
Differential and Overlapping Effects of Melatonin and Its Metabolites on Keratinocyte Function: Bioinformatics and Metabolic Analyses

 ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Treatment
2.3. Reverse Transcription Reaction and Real-Time PCR
2.4. RNA Sequencing and Bioinformatics Analysis
2.5. Bioinformatics
2.6. Analysis of Metabolic Function
2.7. Proteome Profiler and L-Lactate Functional Assays
2.8. Statistical Analysis
3. Results and Discussion
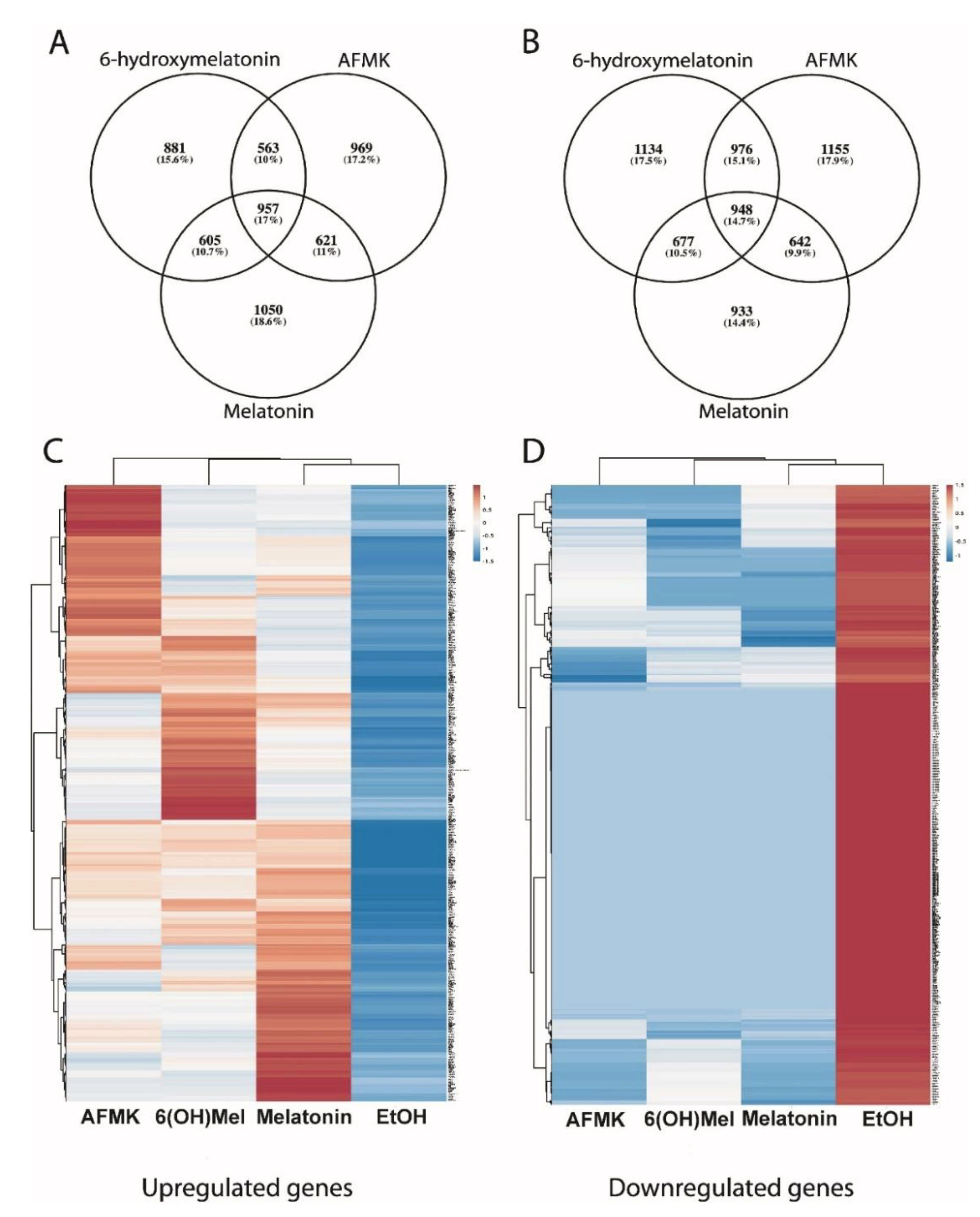
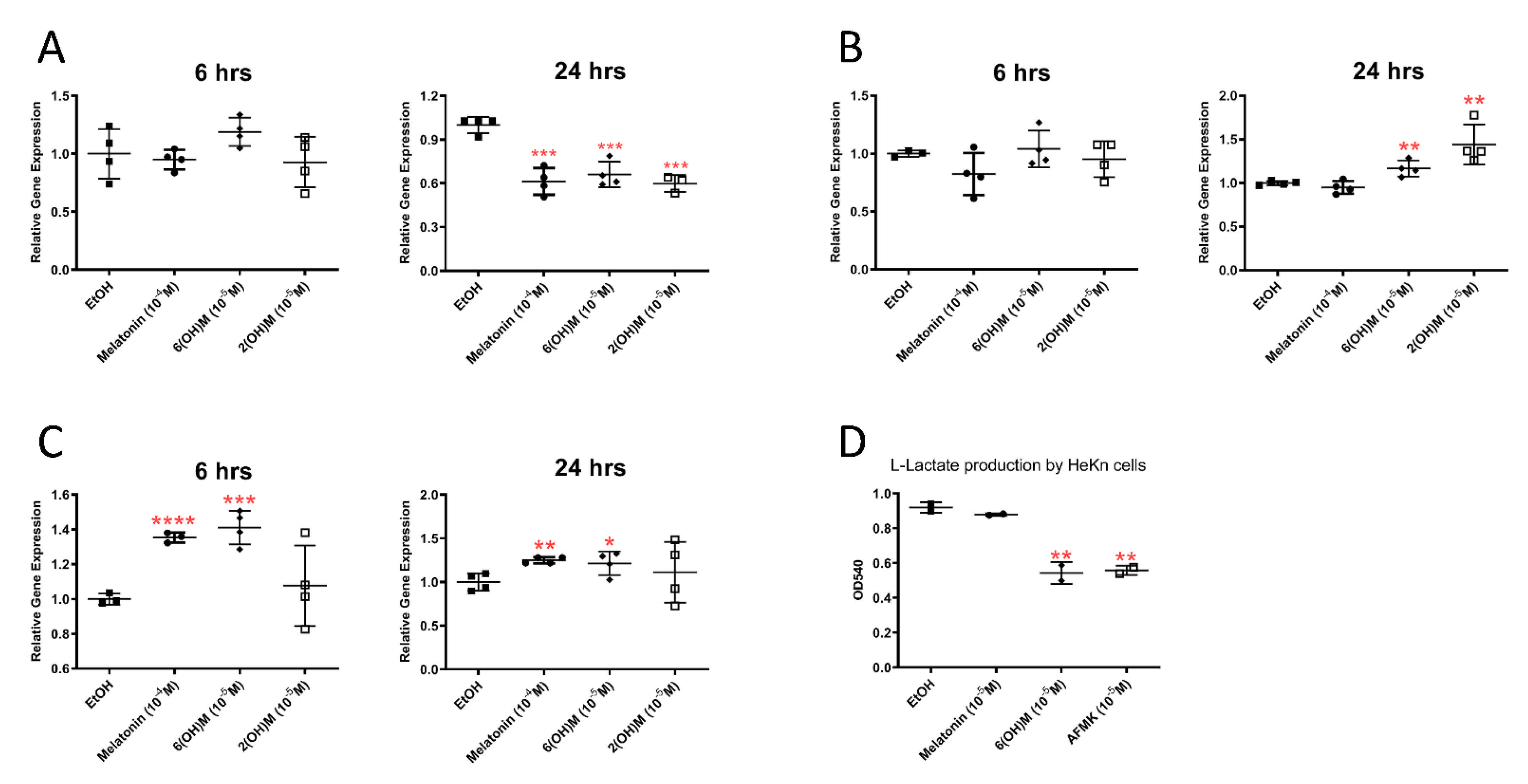
3.1. RNAseq Results
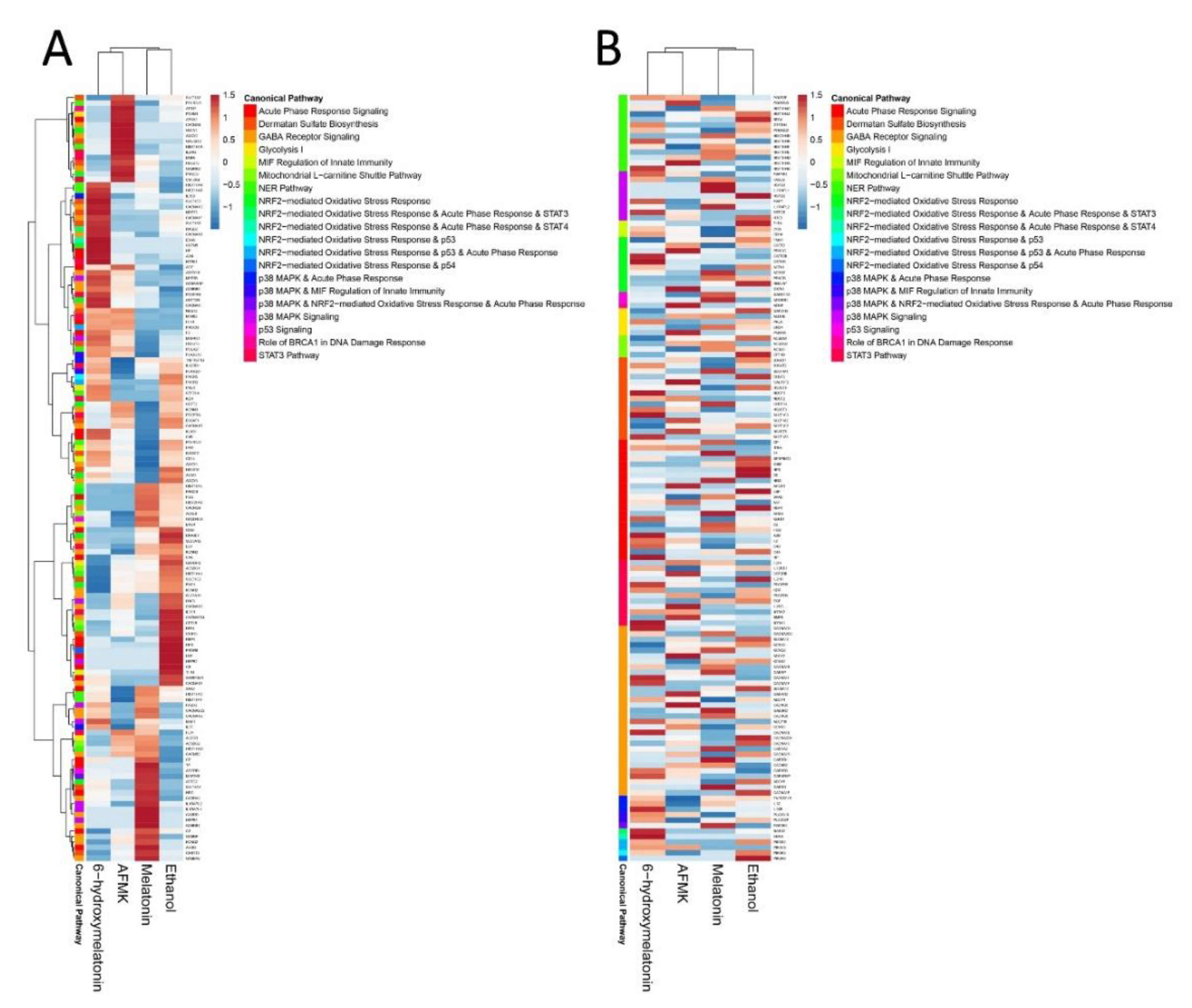
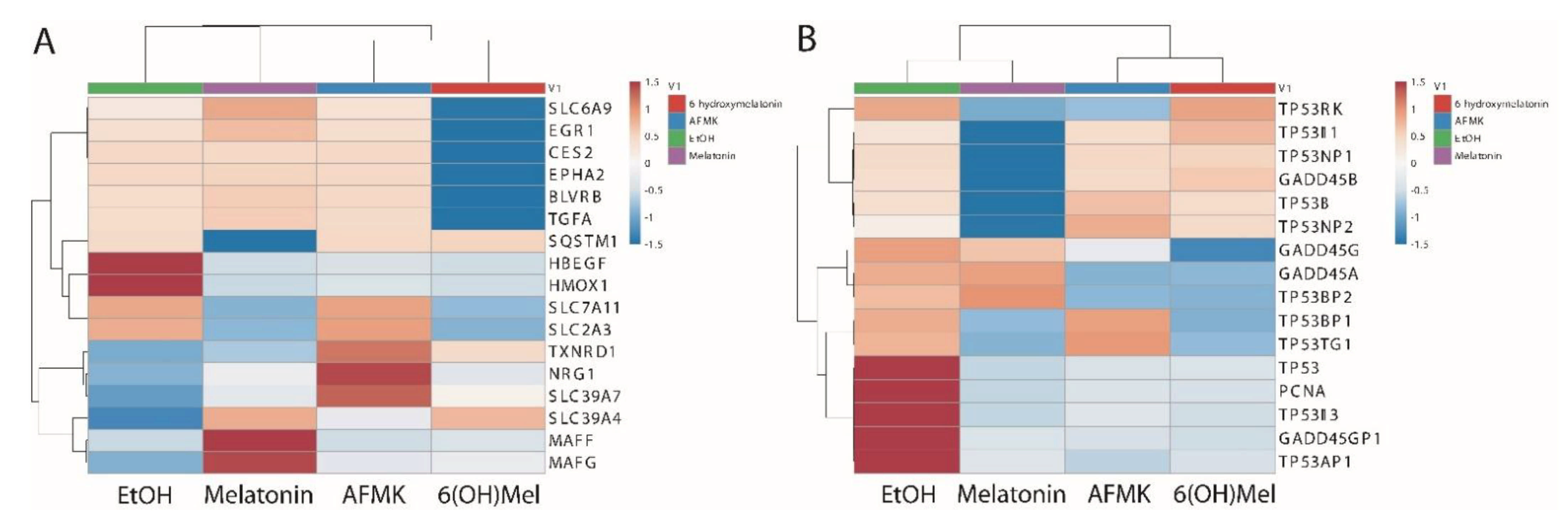
3.2. Bioinformatics Analysis
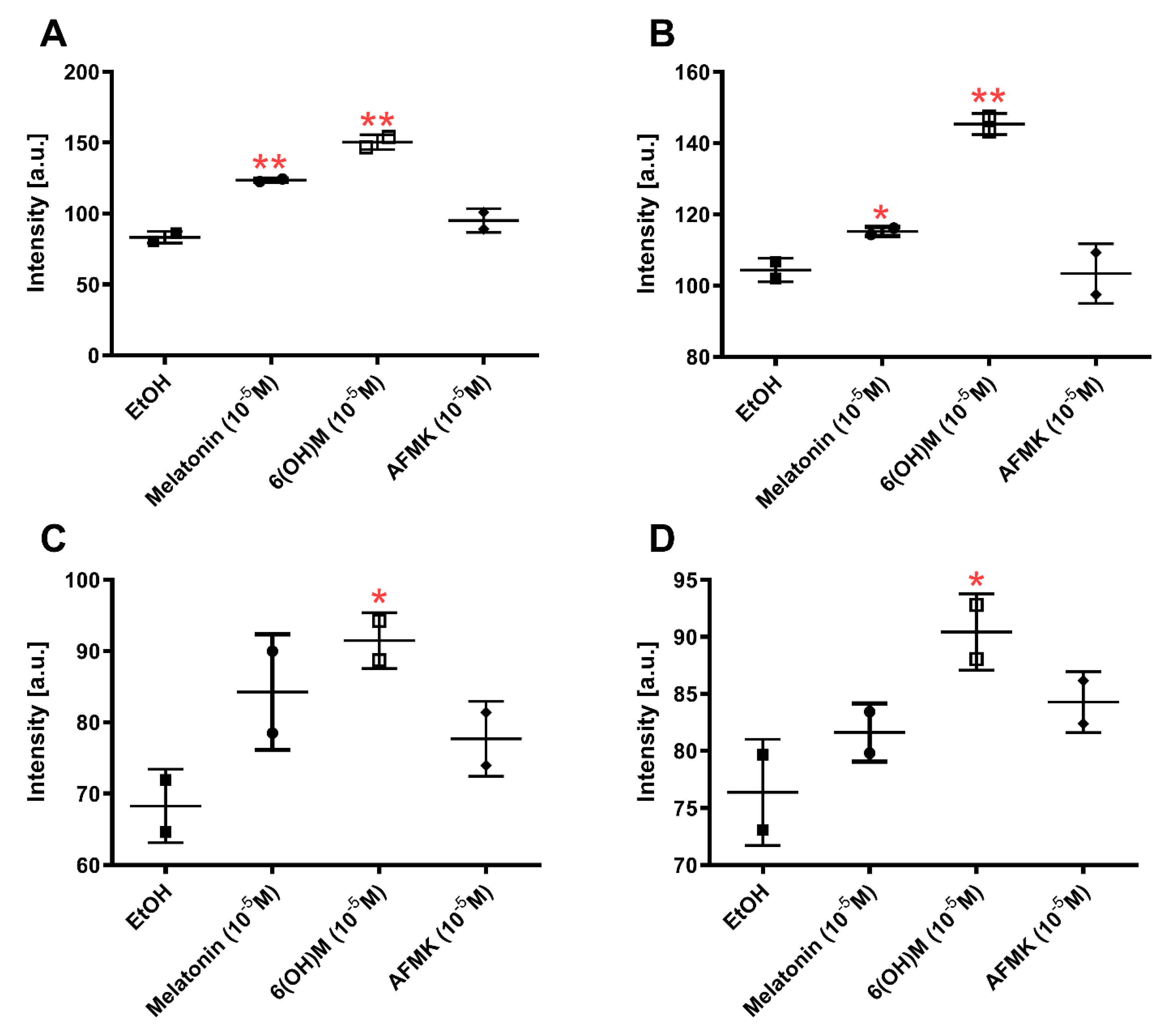
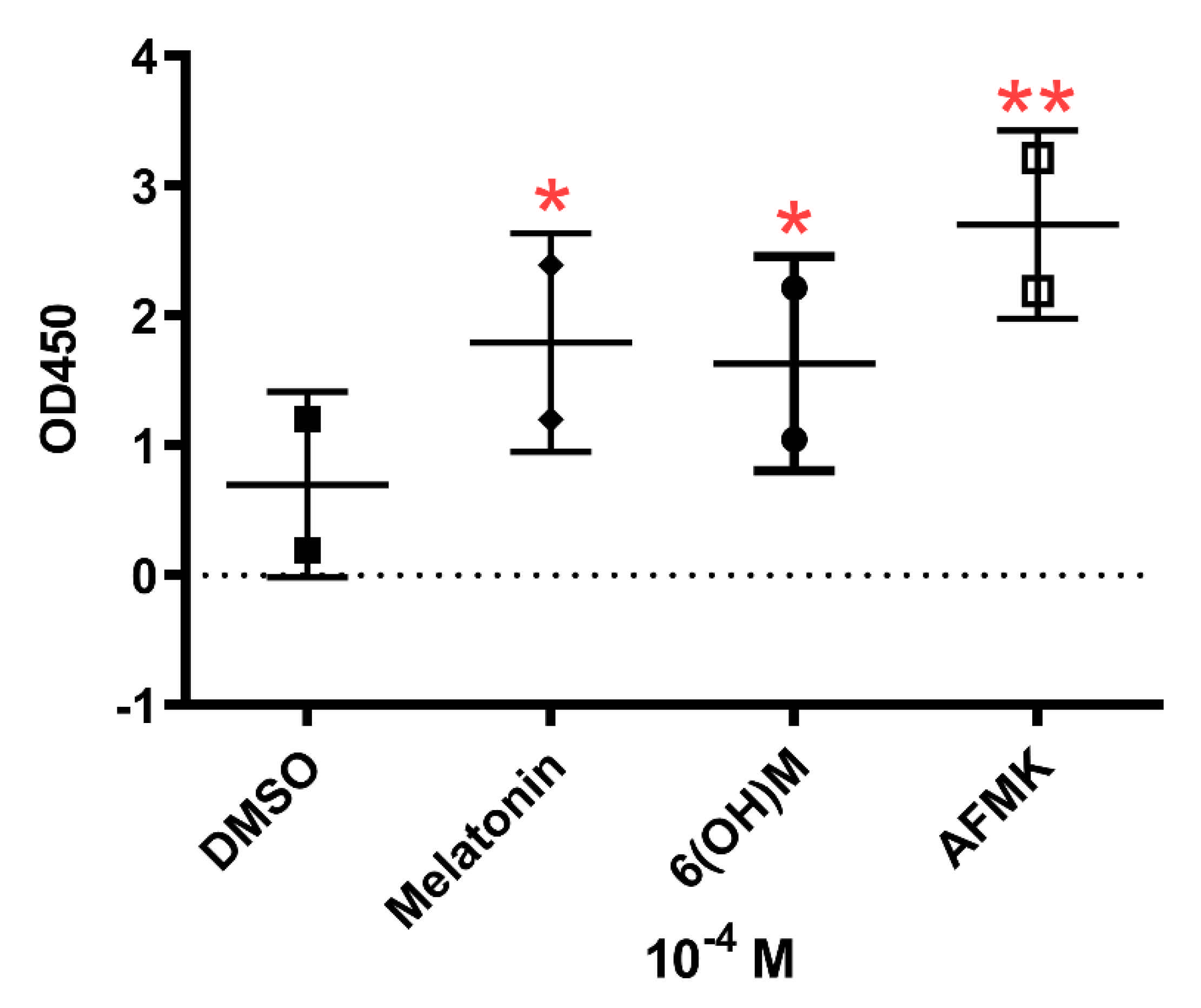
3.3. Cytokine Array Analysis
3.4. Results of Telomerase Assay
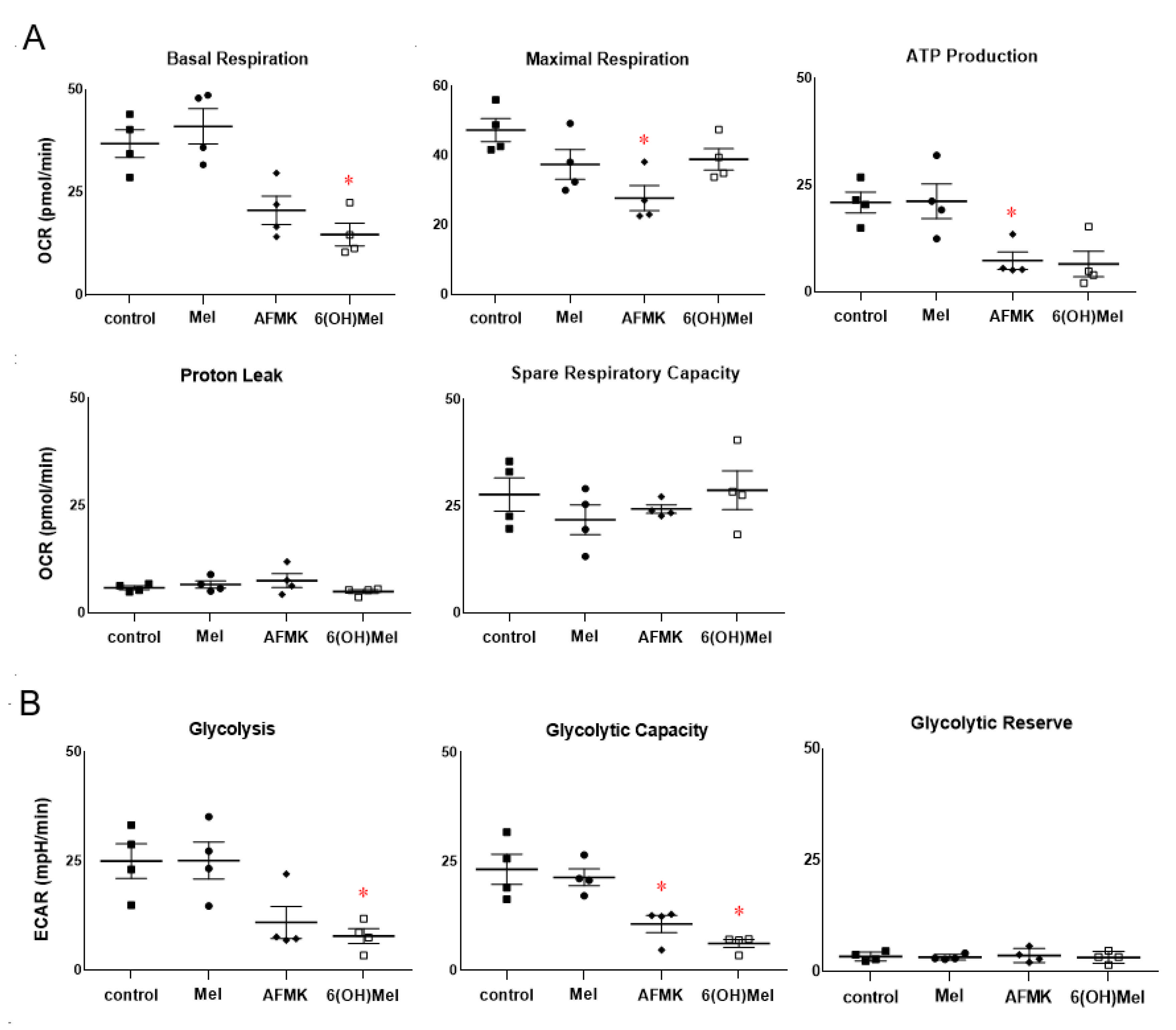
3.5. Energy Yielding Metabolism Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BRCA1 | breast cancer gene 1 |
| DMSO | dimethyl sulfoxide |
| NRF2 | NF-E2–related factor 2 |
| p53 | tumor protein P53 |
References
- Reiter, R.J.; Rosales-Corral, S.A.; Tan, D.X.; Acuna-Castroviejo, D.; Qin, L.; Yang, S.F.; Xu, K. Melatonin, a full service anti-cancer agent: Inhibition of initiation, progression and metastasis. Int. J. Mol. Sci. 2017, 18, 843. [Google Scholar] [CrossRef]
- Tan, D.X.; Reiter, R.J. Mitochondria: The birth place, battle ground and the site of melatonin metabolism in cells. Melatonin Res. 2019, 2, 44–66. [Google Scholar] [CrossRef]
- Cipolla-Neto, J.; Gaspar do Amaral, F. Melatonin as a hormone: New physiological and clinical insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef] [Green Version]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell. Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Tan, D.X.; Rosales-Corral, S.; Manchester, L.C. The universal nature, unequal distribution and antioxidant functions of melatonin and its derivatives. Mini Rev. Med. Chem. 2013, 13, 373–384. [Google Scholar]
- Slominski, A.; Tobin, D.J.; Zmijewski, M.A.; Wortsman, J.; Paus, R. Melatonin in the skin: Synthesis, metabolism and functions. Trends Endocrinol. Metab. 2008, 19, 17–24. [Google Scholar] [CrossRef]
- Fischer, T.W.; Zmijewski, M.A.; Wortsman, J.; Slominski, A. Melatonin maintains mitochondrial membrane potential and attenuates activation of initiator (casp-9) and effector caspases (casp-3/casp-7) and PARP in UVR-exposed HaCaT keratinocytes. J. Pineal Res. 2008, 44, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Jung, B.; Ahmad, N. Melatonin in cancer management: Progress and promise. Cancer Res. 2006, 66, 9789–9793. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Rosales-Corral, S.; Tan, D.X.; Jou, M.J.; Galano, A.; Xu, B. Melatonin as a mitochondria-targeted antioxidant: One of evolution’s best ideas. Cell. Mol. Life Sci. 2017, 74, 3863–3881. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kleszczyński, K.; Semak, I.; Janjetovic, Z.; Zmijewski, M.A.; Kim, T.K.; Slominski, R.M.; Reiter, R.J.; Fischer, T.W. Local melatoninergic system as the protector of skin integrity. Int. J. Mol. Sci. 2014, 15, 17705–17732. [Google Scholar] [CrossRef] [Green Version]
- Fischer, T.W.; Sweatman, T.W.; Semak, I.; Sayre, R.M.; Wortsman, J.; Slominski, A. Constitutive and UV-induced metabolism of melatonin in keratinocytes and cell-free systems. FASEB J. 2006, 20, 1564–1566. [Google Scholar] [CrossRef] [Green Version]
- Janjetovic, Z.; Stuart, G.; Jarrett, S.G.; Lee, E.F.; Duprey, C.; Reiter, R.J.; Slominski, A.T. Melatonin and its metabolites protect human melanocytes against UVB-induced damage: Involvement of NRF2-mediated pathways. Sci. Rep. 2017, 7, 1274. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.K.; Kleszczyński, K.; Janjetovic, Z.; Sweatman, T.; Lin, Z.; Li, W.; Reiter, R.J.; Fischer, T.W.; Slominski, A.T. Metabolism of melatonin and biological activity of intermediates of melatoninergic pathway in human skin cells. FASEB J. 2013, 27, 2742–2755. [Google Scholar] [CrossRef]
- Semak, I.; Naumova, M.; Korik, E.; Terekhovich, V.; Wortsman, J.; Slominski, A. A novel metabolic pathway of melatonin: Oxidation by cytochrome C. Biochemistry 2005, 44, 9300–9307. [Google Scholar] [CrossRef]
- Semak, I.; Korik, E.; Antonova, M.; Wortsman, J.; Slominski, A. Metabolism of melatonin by cytochrome P450s in rat liver mitochondria and microsomes. J. Pineal Res. 2008, 45, 515–523. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.T.; Semak, I.; Fischer, T.W.; Kim, T.K.; Kleszczyński, K.; Hardeland, R.; Reiter, R.J. Metabolism of melatonin in the skin: Why is it important? Exp. Dermatol. 2017, 26, 563–568. [Google Scholar] [CrossRef]
- Fischer, T.W.; Zbytek, B.; Sayre, R.M.; Apostolov, E.O.; Basnakian, A.G.; Sweatman, T.W.; Wortsman, J.; Elsner, P.; Slominski, A. Melatonin increases survival of HaCaT keratinocytes by suppressing UV-induced apoptosis. J. Pineal Res. 2006, 40, 18–26. [Google Scholar] [CrossRef]
- Slominski, A.; Baker, J.; Rosano, T.G.; Guisti, L.W.; Ermak, G.; Grande, M.; Gaudet, S.J. Metabolism of serotonin to N-acetylserotonin, melatonin, and 5-methoxytryptamine in hamster skin culture. J. Biol. Chem. 1996, 271, 12281–12286. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.T.; Kim, T.K.; Kleszczyński, K.; Semak, I.; Janjetovic, Z.; Sweatman, T.; Skobowiat, C.; Steketee, J.D.; Lin, Z.; Postlethwaite, A.; et al. Characterization of serotonin and N-acetylserotonin systems in the human epidermis and skin cells. J. Pineal Res. 2020, 68, e12626. [Google Scholar] [CrossRef]
