
A Large-Scale Multiple Genome Comparison of Acidophilic Archaea (pH ≤ 5.0) Extends Our Understanding of Oxidative Stress Responses in Polyextreme Environments



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Collection
2.2. Phylogeny
2.3. Functional Annotation and Orthologous Groups Identification
2.4. Evolutionary Trajectory Analysis
3. Results
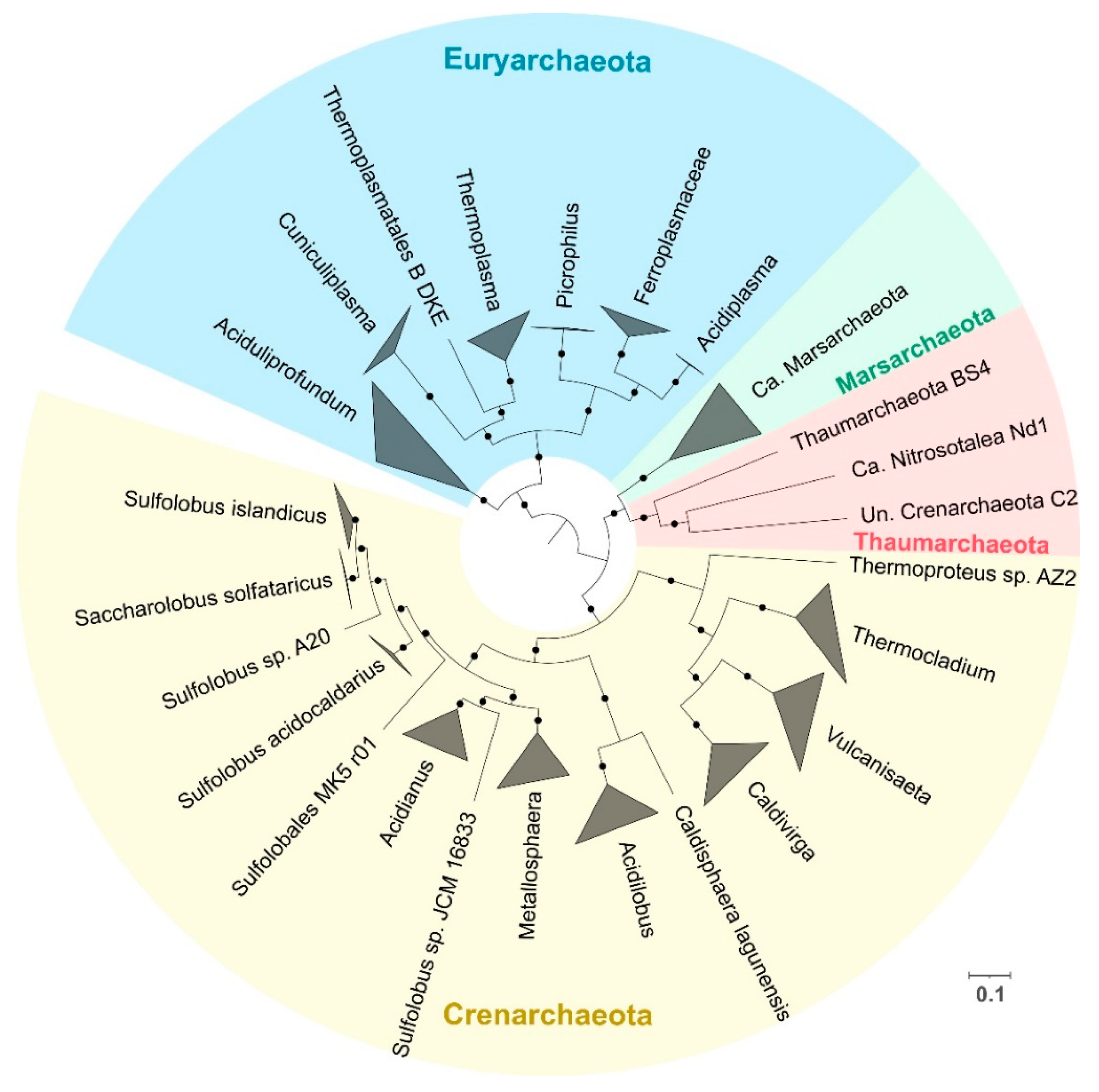
3.1. Phylogeny and Species Clustering
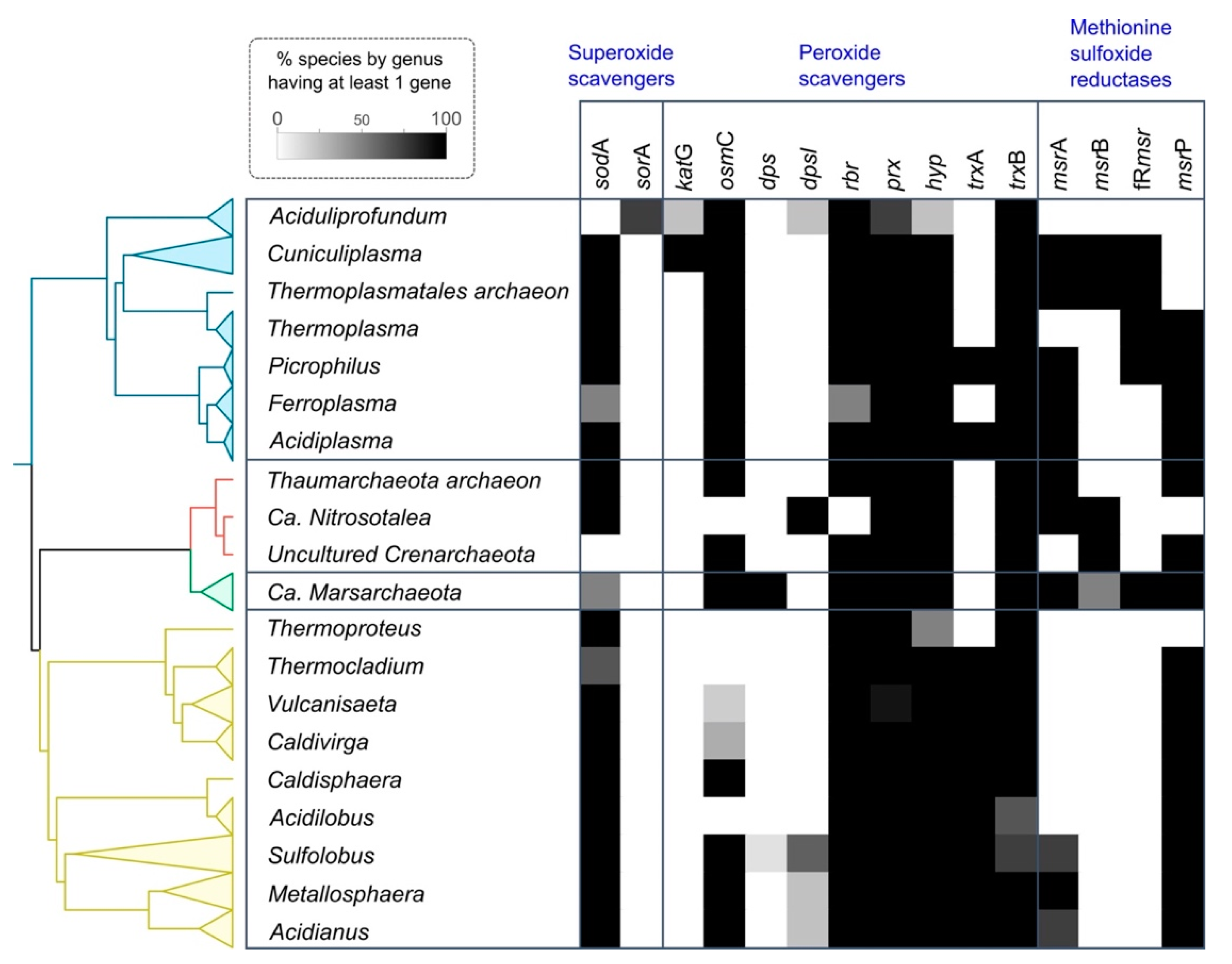
3.2. Superoxide Defense Proteins
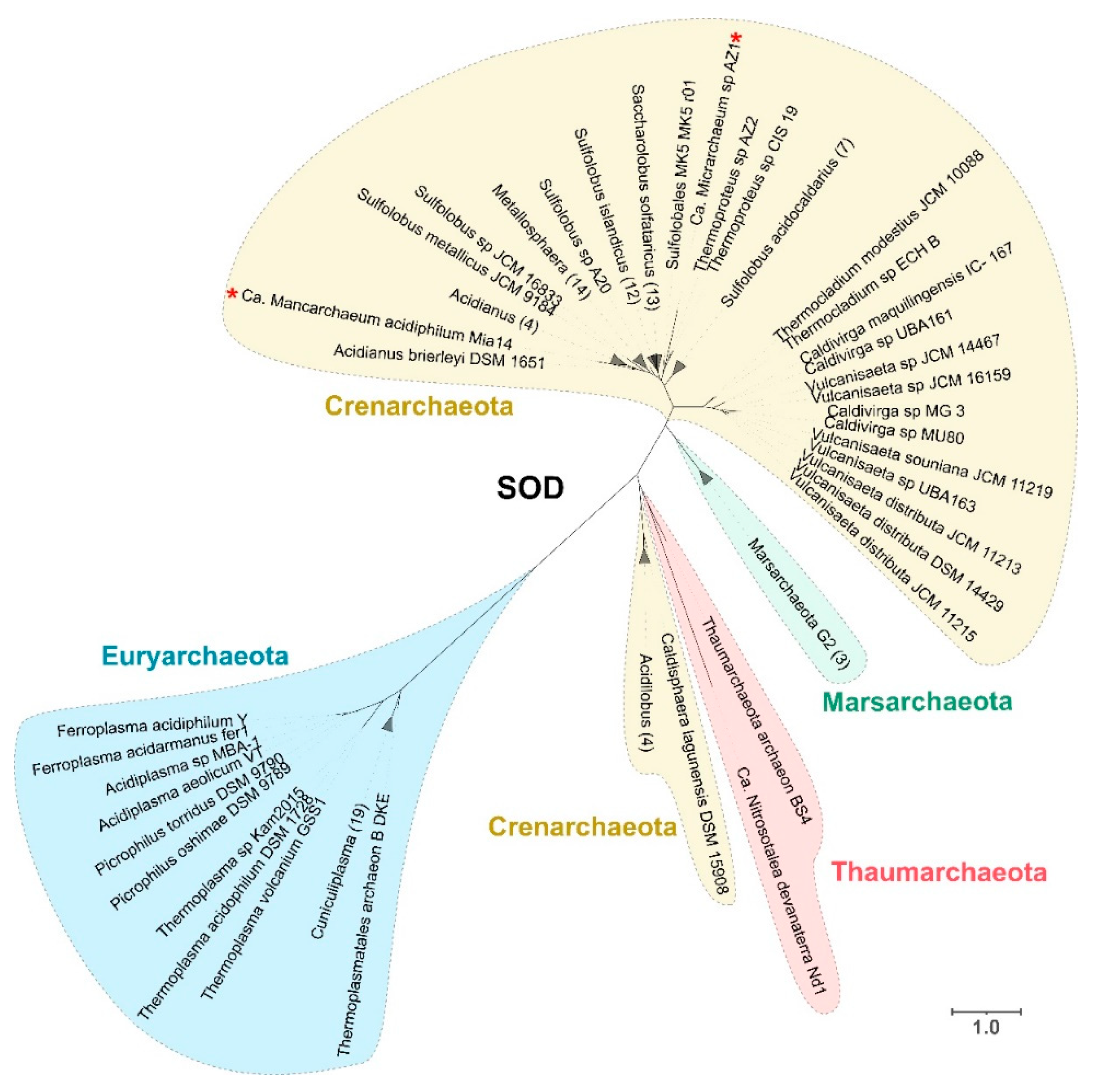
3.2.1. Superoxide Dismutase (SOD)
3.2.2. Superoxide Reductase (SOR)
3.3. Peroxide Scavengers
3.3.1. Catalase
3.3.2. Osmotically Inducible Protein C (OsmC)
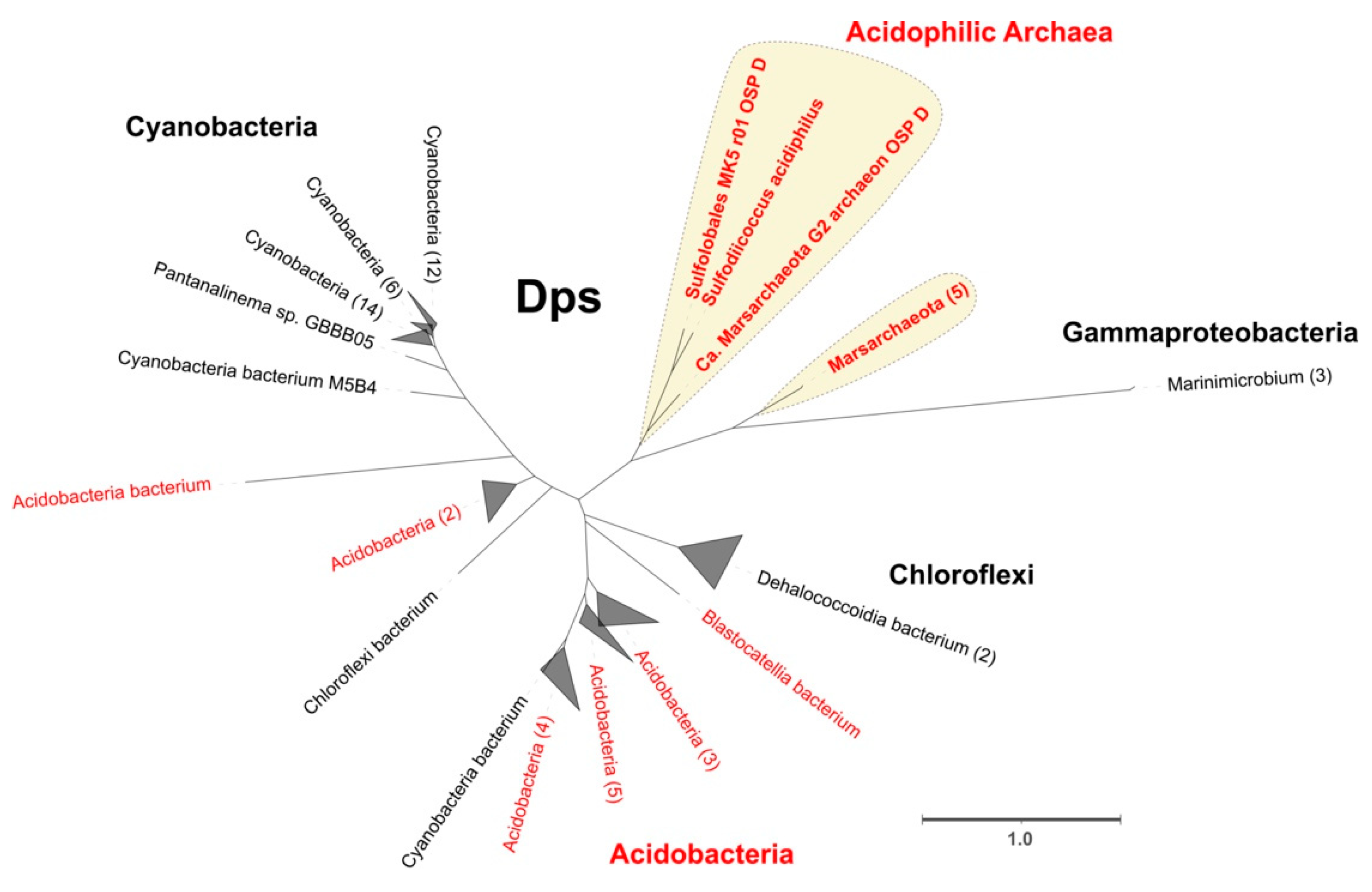
3.3.3. DNA-Binding Protein from Starved Cells
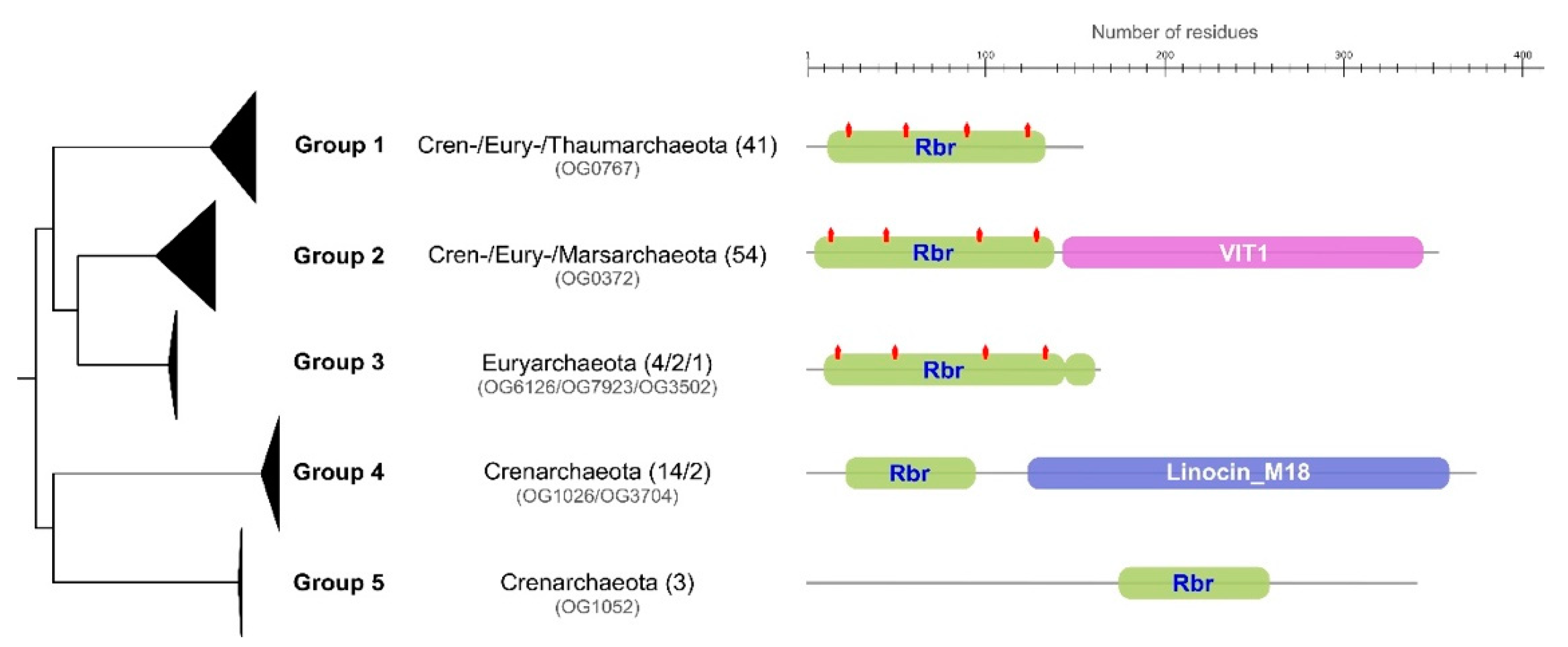
3.3.4. Rubrerythrin (Rbr)
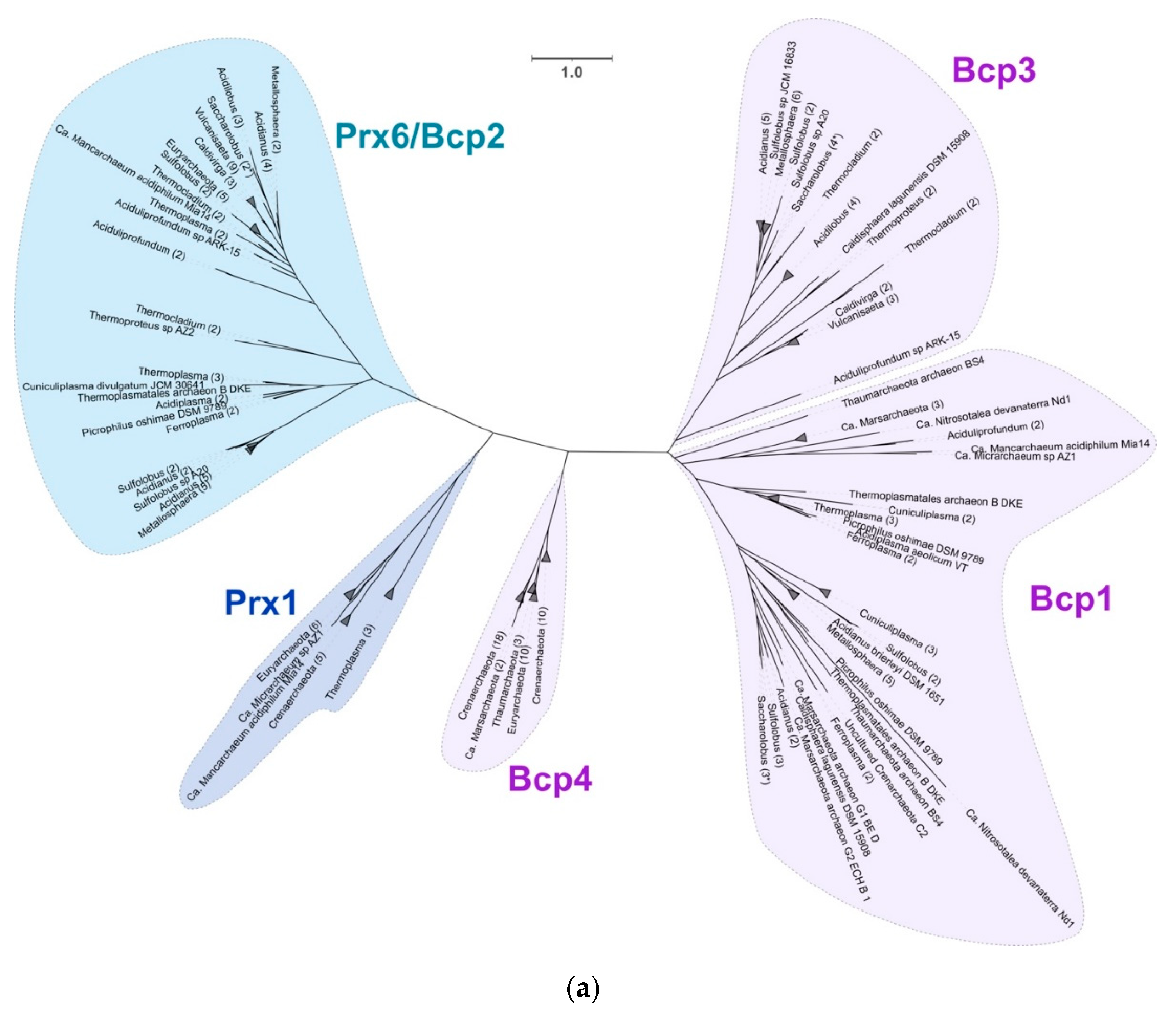
3.3.5. Peroxiredoxins
3.3.6. Thioredoxin
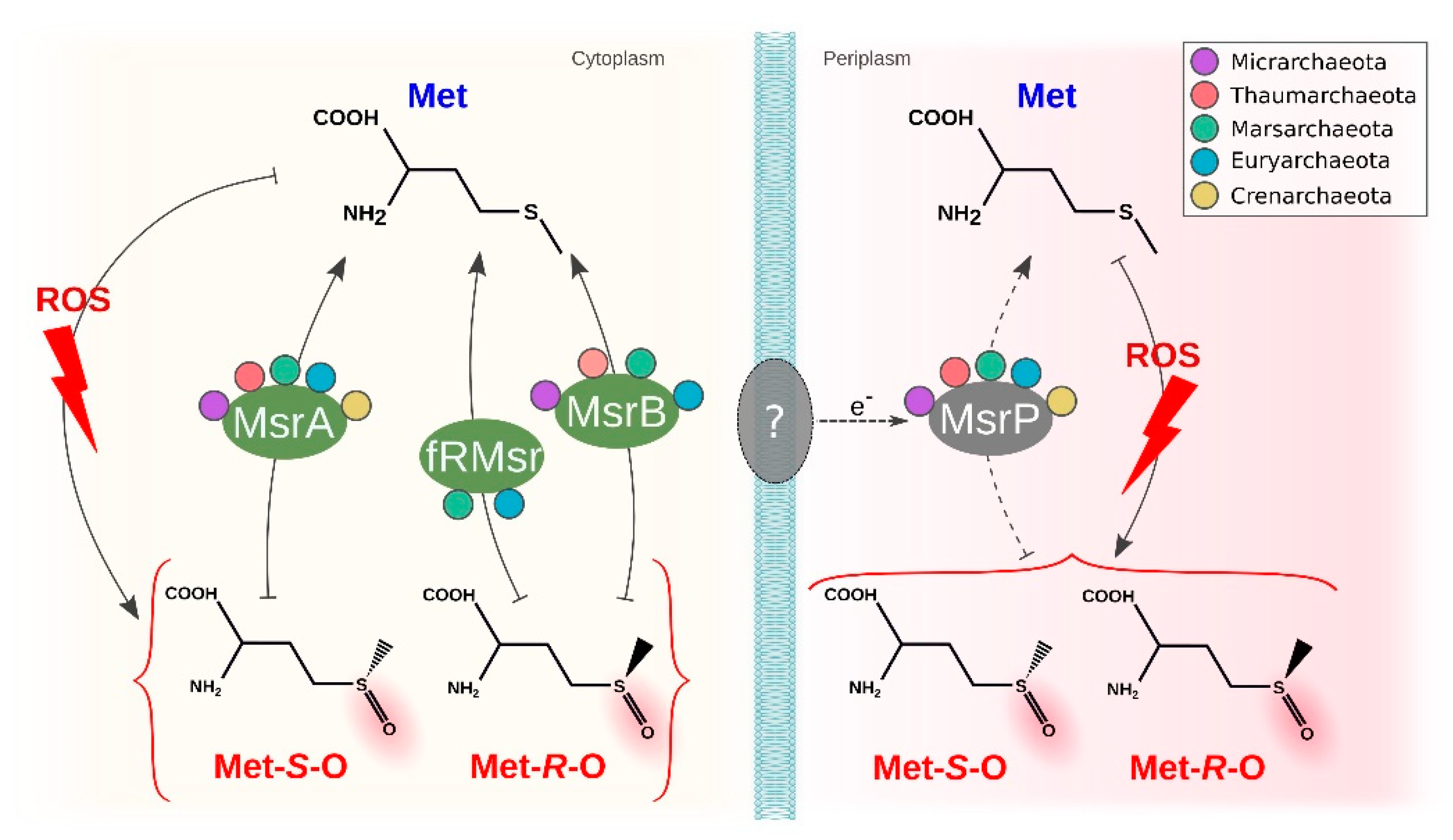
3.4. Methionine Sulfoxide Reductases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Imlay, J.A. Pathways of Oxidative Damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Hemnani, T.; Parihar, M.S. Reactive oxygen species and oxidative DNA damage. Indian J. Physiol. Pharmacol. 1998, 42, 440–452. [Google Scholar] [PubMed]
- Cabiscol, E.; Tamarit, J.; Ros, J. Oxidative Stress in Bacteria and Protein Damage by Reactive Oxygen Species. Int. Microbiol. 2000, 3, 3–8. [Google Scholar] [PubMed]
- Lushchak, V.I. Oxidative Stress and Mechanisms of Protection against It in Bacteria. Biochemistry 2001, 66, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxidative Med. Cell. Longev. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.F. Superoxide Dismutases: Ancient Enzymes and New Insights. FEBS Lett. 2012, 586, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Zámocký, M.; Gasselhuber, B.; Furtmüller, P.G.; Obinger, C. Molecular Evolution of Hydrogen Peroxide Degrading Enzymes. Arch. Biochem. Biophys. 2012, 525, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Farr, S.B.; Kogoma, T. Oxidative Stress Responses in Escherichia Coli and Salmonella Typhimurium. Microbiol. Rev. 1991, 55, 561–585. [Google Scholar] [CrossRef] [PubMed]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.F. Oxidative Stress, Protein Damage and Repair in Bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Johnson, L.A.; Hug, L.A. Distribution of Reactive Oxygen Species Defense Mechanisms across Domain Bacteria. Free Radic. Biol. Med. 2019, 140, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Gribaldo, S.; Brochier-Armanet, C. The Origin and Evolution of Archaea: A State of the Art. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1007–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozubal, M.A.; Romine, M.; Jennings, R.D.; Jay, Z.J.; Tringe, S.G.; Rusch, D.B.; Beam, J.P.; McCue, L.A.; Inskeep, W.P. Geoarchaeota: A New Candidate Phylum in the Archaea from High-Temperature Acidic Iron Mats in Yellowstone National Park. ISME J. 2013, 7, 622–634. [Google Scholar] [CrossRef]
