
Benefits of Exercise and Astaxanthin Supplementation: Are There Additive or Synergistic Effects?

 , and
, and
Abstract
:1. Introduction
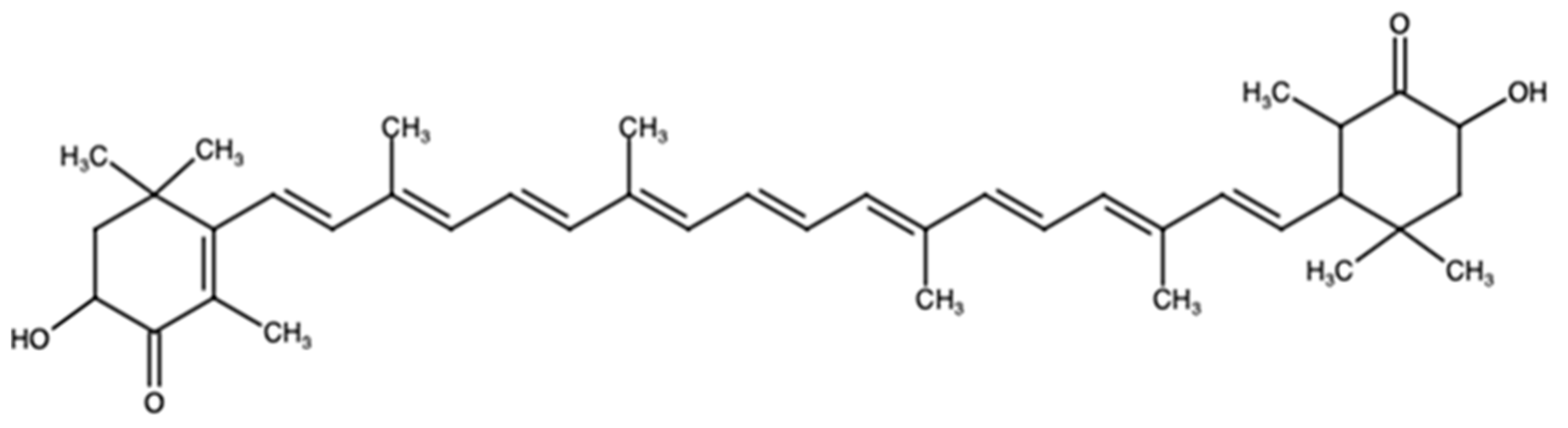
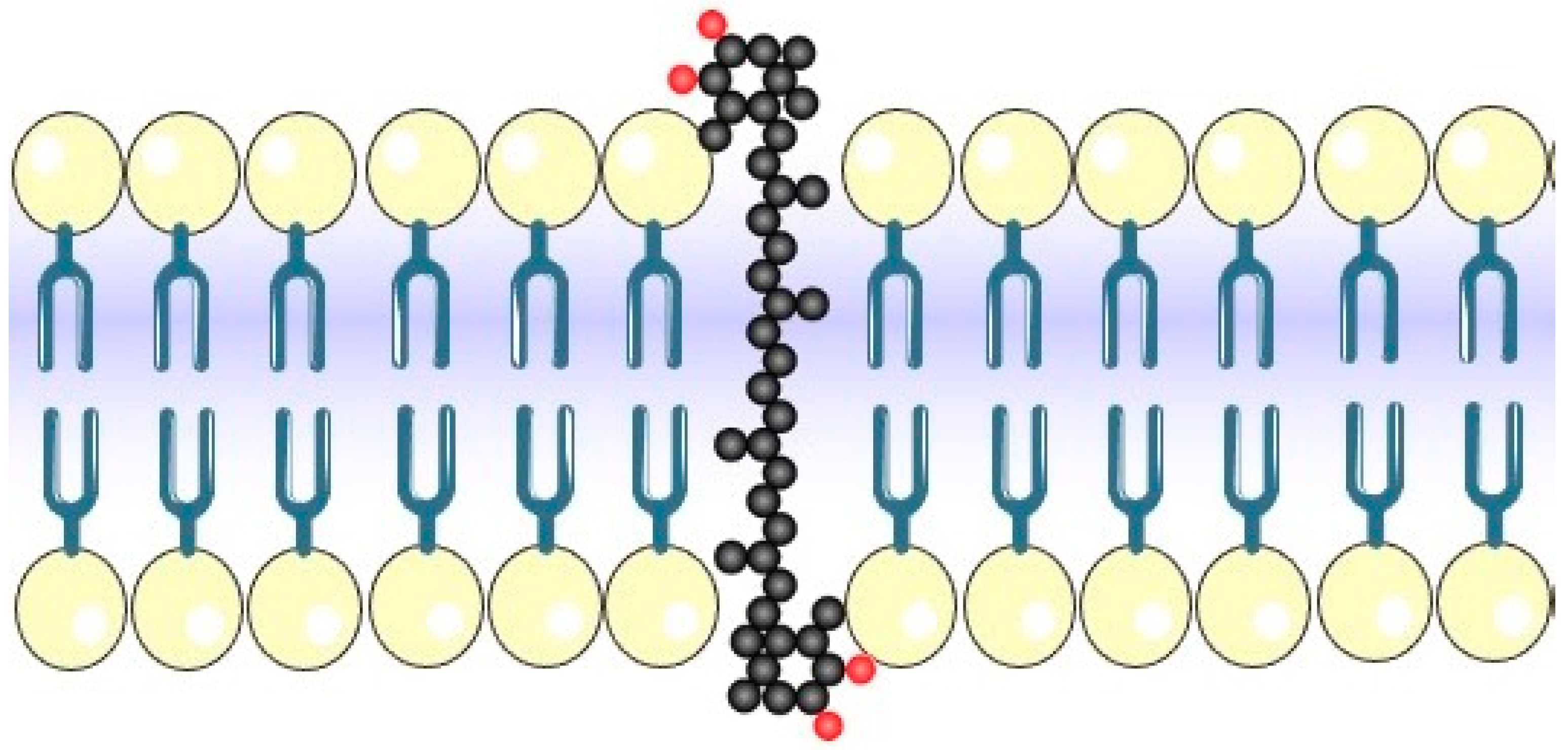
2. Astaxanthin
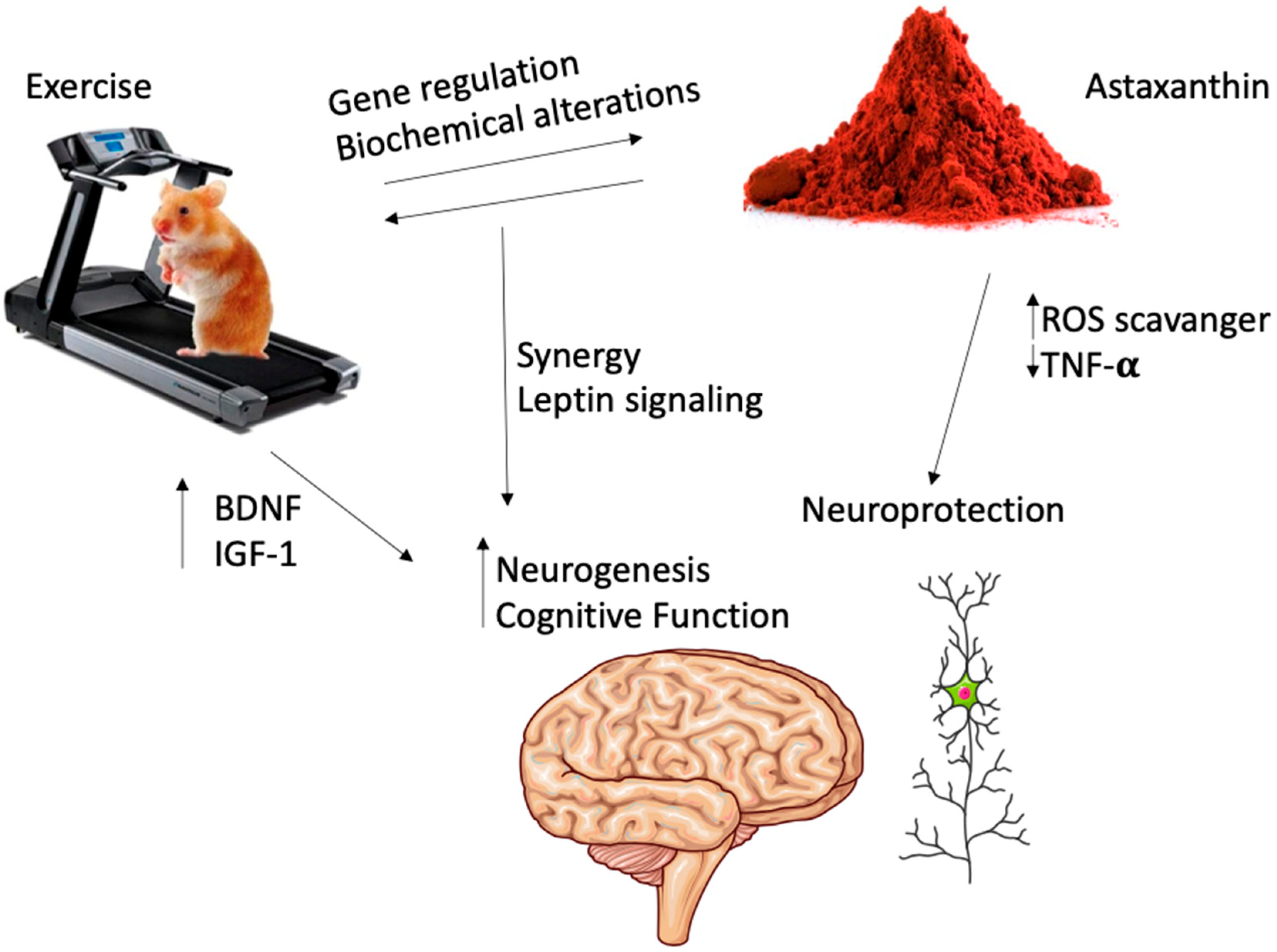
3. Astaxanthin and Exercise: Animal Studies
4. Astaxanthin and Exercise: Human Studies
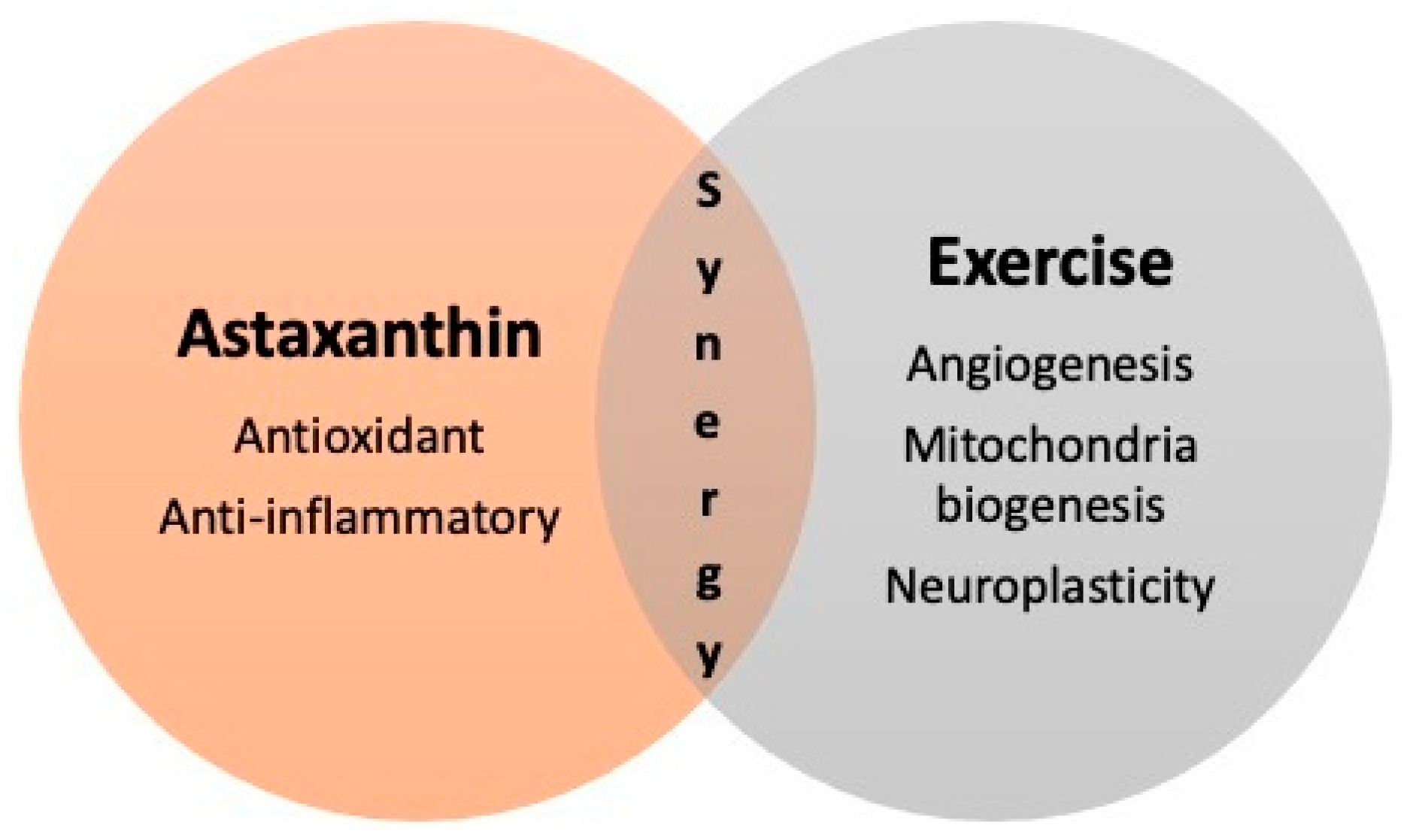
5. Overview of the Current Scientific Literature
6. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zheng, L.; Wu, J.; Wang, G.; Persuitte, G.; Ma, Y.; Zou, L.; Zhang, L.; Zhao, M.; Wang, J.; Lan, Q.; et al. Comparison of control fasting plasma glucose of exercise-only versus exercise-diet among a pre-diabetic population: A meta-analysis. Eur. J. Clin. Nutr. 2016, 70, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Bondyra-Wiśniewska, B.; Myszkowska-Ryciak, J.; Harton, A. Impact of lifestyle intervention programs for children and adolescents with overweight or obesity on body weight and selected cardiometabolic factors—A systematic review. Int. J. Environ. Res. Public Health 2021, 18, 2061. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Delgado, A.; Hernández-Segura, N.; Fernández-Villa, T.; Molina, A.J.; Martín, V. The effect of lifestyle intervention on health-related quality of life in adults with metabolic syndrome: A meta-analysis. Int. J. Environ. Res. Public Health 2021, 18, 887. [Google Scholar] [CrossRef] [PubMed]
- Al-Dashti, Y.A.; Holt, R.R.; Stebbins, C.L.; Keen, C.L.; Hackman, R.M. Dietary flavanols: A review of select effects on vascular function, blood pressure, and exercise performance. J. Am. Coll. Nutr. 2018, 37, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C. Lifestyle modulators of neuroplasticity: How physical activity, mental engagement, and diet promote cognitive health during aging. Neural Plast. 2017, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Baguley, B.; Bolam, K.; Wright, O.; Skinner, T. The effect of nutrition therapy and exercise on cancer-related fatigue and quality of life in men with prostate cancer: A systematic review. Nutrients 2017, 9, 1003. [Google Scholar] [CrossRef] [Green Version]
- Verheggen, R.J.H.M.; Maessen, M.F.H.; Green, D.J.; Hermus, A.R.M.M.; Hopman, M.T.E.; Thijssen, D.H.T. A systematic review and meta-analysis on the effects of exercise training versus hypocaloric diet: Distinct effects on body weight and visceral adipose tissue. Obes. Rev. 2016, 17, 664–690. [Google Scholar] [CrossRef] [PubMed]
- Ismail, H.; McFarlane, J.R.; Nojoumian, A.H.; Dieberg, G.; Smart, N.A. Clinical outcomes and cardiovascular responses to different exercise training intensities in patients with heart failure. a systematic review and meta-analysis. JACC Heart Fail. 2013, 1, 514–522. [Google Scholar] [CrossRef]
- Conte, M.; Baldin, A.; Russo, M.; Storti, L.; Caldara, A.; Cozza, H.; Ciolac, E. Effects of high-intensity interval vs. continuous moderate exercise on intraocular pressure. Int. J. Sports Med. 2014, 35, 874–878. [Google Scholar] [CrossRef]
