoS ONE 2009, 4, e5557. [Google Scholar] [CrossRef]
- Santiago, E.; Martínez, M.P.; Climent, B.; Muñoz, M.; Briones, A.M.; Salaices, M.; García-Sacristán, A.; Rivera, L.; Prieto, D. Augmented oxidative stress and preserved vasoconstriction induced by hydrogen peroxide in coronary arteries in obesity: Role of COX-2. Br. J. Pharmacol. 2016, 173, 3176–3195. [Google Scholar] [CrossRef]
- El Assar, M.; Ruiz de Adana, J.C.; Angulo, J.; Pindado Martínez, M.L.; Hernández Matías, A.; Rodríguez-Mañas, L. Preserved endothelial function in human obesity in the absence of insulin resistance. J. Transl. Med. 2013, 11, 263. [Google Scholar] [CrossRef]
- Muñoz, M.; López-Oliva, M.E.; Rodríguez, C.; Martínez, M.P.; Sáenz-Medina, J.; Sánchez, A.; Climent, B.; Benedito, S.; García-Sacristán, A.; Rivera, L.; et al. Differential contribution of Nox1, Nox2 and Nox4 to kidney vascular oxidative stress and endothelial dysfunction in obesity. Redox Biol. 2020, 28, 101330. [Google Scholar] [CrossRef]
- Matute, A.; Tabart, J.; Cheramy-Bien, J.-P.; Pirotte, B.; Kevers, C.; Auger, C.; Schini-Kerth, V.; Dommes, J.; Defraigne, J.-O.; Pincemail, J. Compared phenolic compound contents of 22 commercial fruit and vegetable juices: Relationship to ex-vivo vascular reactivity and potential in vivo projection. Antioxidants 2020, 9, 92. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Wang, H.; He, Z. Recent advances in polyphenols improving vascular endothelial dysfunction induced by endogenous toxicity. J. Appl. Toxicol. 2021, 41, 701–712. [Google Scholar] [CrossRef]
- Sánchez, M.; Lodi, F.; Vera, R.; Villar, I.C.; Cogolludo, A.; Jiménez, R.; Moreno, L.; Romero, M.; Tamargo, J.; Pérez-Vizcaíno, F.; et al. Quercetin and isorhamnetin prevent endothelial dysfunction, superoxide production, and overexpression of p47 phox induced by angiotensin II in rat aorta. J. Nutr. 2007, 137, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, P.; Khanna, D.; Balakumar, P. Catechin averts experimental diabetes mellitus-induced vascular endothelial structural and functional abnormalities. Cardiovasc. Toxicol. 2014, 14, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.Y.; Lee, H.; Woo, J.S.; Jang, H.H.; Hwang, S.J.; Kim, H.S.; Kim, W.S.; Kim, Y.S.; Choue, R.; Cha, Y.J.; et al. Effect of onion peel extract on endothelial function and endothelial progenitor cells in overweight and obese individuals. Nutrition 2015, 31, 1131–1135. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, H.; Tsuge, N.; Sawada, H.; Higashi, Y. Chronic intake of onion extract containing quercetin improved postprandial endothelial dysfunction in healthy men. J. Am. Coll. Nutr. 2013, 32, 160–164. [Google Scholar] [CrossRef]
