
Blood Plasma Quality Control by Plasma Glutathione Status

 ,
,  , , , ,
, , , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
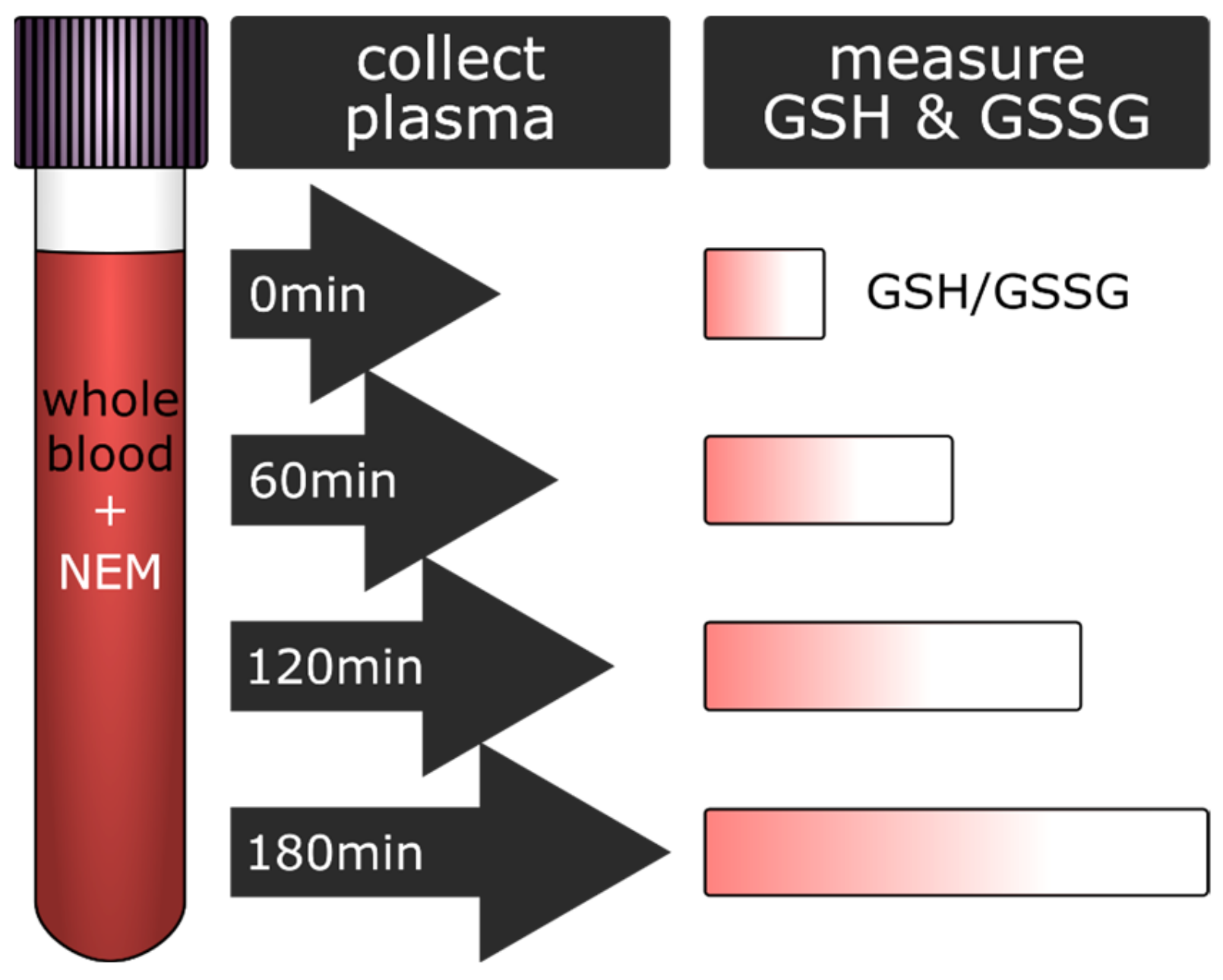
2.1. Study Design
2.2. Sample Collection and Plasma Preparation
2.3. Clinical Routine Hematology
2.4. Glutathione Measurements
2.5. Lactate Dehydrogenase (LDH) Activity Assay
2.6. Statistical Analysis
3. Results
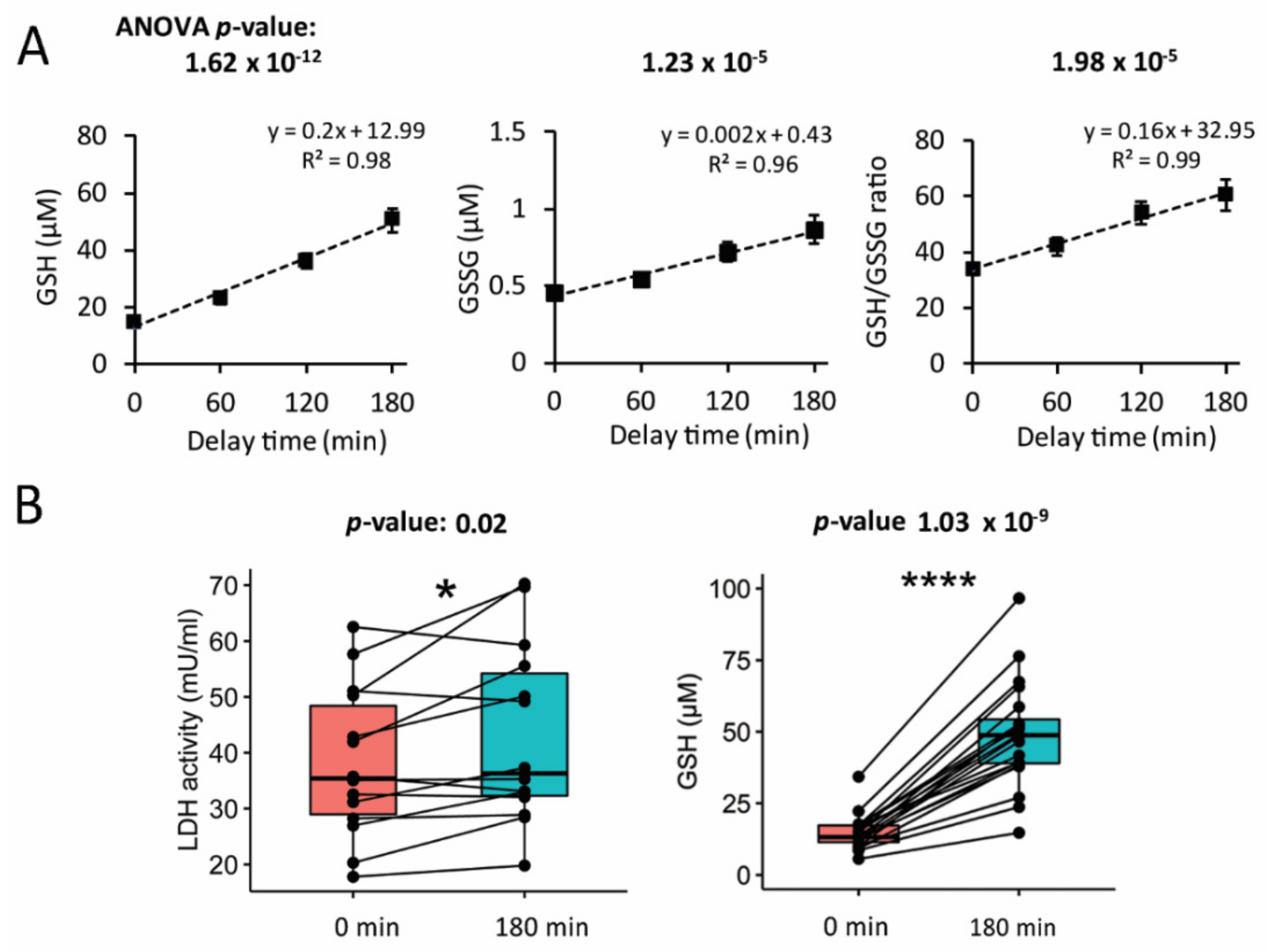
3.1. Glutathione Content in Plasma Increased with the Delay in Plasma Preparation
3.2. Increase in Glutathione Was Accompanied by Higher Lactate Dehydrogenase Activity of the Plasma
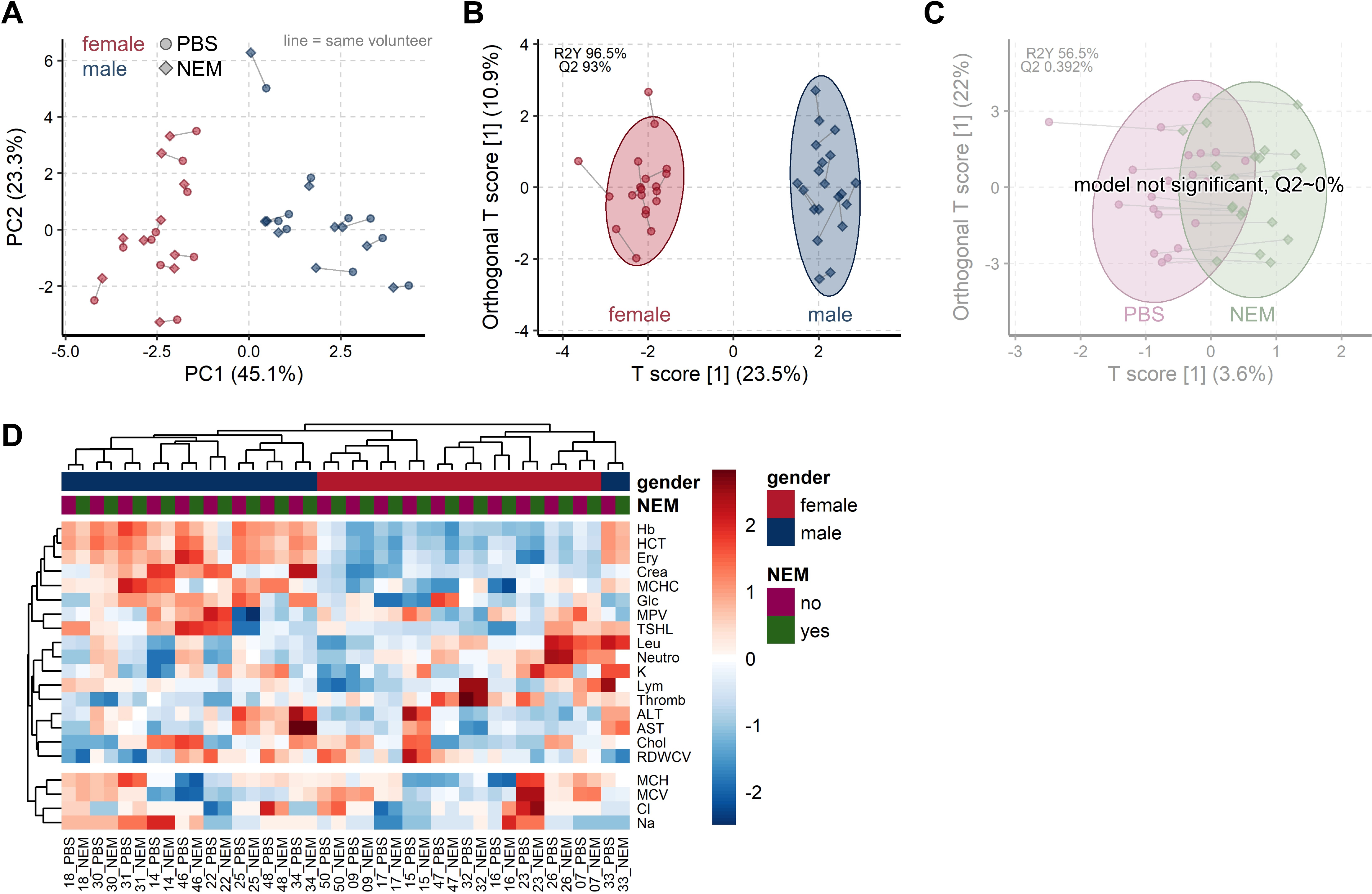
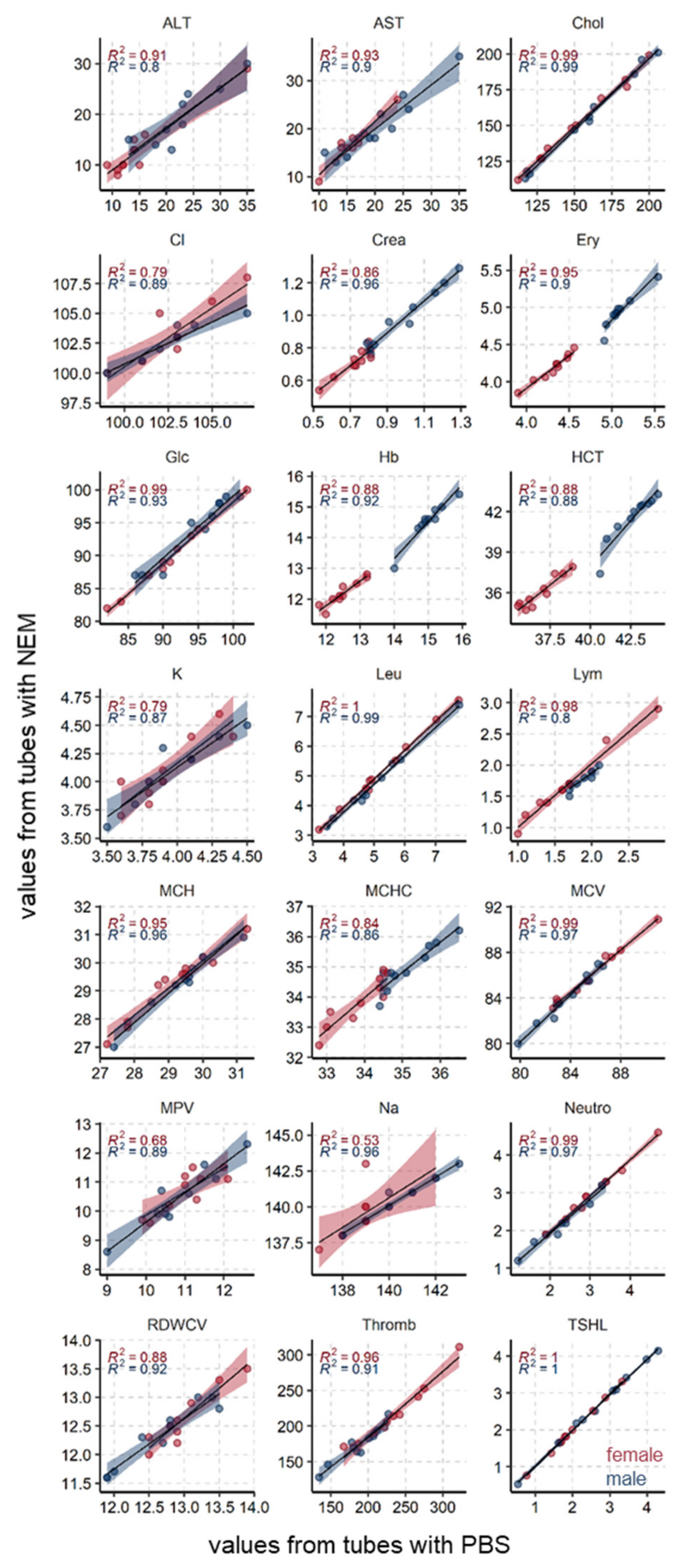
3.3. Common Blood Parameters Were Not Affected by NEM-Spiked Tubes
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuck, M.K.; Chan, D.W.; Chia, D.; Godwin, A.K.; Grizzle, W.E.; Krueger, K.E.; Rom, W.; Sanda, M.; Sorbara, L.; Stass, S.; et