- Lane, N. The Evolution of Oxidative Stress. In Principles of Free Radical Biomedicine; Nova Science Publishers: New York, NY, USA, 2011; Volume 1, pp. 1–17. ISBN 9781612097732. [Google Scholar]
- Anbar, A.D.; Duan, Y.; Lyons, T.W.; Arnold, G.L.; Kendall, B.; Creaser, R.A.; Kaufman, A.J.; Gordon, G.W.; Scott, C.; Garvin, J.; et al. A Whiff of Oxygen before the Great Oxidation Event? Science 2007, 317, 1903–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, M.C.; Romão, C.V.; Folgosa, F.; Borges, P.T.; Frazão, C.; Teixeira, M. How Superoxide Reductases and Flavodiiron Proteins Combat Oxidative Stress in Anaerobes. Free Radic. Biol. Med. 2019, 140, 36–60. [Google Scholar] [CrossRef]
- Fischer, W.W.; Hemp, J.; Valentine, J.S. How Did Life Survive Earth’s Great Oxygenation? Curr. Opin. Chem. Biol. 2016, 31, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Stan-Lotter, H.; Fendrihan, S. Halophilic Archaea: Life with Desiccation, Radiation and Oligotrophy over Geological Times. Life 2015, 5, 1487–1496. [Google Scholar] [CrossRef] [PubMed]
- Korzhenkov, A.A.; Toshchakov, S.V.; Bargiela, R.; Gibbard, H.; Ferrer, M.; Teplyuk, A.V.; Jones, D.L.; Kublanov, I.V.; Golyshin, P.N.; Golyshina, O.V. Archaea Dominate the Microbial Community in an Ecosystem with Low-to-Moderate Temperature and Extreme Acidity. Microbiome 2019, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Blöchl, E.; Rachel, R.; Burggraf, S.; Hafenbradl, D.; Jannasch, H.W.; Stetter, K.O. Pyrolobus Fumarii, Gen. and Sp. Nov., Represents a Novel Group of Archaea, Extending the Upper Temperature Limit for Life to 113 °C. Extremophiles 1997, 1, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Pedone, E.; Fiorentino, G.; Bartolucci, S.; Limauro, D. Enzymatic Antioxidant Signatures in Hyperthermophilic Archaea. Antioxidants 2020, 9, 703. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Dopson, M. Life in Acid: PH Homeostasis in Acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef]
- Dopson, M. Physiological and Phylogenetic Diversity of Acidophilic Bacteria. In Acidophiles: Life in Extremely Acidic Environments; Caister Academic Press: Poole, UK, 2016; pp. 79–92. [Google Scholar]
- Benison, K.C.; O’Neill, W.K.; Blain, D.; Hallsworth, J.E. Water Activities of Acid Brine Lakes Approach the Limit for Life. Astrobiology 2021, 21, 729–740. [Google Scholar] [CrossRef]
- Johnson, D.B. Physiology and Ecology of Acidophilic Microorganisms. In Physiology and Biochemistry of Extremophiles; ASM Press: Washington, DC, USA, 2014; pp. 255–270. [Google Scholar]
- Slonczewski, J.L.; Fujisawa, M.; Dopson, M.; Krulwich, T.A. Cytoplasmic PH Measurement and Homeostasis in Bacteria and Archaea. Adv. Microb. Physiol. 2009, 55, 1–317. [Google Scholar] [CrossRef] [PubMed]
- Orellana, L.H.; Jerez, C.A. A Genomic Island Provides Acidithiobacillus Ferrooxidans ATCC 53993 Additional Copper Resistance: A Possible Competitive Advantage. Appl. Microbiol. Biotechnol. 2011, 92, 761–767. [Google Scholar] [CrossRef]
