
Glycoside Hydrolases and Non-Enzymatic Glycation Inhibitory Potential of Viburnum opulus L. Fruit—In Vitro Studies


Abstract
:1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
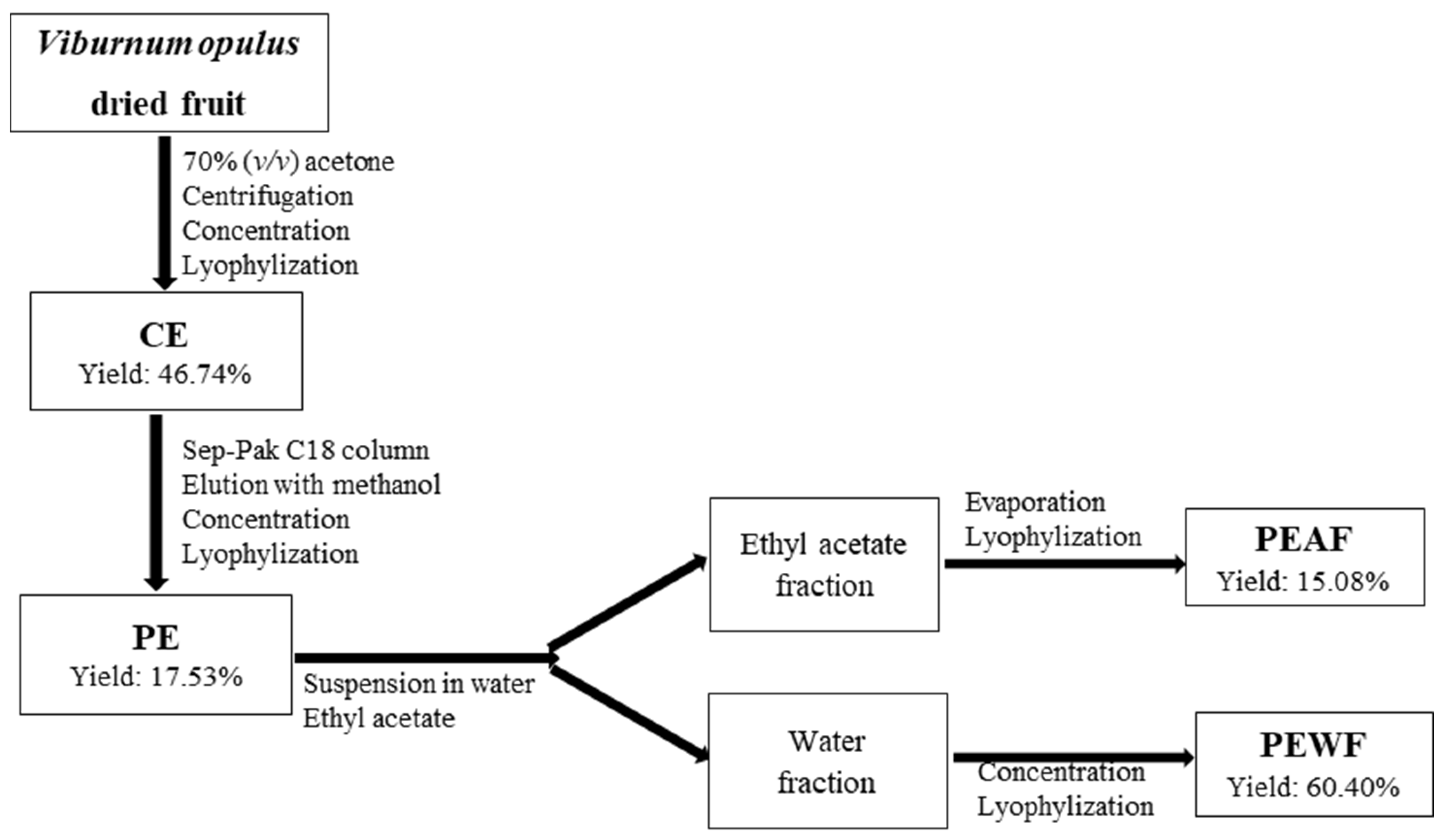
2.2. Plant Material and Samples Preparation
2.3. Quantification of Individual Phenolic Compounds
2.4. Total Proanthocyanidins Content
2.5. α-Amylase Inhibition Assay
2.6. α-Glucosidase Inhibition Assay
2.7. Mode of Enzyme Inhibition
2.8. Fluorescence Measurements
2.9. Protein Glycation Inhibition Assay
2.10. In Vitro Antioxidant Assays
2.11. Statistical Analysis
3. Results
3.1. Effects of V. opulus Fruit Components on α-Amylase and α-Glucosidase Activity
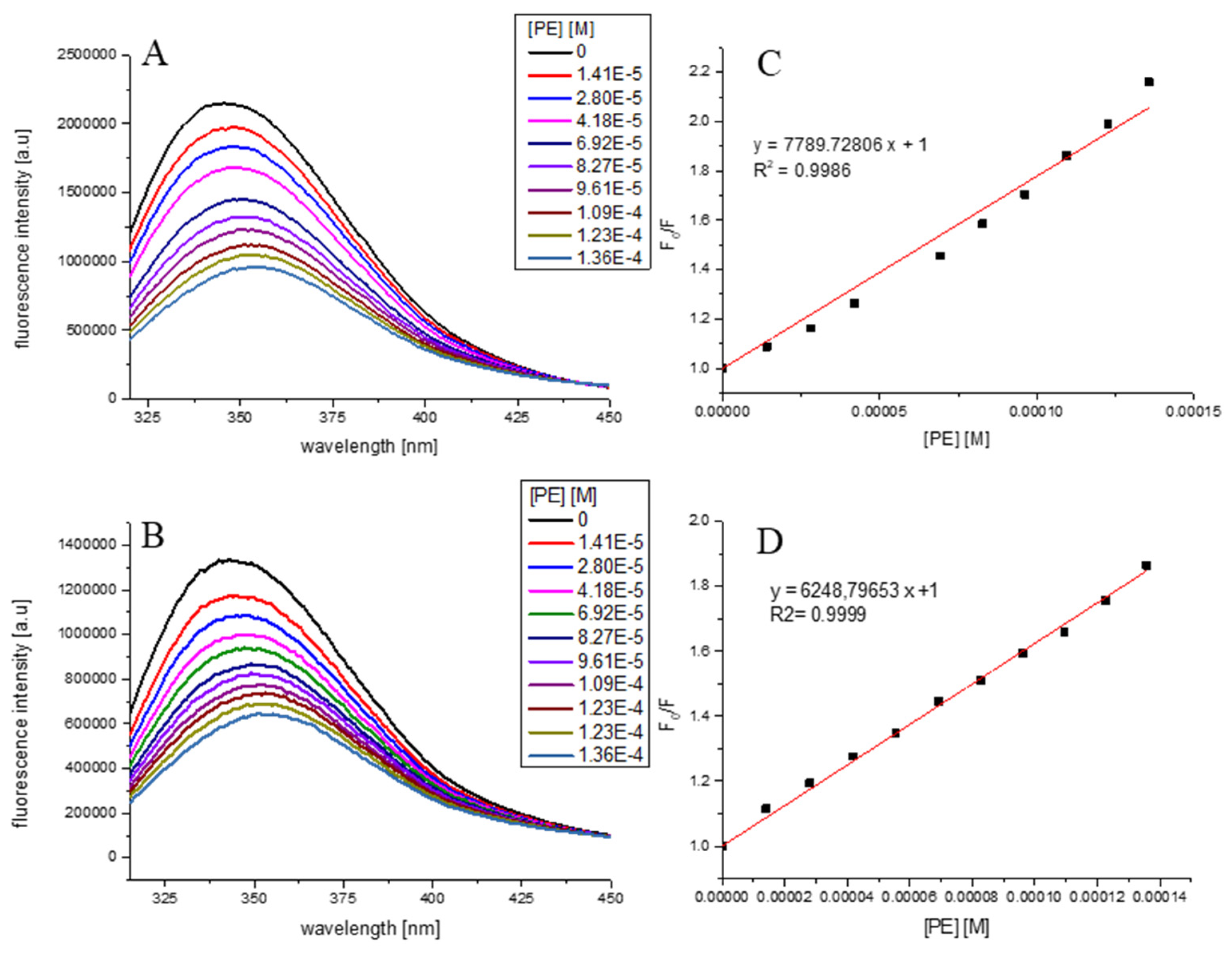
3.2. Effect of PE on α-Amylase and α-Glucosidase Spectra
3.3. Antiglycation Capacity of V. opulus Fruit Samples
3.4. Antioxidant Capacity of V. opulus Fruit Samples
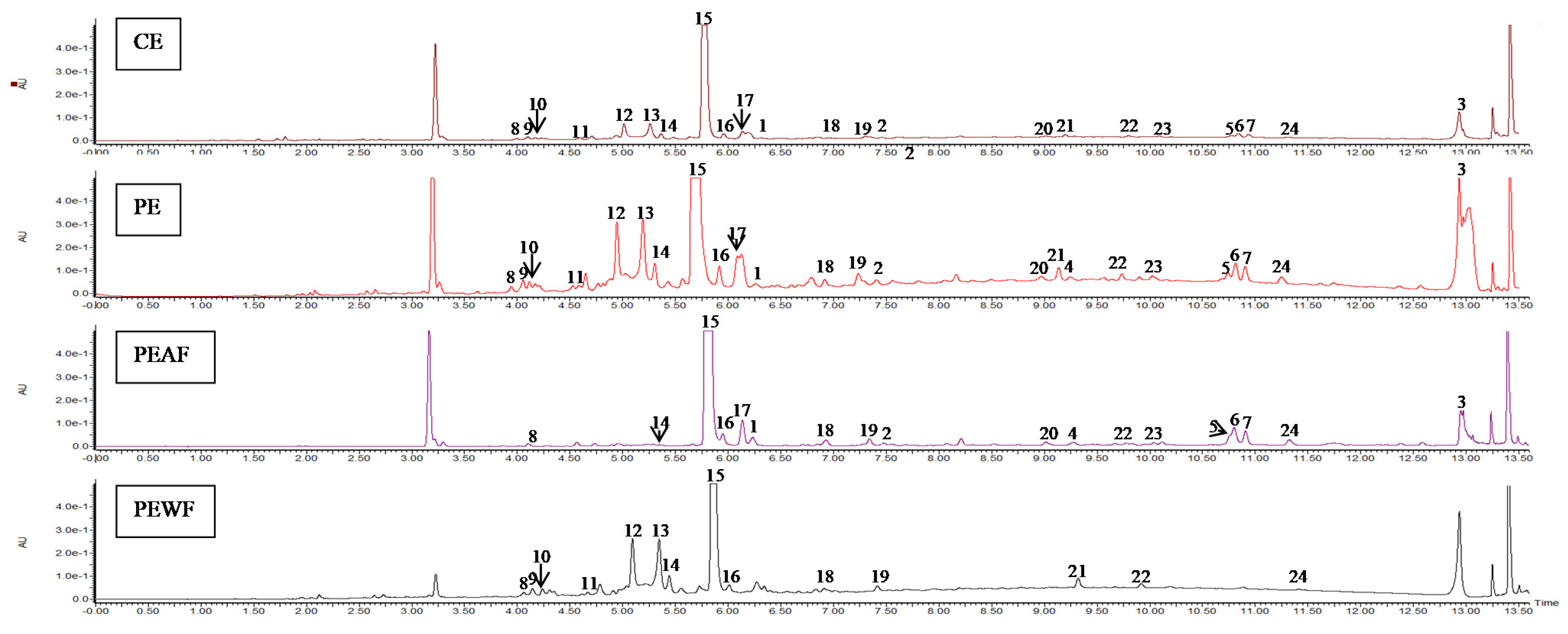
3.5. The Phenolic Compounds Content of V. opulus Fruit Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manaharan, T.; Teng, L.L.; Appleton, D.; Ming, C.H.; Masilamani, T.; Palanisamy, U.D. Antioxidant and antiglycemic potential of Peltophorum pterocarpum plant parts. Food Chem. 2011, 129, 1355–1361. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, X.F.; Chen, J.; Xia, L.; Cao, A.; Zhang, Y.; Pan, A. Combined lifestyle factors and risk of incident type 2 diabetes and prognosis among individuals with type 2 diabetes: A systematic review and meta-analysis of prospective cohort studies. Diabetologia 2020, 63, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassuk, S.S.; Manson, J.E. Lifestyle and risk of cardiovascular disease and type 2 diabetes in women: A review of the epidemiologic evidence. Am. J. Lifestyle Med. 2008, 2, 191–213. [Google Scholar] [CrossRef]
- Haque, M.U.; Sana, S. Antidiabetic activity of plants, fruits and vegetables: A review. Int. J. Biol. Med. Res. 2016, 7, 5452–5458. [Google Scholar]
- Park, J.H.; Kim, R.Y.; Park, E. Antidiabetic activity of fruits and vegetables commonly consumed in Korea: Inhibitory potential against α-glucosidase and insulin-like action in vitro. Food Sci. Biotechnol. 2012, 21, 1187–1193. [Google Scholar] [CrossRef]
- Lee, J.; Noh, S.; Lim, S.; Kim, B. Plant extracts for type 2 diabetes: From traditional medicine to modern drug discovery. Antioxidants 2021, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Semple, S.J.; Simpson, B.S.; Deo, P. Antioxidant and antiglycation activities of Syzygium paniculatum Gaertn and inhibition of digestive enzymes relevant to type 2 diabetes mellitus. Plant Foods Hum. Nutr. 2020, 75, 621–627. [Google Scholar] [CrossRef]
- Spínola, V.; Pinto, J.; Castilho, P.C. Hypoglycemic, anti-glycation and antioxidant in vitro properties of two Vaccinium species from Macaronesia: A relation to their phenolic composition. J. Funct. Foods 2018, 40, 595–605. [Google Scholar] [CrossRef]
- Vinayagam, R.; Jayachandran, M.; Xu, B. Antidiabetic effects of simple phenolic acids: A comprehensive review. Phytother. Res. 2016, 30, 184–199. [Google Scholar] [CrossRef]