- Slominski, A.; Pisarchik, A.; Semak, I.; Sweatman, T.; Wortsman, J.; Szczesniewski, A.; Slugocki, G.; McNulty, J.; Kauser, S.; Tobin, D.J.; et al. Serotoninergic and melatoninergic systems are fully expressed in human skin. FASEB J. 2002, 16, 896–898. [Google Scholar] [CrossRef] [Green Version]
- Slominski, A.; Wortsman, J.; Tobin, D.J. The cutaneous serotoninergic/melatoninergic system: Securing a place under the sun. FASEB J. 2005, 19, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Kim, T.K.; Janjetovic, Z.; Slominski, R.; Zmijewski, J.W. Melatonin, mitochondria, and the skin. Cell. Mol. Life. Sci. 2017, 74, 3913–3925. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Plonka, P.M.; Szaflarski, J.P.; Paus, R. How UV light touches the brain and endocrine system through skin, and why. Endocrinology 2018, 159, 1992–2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.; Wortsman, J. Neuroendocrinology of the skin. Endocr. Rev. 2000, 21, 457–487. [Google Scholar] [CrossRef]
- Fischer, T.W.; Kleszczyński, K.; Hardkop, L.H.; Kruse, N.; Zillikens, D. Melatonin enhances antioxidative enzyme gene expression (CAT, GPx, SOD), prevents their UVR-induced depletion, and protects against the formation of DNA damage (8-hydroxy-2’-deoxyguanosine) in ex vivo human skin. J. Pineal Res. 2013, 54, 303–312. [Google Scholar] [CrossRef]
- Fischer, T.W.; Slominski, A.; Zmijewski, M.A.; Reiter, R.J.; Paus, R. Melatonin as a major skin protectant: From free radical scavenging to DNA damage repair. Exp. Dermatol. 2008, 17, 713–730. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Zillikens, D.; Fischer, T.W. Melatonin enhances mitochondrial ATP synthesis, reduces reactive oxygen species formation, and mediates translocation of the nuclear erythroid 2-related factor 2 resulting in activation of phase-2 antioxidant enzymes (γ-GCS, HO-1, NQO1) in ultraviolet radiation-treated normal human epidermal keratinocytes (NHEK). J. Pineal Res. 2016, 61, 187–197. [Google Scholar]
- Skobowiat, C.; Brożyna, A.A.; Janjetovic, Z.; Jeayeng, S.; Oak, A.S.W.; Kim, T.K.; Panich, U.; Reiter, R.J.; Slominski, A.T. Melatonin and its derivatives counteract the ultraviolet B radiation-induced damage in human and porcine skin ex vivo. J. Pineal Res. 2018, 65, e12501. [Google Scholar] [CrossRef]