- Orell, A.; Remonsellez, F.; Arancibia, R.; Jerez, C.A. Molecular Characterization of Copper and Cadmium Resistance Determinants in the Biomining Thermoacidophilic Archaeon Sulfolobus Metallicus. Archaea 2013, 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dopson, M.; Holmes, D.S. Metal Resistance in Acidophilic Microorganisms and Its Significance for Biotechnologies. Appl. Microbiol. Biotechnol. 2014, 98, 8133–8144. [Google Scholar] [CrossRef]
- Król, A.; Mizerna, K.; Bożym, M. An Assessment of PH-Dependent Release and Mobility of Heavy Metals from Metallurgical Slag. J. Hazard. Mater. 2020, 384, 121502. [Google Scholar] [CrossRef]
- Osorio, H.; Martínez, V.; Veloso, F.A.; Pedroso, I.; Valdés, J.; Jedlicki, E.; Holmes, D.S.; Quatrini, R. Iron Homeostasis Strategies in Acidophilic Iron Oxidizers: Studies in Acidithiobacillus and Leptospirillum. Hydrometallurgy 2008, 94, 175–179. [Google Scholar] [CrossRef]
- Jeon, S.J.; Ishikawa, K. Identification and Characterization of Thioredoxin and Thioredoxin Reductase from Aeropyrum Pernix K1. Eur. J. Biochem. 2002, 269, 5423–5430. [Google Scholar] [CrossRef]
- Amo, T.; Atomi, H.; Imanaka, T. Biochemical Properties and Regulated Gene Expression of the Superoxide Dismutase from the Facultatively Aerobic Hyperthermophile Pyrobaculum Calidifontis. J. Bacteriol. 2003, 185, 6340–6347. [Google Scholar] [CrossRef] [Green Version]
- Dillard, B.D.; Demick, J.M.; Adams, M.W.W.; Lanzilotta, W.N. A Cryo-Crystallographic Time Course for Peroxide Reduction by Rubrerythrin from Pyrococcus Furiosus. J. Biol. Inorg. Chem. 2011, 16, 949–959. [Google Scholar] [CrossRef]
- Fütterer, O.; Angelov, A.; Liesegang, H.; Gottschalk, G.; Schleper, C.; Schepers, B.; Dock, C.; Antranikian, G.; Liebl, W. Genome Sequence of Picrophilus Torridus and Its Implications for Life around pH 0. Proc. Natl. Acad. Sci. USA 2004, 101, 9091–9096. [Google Scholar] [CrossRef] [Green Version]
- Neira, G.; Cortez, D.; Jil, J.; Holmes, D.S. AciDB 1.0: A Database of Acidophilic Organisms, Their Genomic Information and Associated Metadata. Bioinformatics 2020, 36, 4970–4971. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI Reference Sequences (RefSeq): A Curated Non-Redundant Sequence Database of Genomes, Transcripts and Proteins. Nucleic Acids Res. 2007, 35, D61–D65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Asnicar, F.; Thomas, A.M.; Beghini, F.; Mengoni, C.; Manara, S.; Manghi, P.; Zhu, Q.; Bolzan, M.; Cumbo, F.; May, U.; et al. Precise Phylogenetic Analysis of Microbial Isolates and Genomes from Metagenomes Using PhyloPhlAn 3.0. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2014, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High Throughput ANI Analysis of 90K Prokaryotic Genomes Reveals Clear Species Boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The Protein Families Database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, S.; Apweiler, R.; Attwood, T.K.; Bairoch, A.; Bateman, A.; Binns, D.; Bork, P.; Das, U.; Daugherty, L.; Duquenne, L.; et al. InterPro: The Integrative Protein Signature Database. Nucleic Acids Res. 2009, 37, D211–D215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale Protein Function Classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic Orthology Inference for Comparative Genomics. Genome Biol. 2019, 20, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.; Maini, P.K. DendroBLAST: Approximate Phylogenetic Trees in the Absence of Multiple Sequence Alignments. PLoS ONE 2013, 8, e58537. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved Protein Subcellular Localization Prediction with Refined Localization Subcategories and Predictive Capabilities for All Prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Ravenhall, M.; Škunca, N.; Lassalle, F.; Dessimoz, C. Inferring Horizontal Gene Transfer. PLoS Comput. Biol. 2015, 11, e1004095. [Google Scholar] [CrossRef] [Green Version]
- Nelson, K.E.; Clayton, R.A.; Gill, S.R.; Gwinn, M.L.; Dodson, R.J.; Haft, D.H.; Hickey, E.K.; Peterson, J.D.; Nelson, W.C.; Ketchum, K.A.; et al. Evidence for Lateral Gene Transfer between Archaea and Bacteria from Genome Sequence of Thermotoga Maritima. Nature 1999, 399, 323–329. [Google Scholar] [CrossRef]
- Koonin, E.V.; Makarova, K.S.; Aravind, L. Horizontal Gene Transfer in Prokaryotes: Quantification and Classification. Annu. Rev. Microbiol. 2001, 55, 709–742. [Google Scholar] [CrossRef]
- Maglott, D.; Ostell, J.; Pruitt, K.D.; Tatusova, T. Entrez Gene: Gene-Centered Information at NCBI. Nucleic Acids Res. 2010, 39, D52–D57. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Harrison, K.J.; de Crécy-Lagard, V.; Zallot, R. Gene Graphics: A Genomic Neighborhood Data Visualization Web Application. Bioinformatics 2018, 34, 1406–1408. [Google Scholar] [CrossRef] [PubMed]
- Golyshina, O.V.; Toshchakov, S.V.; Makarova, K.S.; Gavrilov, S.N.; Korzhenkov, A.A.; la Cono, V.; Arcadi, E.; Nechitaylo, T.Y.; Ferrer, M.; Kublanov, I.V.; et al. “ARMAN” Archaea Depend on Association with Euryarchaeal Host in Culture and In Situ. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Murray, A.E.; Freudenstein, J.; Gribaldo, S.; Hatzenpichler, R.; Hugenholtz, P.; Kämpfer, P.; Konstantinidis, K.T.; Lane, C.E.; Papke, R.T.; Parks, D.H.; et al. Roadmap for Naming Uncultivated Archaea and Bacteria. Nat. Microbiol. 2020, 5, 987–994. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Chaumeil, P.A.; Rinke, C.; Mussig, A.J.; Hugenholtz, P. A Complete Domain-to-Species Taxonomy for Bacteria and Archaea. Nat. Biotechnol. 2020, 38, 1079–1086. [Google Scholar] [CrossRef]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.F.; Teixeira, M.; Valentine, J.S. Superoxide Dismutases and Superoxide Reductases. Chem. Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef]
- dello Russo, A.; Rullo, R.; Nitti, G.; Masullo, M.; Bocchini, V. Iron Superoxide Dismutase from the Archaeon Sulfolobus Solfataricus: Average Hydrophobicity and Amino Acid Weight Are Involved in the Adaptation of Proteins to Extreme Environments. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1997, 1343, 23–30. [Google Scholar] [CrossRef]