- Meneses, M.J.; Silva, B.M.; Sousa, M.; Sa, R.; Oliveira, P.F.; Alves, M.G. Antidiabetic drugs: Mechanisms of action and potential outcomes on cellular metabolism. Curr. Pharm. Des. 2015, 21, 3606–3620. [Google Scholar] [CrossRef]
- Prpa, E.J.; Bajka, B.H.; Ellis, P.R.; Butterworth, P.J.; Corpe, C.P.; Hall, W.L. A systematic review of in vitro studies evaluating the inhibitory effects of polyphenol-rich fruit extracts on carbohydrate digestive enzymes activity: A focus on culinary fruits consumed in Europe. Crit. Rev. Food Sci. Nutr. 2020, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Du, Y.J.; Song, H.C. α-Glucosidase and α-amylase inhibitory activities of guava leaves. Food Chem. 2010, 123, 6–13. [Google Scholar] [CrossRef]
- Alam, F.; Shafique, Z.; Amjad, S.T.; Bin Asad, M.H.H. Enzymes inhibitors from natural sources with antidiabetic activity: A review. Phytother. Res. 2019, 33, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantidos, N.; Boath, A.; Lund, V.; Conner, S.; McDougall, G.J. Phenolic-rich extracts from the edible seaweed, Ascophyllum nodosum, inhibit α-amylase and α-glucosidase: Potential anti-hyperglycemic effects. J. Funct. Food 2014, 10, 201–209. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Rodríguez-Werner, M.; Schlösser, A.; Winterhalter, P.; Rimbach, G. Fractionation, enzyme inhibitory and cellular antioxidant activity of bioactives from purple sweet potato (Ipomoea batatas). Food Chem. 2017, 221, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Ahangarpour, A.; Sayahi, M.; Sayahi, M. The antidiabetic and antioxidant properties of some phenolic phytochemicals: A review study. Diabetes Metab. Syndr. 2019, 13, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Aba, P.E.; Asuzu, I.U. Mechanisms of actions of some bioactive anti-diabetic principles from phytochemicals of medicinal plants: A review. Indian J. Nat. Prod. Resour. 2018, 9, 85–96. [Google Scholar]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycemic level: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef]
- Cosme, P.; Rodríguez, A.B.; Espino, J.; Garrido, M. Plant phenolics: Bioavailability as a key determinant of their potential health-promoting applications. Antioxidants 2020, 9, 1263. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.R.; Alves, M.G.; Casal, S.; Oliveira, P.F.; Silva, B.M. Promising potential of dietary (poly)phenolic compounds in the prevention and treatment of diabetes mellitus. Curr. Med. Chem. 2017, 24, 334–354. [Google Scholar]
- Soman, S.; Rauf, A.A.; Indira, M.; Rajamanickam, C. Antioxidant and antiglycative potential of ethyl acetate fraction of Psidium guajava leaf extract in streptozotocin-induced diabetic rats. Plant Foods Hum. Nutr. 2010, 65, 386–391. [Google Scholar] [CrossRef]
- Wu, T.; Luo, J.; Xu, B. In vitro antidiabetic effects of selected fruits and vegetables against glycosidase and aldose reductase. Food Sci. Nutr. 2015, 3, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Chen, X. Structures required of flavonoids for inhibiting digestive enzymes. Anti-Cancer Agents Med. Chem. 2012, 12, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Simsek, M.; Quezada-Calvillo, R.; Ferruzzi, M.G.; Nichols, B.L.; Hamaker, B.R. Dietary phenolic compounds selectively inhibit the individual subunits of maltase-glucoamylase and sucrase-isomaltase with the potential of modulating glucose release. J. Agric. Food Chem. 2015, 63, 3873–3879. [Google Scholar] [CrossRef] [PubMed]