- Kim, T.K.; Lin, Z.; Li, W.; Reiter, R.J.; Slominski, A.T. N1-Acetyl-5-Methoxykynuramine (AMK) is produced in the human epidermis and shows antiproliferative effects. Endocrinology 2015, 156, 1630–1636. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.K.; Lin, Z.; Tidwell, W.J.; Li, W.; Slominski, A.T. Melatonin and its metabolites accumulate in the human epidermis in vivo and inhibit proliferation and tyrosinase activity in epidermal melanocytes in vitro. Mol. Cell. Endocrinol. 2015, 404, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Milani, M.; Sparavigna, A. Antiaging efficacy of melatonin-based day and night creams: A randomized, split-face, assessor-blinded proof-of-concept trial. Clin. Cosmet. Investig. Dermatol. 2018, 11, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Scheuer, C.; Pommergaard, H.C.; Rosenberg, J.; Gögenur, I. Dose dependent sun protective effect of topical melatonin: A randomized, placebo-controlled, double-blind study. J. Dermatol. Sci. 2016, 84, 178–185. [Google Scholar] [CrossRef]
- Ren, W.; Liu, G.; Chen, S.; Yin, J.; Wang, J.; Tan, B.; Wu, G.; Bazer, F.W.; Peng, Y.; Li, T.; et al. Melatonin signaling in T cells: Functions and applications. J. Pineal Res. 2017, 62, e12394. [Google Scholar] [CrossRef] [Green Version]
- Tamtaji, O.R.; Mobini, M.; Reiter, R.J.; Azami, A.; Gholami, M.S.; Asemi, Z. Melatonin, a toll-like receptor inhibitor: Current status and future perspectives. J. Cell. Physiol. 2019, 234, 7788–7795. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, S.; Zeng, S.; Zhao, Y.; Zhu, C.; Deng, B.; Zhu, G.; Yin, Y.; Wang, W.; Hardeland, R.; et al. Melatonin in macrophage biology: Current understanding and future perspectives. J. Pineal Res. 2019, 66, e12547. [Google Scholar] [CrossRef] [Green Version]
- Kleszczyński, K.; Slominski, A.T.; Steinbrink, K.; Reiter, R.J. Clinical trials for use of melatonin to fight against COVID-19 are urgently needed. Nutrients 2020, 12, 2561. [Google Scholar] [CrossRef]
- Reiter, R.J.; Ma, A.; Sharma, R. Treatment of ebola and other infectious diseases: Melatonin “goes viral”. Melatonin Res. 2020, 3, 43–57. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R. Potential utility of melatonin in deadly infectious diseases related to the overreaction of innate immune response and destructive inflammation: Focus on COVID-19. Melatonin Res. 2020, 3, 120–143. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin and inflammation-Story of a double-edged blade. J. Pineal Res. 2018, 65, e12525. [Google Scholar] [CrossRef] [Green Version]
- Hardeland, R. Aging, melatonin, and the pro- and anti-inflammatory networks. Int. J. Mol. Sci. 2019, 20, 1223. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, I.J.; Huang, C.C.; Liu, S.C.; Tang, C.H. Reconsidering the role of melatonin in rheumatoid arthritis. Int. J. Mol. Sci. 2020, 21, 2877. [Google Scholar] [CrossRef] [PubMed]