- Felice, F.; Francini, A.; Domenici, V.; Cifelli, M.; Belardinelli, E.; Sebastiani, L.; Cantini, C.; Di Stefano, R. Effects of extra virgin olive oil and apples enriched-dark chocolate on endothelial progenitor cells in patients with cardiovascular risk factors: A randomized cross-over trial. Antioxidants 2019, 8, 88. [Google Scholar] [CrossRef]
- Jia, M.; Ren, D.; Nie, Y.; Yang, X. Beneficial effects of apple peel polyphenols on vascular endothelial dysfunction and liver injury in high choline-fed mice. Food Funct. 2017, 8, 1282. [Google Scholar] [CrossRef]
- Bolea, G.; Philouze, C.; Dubois, M.; Risdon, S.; Humberclaude, A.; Ginies, C.; Charles, A.L.; Geny, B.; Reboul, C.; Arnaud, C.; et al. Digestive n-6 lipid oxidation, a key trigger of vascular dysfunction and atherosclerosis in the Western diet: Protective effects of apple polyphenols. Mol. Nutr. Food Res. 2021, 65, 2000487. [Google Scholar] [CrossRef]
- Vendrame, S.; Kristo, A.S.; Schuschke, D.A.; Klimis-Zacas, D. Wild blueberry consumption affects aortic vascular function in the obese Zucker rat. Appl. Physiol. Nutr. Metab. 2014, 39, 255–261. [Google Scholar] [CrossRef]
- Yu, Y.; Xiong, Y.; Montani, J.P.; Yang, Z.; Ming, X.F. En face detection of nitric oxide and superoxide in endothelial layer of intact arteries. J. Vis. Exp. 2016, 108, 53718. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rodríguez, P.; Ragusky, K.; Phuthong, S.; Ruvira, S.; Ramiro-Cortijo, D.; Cañas, S.; Rebollo-Hernanz, M.; Morales, M.D.; López de Pablo, A.L.; Martín-Cabrejas, M.A.; et al. Vasoactive properties of a cocoa shell extract: Mechanism of action and effect on endothelial dysfunction in aged rats. Antioxidants 2022, 11, 429. [Google Scholar] [CrossRef]
- Rozentsvit, A.; Vinokur, K.; Samuel, S.; Li, Y.; Gerdes, A.M.; Carrillo-Sepulveda, M.A. Ellagic acid reduces high glucose-induced vascular oxidative stress through ERK1/2/NOX4 signaling pathway. Cell. Physiol. Biochem. 2017, 44, 1174–1187. [Google Scholar] [CrossRef]
- Vandanmagsar, B.; Youm, Y.H.; Ravussin, A.; Galgani, J.E.; Stadler, K.; Mynatt, R.L.; Ravussin, E.; Stephens, J.M.; Dixit, V.D. The NLRP3 inflammasome instigates obesity-induced inflammation and insulin resistance. Nat. Med. 2011, 17, 179–188. [Google Scholar] [CrossRef]
- Abderrazak, A.; Syrovets, T.; Couchie, D.; El Hadri, K.; Friguet, B.; Simmet, T.; Rouis, M. NLRP3 inflammasome: From a danger signal sensor to a regulatory node of oxidative stress and inflammatory diseases. Redox Biol. 2015, 4, 296–307. [Google Scholar] [CrossRef]
- Rahman, I.; Marwick, J.; Kirkham, P. Redox modulation of chromatin remodeling: Impact on histone acetylation and deacetylation, NF-kappaB and pro-inflammatory gene expression. Biochem. Pharmacol. 2004, 68, 1255–1267. [Google Scholar] [CrossRef]