- Nieman, D.C.; Miller, A.R.; Henson, D.A.; Warren, B.J.; Gusewitch, G.; Johnson, R.L.; Davis, J.M.; Butterworth, D.E.; Nehlsen-Cannarella, S.L. Effects of high- vs moderate-intensity exercise on natural killer cell activity. Med. Sci. Sports Exerc. 1993, 25, 1126–1134. [Google Scholar] [CrossRef]
- Shannon, O.M.; Ashor, A.W.; Scialo, F.; Saretzki, G.; Martin-Ruiz, C.; Lara, J.; Matu, J.; Griffiths, A.; Robinson, N.; Lillà, L.; et al. Mediterranean diet and the hallmarks of ageing. Eur. J. Clin. Nutr. 2021. [Google Scholar] [CrossRef]
- Bhatt, T.; Patel, K. Carotenoids: Potent to prevent diseases review. Nat. Products Bioprospect. 2020, 10, 109–117. [Google Scholar] [CrossRef]
- Jeukendrup, A.E. Periodized nutrition for athletes. Sports Med. 2017, 47, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Sztretye, M.; Dienes, B.; Gönczi, M.; Czirják, T.; Csernoch, L.; Dux, L.; Szentesi, P.; Keller-Pintér, A. Astaxanthin: A potential mitochondrial-targeted antioxidant treatment in diseases and with aging. Oxid. Med. Cell. Longev. 2019. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef]
- Higuera-Ciapara, I.; Félix-Valenzuela, L.; Goycoolea, F.M. Astaxanthin: A Review of its chemistry and applications. Crit. Rev. Food Sci. Nutr. 2006, 46, 185–196. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Polyakov, N.E.; Magyar, A.; Kispert, L.D. Photochemical and optical properties of water-soluble xanthophyll antioxidants: Aggregation vs complexation. J. Phys. Chem. B 2013, 117, 10173–10182. [Google Scholar] [CrossRef]
- Miki, W. Biological functions and activities of animal carotenoids. Pure Appl. Chem. 1991, 63, 141–146. [Google Scholar] [CrossRef]
- Shimidzu, N.; Goto, M.; Miki, W. Carotenoids as singlet oxygen quenchers in marine organisms. Fish. Sci. 1996, 62, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Rohmer, M.; Bouvier, P.; Ourisson, G. Molecular evolution of biomembranes: Structural equivalents and phylogenetic precursors of sterols. Proc. Natl. Acad. Sci. USA 1979, 76, 847–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widomska, J.; Zareba, M.; Subczynski, W. Can xanthophyll-membrane interactions explain their selective presence in the retina and brain? Foods 2016, 5, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vershinin, A. Biological functions of carotenoids—Diversity and evolution. BioFactors 1999, 10, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Oslan, S.N.H.; Shoparwe, N.F.; Yusoff, A.H.; Rahim, A.A.; Chang, C.S.; Tan, J.S.; Oslan, S.N.; Arumugam, K.; Ariff, A.B.; Sulaiman, A.Z.; et al. A review on haematococcus pluvialis bioprocess optimization of green and red stage culture conditions for the production of natural astaxanthin. Biomolecules 2021, 11, 256. [Google Scholar] [CrossRef] [PubMed]
- Capelli, B.; Bagchi, D.; Cysewski, G.R. Synthetic astaxanthin is significantly inferior to algal-based astaxanthin as an antioxidant and may not be suitable as a human nutraceutical supplement. Nutrafoods 2013, 12, 145–152. [Google Scholar] [CrossRef]