- Pollini, L.; Riccio, A.; Juan, C.; Tringaniello, C.; Ianni, F.; Blasi, F.; Cossignani, L. Phenolic acids from Lycium barbarum leaves: In vitro and in silico studies of the inhibitory activity against porcine pancreatic α-amylase. Processes 2020, 8, 1388. [Google Scholar] [CrossRef]
- Shen, W.; Xu, Y.; Lu, Y.H. Inhibitory effects of citrus flavonoids on starch digestion and antihyperglycemic effects in HepG2 cells. J. Agric. Food Chem. 2012, 60, 9609–9619. [Google Scholar] [CrossRef]
- Zhou, Q.; Cheng, K.W.; Xiao, J.; Wang, M. The multifunctional roles of flavonoids against the formation of advanced glycation end products (AGEs) and AGEs-induced harmful effects. Trends Food Sci. Technol. 2020, 103, 333–347. [Google Scholar] [CrossRef]
- Matsuda, H.; Wang, T.; Managi, H.; Yoshikawa, M. Structural requirements of flavonoids for inhibition of protein glycation and radical scavenging activities. Bioorg. Med. Chem. 2003, 11, 5317–5323. [Google Scholar] [CrossRef]
- Cho, S.; Lee, H.; Han, J.; Lee, H.; Kattia, R.O.; Nelson, Z.V.; Jeong, T.S. Viburnum stellato-tomentosum extract suppresses obesity and hyperglycemia through regulation of lipid metabolism in high-fat diet-fed mice. Molecules 2021, 26, 1052. [Google Scholar] [CrossRef]
- Iwai, K.; Kim, M.Y.; Onodera, A.; Matsue, H. α-Glucosidase inhibitory and antihyperglycemic effects of polyphenols in the fruit of Viburnum dilatatum thunb. J. Agric. Food Chem. 2006, 54, 4588–4592. [Google Scholar] [CrossRef]
- Altun, M.L.; Özbek, H.; Çitoğlu, G.S.; Yilmaz, B.S.; Bayram, I.; Cengiz, N. Hepatoprotective and hypoglycemic activities of Viburnum opulus L. Turk. J. Pharm. Sci. 2010, 7, 35–48. [Google Scholar]
- Zakłos-Szyda, M.; Pawlik, N.; Polka, D.; Nowak, A.; Koziołkiewicz, M.; Podsędek, A. Viburnum opulus fruit phenolic compounds as cytoprotective agents able to decrease free fatty acids and glucose uptake by Caco-2 cells. Antioxidants 2019, 8, 262. [Google Scholar] [CrossRef] [Green Version]
- Zakłos-Szyda, M.; Kowalska-Baron, A.; Pietrzyk, N.; Drzazga, A.; Podsędek, A. Evaluation of Viburnum opulus L. fruit phenolics cytoprotective potential on insulinoma MIN6 cells relevant for diabetes mellitus and obesity. Antioxidants 2020, 9, 433. [Google Scholar] [CrossRef]
- Zakłos-Szyda, M.; Majewska, I.; Redzynia, M.; Koziołkiewicz, M. Antidiabetic effect of polyphenolic extracts from selected edible plants as α-amylase, α-glucosidase and PTP1B inhibitors, and β-pancreatic cells cytoprotective agents—A comparative study. Curr. Top. Med. Chem. 2015, 15, 2431–2444. [Google Scholar] [CrossRef] [PubMed]
- Kajszczak, D.; Zakłos-Szyda, M.; Podsędek, A. Viburnum opulus L.—A review of phytochemistry and biological effects. Nutrients 2020, 12, 3398. [Google Scholar] [CrossRef] [PubMed]
- Dienaitė, L.; Pukalskienė, M.; Pereira, C.V.; Matias, A.A.; Venskutonis, P.R. Valorization of European cranberry bush (Viburnum opulus L.) berry pomace extracts isolated with pressurized ethanol and water by assessing their phytochemical composition, antioxidant, and antiproliferative activities. Foods 2020, 9, 1413. [Google Scholar] [CrossRef]