- Guo, X.D.; Zhang, D.Y.; Gao, X.J.; Parry, J.; Liu, K.; Liu, B.L.; Wang, M. Quercetin and quercetin-3-O-glucuronide are equally effective in ameliorating endothelial insulin resistance through inhibition of reactive oxygen species-associated inflammation. Mol. Nutr. Food Res. 2013, 57, 1037–1045. [Google Scholar] [CrossRef]
- Xu, Z.R.; Li, J.Y.; Dong, X.W.; Tan, Z.J.; Wu, W.Z.; Xie, Q.M.; Yang, Y.M. Apple polyphenols decrease atherosclerosis and hepatic steatosis in ApoE−/− mice through the ROS/MAPK/NF-κB pathway. Nutrients 2015, 7, 7085–7105. [Google Scholar] [CrossRef]
- Xiao, J.; Ho, C.T.; Liong, E.C.; Nanji, A.A.; Leung, T.M.; Lau, T.Y.H.; Fung, M.L.; Tipoe, G.L. Epigallocatechin gallate attenuates fibrosis, oxidative stress, and inflammation in non-alcoholic fatty liver disease rat model through TGF/SMAD, PI3 K/Akt/FoxO1, and NF-kappa B pathways. Eur. J. Nutr. 2014, 53, 187–199. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Onion Powder | Apple Powder | |
|---|---|---|
| Protein (g/100 g) | 9.87 ± 0.07 | 1.60 ± 0.01 |
| Lipids (g/100 g) | 1.35 ± 0.05 | 1.45 ± 0.07 |
| Carbohydrates (g/100 g) | 79.12 ± 3.18 | 85.67 ± 3.84 |
| Glucose (g/100 g) | 32.81 ± 1.25 | 15.23 ± 1.81 |
| Fructose (g/100 g) | 19.04 ± 2.50 | 31.37 ± 3.08 |
| Sucrose (g/100 g) | 7.74 ± 1.28 | 5.93 ± 0.85 |
| Total dietary fibre (g/100 g) | 16.86 ± 1.23 | 14.43 ± 1.89 |
| Soluble fibre (g/100 g) | 3.71 ± 0.07 | 3.14 ± 0.56 |
| Insoluble fibre (g/100 g) | 13.15 ± 0.97 | 11.29 ± 1.63 |
| Pectin (g/100 g) | ND | 2.67 ± 0.67 |
| Ash (g/100 g) | 3.59 ± 0.15 | 0.98 ± 0.04 |
| Total phenols (mg GAE/100 g) | 948.78 ± 39.75 | 2077.03 ± 74.42 |
| Quercetin (mg/100 g) | 1.56 ± 0.56 | ND |
| Quercetin 3-O-rutinoside (mg/100 g) | ND | 0.33 ± 0.07 |
| Quercetin 3-O-arabinoside (mg/100 g) | ND | 2.39 ± 0.30 |
| Quercetin 3-O-xyloside (mg/100 g) | ND | 3.82 ± 0.004 |
| Quercetin 3-O-rhamnoside (mg/100 g) | ND | 5.87 ± 0.05 |
| Quercetin 3-O-glucoside (mg/100 g) | 10.62 ± 0.08 | 2.45 ± 0.60 |
| Quercetin 3-O-galactoside (mg/100 g) | ND | 5.59 ± 0.79 |
| Quercetin 4′-O-glucoside (mg/100 g) | 313.53 ± 2.12 | ND |
| Quercetin 3,4′-di-O-glucoside (mg/100 g) | 1571.16 ± 6.75 | 7.44 ± 0.36 |
| Quercetin 7,4′-O-diglucoside (mg/100 g) | 37.95 ± 0.75 | ND |
| Quercetin 3,7,4′-O-triglucoside (mg/100 g) | 12.29 ± 0.35 | ND |
| Isorhamnetin 3-O-glucoside (mg/100 g) | 0.45 ± 0.001 | ND |
| Isorhamnetin 4′-glucoside (mg/100 g) | 12.95 ± 1.16 | ND |
| Isorhamnetin 3,4′-diglucoside (mg/100 g) | 3.18 ± 1.07 | ND |
| Phloridzin (mg/100 g) | 18.15 ± 1.57 | 6.35 ± 0.45 |
| Phloretin (mg/100 g) | ND | 0.17 ± 0.01 |
| Phloretin 2′-xyloglucoside (mg/100 g) | ND | 3.60 ± 0.14 |
| Epicatechin (mg/100 g) | 10.37 ± 0.35 | 20.81 ± 0.44 |
| Catechin (mg/100 g) | ND | 0.41 ± 0.004 |
| Epicatechin dimer 1 (mg/100 g) | ND | 4.05 ± 0.04 |
| Epicatechin dimer 2 (mg/100 g) | ND | 27.00 ± 0.90 |
| Epigallocatechin (mg/100 g) | ND | 11.18 ± 1.29 |
| Procyanidin B2 (mg/100 g) | ND | 26.84 ± 7.87 |