- Rüfer, C.E.; Moeseneder, J.; Briviba, K.; Rechkemmer, G.; Bub, A. Bioavailability of astaxanthin stereoisomers from wild (Oncorhynchus spp.) and aquacultured (Salmo salar) salmon in healthy men: A randomised, double-blind study. Br. J. Nutr. 2008, 99, 1048–1054. [Google Scholar] [CrossRef] [Green Version]
- Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Safety of astaxanthin for its use as a novel food in food supplements. EFSA J. 2020, 18. [Google Scholar] [CrossRef] [Green Version]
- Brendler, T.; Williamson, E.M. Astaxanthin: How much is too much? A safety review. Phyther. Res. 2019, 33, 3090–3111. [Google Scholar] [CrossRef]
- Totland, M.Z.; Rasmussen, N.L.; Knudsen, L.M.; Leithe, E. Regulation of gap junction intercellular communication by connexin ubiquitination: Physiological and pathophysiological implications. Cell. Mol. Life Sci. 2020, 77, 573–591. [Google Scholar] [CrossRef] [Green Version]
- Giannaccare, G.; Pellegrini, M.; Senni, C.; Bernabei, F.; Scorcia, V.; Cicero, A.F.G. Clinical Applications of Astaxanthin in the Treatment of Ocular Diseases: Emerging Insights. Mar. Drugs 2020, 18, 239. [Google Scholar] [CrossRef]
- Brown, D.R.; Gough, L.A.; Deb, S.K.; Sparks, S.A.; McNaughton, L.R. Astaxanthin in Exercise Metabolism, Performance and Recovery: A Review. Front. Nutr. 2018, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.N.; Patil, S.; Barkate, H. Protective effects of astaxanthin on skin: Recent scientific evidence, possible mechanisms, and potential indications. J. Cosmet. Dermatol. 2020, 19, 22–27. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Koyama, T.; Takahashi, J.; Yazawa, K. Effects of astaxanthin supplementation on exercise-induced fatigue in mice. Biol. Pharm. Bull. 2006, 29, 2106–2110. [Google Scholar] [CrossRef] [Green Version]
- Aoi, W.; Naito, Y.; Takanami, Y.; Ishii, T.; Kawai, Y.; Akagiri, S.; Kato, Y.; Osawa, T.; Yoshikawa, T. Astaxanthin improves muscle lipid metabolism in exercise via inhibitory effect of oxidative CPT I modification. Biochem. Biophys. Res. Commun. 2008, 366, 892–897. [Google Scholar] [CrossRef]
- Polotow, T.G.; Vardaris, C.V.; Mihaliuc, A.R.; Gonçalves, M.S.; Pereira, B.; Ganini, D.; Barros, M.P. Astaxanthin supplementation delays physical exhaustion and prevents redox imbalances in plasma and soleus muscles of wistar rats. Nutrients 2014, 6, 5819–5838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoi, W.; Maoka, T.; Abe, R.; Fujishita, M.; Tominaga, K. Comparison of the effect of non-esterified and esterified astaxanthins on endurance performance in mice. J. Clin. Biochem. Nutr. 2018, 62, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibaguchi, T.; Yamaguchi, Y.; Miyaji, N.; Yoshihara, T.; Naito, H.; Goto, K.; Ohmori, D.; Yoshioka, T.; Sugiura, T. Astaxanthin intake attenuates muscle atrophy caused by immobilization in rats. Physiol. Rep. 2016, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.H.; Aoi, W.; Takami, M.; Terajima, H.; Tanimura, Y.; Naito, Y.; Itoh, Y.; Yoshikawa, T. The astaxanthin-induced improvement in lipid metabolism during exercise is mediated by a PGC-1α increase in skeletal muscle. J. Clin. Biochem. Nutr. 2014, 54, 86–89. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Baker, J.S.; Chen, X.; Wang, Y.; Chen, H.; Davison, G.W.; Yan, X. High-dose astaxanthin supplementation suppresses antioxidant enzyme activity during moderate-intensity swimming training in mice. Nutrients 2019, 11, 1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, Y.; Nawaz, A.; Kado, T.; Takikawa, A.; Igarashi, Y.; Onogi, Y.; Wada, T.; Sasaoka, T.; Yamamoto, S.; Sasahara, M.; et al. Astaxanthin stimulates mitochondrial biogenesis in insulin resistant muscle via activation of AMPK pathway. J. Cachexia Sarcopenia Muscle 2020, 11, 241–258. [Google Scholar] [CrossRef] [Green Version]
- Yook, J.S.; Okamoto, M.; Rakwal, R.; Shibato, J.; Lee, M.C.; Matsui, T.; Chang, H.; Cho, J.Y.; Soya, H. Astaxanthin supplementation enhances adult hippocampal neurogenesis and spatial memory in mice. Mol. Nutr. Food Res. 2016, 60, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Yook, J.S.; Rakwal, R.; Shibato, J.; Takahashi, K.; Koizumi, H.; Shima, T.; Ikemoto, M.J.; Oharomari, L.K.; McEwen, B.S.; Soya, H. Leptin in hippocampus mediates benefits of mild exercise by an antioxidant on neurogenesis and memory. Proc. Natl. Acad. Sci. USA 2019, 166, 10988–10993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambati, R.; Phang, S.-M.; Ravi, S.; Aswathanarayana, R. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Raz, L.; Knoefel, J.; Bhaskar, K. The neuropathology and cerebrovascular mechanisms of dementia. J. Cereb. Blood Flow Metab. 2016, 36, 172–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hongo, N.; Takamura, Y.; Nishimaru, H.; Matsumoto, J.; Tobe, K.; Saito, T.; Saido, T.C.; Nishijo, H. Astaxanthin ameliorated parvalbumin-positive neuron deficits and Alzheimer’s Disease-related pathological progression in the hippocampus of AppNL-G-F/NL-G-F Mice. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Matsuba, Y.; Mihira, N.; Takano, J.; Nilsson, P.; Itohara, S.; Iwata, N.; Saido, T.C. Single App knock-in mouse models of Alzheimer’s disease. Nat. Neurosci. 2014, 17, 661–663. [Google Scholar] [CrossRef]
- Choi, H.D.; Kang, H.E.; Yang, S.H.; Lee, M.G.; Shin, W.G. Pharmacokinetics and first-pass metabolism of astaxanthin in rats. Br. J. Nutr. 2011, 105, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Maoka, T.; Tsuji, S.; Hayashi, M.; Shimazawa, M.; Hara, H. Central nervous system migration of astaxanthin and adonixanthin following their oral administration in cynomolgus monkeys. J. Nutr. Sci. Vitaminol. 2020, 66, 488–494. [Google Scholar] [CrossRef]