- Kleszczyński, K.; Bilska, B.; Stegemann, A.; Flis, D.J.; Ziolkowski, W.; Pyza, E.; Luger, T.A.; Reiter, R.J.; Böhm, M.; Slominski, A.T. Melatonin and its metabolites ameliorate UVR-induced mitochondrial oxidative stress in human MNT-1 melanoma cells. Int. J. Mol. Sci. 2018, 19, 3786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bocheva, G.; Slominski, R.M.; Slominski, A.T. Neuroendocrine aspects of skin aging. Int. J. Mol. Sci. 2019, 20, 2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janjetovic, Z.; Nahmias, Z.P.; Hanna, S.; Jarrett, S.G.; Kim, T.K.; Reiter, R.J.; Slominski, A.T. Melatonin and its metabolites ameliorate ultraviolet B-induced damage in human epidermal keratinocytes. J. Pineal Res. 2014, 57, 90–102. [Google Scholar] [CrossRef]
- Lin, S.; Hoffmann, K.; Gao, C.; Petrulionis, M.; Herr, I.; Schemmer, P. Melatonin promotes sorafenib-induced apoptosis through synergistic activation of JNK/c-jun pathway in human hepatocellular carcinoma. J. Pineal Res. 2017, 62, e12398. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Fischer, T.W. Melatonin and human skin aging. Dermatoendocrinology 2012, 4, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Kleszczyński, K.; Tukaj, S.; Kruse, N.; Zillikens, D.; Fischer, T.W. Melatonin prevents ultraviolet radiation-induced alterations in plasma membrane potential and intracellular pH in human keratinocytes. J. Pineal Res. 2013, 54, 89–99. [Google Scholar] [CrossRef]
- Le, P.H.; Kuo, C.J.; Cheng, H.T.; Wu, R.C.; Chen, T.H.; Lin, C.J.; Chiang, K.C.; Hsu, J.T. Melatonin attenuates acute pancreatitis-induced liver damage through Akt-dependent PPAR-γ pathway. J. Surg. Res. 2019, 236, 311–318. [Google Scholar] [CrossRef]
- Matsui, M.S.; Pelle, E.; Dong, K.; Pernodet, N. Biological rhythms in the skin. Int. J. Mol. Sci. 2016, 17, 801. [Google Scholar] [CrossRef]
- Pappas, A. Epidermal surface lipids. Dermatoendocrinology 2009, 1, 72–76. [Google Scholar] [CrossRef] [Green Version]
- Zouboulis, C.C. Acne and sebaceous gland function. Clin. Dermatol. 2004, 22, 360–366. [Google Scholar] [CrossRef]
- Rosenstein, R.E.; Cardinali, D.P. Central gabaergic mechanisms as targets for melatonin activity in brain. Neurochem. Int. 1990, 17, 373–379. [Google Scholar] [CrossRef]
- Guo, J.Y.; Li, F.; Wen, Y.B.; Cui, H.X.; Guo, M.L.; Zhang, L.; Zhang, Y.F.; Guo, Y.J.; Guo, Y.X. Melatonin inhibits Sirt1-dependent NAMPT and NFAT5 signaling in chondrocytes to attenuate osteoarthritis. Oncotarget 2017, 8, 55967–55983. [Google Scholar] [CrossRef] [Green Version]
- Hosseinzadeh, A.; Kamrava, S.K.; Joghataei, M.T.; Darabi, R.; Shakeri-Zadeh, A.; Shahriari, M.; Reiter, R.J.; Ghaznavi, H.; Mehrzadi, S. Apoptosis signaling pathways in osteoarthritis and possible protective role of melatonin. J. Pineal Res. 2016, 61, 411–425. [Google Scholar] [CrossRef]