| Chlorogenic acid (mg/100 g) | ND | 28.15 ± 0.89 |
| Neochlorogenic acid (mg/100 g) | ND | 1.36 ± 0.003 |
| Cryptochlorogenic acid (mg/100 g) | ND | 1.67 ± 0.13 |
| p-Coumaric acid (mg/100 g) | ND | 0.08 ± 0.003 |
| Propionaldehyde (mg/100 g) | 15.59 ± 0.89 | ND |
| 1-Propanethiol (mg/100 g) | 2.50 ± 0.15 | ND |
| Hexanal (mg/100 g) | 0.26 ± 0.02 | 2.17 ± 0.05 |
| 2-Methyl 2-pentenal (mg/100 g) | 0.47 ± 0.05 | ND |
| Propyl thioacetate (mg/100 g) | 0.038 ± 0.0002 | ND |
| Dimethyl trisulphide (mg/100 g) | 0.057 ± 0.0004 | ND |
| Dipropyl disulphide (mg/100 g) | 2.70 ± 0.37 | ND |
| Methyl propyl trisulphide (mg/100 g) | 0.30 ± 0.02 | ND |
| Dipropyl trisulphide (mg/100 g) | 1.00 ± 0.09 | ND |
| Ascorbic acid (mg/100 g) | 58.75 ± 0.30 | 51.33 ± 0.80 |
| Total vitamin C (mg/100 g) | 71.25 ± 0.20 | 82.67 ± 0.90 |
| Peroxidase activity (ΔOD/min/100 g) | 7.66 ± 1.24 | 20.74 ± 0.89 |
| Polyphenoloxidase activity (ΔOD/min/100 g) | 0.38 ± 0.09 | 0.91 ± 0.04 |
| ABTS•+ (μmol TE/100 g) | 1509.45 ± 35.05 | 1821.79 ± 154.65 |
| DPPH• (μmol TE/100 g) | 1129.50 ± 153.22 | 2589.74 ± 185.31 |
| FRAP (μmol TE/100 g) | 1122.55 ± 52.03 | 1582.95 ± 110.57 |
| Component (g/kg) | LC/OC Diet | OO Diet | OA Diet |
|---|---|---|---|
| Onion powder | − | 100 | − |
| Apple powder | − | − | 100 |
| Casein | 200 | 200 | 200 |
| Sucrose | 100 | 100 | 100 |
| Maize starch | 470.49 | 386.14 | 386.14 |
| Soya oil | 50 | 50 | 50 |
| Maize oil | 80 | 80 | 80 |
| Mineral mixture a | 35 | 35 | 35 |
| Vitamin mixture b | 10 | 10 | 10 |
| Cellulose powder | 50 | 34.35 | 34.35 |
| Choline bitartrate | 2.5 | 2.5 | 2.5 |
| tert-butylhydroquinone | 0.01 | 0.01 | 0.01 |
| L-cystine | 2 | 2 | 2 |
| LC | OC | OO | OA | |
|---|---|---|---|---|
| Food intake (g/day/rat) | 18.44 ± 0.35 a | 23.99 ± 1.23 b | 24.98 ± 2.21 b | 31.99 ± 1.07 c |
| Initial body weight (g) | 197.75 ± 17.69 a | 221.88 ± 16.49 b | 225.63 ± 20.59 b | 226.88 ± 19.89 b |
| Final body weight (g) | 357.78 ± 38.45 a | 457.20 ± 20.10 b | 424.40 ± 30.03 b | 462.30 ± 51.55 b |
| Body weight gain (g) | 160.03 ± 27.08 a | 235.33 ± 15.80 b | 198.78 ± 27.68 b | 235.43 ± 40.12 b |
| Food efficiency ratio † | 15.49 ± 2.62 ab | 17.51 ± 1.17 b | 14.21 ± 1.98 a | 13.14 ± 2.24 a |
| Faecal weight (g fresh matter/day) | 2.91 ± 0.39 a | 3.89 ± 0.56 b | 3.77 ± 0.27 b | 4.11 ± 0.52 b |
| Apparent diet digestibility § | 88.01 ± 3.50 a | 89.59 ± 1.72 a | 87.34 ± 1.71 a | 87.43 ± 1.97 a |
| LC | OC | OO | OA | |
|---|---|---|---|---|
| Total cholesterol (mmol/l) | 2.65 ± 0.33 a | 6.93 ± 1.80 c | 4.53 ± 1.18 b | 5.18 ± 1.78 bc |
| HDL-cholesterol (mmol/l) | 1.50 ± 0.22 a | 2.66 ± 0.76 b | 3.52 ± 0.78 c | 2.84 ± 0.39 bc |
| LDL-cholesterol (mmol/l) | 0.28 ± 0.003 a | 1.03 ± 0.11 c | 0.89 ± 0.08 b | 0.98 ± 0.09 bc |
| AI (1) | 0.19 ± 0.02 a | 0.41 ± 0.11 c | 0.26 ± 0.04 b | 0.35 ± 0.04 c |