- Ahlskog, J.E.; Geda, Y.E.; Graff-Radford, N.R.; Petersen, R.C. Physical exercise as a preventive or disease-modifying treatment of dementia and brain aging. Mayo Clin. Proc. 2011, 86, 876–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenco, M.V.; Frozza, R.L.; de Freitas, G.B.; Zhang, H.; Kincheski, G.C.; Ribeiro, F.C.; Gonçalves, R.A.; Clarke, J.R.; Beckman, D.; Staniszewski, A.; et al. Exercise-linked FNDC5/irisin rescues synaptic plasticity and memory defects in Alzheimer’s models. Nat. Med. 2019, 25, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Bylykbashi, E.; Chatila, Z.K.; Lee, S.W.; Pulli, B.; Clemenson, G.D.; Kim, E.; Rompala, A.; Oram, M.K.; Asselin, C.; et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science 2018, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titus, J.; Bray, N.W.; Kamkar, N.; Camicioli, R.; Nagamatsu, L.S.; Speechley, M.; Montero-Odasso, M. The role of physical exercise in modulating peripheral inflammatory and neurotrophic biomarkers in older adults: A systematic review and meta-analysis. Mech. Ageing Dev. 2021, 194, 111431. [Google Scholar] [CrossRef]
- Koizumi, H.; Hiraga, T.; Oharomari, L.K.; Hata, T.; Shima, T.; Yook, J.S.; Okamoto, M.; Mouri, A.; Nabeshima, T.; Soya, H. Preventive role of regular low-intensity exercise during adolescence in schizophrenia model mice with abnormal behaviors. Biochem. Biophys. Res. Commun. 2021, 534, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Samieri, C.; Perier, M.C.; Gaye, B.; Proust-Lima, C.; Helmer, C.; Dartigues, J.F.; Berr, C.; Tzourio, C.; Empana, J.P. Association of cardiovascular health level in older age with cognitive decline and incident dementia. JAMA J. Am. Med. Assoc. 2018, 320, 657–664. [Google Scholar] [CrossRef]
- Gómez-Pinilla, F. Brain foods: The effects of nutrients on brain function. Nat. Rev. Neurosci. 2008, 9, 568–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fares, J.; Bou Diab, Z.; Nabha, S.; Fares, Y. Neurogenesis in the adult hippocampus: History, regulation, and prospective roles. Int. J. Neurosci. 2019, 129, 598–611. [Google Scholar] [CrossRef] [PubMed]
- McGregor, G.; Harvey, J. Regulation of hippocampal synaptic function by the metabolic hormone, leptin: Implications for health and neurodegenerative disease. Front. Cell. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Fedewa, M.V.; Hathaway, E.D.; Ward-Ritacco, C.L.; Williams, T.D.; Dobbs, W.C. The effect of chronic exercise training on leptin: A systematic review and meta-analysis of randomized controlled trials. Sports Med. 2018, 48, 1437–1450. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Fry, A.; Schilling, B.; Chiu, L.; Hori, N.; Weiss, L. Astaxanthin supplementation does not attenuate muscle injury following eccentric exercise in resistance-trained men. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Res, P.T.; Cermak, N.M.; Stinkens, R.; Tollakson, T.J.; Haenen, G.R.; Bast, A.; Van Loon, L.J.C. Astaxanthin supplementation does not augment fat use or improve endurance performance. Med. Sci. Sports Exerc. 2013, 45, 1158–1165. [Google Scholar] [CrossRef]
- Klinkenberg, L.J.J.; Res, P.T.; Haenen, G.R.; Bast, A.; Van Loon, L.J.C.; Van Dieijen-Visser, M.P.; Meex, S.J.R. Effect of antioxidant supplementation on exercise-induced cardiac troponin release in cyclists: A randomized trial. PLoS ONE 2013, 8, e79280. [Google Scholar] [CrossRef]