- Zakłos-Szyda, M.; Nowak, A.; Pietrzyk, N.; Podsędek, A. Viburnum opulus L. juice phenolic compounds influence osteogenic differentiation in human osteosarcoma Saos-2 cells. Inter. J. Mol. Sci. 2020, 21, 4909. [Google Scholar] [CrossRef] [PubMed]
- Wójcik-Bojek, U.; Rywaniak, J.; Bernat, P.; Podsędek, A.; Kajszczak, D.; Sadowska, B. An in vitro study of the effect of Viburnum opulus extracts on key processes in the development of Staphylococcal infections. Molecules 2021, 26, 1758. [Google Scholar] [CrossRef]
- Rösch, D.; Bergmann, M.; Knorr, D.; Kroh, L.W. Structure antioxidant efficiency relationships of phenolic compounds and their contribution to the antioxidant activity of sea buckthorn juice. J. Agric. Food Chem. 2003, 51, 4233–4239. [Google Scholar] [CrossRef]
- Podsędek, A.; Majewska, I.; Redzynia, M.; Sosnowska, D.; Koziołkiewicz, M. In vitro inhibitory effect on digestive enzymes and antioxidant potential of commonly consumed fruits. J. Agric. Food Chem. 2014, 62, 4610–4617. [Google Scholar] [CrossRef] [PubMed]
- Adisakwattana, S.; Charoenlertkul, P.; Yibchok-Anun, S. A-Glucosidase Inhibitory activity of cyanidin-3-galactoside and synergistic effect with acarbose. J. Enzyme Inhib. Med. Chem. 2009, 24, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Huang, Y.N.; Gao, B.; Li, P.; Inagaki, C.; Kawabata, J. Inhibitory effect on α-glucosidase by Adhatoda vasica Nees. Food Chem. 2008, 108, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Khalifa, I.; Wang, R.; Li, C. Persimmon highly galloylated-tannins in vitro mitigated α-amylase and α-glucosidase via statically binding with their catalytic-closed sides and altering their secondary structure elements. J. Food. Biochem. 2020, 44, 1–12. [Google Scholar] [CrossRef]
- Błaszczak, W.; Jeż, M.; Szwengiel, A. Polyphenols and inhibitory effects of crude and purified extracts from tomato varieties on the formation of advanced glycation end products and the activity of angiotensin-converting and acetylcholinesterase enzymes. Food Chem. 2020, 314, 126181. [Google Scholar] [CrossRef] [PubMed]
- Polka, D.; Podsędek, A.; Koziołkiewicz, M. Comparison of chemical composition and antioxidant capacity of fruit, flower and bark of Viburnum opulus. Plant Food. Hum. Nutr. 2019, 74, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Chen, S. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Martín, C. Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, Y.; Wang, Y.; Zhang, Y. Bioassay-guided screening and isolation of α-glucosidase and tyrosinase inhibitors from leaves of Morus alba. Food Chem. 2012, 131, 617–625. [Google Scholar] [CrossRef]
- Rubilar, M.; Jara, C.; Poo, Y.; Acevedo, F.; Gutierrez, C.; Sineiro, J.; Shene, C. Extracts of Maqui (Aristotelia chilensis) and Murta (Ugni molinae Turcz.): Sources of antioxidant compounds and α-glucosidase/α-amylase inhibitors. J. Agric. Food Chem. 2011, 59, 1630–1637. [Google Scholar] [CrossRef] [PubMed]