al. Standard Operating Procedures for Serum and Plasma Collection: Early Detection Research Network Consensus Statement Standard Operating Procedure Integration Working Group. J. Proteome Res. 2009, 8, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Godoy, L.M.; Enríquez-Cárcamo, V.; Suárez-Roa, L.; Lopez-Castro, M.L.; Santamaría, A.; Orozco-Morales, M.; Colín-González, A.L. Identification of Specific Pre-analytical Quality Control Markers in Plasma and Serum Samples. Anal. Methods 2019, 11, 2259–2271. [Google Scholar] [CrossRef]
- Kamlage, B.; Maldonado, S.G.; Bethan, B.; Peter, E.; Schmitz, O.; Liebenberg, V.; Schatz, P. Quality Markers Addressing Preanalytical Variations of Blood and Plasma Processing Identified by Broad and Targeted Metabolite Profiling. Clin. Chem. 2014, 60, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Bernini, P.; Bertini, I.; Luchinat, C.; Nincheri, P.; Staderini, S.; Turano, P. Standard Operating Procedures for Pre-analytical Handling of Blood and Urine for Metabolomic Studies and Biobanks. J. Biomol. NMR 2011, 49, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.-Y.; Chen, R.-K.; Pan, Y.-H.; Lee, H.-L. Systematical Evaluation of the Effects of Sample Collection Procedures on Low-molecular-weight Serum/Plasma Proteome Profiling. Proteomics 2006, 6, 3189–3198. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Hong, M.; Park, S.-K.; Yu, J.-I.; Wang, J.-S.; Shin, H.; Kim, J.-W.; Han, B.-G.; Shin, S.-Y. Inorganic Phosphorus and Potassium Are Putative Indicators of Delayed Separation of Whole Blood. Osong Public Heal. Res. Perspect. 2016, 7, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-E.; Kim, J.-W.; Han, B.-G.; Shin, S.