- Lim, H.D.; Kim, Y.S.; Ko, S.H.; Yoon, I.J.; Cho, S.G.; Chun, Y.H.; Choi, B.J.; Kim, E.C. Cytoprotective and anti-inflammatory effects of melatonin in hydrogen peroxide-stimulated CHON-001 human chondrocyte cell line and rabbit model of osteoarthritis via the SIRT1 pathway. J. Pineal Res. 2012, 53, 225–237. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, J.; Zhou, X.; Chen, X.; Chen, A.C.; Pi, B.; Pan, G.; Pei, M.; Yang, H.; Liu, T.; et al. Melatonin prevents osteoarthritis-induced cartilage degradation via targeting microRNA-140. Oxid. Med. Cell. Longev. 2019, 2019, 9705929. [Google Scholar] [CrossRef] [Green Version]
- Ansel, J.; Perry, P.; Brown, J.; Damm, D.; Phan, T.; Hart, C.; Luger, T.; Hefeneider, S. Cytokine modulation of keratinocyte cytokines. J. Investig. Dermatol. 1990, 94, 101S–107S. [Google Scholar] [CrossRef] [Green Version]
- Kupper, T.S. Interleukin 1 and other human keratinocyte cytokines: Molecular and functional characterization. Adv. Dermatol. 1988, 3, 293–307. [Google Scholar]
- Kupper, T.S.; Chua, A.O.; Flood, P.; McGuire, J.; Gubler, U. Interleukin 1 gene expression in cultured human keratinocytes is augmented by ultraviolet irradiation. J. Clin. Investig. 1987, 80, 430–436. [Google Scholar] [CrossRef]
- Gabay, C.; Lamacchia, C.; Palmer, G. IL-1 pathways in inflammation and human diseases. Nat. Rev. Rheumatol. 2010, 6, 232–241. [Google Scholar] [CrossRef]
- Providence, K.M.; Higgins, S.P.; Mullen, A.; Battista, A.; Samarakoon, R.; Higgins, C.E.; Wilkins-Port, C.E.; Higgins, P.J. SERPINE1 (PAI-1) is deposited into keratinocyte migration “trails” and required for optimal monolayer wound repair. Arch. Dermatol. Res. 2008, 300, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Gnanadurai, C.W.; Fu, Z. Critical roles of chemokines and cytokines in antiviral innate immune responses during rabies virus infection. Front. Agr. Sci. Eng. 2017, 4, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Juybari, K.B.; Pourhanifeh, M.H.; Hosseinzadeh, A.; Hemati, K.; Mehrzadi, S. Melatonin potentials against viral infections including COVID-19: Current evidence and new findings. Virus Res. 2020, 287, 198108. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Sharma, R.; Ma, Q. Switching diseased cells from cytosolic aerobic glycolysis to mitochondrial oxidative phosphorylation: A metabolic rhythm regulated by melatonin? J. Pineal Res. 2021, 70, e12677. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Top Diseases and Biofunctions | Melatonin | AFMK | 6(OH)Mel | |||
|---|---|---|---|---|---|---|
| p value range | Molecules | p value range | Molecules | p value range | Molecules |
| Cancer | 1.39 × 10−3 ÷ 4.30 × 10−21 | 1313 | 3.11 × 10−3 ÷ 2.06 × 10−23 | 1419 | 5.59 × 10−3 ÷ 8.62 × 10−21 | 1211 |
| Dermatological diseases and conditions | 2.99 × 10−4 ÷ 4.30 × 10−21 | 1073 | 1.60 × 10−3 ÷ 2.06 × 10−23 | 1127 | 5.22 × 10−3 ÷ 8.62 × 10−21 | 1065 |