- Aksornchu, P.; Chamnansilpa, N.; Adisakwattana, S.; Thilavech, T.; Choosak, C.; Marnpae, M.; Ngamukote, S. Inhibitory effect of Antidesma bunius fruit extract on carbohydrate digestive enzymes activity and protein glycation in vitro. Antioxidants 2021, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Justino, A.B.; de Moura, F.R.B.; Franco, R.R.; Espindola, F.S. α-Glucosidase and non-enzymatic glycation inhibitory potential of Eugenia dysenterica fruit pulp extracts. Food Biosci. 2020, 35, 100573. [Google Scholar] [CrossRef]
- Saucier, C.; Mirabel, M.; Daviaud, F.; Longieras, A.; Glories, Y. Rapid fractionation of grape seed proanthocyanidins. J. Agric. Food Chem. 2001, 49, 5732–5735. [Google Scholar] [CrossRef]
- Perova, I.B.; Zhogova, A.A.; Cherkashin, A.V.; Éller, K.I.; Ramenskaya, G.V.; Samylina, I.A. Biologically active substances from European guelder berry fruits. Pharm. Chem. J. 2014, 48, 332–339. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Ekici, L.; Poyrazoglu, E.S. Phenolic composition of European cranberrybush (Viburnum opulus L.) berries and astringency removal of its commercial juice. Int. J. Food Sci. Technol 2006, 9205, 1011–1015. [Google Scholar] [CrossRef]
- Zakłos-Szyda, M.; Pietrzyk, N.; Szustak, M.; Podsędek, A. Viburnum opulus L. juice phenolics inhibit mouse 3T3-L1 cells adipogenesis and pancreatic lipase activity. Nutrients 2020, 12, 2003. [Google Scholar] [CrossRef] [PubMed]
- Barak, T.H.; Celep, E.; Yesilada, E. Influence of in vitro human digestion on the bioavailability of phenolic content and antioxidant activity of Viburnum opulus L. (European cranberry) fruit extracts. Ind. Crop. Prod. 2019, 131, 62–69. [Google Scholar] [CrossRef]
- Kam, A.; Li, K.M.; Razmovski-Naumovski, V.; Nammi, S.; Shi, J.; Chan, K.; Li, G.Q. A comparative study on the inhibitory effects of different parts and chemical constituents of pomegranate on α-amylase and α-glucosidase. Phytother. Res. 2013, 27, 1614–1620. [Google Scholar] [CrossRef]
- Kato, E. Bioactive compounds in plant materials for the prevention of diabetes and obesity. Biosci. Biotechnol. Biochem. 2019, 83, 975–985. [Google Scholar] [CrossRef]
- Zhong, H.; Xue, Y.; Lu, X.; Shao, Q.; Cao, Y.; Wu, Z.; Chen, G. The effects of different degrees of procyanidin polymerization on the nutrient absorption and digestive enzyme activity in mice. Molecules 2018, 23, 2916. [Google Scholar] [CrossRef] [Green Version]
- Narita, Y.; Inouye, K. Inhibitory effects of chlorogenic acids from green coffee beans and cinnamate derivatives on the activity of porcine pancreas α-amylase isozyme I. Food Chem. 2011, 127, 1532–1539. [Google Scholar] [CrossRef] [Green Version]
- Oki, T.; Matsui, T.; Osajima, Y. Inhibitory effect of α-glucosidase inhibitors varies according to its origin. J. Agric. Food Chem. 1999, 47, 550–553. [Google Scholar] [CrossRef]