- Ferrer, M.; Golyshina, O.V.; Beloqui, A.; Golyshin, P.N.; Timmis, K.N. The Cellular Machinery of Ferroplasma Acidiphilum Is Iron-Protein-Dominated. Nature 2007, 445, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Wheaton, G.H.; Mukherjee, A.; Kelly, R.M. Transcriptomes of the Extremely Thermoacidophilic Archaeon Metallosphaera Sedula Exposed to Metal “Shock” Reveal Generic and Specific Metal Responses. Appl. Environ. Microbiol. 2016, 82, 4613–4627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamano, S.; Sako, Y.; Nomura, N.; Maruyama, T. A Cambialistic SOD in a Strictly Aerobic Hyperthermophilic Archaeon, Aeropyrum Pernix. J. Biochem. 1999, 126, 218–225. [Google Scholar] [CrossRef]
- Whittaker, M.M.; Whittaker, J.W. Recombinant Superoxide Dismutase from a Hyperthermophilic Archaeon, Pyrobaculum Aerophilium. J. Biol. Inorg. Chem. JBIC A Publ. Soc. Biol. Inorg. Chem. 2000, 5, 402–408. [Google Scholar]
- Jay, Z.J.; Beam, J.P.; Dlakić, M.; Rusch, D.B.; Kozubal, M.A.; Inskeep, W.P. Marsarchaeota Are an Aerobic Archaeal Lineage Abundant in Geothermal Iron Oxide Microbial Mats. Nat. Microbiol. 2018, 3, 732–740. [Google Scholar] [CrossRef]
- Dombrowski, N.; Lee, J.H.; Williams, T.A.; Offre, P.; Spang, A. Genomic Diversity, Lifestyles and Evolutionary Origins of DPANN Archaea. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef] [Green Version]
- Dombrowski, N.; Williams, T.A.; Sun, J.; Woodcroft, B.J.; Lee, J.H.; Minh, B.Q.; Rinke, C.; Spang, A. Undinarchaeota Illuminate DPANN Phylogeny and the Impact of Gene Transfer on Archaeal Evolution. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Lucchetti-Miganeh, C.; Goudenège, D.; Thybert, D.; Salbert, G.; Barloy-Hubler, F. SORGOdb: Superoxide Reductase Gene Ontology Curated DataBase. BMC Microbiol. 2011, 11, 105. [Google Scholar] [CrossRef]
- Nivière, V.; Fontecave, M. Discovery of Superoxide Reductase: An Historical Perspective. J. Biol. Inorg. Chem. 2004, 9, 119–123. [Google Scholar] [CrossRef]
- Grunden, A.M.; Jenney, F.E.; Ma, K.; Ji, M.; Weinberg, M.V.; Adams, M.W.W. In Vitro Reconstitution of an NADPH-Dependent Superoxide Reduction Pathway from Pyrococcus Furiosus. Appl. Environ. Microbiol. 2005, 71, 1522–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cárdenas, J.P.; Moya, F.; Covarrubias, P.; Shmaryahu, A.; Levicán, G.; Holmes, D.S.; Quatrini, R. Comparative Genomics of the Oxidative Stress Response in Bioleaching Microorganisms. Hydrometallurgy 2012, 127–128, 162–167. [Google Scholar] [CrossRef]
- Zamocky, M.; Furtmüller, P.G.; Obinger, C. Evolution of Catalases from Bacteria to Humans. Antioxid. Redox Signal. 2008, 10, 1527–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passardi, F.; Zamocky, M.; Favet, J.; Jakopitsch, C.; Penel, C.; Obinger, C.; Dunand, C. Phylogenetic Distribution of Catalase-Peroxidases: Are There Patches of Order in Chaos? Gene 2007, 397, 101–113. [Google Scholar] [CrossRef]