| Organismal injury and abnormalities | 1.48 × 10−3 ÷ 4.30 × 10−21 | 1454 | 3.14 × 10−3 ÷ 2.06 × 10−23 | 1528 | 5.94 × 10−3 ÷ 8.62 × 10−21 | 1330 |
| Reproductive system disease | 1.48 × 10−3 ÷ 3.49 × 10−13 | 104 | 1.13 × 10−10 | 19 | 5.78 × 10−3 ÷ 6.37 × 10−9 | 47 |
| Endocrine system disorders | 1.47 × 10−3 ÷ 9.57 × 10−13 | 598 | 3.14 × 10−03 ÷ 1.56 × 10−12 | 252 | 6.1 × 10−8 | 194 |
| Gastrointestinal disease | 6.37 × 10−8 | 26 | 3.11 × 10−3 ÷ 1.56 × 10−12 | 404 | 1.6 × 10−7 | 86 |
| Psychological disorders | 1.7 × 10−7 | 18 | 1.35 × 10−5 | 31 | 5.93 × 10−3 ÷ 4.25 × 10−8 | 170 |
| p value range | Molecules | p value range | Molecules | p value range | Molecules |
| Cell to cell signaling and interaction | 1.47 × 10−3 ÷ 2.19 × 10−9 | 415 | 1.2 × 10−5 | 86 | 5.96 × 10−3 ÷ 1.71 × 10−9 | 416 |
| Gene expression | 5.80 × 10−8 ÷ 5.80 × 10−8 | 25 | 1.96 × 10−6 | 23 | 5.15 × 10−5 | 20 |
| Cell signaling | 1.39 × 10−3 ÷ 7.55 × 10−7 | 172 | 3.11 × 10−3 ÷ 2.71 × 10−7 | 191 | 5.96 × 10−3 ÷ 1.23 × 10−8 | 205 |
| Molecular transport | 1.43 × 10−3 ÷ 7.55 × 10−7 | 267 | 3.12 × 10x−3 ÷ 7.72 × 10−10 | 375 | 5.96 × 10−3 ÷ 3.08 × 10−10 | 349 |
| Vitamin and mineral metabolism | 7.29 × 10−4 ÷ 7.55 × 10−7 | 138 | 2.43 × 10−3 ÷ 2.71 × 10−7 | 150 | 5.96 × 10−3 ÷ 1.23 × 10−8 | 169 |
| Cellular function and maintenance | 1.49 × 10−6 | 69 | 3.12 × 10−3 ÷ 1.06 × 10−9 | 157 | 2.51 × 10−7 | 18 |
| Cellular movement | 3.95 × 10−6 | 37 | 2.95 × 10−3 ÷ 4.56 × 10−7 | 208 | 7.3 × 10−6 | 62 |
| Cell morphology | 9.84 × 10−4 | 7 | 5.36 × 10−5 | 15 | 5.34 × 10−3 ÷ 2.51 × 10−7 | 97 |
| Reactome Pathway | GSEA for Melatonin | GSEA for AFMK | GSEA for 6(OH)Mel | |||
|---|---|---|---|---|---|---|
| NES | Direction | NES | Direction | NES | Direction | |
| Detoxification of reactive oxygen species | 1.09 | ↑ | 1.10 | ↑ | 1.10 | ↑ |
| DNA double-strand break response | 1.03 | ↑ | 1.04 | ↑ | 1.03 | ↑ |
| Base excision repair | 0.99 | ↑ | × | × | 0.99 | ↑ |
| p53-dependent G1/S DNA damage checkpoint | 1.11 | ↑ | 1.09 | ↑ | × | × |
| DNA double-strand break repair | 0.99 | ↑ | 1.00 | ↑ | 0.99 | ↑ |
| p53-independent G1/S DNA damage checkpoint | 1.10 | ↑ | 1.12 | ↑ | 1.11 | ↑ |
| TP53 regulates transcription of DNA repair genes | 1.01 | ↑ | 1.002 | ↑ | 0.99 | ↑ |
| Regulation of TP53 activity through methylation | 1.07 | ↑ | 1.10 | ↑ | 1.08 | ↑ |
| Regulation of TP53 activity through phosphorylation | 1.00 | ↑ | × | × | × | × |
| Recognition of DNA damage by PCNA-containing replication complex | 1.05 | ↑ | × | × | 1.06 | ↑ |
| Dual incision in GG-NER | 1.05 | ↑ | × | × | × | × |
| Gap-filling DNA repair synthesis and ligation in GG-NER | 1.05 | ↑ | × | × | 1.05 | ↑ |