- Earnest, C.P.; Lupo, M.; White, K.M.; Church, T.S. Effect of astaxanthin on cycling time trial performance. Int. J. Sports Med. 2011, 32, 882–888. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Warner, A.R.; Deb, S.K.; Gough, L.A.; Sparks, S.A.; McNaughton, L.R. The effect of astaxanthin supplementation on performance and fat oxidation during a 40 km cycling time trial. J. Sci. Med. Sport 2021, 24, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, B.; Baralic, I.; Kotur-Stevuljevic, J.; Stefanovic, A.; Ivanisevic, J.; Radivojevic, N.; Andjelkovic, M.; Dikic, N. Effect of astaxanthin supplementation on muscle damage and oxidative stress markers in elite young soccer players. J. Sports Med. Phys. Fit. 2012, 52, 382–392. [Google Scholar]
- Madhavi, D.; Kagan, D.; Seshadri, S. A Study on the bioavailability of a proprietary, sustained-release formulation of astaxanthin. Integr. Med. 2018, 17, 38–42. [Google Scholar]
- Yang, L.; Qiao, X.; Gu, J.; Li, X.; Cao, Y.; Xu, J.; Xue, C. Influence of molecular structure of astaxanthin esters on their stability and bioavailability. Food Chem. 2021, 343, 128497. [Google Scholar] [CrossRef]
- Qiao, X.; Yang, L.; Zhang, T.; Zhou, Q.; Wang, Y.; Xu, J.; Xue, C. Synthesis, stability and bioavailability of astaxanthin succinate diester. J. Sci. Food Agric. 2018, 98, 3182–3189. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, S.; McClements, D.J.; Wang, D.; Xu, Y. Design of astaxanthin-loaded core–shell nanoparticles consisting of chitosan oligosaccharides and Poly(lactic-co-glycolic acid): Enhancement of water solubility, stability, and bioavailability. J. Agric. Food Chem. 2019, 67, 5113–5121. [Google Scholar] [CrossRef]
- Martínez-Álvarez, Ó.; Calvo, M.M.; Gómez-Estaca, J. Recent advances in astaxanthin micro/nanoencapsulation to improve its stability and functionality as a food ingredient. Mar. Drugs 2020, 18, 406. [Google Scholar] [CrossRef]
- Mercke Odeberg, J.; Lignell, Å.; Pettersson, A.; Höglund, P. Oral bioavailability of the antioxidant astaxanthin in humans is enhanced by incorporation of lipid based formulations. Eur. J. Pharm. Sci. 2003, 19, 299–304. [Google Scholar] [CrossRef]
- Visioli, F.; Artaria, C. Astaxanthin in cardiovascular health and disease: Mechanisms of action, therapeutic merits, and knowledge gaps. Food Funct. 2017, 8, 39–63. [Google Scholar] [CrossRef]
- Kistler, A.; Liechti, H.; Pichard, L.; Wolz, E.; Oesterhelt, G.; Hayes, A.; Maurel, P. Metabolism and CYP-inducer properties of astaxanthin in man and primary human hepatocytes. Arch. Toxicol. 2002, 75, 665–675. [Google Scholar] [CrossRef]
- Clark, R.M.; Yao, L.; She, L.; Furr, H.C. A comparison of lycopene and astaxanthin absorption from corn oil and olive oil emulsions. Lipids 2000, 35, 803–806. [Google Scholar] [CrossRef]
- OKADA, Y.; ISHIKURA, M.; MAOKA, T. Bioavailability of astaxanthin in haematococcus Algal Extract: The effects of timing of diet and smoking habits. Biosci. Biotechnol. Biochem. 2009, 73, 1928–1932. [Google Scholar] [CrossRef] [Green Version]