- Karim, Z.; Holmes, M.; Orfila, C. Inhibitory effect of chlorogenic acid on digestion of potato starch. Food Chem. 2017, 217, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Chen, W.; Meng, Y.; Yang, X.; Yuan, L.; Guo, Y. Interactions between polyphenols in thinned young apples and porcine pancreatic α-amylase: Inhibition, detailed kinetics and fluorescence quenching. Food Chem. 2016, 208, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Nájera, V.C.; Gutiérrez-Uribe, J.A.; Antunes-Ricardo, M.; Hidalgo-Figueroa, S.; Del-Toro-Sánchez, C.L.; Salazar-Olivo, L.A.; Lugo-Cervantes, E. Smilax aristolochiifolia root extract and its compounds chlorogenic acid and astilbin inhibit the activity of α-amylase and α-glucosidase enzymes. J. Evid. Based Complementary Altern. Med. 2018, 2018, 6247306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Wang, Q.; Zhang, W.; Hu, B.; Zhou, L.; Zeng, X.; Sun, Y. Inhibitory activities of caffeoylquinic acid derivatives from Ilex kudingcha CJ Tseng on α-glucosidase from Saccharomyces cerevisiae. J. Agric. Food Chem. 2015, 63, 3694–3703. [Google Scholar] [CrossRef]
- Lakowicz, J.R.; Masters, B.R. Principles of fluorescence spectroscopy. J. Biomed. Opt. 2008, 13, 9901. [Google Scholar] [CrossRef]
- Yuan, T.; Yang, T.; Chen, H.; Fu, D.; Hu, Y.; Wang, J.; Xie, X. New insights into oxidative stress and inflammation during diabetes mellitus-accelerated atherosclerosis. Redox Biol. 2019, 20, 247–260. [Google Scholar] [CrossRef]
- Dos Santos, J.M.; Tewari, S.; Mendes, R.H. The role of oxidative stress in the development of diabetes mellitus and its complications. J. Evid. Based Complementary Altern. Med. 2019, 2019, 4189813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.G.; Hu, Q.P.; Liu, Y. Antioxidant and DNA-protective activities of chlorogenic acid isomers. J. Agric. Food Chem. 2012, 60, 11625–11630. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G. Prevention of protein glycation by natural compounds. Molecules 2015, 20, 3309–3334. [Google Scholar] [CrossRef]
- Jang, D.S.; Yoo, N.H.; Kim, N.H.; Lee, Y.M.; Kim, C.S.; Kim, J.; Kim, J.S. 3, 5-Di-O-caffeoyl-epi-quinic acid from the leaves and stems of Erigeron annuus inhibits protein glycation, aldose reductase, and cataractogenesis. Biol. Pharm. Bull. 2010, 33, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Jeong, I.H.; Kim, C.S.; Lee, Y.M.; Kim, J.M.; Kim, J.S. Chlorogenic acid inhibits the formation of advanced glycation end products and associated protein cross-linking. Arch. Pharm. Res. 2011, 34, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Gugliucci, A.; Bastos, D.H.M.; Schulze, J.; Souza, M.F.F. Caffeic and chlorogenic acids in Ilex paraguariensis extracts are the main inhibitors of AGE generation by methylglyoxal in model proteins. Fitoterapia 2009, 80, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; McIntyre, K.; Saleem, A.; Haddad, P.S.; Arnason, J.T. The relationship between antiglycation activity and procyanidin and phenolic content in commercial grape seed products. Can. J. Physiol. Pharmacol. 2012, 90, 167–174. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample | α-Amylase | α-Glucosidase | ||
|---|---|---|---|---|
| Substrate | ||||
| Potato Starch | Starch from Rice | Maltose | Sucrose | |
| CE | 400.70 ± 8.29d | 572.12 ± 19.41d | 1922.83 ± 77.14e | 3244.88 ± 111.28d |
| PE | 61.51 ± 0.72b | 132.42 ± 3.33b | 180.09 ± 4.67cd | 447.60 ± 29.22c |
| PEAF | 378.71 ± 3.64c | 509.22 ± 1.81c | 61.87 ± 1.97ab | 110.06 ± 1.12b |
| PEWF | 24.17 ± 0.23a | 124.90 ± 2.70b | 253.71 ± 6.79d | 324.93 ± 11.32c |
| Chlorogenic acid | 833.61 ± 16.34e | 1963.57 ± 30.49e | 103.37 ± 5.29bc | 99.81 ± 0.75ab |