- Saikolappan, S.; Das, K.; Sasindran, S.J.; Jagannath, C.; Dhandayuthapani, S. OsmC Proteins of Mycobacterium Tuberculosis and Mycobacterium Smegmatis Protect against Organic Hydroperoxide Stress. Tuberculosis 2011, 91, S119–S127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.C.; Pham, B.P.; van Duyet, L.; Jia, B.; Lee, S.; Yu, R.; Woo Han, S.; Yang, J.K.; Hahm, K.S.; Cheong, G.W. Structural and Functional Characterization of Osmotically Inducible Protein C (OsmC) from Thermococcus Kodakaraensis KOD1. Biochim. Biophys. Acta Proteins Proteom. 2008, 1784, 783–788. [Google Scholar] [CrossRef]
- Gauss, G.H.; Benas, P.; Wiedenheft, B.; Young, M.; Douglas, T.; Lawrence, C.M. Structure of the DPS-like Protein from Sulfolobus Solfataricus Reveals a Bacterioferritin-like Dimetal Binding Site within a DPS-like Dodecameric Assembly. Biochemistry 2006, 45, 10815–10827. [Google Scholar] [CrossRef] [Green Version]
- Wiedenheft, B.; Mosolf, J.; Willits, D.; Yeager, M.; Dryden, K.A.; Young, M.; Douglas, T. An Archaeal Antioxidant: Characterization of a Dps-like Protein from Sulfolobus Solfataricus. Proc. Natl. Acad. Sci. USA 2005, 102, 10551–10556. [Google Scholar] [CrossRef] [Green Version]
- Wiedenbeck, J.; Cohan, F.M. Origins of Bacterial Diversity through Horizontal Genetic Transfer and Adaptation to New Ecological Niches. FEMS Microbiol. Rev. 2011, 35, 957–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinberg, M.V.; Jenney, F.E.; Cui, X.; Adams, M.W.W. Rubrerythrin from the Hyperthermophilic Archaeon Pyrococcus Furiosus Is a Rubredoxin-Dependent, Iron-Containing Peroxidase. J. Bacteriol. 2004, 186, 7888–7895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.A.; Punshon, T.; Lanzirotti, A.; Li, A.; Alonso, J.M.; Ecker, J.R.; Kaplan, J.; Guerinot, M. lou Localization of Iron in Arabidopsis Seed Requires the Vacuolar Membrane Transporter VIT1. Science 2006, 314, 1295–1298. [Google Scholar] [CrossRef]
- Cardenas, J.P.; Quatrini, R.; Holmes, D.S. Aerobic Lineage of the Oxidative Stress Response Protein Rubrerythrin Emerged in an Ancient Microaerobic, (Hyper)Thermophilic Environment. Front. Microbiol. 2016, 7, 1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, T.; Kumazaki, K.; Wada, M.; Taniguchi, R.; Nakane, T.; Yamashita, K.; Hirata, K.; Ishitani, R.; Ito, K.; Nishizawa, T.; et al. Crystal Structure of Plant Vacuolar Iron Transporter VIT1. Nat. Plants 2019, 5, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Sorribes-Dauden, R.; Peris, D.; Martínez-Pastor, M.T.; Puig, S. Structure and Function of the Vacuolar Ccc1/VIT1 Family of Iron Transporters and Its Regulation in Fungi. Comput. Struct. Biotechnol. J. 2020, 18, 3712–3722. [Google Scholar] [CrossRef] [PubMed]
- Valdes-Stauber, N.; Scherer, S. Isolation and Characterization of Linocin M18, a Bacteriocin Produced by Brevibacterium Linens. Appl. Environ. Microbiol. 1994, 60, 3809–3814. [Google Scholar] [CrossRef] [Green Version]