-Y. Impact of Whole-Blood Processing Conditions on Plasma and Serum Concentrations of Cytokines. Biopreservation Biobanking 2016, 14, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Giustarini, D.; Colombo, G.; Garavaglia, M.L.; Astori, E.; Portinaro, N.M.; Reggiani, F.; Badalamenti, S.; Aloisi, A.M.; Santucci, A.; Rossi, R.; et al. Assessment of Glutathione/Glutathione Disulphide Ratio and S-glutathionylated Proteins in Human Blood, Solid Tissues, and Cultured Cells. Free Radic. Biol. Med. 2017, 112, 360–375. [Google Scholar] [CrossRef] [PubMed]
- Tomin, T.; Schittmayer, M.; Birner-Gruenberger, R. Addressing Glutathione Redox Status in Clinical Samples by Two-Step Alkylation with N-ethylmaleimide Isotopologues. Metabolites 2020, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Giustarini, D.; Galvagni, F.; Orlandini, M.; Fanti, P.; Rossi, R. Immediate Stabilization of Human Blood for Delayed Quantification of Endogenous Thiols and Disulfides. J. Chromatogr. B 2016, 1019, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Sutton, T.; Minnion, M.; Barbarino, F.; Koster, G.; Fernandez, B.; Cumpstey, A.; Wischmann, P.; Madhani, M.; Frenneaux, M.; Postle, A.; et al. A Robust and Versatile Mass Spectrometry Platform for Comprehensive Assessment of the Thiol Redox Metabolome. Redox Biol. 2018, 16, 359–380. [Google Scholar] [CrossRef]
- Paulech, J.; Solis, N.; Cordwell, S. Characterization of Reaction Conditions Providing Rapid and Specific Cysteine Alkylation for Peptide-based Mass Spectrometry. Biochim. Biophys. Acta Proteins Proteom. 2013, 1834, 372–379. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Raymaekers, J.; Rousseeuw, P. cellWise: Analyzing Data with Cellwise Outliers, R Package Version 2.2.3. 2020. Available online: https://CRAN.R-project.org/package=cellWise (accessed on 19 May 2021).