| AI (2) | 1.78 ± 0.21 b | 2.66 ± 0.45 c | 1.29 ± 0.18 a | 1.80 ± 0.49 b |
| Triacylglycerols (mmol/l) | 3.81 ± 0.82 a | 35.77 ± 3.89 c | 28.05 ± 4.42 b | 32.55 ± 2.82 bc |
| Glucose (mmol/l) | 7.16 ± 1.61 a | 26.20 ± 2.94 c | 17.14 ± 6.74 b | 20.19 ± 7.63 bc |
| Insulin (pmol/l) | 88.31 ± 12.50 a | 651.04 ± 220.96 c | 332.26 ± 124.17 b | 430.25 ± 150.88 bc |
| HOMA-IR | 3.95 ± 0.42 a | 112.26 ± 48.25 d | 31.91 ± 7.22 b | 50.30 ± 14.82 bc |
| Triglyceride–Glucose Index | 9.94 ± 0.34 a | 13.51 ± 0.17 c | 12.76 ± 0.54 b | 13.08 ± 0.47 bc |
| Glucose/Insulin Ratio | 10.57 ± 3.84 a | 5.54 ± 1.80 a | 8.67 ± 6.99 a | 7.40 ± 5.46 a |
| Urea (mmol/l) | 13.39 ± 0.89 a | 12.99 ± 2.96 a | 12.09 ± 3.03 a | 11.20 ± 3.15 a |
| Uric acid (µmol/l) | 31.23 ± 10.42 a | 32.71 ± 11.90 a | 23.79 ± 6.36 a | 23.05 ± 7.41 a |
| Creatinine (µmol/l) | 59.67 ± 4.09 a | 78.45 ± 22.88 a | 75.14 ± 29.51 a | 81.77 ± 34.96 a |
| Albumin (g/l) | 40.75 ± 1.98 a | 55.87 ± 9.52 b | 55.50 ± 9.69 b | 57.62 ± 9.88 b |
| ALT (U/l) | 77.25 ± 19.56 a | 170.12 ± 43.09 b | 90.12 ± 26.35 a | 104.25 ± 23.19 a |
| AST (U/l) | 284.50 ± 55.46 a | 459.12 ±72.97 b | 333.87 ± 40.48 a | 344.37 ± 48.59 a |
| GGT (U/l) | 13.60 ± 1.94 a | 29.42 ± 3.62 c | 22.85 ± 1.78 b | 25.01 ± 1.73 b |
| Total bilirubin (µmol/l) | 18.38 ± 5.31 a | 33.77 ± 9.08 b | 31.85 ± 5.92 b | 26.50 ± 5.71 b |
| LC | OC | OO | OA | |
|---|---|---|---|---|
| Plasma ABTS•+ (μmol TE/L) | 1727.84 ± 85.39 b | 1504.42 ± 44.88 a | 1686.67 ± 24.73 b | 1757.08 ± 77.48 b |
| Plasma FRAP (μmol TE/L) | 828.61 ± 59.87 b | 581.59 ± 54.25 a | 872.99 ± 57.30 b | 843.04 ± 55.98 b |
| Erythrocyte catalase (CAT) activity (nmol/min/mL) | 1759.24 ± 12.06 b | 1696.97 ± 28.65 a | 1788.10 ± 5.18 c | 1696.27 ± 8.69 a |
| Liver catalase (CAT) activity (nmol/min/mg protein) | 390.81 ± 36.24 b | 188.15 ± 6.39 a | 206.02 ± 24.17 a | 194.01 ± 8.20 a |
| Erythrocyte superoxide dismutase (SOD) activity (U/mL) | 117.83 ± 2.87 b | 104.10 ± 2.68 a | 143.07 ± 0.61 c | 123.32 ± 14.50 b |
| Liver superoxide dismutase (SOD) activity (U/mg protein) | 0.014 ± 0.002 c | 0.0042 ± 0.0005 ab | 0.0046 ± 0.004 b | 0.0037 ± 0.001 a |
| Erythrocyte glutathione peroxidase (GPx) activity (nmol/min/mL) | 35.66 ± 3.85 c | 17.93 ± 0.17 a | 25.67 ± 1.02 b | 22.92 ± 2.26 b |
| Liver glutathione peroxidase (GPx) activity (nmol/min/mg protein) | 2.42 ± 0.08 c | 1.61 ± 0.10 a | 2.12 ± 0.11 b | 2.06 ± 0.02 b |
| Erythrocyte glutathione/glutathione disulphide (GSH/GSSH) ratio | 0.49 ± 0.01 c | 0.31 ± 0.03 a | 0.45 ± 0.02 b | 0.52 ± 0.02 d |
| Liver glutathione/glutathione disulphide (GSH/GSSH) ratio | 3.59 ± 0.16 c | 1.75 ± 0.10 a | 2.25 ± 0.12 b | 2.21 ± 0.14 b |
| Plasma protein carbonyls (nmol/mL) | 21.52 ± 1.46 a | 29.89 ± 2.40 c | 25.45 ± 0.84 b | 21.36 ± 1.63 a |