- Soito, L.; Williamson, C.; Knutson, S.T.; Fetrow, J.S.; Poole, L.B.; Nelson, K.J. PREX: PeroxiRedoxin Classification IndEX, a Database of Subfamily Assignments across the Diverse Peroxiredoxin Family. Nucleic Acids Res. 2011, 39, D332–D337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limauro, D.; Pedone, E.; Galdi, I.; Bartolucci, S. Peroxiredoxins as Cellular Guardians in Sulfolobus Solfataricus- Characterization of Bcp1, Bcp3 and Bcp4. FEBS J. 2008, 275, 2067–2077. [Google Scholar] [CrossRef]
- D’Ambrosio, K.; Limauro, D.; Pedone, E.; Galdi, I.; Pedone, C.; Bartolucci, S.; de Simone, G. Insights into the Catalytic Mechanism of the Bcp Family: Functional and Structural Analysis of Bcp1 from Sulfolobus Solfataricus. Proteins Struct. Funct. Bioinform. 2009, 76, 995–1006. [Google Scholar] [CrossRef]
- Sun, C.C.; Dong, W.R.; Shao, T.; Li, J.Y.; Zhao, J.; Nie, L.; Xiang, L.X.; Zhu, G.; Shao, J.Z. Peroxiredoxin 1 (Prx1) Is a Dual-Function Enzyme by Possessing Cys-Independent Catalase-like Activity. Biochem. J. 2017, 474, 1373–1394. [Google Scholar] [CrossRef] [Green Version]
- Bang, Y.J.; Oh, M.H.; Choi, S.H. Distinct Characteristics of Two 2-Cys Peroxiredoxins of Vibrio Vulnificus Suggesting Differential Roles in Detoxifying Oxidative Stress. J. Biol. Chem. 2012, 287, 42516–42524. [Google Scholar] [CrossRef] [Green Version]
- Groitl, B.; Jakob, U. Thiol-Based Redox Switches. Biochim. Biophys. Acta Proteins Proteom. 2014, 1844, 1335–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maupin-Furlow, J.A. Methionine Sulfoxide Reductases of Archaea. Antioxidants 2018, 7, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Kwak, G.H.; Lee, K.; Jo, C.H.; Hwang, K.Y.; Kim, H.Y. Structural and Biochemical Analysis of a Type II Free Methionine-R- Sulfoxide Reductase from Thermoplasma Acidophilum. Arch. Biochem. Biophys. 2014, 560, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Juillan-Binard, C.; Picciocchi, A.; Andrieu, J.P.; Dupuy, J.; Petit-Hartlein, I.; Caux-Thang, C.; Vivès, C.; Nivière, V.; Fieschi, F. A Two-Component NADPH Oxidase (NOX)-like System in Bacteria Is Involved in the Electron Transfer Chain to the Methionine Sulfoxide Reductase MsrP. J. Biol. Chem. 2017, 292, 2485–2494. [Google Scholar] [CrossRef] [Green Version]
- Case, A.J. On the Origin of Superoxide Dismutase: An Evolutionary Perspective of Superoxide-Mediated Redox Signaling. Antioxidants 2017, 6, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenney, F.E.; Verhagen, M.F.J.M.; Cui, X.; Adams, M.W.W. Anaerobic Microbes: Oxygen Detoxification Without Superoxide Dismutase. Science 1999, 286, 306–309. [Google Scholar] [CrossRef]
- Koonin, E.V.; Wolf, Y.I. Genomics of Bacteria and Archaea: The Emerging Dynamic View of the Prokaryotic World. Nucleic Acids Res. 2008, 36, 6688–6719. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.; Whitaker, R.J.; Krause, D.J.; Heilers, J.H.; van Wolferen, M.; van der Does, C.; Albers, S.V. Mechanisms of Gene Flow in Archaea. Nat. Rev. Microbiol. 2017, 15, 492–501. [Google Scholar] [CrossRef]