- Chong, J.; Xia, J. MetaboAnalystR: An R Package for Flexible and Reproducible Analysis of Metabolomics Data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef] [Green Version]
- Hubert, M.; Rousseeuw, P.J.; Bossche, W.V.D. MacroPCA: An All-in-One PCA Method Allowing for Missing Values as Well as Cellwise and Rowwise Outliers. Technometrics 2019, 61, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Erve, T.J.V.T.; Wagner, B.A.; Ryckman, K.K.; Raife, T.J.; Buettner, G.R. The Concentration of Glutathione in Human Erythrocytes is a Heritable Trait. Free Radic. Biol. Med. 2013, 65, 742–749. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the Lactate Dehydrogenase Assay. Cold Spring Harb. Protoc. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Greening, D.W.; Simpson, R.J. Serum/Plasma Proteomics; Springer: New York, NY, USA, 2017. [Google Scholar]
- Makhro, A.; Huisjes, R.; Verhagen, L.P.; Mañú-Pereira, M.D.M.; Llaudet-Planas, E.; Petkova-Kirova, P.; Wang, J.; Eichler, H.; Bogdanova, A.; Van Wijk, R.; et al. Red Cell Properties after Different Modes of Blood Transportation. Front. Physiol. 2016, 7, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Zhu, W.; Wang, Y.; Wei, X.; Li, J.; Peng, L.; Zhang, K.; Bai, B. A Reference Chart for Clinical Biochemical Tests of Hemolyzed Serum Samples. J. Clin. Lab. Anal. 2021, 35, 23561. [Google Scholar] [CrossRef]
- Ballatori, N.; Krance, S.M.; Marchan, R.; Hammond, C.L. Plasma Membrane Glutathione Transporters and Their Roles in Cell Physiology and Pathophysiology. Mol. Asp. Med. 2009, 30, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Prenesti, E.; Berto, S.; Gosmaro, F.; Bagnati, M.; Bellomo, G. Biomolecules Responsible for the Total Antioxidant Capacity (TAC) of Human Plasma in Healthy and Cardiopathic Individuals: A Chemical Speciation Model. Antioxidants 2021, 10, 656. [Google Scholar] [CrossRef] [PubMed]
- Ricós, V.A.C. Current Databases on Biological Variation: Pros, Cons and Progress. Scand. J. Clin. Lab. Investig. 1999, 59, 491–500. [Google Scholar] [CrossRef]
- Perich, C.; Minchinela, J.; Ricós, C.; Fernández-Calle, P.; Alvarez, V.; Doménech, M.V.; Simón, M.; Biosca, C.; Boned, B.; García-Lario, J.V.; et al. Biological Variation Database: Structure and Criteria Used for Generation and Update. Clin. Chem. Lab. Med. 2015, 53, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Carobene, A.; Braga, F.; Roraas, T.; Sandberg, S.; Bartlett, W.A. A Systematic Review of Data on Biological Variation for Alanine Aminotransferase, Aspartate Aminotransferase and γ-glutamyl Transferase. Clin. Chem. Lab. Med. 2013, 51, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Coşkun, A.; Carobene, A.; Kilercik, M.; Serteser, M.; Sandberg, S.; Aarsand, A.K.; Fernandez-Calle, P.; Jonker, N.; Bartlett, W.A.; Díaz-Garzón, J.; et al. Within-subject and between-subject Biological Variation Estimates of 21 Hematological Parameters in 30 Healthy Subjects. Clin. Chem. Lab. Med. 2018, 56, 1309–1318. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Parent Ion | Product Ion | Collision Energy |
|---|---|---|---|