- Mason, S.A.; Trewin, A.J.; Parker, L.; Wadley, G.D. Antioxidant supplements and endurance exercise: Current evidence and mechanistic insights. Redox Biol. 2020, 35, 101471. [Google Scholar] [CrossRef]
- Pasanen, T.; Tolvanen, S.; Heinonen, A.; Kujala, U.M. Exercise therapy for functional capacity in chronic diseases: An overview of meta-analyses of randomised controlled trials. Br. J. Sports Med. 2017, 51, 1459–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimmig, B.; Kim, S.H.; Nash, K.; Bickford, P.C.; Douglas Shytle, R. Neuroprotective mechanisms of astaxanthin: A potential therapeutic role in preserving cognitive function in age and neurodegeneration. GeroScience 2017, 39, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Ludyga, S.; Gerber, M.; Pühse, U.; Looser, V.N.; Kamijo, K. Systematic review and meta-analysis investigating moderators of long-term effects of exercise on cognition in healthy individuals. Nat. Hum. Behav. 2020, 4, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Kivipelto, M.; Mangialasche, F.; Ngandu, T. Lifestyle interventions to prevent cognitive impairment, dementia and Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 653–666. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author (Year) | Animal (Age) | Intervention (Duration) | Primary Outcome | Results |
|---|---|---|---|---|
| M. Ikeuchi (2006) | ddY mice (4 weeks) | ASX: 30 mg/kg b.w. (gavage) Exe: swimming to exhaustion (5 weeks) | Endurance performance | Increased time to exhaustion by increasing lipid metabolism [33] |
| W. Aoi (2008) | ICR mice (7 weeks) | ASX: 0.02% w/w Exe: 2/wk, 18 m/min—5 min (4 weeks) | Endurance performance | Increased time to exhaustion by increasing lipid metabolism [34] |
| H. Liu (2014) | ICR mice (7 weeks) | ASX: 0.02% w/w Exe: 30 min 25 m/min treadmill (2 weeks) | Lipid metabolism | Increased PGC-1alpha in skeletal muscle [38] |
| T. G. Polotow (2014) | Wistar rats (NA) | ASX: 1 mg/kg bw (gavage) Exe: swimming to exhaustion (6 weeks) | Endurance performance | Increased time to exhaustion by redox balance [35] |
| T. Shibaguchi (2016) | Wistar rats (14 weeks) | ASX: 0.04% w/w (6 weeks) | Muscle atrophy | Attenuated skeletal muscle atrophy by redox balance [37] |
|
W. Aoi (2017) |
ICR mice (8 weeks) |
ASX: 0.02% w/w Exe: 3/wk, 25 m/min—5 min (5 weeks) |
Endurance performance | Increased time to exhaustion [36] |
| Y Zhou (2019) | C57BL/6J mice (7 weeks) | ASX: 30 mg/kg bw (gavage) Exe: 45 min moderate swimming (4 weeks) | Redox status | Suppressed antioxidant enzyme activity [39] |
| Y. Nishida (2020) | C57BL/6J mice (6 weeks) | ASX: 0.02% w/w (24 weeks) | Insulin resistance | Increased mitochondria biogenesis via AMPK pathway [40] |
| J. S. Yook (2016) | C57BL/6J mice (11 weeks) | ASX: 0.5% w/w (4 weeks) | Cognitive function | Increased spatial memory by increasing hippocampal neurogenesis [41] |
| J. S. Yook (2019) | C57BL/6J mice (11 weeks) | ASX: 0.5% w/w Exe: mild treadmill running (4 weeks) | Cognitive function | Increased spatial memory with increasing hippocampal neurogenesis [42] |