- Beard, S.; Ossandon, F.J.; Rawlings, D.E.; Quatrini, R. The Flexible Genome of Acidophilic Prokaryotes. Curr. Issues Mol. Biol. 2020, 40, 231–266. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, X.; Liang, Y.; Guo, X.; Xiao, Y.; Ma, L.; Miao, B.; Liu, H.; Peng, D.; Huang, W.; et al. Adaptive Evolution of Extreme Acidophile Sulfobacillus Thermosulfidooxidans Potentially Driven by Horizontal Gene Transfer and Gene Loss. Appl. Environ. Microbiol. 2017, 83, e03098-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergara, E.; Neira, G.; González, C.; Cortez, D.; Dopson, M.; Holmes, D.S. Evolution of Predicted Acid Resistance Mechanisms in the Extremely Acidophilic Leptospirillum Genus. Genes 2020, 11, 389. [Google Scholar] [CrossRef] [Green Version]
- Chiancone, E.; Ceci, P. The Multifaceted Capacity of Dps Proteins to Combat Bacterial Stress Conditions: Detoxification of Iron and Hydrogen Peroxide and DNA Binding. Biochim. Biophys. Acta Gen. Subj. 2010, 1800, 798–805. [Google Scholar] [CrossRef]
- Yuan, F.; Yin, S.; Xu, Y.; Xiang, L.; Wang, H.; Li, Z.; Fan, K.; Pan, G. The Richness and Diversity of Catalases in Bacteria. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Canfield, D.E.; Rosing, M.T.; Bjerrum, C. Early Anaerobic Metabolisms. Proc. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1819–1836. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.F.; Sousa, F.L. Early Microbial Evolution: The Age of Anaerobes. Cold Spring Harb. Perspect. Biol. 2016, 8, a018127. [Google Scholar] [CrossRef]
- Ouzounis, C.A.; Kunin, V.; Darzentas, N.; Goldovsky, L.A. Minimal Estimate for the Gene Content of the Last Universal Common Ancestor—Exobiology from a Terrestrial Perspective. Res. Microbiol. 2006, 157, 57–68. [Google Scholar] [CrossRef]
- Ślesak, I.; Ślesak, H.; Kruk, J. RubisCO Early Oxygenase Activity: A Kinetic and Evolutionary Perspective. BioEssays 2017, 39, 1700071. [Google Scholar] [CrossRef] [PubMed]
- Planavsky, N.J.; Asael, D.; Hofmann, A.; Reinhard, C.T.; Lalonde, S.V.; Knudsen, A.; Wang, X.; Ossa Ossa, F.; Pecoits, E.; Smith, A.J.B.; et al. Evidence for Oxygenic Photosynthesis Half a Billion Years before the Great Oxidation Event. Nat. Geosci. 2014, 7, 283–286. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neira, G.; Vergara, E.; Cortez, D.; Holmes, D.S. A Large-Scale Multiple Genome Comparison of Acidophilic Archaea (pH ≤ 5.0) Extends Our Understanding of Oxidative Stress Responses in Polyextreme Environments. Antioxidants 2022, 11, 59. https://doi.org/10.3390/antiox11010059
Neira G, Vergara E, Cortez D, Holmes DS. A Large-Scale Multiple Genome Comparison of Acidophilic Archaea (pH ≤ 5.0) Extends Our Understanding of Oxidative Stress Responses in Polyextreme Environments. Antioxidants. 2022; 11(1):59. https://doi.org/10.3390/antiox11010059
Chicago/Turabian StyleNeira, Gonzalo, Eva Vergara, Diego Cortez, and David S. Holmes. 2022. "A Large-Scale Multiple Genome Comparison of Acidophilic Archaea (pH ≤ 5.0) Extends Our Understanding of Oxidative Stress Responses in Polyextreme Environments" Antioxidants 11, no. 1: 59. https://doi.org/10.3390/antiox11010059