| Acarbose | 13.33 ± 0.17a | 30.57 ± 1.19a | 0.051 ± 0.001a | 1.37 ± 0.04a |
| Enzyme | Substrate | Sample | Concentration (μg/mL) | Km (μg/mL) | Vmax (OD/min) | Ki (μg/mL) | Inhibition Type |
|---|---|---|---|---|---|---|---|
| α-amylase | Potato starch | Control PEWF | - 25.64 | 1.25 0.32 | 1.32 0.40 | / 11.14 | uncompetitive |
| Control PE | - 54.95 | 1.00 0.21 | 1.10 0.24 | / 15.22 | uncompetitive | ||
| Control Chlorogenic acid | - 769.23 | 0.59 0.83 | 0.80 0.80 | / 1225.78 | competitive | ||
| Control Acarbose | - 6.62 | 1.43 0.20 | 1.41 0.22 | / 1.24 | uncompetitive | ||
| Rice starch | Control PEWF | - 153.86 | 1.25 0.18 | 0.63 0.08 | / 20.82 | uncompetitive | |
| Control PE | - 153.86 | 1.00 0.15 | 0.51 0.07 | / 22.84 | uncompetitive | ||
| Control Chlorogenic acid | - 1923.1 | 1.67 3.33 | 0.60 0.60 | / 1886.20 | competitive | ||
| Control Acarbose | - 19.86 | 2.50 0.19 | 1.27 0.10 | / 1.78 | uncompetitive | ||
| α-glucosidase | Maltose | Control PEAF | - 61.73 | 0.40 0.63 | 0.021 0.014 | / 46.44 | mixed |
| Control PE | - 185.19 | 0.18 0.69 | 0.023 0.013 | / 89.25 | mixed | ||
| Control Chlorogenic acid | - 191.31 | 0.40 0.40 | 0.021 0.009 | / 145.41 | noncompetitive | ||
| Control Acarbose | - 0.05 | 0.40 0.69 | 0.021 0.012 | / 0.021 | mixed | ||
| Sucrose | Control PEAF | - 111.11 | 6.67 9.09 | 0.006 0.005 | / 159.10 | mixed | |
| Control PE | - 476.19 | 5.56 6.67 | 0.008 0.004 | / 379.75 | mixed | ||
| Control Chlorogenic acid | - 191.31 | 7.69 7.69 | 0.005 0.003 | / 181.21 | noncompetitive | ||
| Control Acarbose | - 1.35 | 6.67 8.33 | 0.006 0.004 | / 1.52 | mixed |
| Sample | τ1 [ns] | f1 [%] | τ2 [ns] | f2 [%] | τ3 [ns] | f3 [%] | <τ> * [ns] | χ2 |
|---|---|---|---|---|---|---|---|---|
| α-Amylase | 0.89 ± 0.04 | 13.73 | 3.13 ± 0.05 | 58.35 | 8.90 ± 0.11 | 27.92 | 12.40398 | 1.169 |
| α-Amylase + PE | 0.84 ± 0.04 | 14.18 | 3.10 ± 0.05 | 58.66 | 8.83 ± 0.11 | 27.17 | 12.12822 | 1.210 |
| α-Glucosidase | 0.62 ± 0.03 | 11.51 | 3.02 ± 0.04 | 62.61 | 7.02 ± 0.11 | 25.88 | 11.00854 | 1.196 |
| α-Glucosidase + PE | 0.66 ± 0.03 | 12.29 | 3.12 ± 0.05 | 64.20 | 7.19 ± 0.13 | 23.51 | 11.50588 | 1.028 |
| Sample | BSA-Fructose Model | BSA-Glucose Model |
|---|---|---|
| CE | 182.03 ± 2.52e | 4966.22 ± 110.32d |
| PE | 40.78 ± 0.02b | 895.33 ± 9.39c |
| PEAF | 32.32 ± 0.28a | 577.50 ± 5.32b |
| PEWF | 33.96 ± 0.27a | 803.27 ± 4.79c |
| Chlorogenic Acid | 58.87 ± 0.93c | 869.94 ± 6.25c |
| Aminoguanidine | 106.08 ± 3.17d | 137.80 ± 3.44a |
| Sample | ABTS | FRAP | ORAC |
|---|---|---|---|
| CE | 0.61 ± 0.02a | 0.40 ± 0.01a | 1.15 ± 0.03a |
| PE | 2.24 ± 0.03b | 1.80 ± 0.08b | 3.33 ± 0.10b |
| PEAF | 6.41 ± 0.25d | 5.18 ± 0.06c | 24.12 ± 0.91d |
| PEWF | 5.13 ± 0.08c | 5.02 ± 0.15c | 10.70 ± 0.40c |
| Phenolic Compounds | V. opulus fruit Sample | |||
|---|---|---|---|---|
| CE | PE | PEAF | PEWF | |
| Flavanols 1 | 11.53 ± 0.07a | 52.63 ± 1.76b | 98.50 ± 3.90c | - |
| Flavalignans 1 | 1.13 ± 0.04a | 8.42 ± 0.20b | 40.70 ± 0.27c | - |
| Hydroxycinnamic acids 1 | 73.90 ± 0.51a | 352.26 ± 3.57c | 829.26 ± 0.51d | 201.81 ± 0.63b |
| Flavonols 1 | 0.86 ± 0.01a | 5.08 ± 0.12b | 8.62 ± 0.17c | 5.06 ± 0.05b |