| Liver protein carbonyls (nmol/mL/mg protein) | 0.97 ± 0.10 a | 1.71 ± 0.27 c | 1.28 ± 0.09 b | 1.30 ± 0.05 b |
| Urine 8-hydroxy-2′-deoxyguanosine (ng/mL)-week 0 | 48.33 ± 0.75 aB | 54.19 ± 2.56 bA | 52.32 ± 0.82 bB | 52.97 ± 3.53 bB |
| Urine 8-hydroxy-2′-deoxyguanosine (ng/mL)-week 8 | 45.02 ± 0.32 aA | 60.48 ± 0.64 cB | 46.29 ± 1.89 aA | 49.53 ± 1.08 bA |
| Urine 8-epi-prostaglandin F2α (pg/mL)-week 0 | 6.02 ± 1.05 aA | 7.79 ± 0.16 bA | 7.49 ± 0.41 bA | 7.39 ± 0.64 bA |
| Urine 8-epi-prostaglandin F2α (pg/mL)-week 8 | 12.03 ± 0.59 aB | 27.28 ± 0.32 bB | 11.65 ± 0.42 aB | 11.42 ± 0.73 aB |
| Plasma nitrate/nitrite (µmol/mL) | 55.70 ± 3.20 b | 36.35 ± 2.66 a | 55.53 ± 1.90 b | 55.22 ± 1.67 b |
| Urine nitrate (NO3−)/nitrite (NO2−) (µmol/mL)-week 0 | 1072.78 ± 21.97 bB | 557.50 ± 37.86 aA | 574.17 ± 33.41 aA | 585.10 ± 7.36 aA |
| Urine nitrate (NO3−)/nitrite (NO2−) (µmol/mL)-week 8 | 1024.17 ± 37.86 cA | 789.44 ± 36.00 aB | 1057.50 ± 20.04 cB | 907.50 ± 2.23 bB |
| PAI-1 (ng/mL) | 0.41 ± 0.02 a | 0.54 ± 0.03 c | 0.45 ± 0.03 ab | 0.46 ± 0.03 b |
| TIMP-1 (ng/mL) | 11.05 ± 0.76 a | 15.55 ± 1.38 c | 11.46 ± 1.08 ab | 13.03 ± 1.23 b |
| VEGF (ng/mL) | 0.13 ± 0.01 ab | 0.16 ± 0.01 c | 0.12 ± 0.006 a | 0.13 ± 0.004 b |
| sICAM-1 (ng/mL) | 0.0242 ± 0.001 a | 0.0419 ± 0.002 c | 0.0345 ± 0.002 b | 0.0245 ± 0.002 a |
| sE-Selectin (ng/mL) | 1.15 ± 0.09 a | 1.30 ± 0.05 b | 1.14 ± 0.09 a | 1.06 ± 0.10 a |
| MCP-1 (pg/mL) | 517.90 ± 30.22 ab | 761.27 ± 75.24 c | 570.24 ± 42.80 b | 506.16 ± 35.58 ab |
| Adiponectin (ng/mL) | 15.57 ± 1.05 b | 13.13 ± 0.41 a | 13.83 ± 0.79 a | 13.41 ± 0.92 a |
| GLP-1 (pg/mL) | 80.42 ± 3.65 d | 39.08 ± 2.53 a | 55.20 ± 2.52 c | 47.66 ± 3.60 b |
| Leptin (pg/mL) | 2516.58 ± 232.93 a | 7561.51 ± 529.90 c | 6813.08 ± 351.68 b | 7042.35 ± 274.07 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balderas, C.; Angulo, J.; Sevilleja-Ortiz, A.; Peiró, C.; Vallejo, S.; Dongil, P.; Ancos, B.d.; Sánchez-Moreno, C. Onion and Apple Functional Ingredients Intake Improves Antioxidant and Inflammatory Status and Vascular Injury in Obese Zucker Rats. Antioxidants 2022, 11, 1953. https://doi.org/10.3390/antiox11101953
Balderas C, Angulo J, Sevilleja-Ortiz A, Peiró C, Vallejo S, Dongil P, Ancos Bd, Sánchez-Moreno C. Onion and Apple Functional Ingredients Intake Improves Antioxidant and Inflammatory Status and Vascular Injury in Obese Zucker Rats. Antioxidants. 2022; 11(10):1953. https://doi.org/10.3390/antiox11101953
Chicago/Turabian StyleBalderas, Claudia, Javier Angulo, Alejandro Sevilleja-Ortiz, Concepción Peiró, Susana Vallejo, Pilar Dongil, Begoña de Ancos, and Concepción Sánchez-Moreno. 2022. "Onion and Apple Functional Ingredients Intake Improves Antioxidant and Inflammatory Status and Vascular Injury in Obese Zucker Rats" Antioxidants 11, no. 10: 1953. https://doi.org/10.3390/antiox11101953