| GSH | 308 | 179 | 19 |
| GSH-NEM (Transition 1) | 433.16 | 201 | 22 |
| GSH-NEM (Transition 2) | 433.16 | 304 | 15 |
| GSH-d5-NEM (Transition 1) | 438.16 | 206 | 22 |
| GSH-d5-NEM (Transition 2) | 438.16 | 309 | 15 |
| 13C2, 15N-GSH-d5-NEM (IS) | 441.16 | 206 | 22 |
| GSSG | 613.2 | 355.3 | 10 |
| Abbreviation | Full Name | Mean (PBS) | Mean (NEM) | CVw (%) | Reference Range | Unit | |
|---|---|---|---|---|---|---|---|
| Male | Female | ||||||
| Ery | erythrocytes | 4.7 ± 0.4 | 4.6 ± 0.4 | 2.1 | 4.5–5.9 | 4.1–5.1 | 1012/L |
| Leu | leucocytes | 5.1 ± 1.3 | 5.0 ± 1.2 | 2.6 | 4.4–11.3 | 109/L | |
| Lym | lymphocytes | 1.8 ± 0.5 | 1.7 ± 0.6 | 3.1 | 1–4.8 | 109/L | |
| Mono | monocytes | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.2–1.0 | 109/L | ||
| Eos | eosinophils | 0.2 ± 0.2 | 0.2 ± 0.2 | 0–0.7 | 109/L | ||
| Baso | basophils | 0.0 ± 0.0 | 0.0 ± 0.0 | 0–0.2 | 109/L | ||
| Neutro | neutrophils | 2.8 ± 0.8 | 2.6 ± 1.0 | 2.8 | 1.8–7.7 | 109/L | |
| Thromb | platelets | 210 ± 45 | 195 ± 41 | 5.5 | 140–440 | 109/L | |
| ALT | alanine amino transferase | 18.5 ± 7.8 | 16.1 ± 6.7 | 11.7 | 0–45 | 0–35 | U/L |
| AST | aspartate amino transferase | 18.5 ± 5.9 | 19.0 ± 5.7 | 5.7 | 0–35 | 0–30 | U/L |
| Crea | creatinine | 0.9 ± 0.2 | 0.8 ± 0.2 | 2.3 | 0 - 1.2 | 0–1 | mg/dL |
| Cl | chloride ions | 103 ± 2 | 103 ± 2 | 0.4 | 95–110 | mmol/L | |
| K | potassium ions | 4.0 ± 0.3 | 4.1 ± 0.3 | 2.6 | 3.5–5.0 | mmol/L | |
| Na | sodium ions | 140 ± 2 | 140 ± 2 | 0.2 | 135 | 145 | mmol/L |
| Chol | cholesterol | 156 ± 30 | 154 ± 30 | 1.1 | 0–200 | mg/dL | |
| Glc | glucose | 92.7 ± 5.6 | 91.8 ± 5.5 | 0.8 | 70–100 | mg/dL | |
| Hb | hemoglobin | 13.8 ± 1.4 | 13.4 ± 1.3 | 2.1 | 13–17.5 | 12–15.3 | g/dL |
| HCT | hematocrit | 39.8 ± 3.2 | 38.8 ± 3.2 | 1.8 | 40–50 | 35–45 | % |
| MCH | mean cell hemoglobin | 29.3 ± 1.1 | 29.3 ± 1.1 | 0.5 | 28–33 | pg | |
| MCHC | mean corpuscular hemoglobin concentration | 34.5 ± 1.0 | 34.4 ± 1.0 | 0.5 | 33–36 | g/dL | |
| MCV | mean corpuscular volume | 84.7 ± 2.6 | 85.1 ± 2.5 | 0.3 | 80–98 | fL | |
| MPV | mean platelet volume | 10.9 ± 0.8 | 10.6 ± 0.9 | 2.9 | 7–13 | fL | |
| RDWCV | Red blood cell distribution width | 12.8 ± 0.5 | 12.5 ± 0.5 | 1.9 | 11–16 | % | |
| TSHL | thyroid - stimulating hormone | 2.4 ± 1.0 | 2.3 ± 1.0 | 1.1 | 0.10–4.00 | µU/mL | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomin, T.; Bordag, N.; Zügner, E.; Al-Baghdadi, A.; Schinagl, M.; Birner-Gruenberger, R.; Schittmayer, M. Blood Plasma Quality Control by Plasma Glutathione Status. Antioxidants 2021, 10, 864. https://doi.org/10.3390/antiox10060864
Tomin T, Bordag N, Zügner E, Al-Baghdadi A, Schinagl M, Birner-Gruenberger R, Schittmayer M. Blood Plasma Quality Control by Plasma Glutathione Status. Antioxidants. 2021; 10(6):864. https://doi.org/10.3390/antiox10060864
Chicago/Turabian StyleTomin, Tamara, Natalie Bordag, Elmar Zügner, Abdullah Al-Baghdadi, Maximilian Schinagl, Ruth Birner-Gruenberger, and Matthias Schittmayer. 2021. "Blood Plasma Quality Control by Plasma Glutathione Status" Antioxidants 10, no. 6: 864. https://doi.org/10.3390/antiox10060864