| Author (Year) | Subjects | Intervention | Primary Outcome | Results |
|---|---|---|---|---|
| R.J. Bloomer (2005) | Resistance-trained males | 4 mg/d, 3 weeks | Muscle performance | No difference [59] |
| C. P. Earnest (2011) | Amateur endurance-trained males | 4 mg/d, 4 weeks | TT performance | Improved performance [62] |
| B. Djordjevic (2012) | Male elite soccer players | 4 mg/d, 12 weeks | Redox status | Stress oxidative prevention [64] |
| P.T. Res (2012) | Well-trained male cyclists or triathletes | 20 mg/d, 4 weeks | TT performance | No effect [60] |
| L.J.J. Klinkenberg (2013) | Well-trained male cyclists | 20 mg/d, 4 weeks | Redox status | No effect [61] |
| D.R. Brown (2021) | Trained male cyclists | 12 mg/d, 1 weeks | TT performance | Improved performance [63] |
| Author (Year) | Purpose of the Study | Practical Value |
|---|---|---|
| S. A. Mason (2020) | The authors performed an extensive and critical literature review regarding the most common antioxidant supplements for athletes (e.g., ASX, catechins, curcumin, quercetin, resveratrol, vitamin C., etc.) | After considering ASX evidence, the authors concluded that there is a lack of evidence to support it as a supplement [75] |
| D. R. Brown (2018) | In this review, the authors rigorously interpreted scientific literature regarding the ergogenic effects of ASX. Although they recognized the promising data found in in vitro and in vivo research, human studies were not satisfactory. | Their final message was that future investigation is needed regarding ASX ergogenic effects in humans [31]. |
| B. Grimmig (2017) | In this narrative review, the authors introduced a wide range of evidence from basic studies to clinical trials for the possible effects of ASX on cognitive function. | After the discussion, the authors conclude that ASX is a promising therapeutic agent for neurodegenerative diseases [77]. |
| J.P. Yaun (2011) | Using a broad approach, ASX’s biological effects were reviewed. The authors summarized the scientific evidence for ASX’s effect on several disease conditions. | Finally, the authors stated that although it is encouraging, more extensive and well-controlled clinical trials are necessary [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oharomari, L.K.; Ikemoto, M.J.; Hwang, D.J.; Koizumi, H.; Soya, H. Benefits of Exercise and Astaxanthin Supplementation: Are There Additive or Synergistic Effects? Antioxidants 2021, 10, 870. https://doi.org/10.3390/antiox10060870
Oharomari LK, Ikemoto MJ, Hwang DJ, Koizumi H, Soya H. Benefits of Exercise and Astaxanthin Supplementation: Are There Additive or Synergistic Effects? Antioxidants. 2021; 10(6):870. https://doi.org/10.3390/antiox10060870
Chicago/Turabian StyleOharomari, Leandro Kansuke, Mitsushi J. Ikemoto, Dong Joo Hwang, Hikaru Koizumi, and Hideaki Soya. 2021. "Benefits of Exercise and Astaxanthin Supplementation: Are There Additive or Synergistic Effects?" Antioxidants 10, no. 6: 870. https://doi.org/10.3390/antiox10060870