| Total phenolics 1 | 87.42 ± 0.63a | 418.39 ± 5.65c | 977.08 ± 4.85d | 206.87 ± 0.68b |
| Total proanthocyanidins 2 | 12.26 ± 0.46a | 74.72 ± 7.90d | 21.96 ± 1.35b | 57.33 ± 1.85c |
| Peak | Phenolic Compound | CE | PE | PEAF | PEWF |
|---|---|---|---|---|---|
| mg/g of Extract | mg/g of Fraction | ||||
| Flavanols | |||||
| 1 | Procyanidin tetramer a | 0.23 ± 0.00a | 1.81 ± 0.03c | 1.28 ± 0.04b | - |
| 2 | Procyanidin C1 | 0.80 ± 0.00a | 2.03 ± 0.11c | 0.99 ± 0.03b | - |
| 3 | (Epi)-catechin hexoside b | 10.50 ± 0.07a | 48.79 ± 1.62b | 96.23 ± 3.83c | - |
| Flavalignans | |||||
| 4 | Cinchonain Ix c | - | 0.72 ± 0.02a | 3.88 ± 0.02b | - |
| 5 | Cinchonain Ix c | 0.19 ± 0.03a | 1.90 ± 0.06b | 23.13 ± 0.21c | - |
| 6 | Cinchonain Ix c | 0.54 ± 0.01a | 3.23 ± 0.07b | 11.55 ± 0.04c | - |
| 7 | Cinchonain Ix c | 0.40 ± 0.00a | 2.57 ± 0.05c | 2.14 ± 0.00b | - |
| Hydroxycinnamic acids | |||||
| 8 | Neochlorogenic acid | 0.20 ± 0.00a | 1.41 ± 0.05b | 1.97 ± 0.03c | 1.39 ± 0.00b |
| 9 | Dicaffeolquinic acid I d | 0.20 ± 0.00a | 1.69 ± 0.08b | - | 1.66 ± 0.00b |
| 10 | Dicaffeolquinic acid II d | 0.09 ± 0.00a | 1.38 ± 0.05b | - | 1.58 ± 0.00c |
| 11 | Dicaffeolquinic acid III d | 0.23 ± 0.00a | 0.55 ± 0.02b | - | 1.22 ± 0.00c |
| 12 | Dicaffeolquinic acid IV d | 2.39 ± 0.01a | 15.14 ± 0.47b | - | 17.35 ± 0.01c |
| 13 | Dicaffeolquinic acid V d | 2.92 ± 0.00a | 20.04 ± 0.20c | - | 19.68 ± 0.04b |
| 14 | Dicaffeolquinic acid VI d | 0.68 ± 0.00b | 4.37 ± 0.12c | 0.37 ± 0.00a | 4.40 ± 0.01c |
| 15 | Chlorogenic acid | 65.62 ± 0.49a | 300.23 ± 2.33c | 795.72 ± 0.03d | 152.05 ± 0.52b |
| 16 | Cryptochlorogenic acid | 0.40 ± 0.00a | 1.81 ± 0.05c | 4.78 ± 0.01d | 0.97 ± 0.01b |
| 17 | Caffeic acid | 0.38 ± 0.01a | 1.67 ± 0.02b | 8.37 ± 0.11c | - |
| 18 | Caffeoylquinic acid d | 0.10 ± 0.00a | 0.66 ± 0.06b | 3.84 ± 0.24c | 0.40 ± 0.02ab |
| 19 | Coumaroylquinic acid d | 0.46 ± 0.00a | 2.48 ± 0.09c | 9.99 ± 0.01d | 1.11 ± 0.02b |
| 20 | Feruoylquinic acid d | 0.23 ± 0.00a | 0.83 ± 0.03b | 4.22 ± 0.08c | - |
| Flavonols | |||||
| 21 | Quercetin 3-sambubioside e | 0.27 ± 0.00a | 1.35 ± 0.04b | - | 2.01 ± 0.00c |
| 22 | Quercetin 3-rutinoside | 0.41 ± 0.00a | 2.22 ± 0.02c | 1.70 ± 0.15b | 2.80 ± 0.03d |
| 23 | Quercetin 3-glucoside | 0.07 ± 0.01a | 0.46 ± 0.03b | 2.05 ± 0.01c | - |
| 24 | Quercetin 3-rhamnoside | 0.11 ± 0.00a | 1.05 ± 0.03c | 4.87 ± 0.01d | 0.25 ± 0.02b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kajszczak, D.; Kowalska-Baron, A.; Podsędek, A. Glycoside Hydrolases and Non-Enzymatic Glycation Inhibitory Potential of Viburnum opulus L. Fruit—In Vitro Studies. Antioxidants 2021, 10, 989. https://doi.org/10.3390/antiox10060989
Kajszczak D, Kowalska-Baron A, Podsędek A. Glycoside Hydrolases and Non-Enzymatic Glycation Inhibitory Potential of Viburnum opulus L. Fruit—In Vitro Studies. Antioxidants. 2021; 10(6):989. https://doi.org/10.3390/antiox10060989
Chicago/Turabian StyleKajszczak, Dominika, Agnieszka Kowalska-Baron, and Anna Podsędek. 2021. "Glycoside Hydrolases and Non-Enzymatic Glycation Inhibitory Potential of Viburnum opulus L. Fruit—In Vitro Studies" Antioxidants 10, no. 6: 989. https://doi.org/10.3390/antiox10060989