| PCNA-dependent long patch base excision repair | 1.03 | ↑ | × | × | × | × |
| DNA damage recognition in GG-NER | × | × | 1.08 | ↑ | 1.07 | ↑ |
| Gap-filling DNA repair synthesis and ligation in TC-NER | × | × | 1.04 | ↑ | 1.04 | ↑ |
| G2/M DNA damage checkpoints | × | × | 1.02 | ↑ | × | × |
| G1/S DNA damage checkpoints | × | × | 1.09 | ↑ | 1.10 | ↑ |
| Stabilization of p53 | × | × | 1.12 | ↑ | × | × |
| DNA damage bypass | × | × | 1.05 | ↑ | × | × |
| Nucleotide excision repair | × | × | × | × | 1.04 | ↑ |
| Influence of Melatonin Metabolites on Mitochondrial Metabolism | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Reactome Pathway | GSEA for Melatonin | GSEA for 6(OH)Mel | GSEA for AFMK | ||||||
| NES | FDR | Direction | NES | FDR | Direction | NES | FDR | Direction | |
| Citric acid cycle | 1.06 | 0.50 | ↑ | 1.07 | 0.49 | ↑ | 1.06 | 0.52 | ↑ |
| Glucose metabolism | 1.06 | 0.50 | ↑ | 1.05 | 0.51 | ↑ | 1.05 | 0.51 | ↑ |
| Mitochondrial translation | 1.07 | 0.50 | ↑ | 1.07 | 0.49 | ↑ | 1.07 | 0.50 | ↑ |
| Cristae formation | 1.07 | 0.50 | ↑ | 1.07 | 0.49 | ↑ | 1.09 | 0.54 | ↑ |
| Pyruvate metabolism and citric acid cycle | 1.05 | 0.50 | ↑ | 1.05 | 0.51 | ↑ | 1.05 | 0.51 | ↑ |
| Mitochondrial translation elongation | 1.08 | 0.51 | ↑ | 1.09 | 0.57 | ↑ | 1.09 | 0.53 | ↑ |
| Pyruvate metabolism | 1.05 | 0.52 | ↑ | 1.03 | 0.53 | ↑ | 1.06 | 0.50 | ↑ |
| Mitochondrial protein import | 1.04 | 0.53 | ↑ | 1.05 | 0.51 | ↑ | 1.03 | 0.54 | ↑ |
| Mitochondrial biogenesis | 1.04 | 0.53 | ↑ | 1.05 | 0.51 | ↑ | 1.03 | 0.54 | ↑ |
| Mitochondrial translation initiation | 1.09 | 0.54 | ↑ | 1.07 | 0.48 | ↑ | 1.08 | 0.51 | ↑ |
| Mitochondrial fatty acid β-oxidation | 1.03 | 0.56 | ↑ | × | × | × | × | × | × |
| The citric acid (TCA) cycle and respiratory electron transport | 1.09 | 0.57 | ↑ | 1.09 | 0.56 | ↑ | 1.08 | 0.52 | ↑ |
| Gluconeogenesis | 1.12 | 0.85 | ↑ | 1.13 | 0.77 | ↑ | 1.12 | 0.86 | ↑ |
| Glycogen metabolism | 1.11 | 0.76 | ↑ | 1.11 | 0.72 | ↑ | 1.11 | 0.79 | ↑ |
| Mitochondrial tRNA aminoacylation | 1.00 | 0.61 | ↑ | 1.00 | 0.59 | ↑ | 1.00 | 0.60 | ↑ |
| Respiratory electron transport | 1.10 | × | × | 1.10 | 0.63 | ↑ | 1.09 | 0.57 | ↑ |
| Respiratory electron transport, ATP synthesis by chemiosmotic coupling, and heat production by uncoupling proteins | × | × | × | 1.10 | 0.68 | ↑ | 1.11 | 0.78 | ↑ |
| Glycogen synthesis | 1.10 | 0.67 | ↑ | × | × | × | 1.09 | 0.57 | ↑ |
| Therapeutic Effects of Melatonin and Its Metabolites | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Reactome Pathway | GSEA for Melatonin | GSEA for 6(OH)Mel | GSEA for AFMK | ||||||
| NES | FDR | Direction | NES | FDR | Direction | NES | FDR | Direction | |
| Antiviral mechanism by IFN-stimulated genes | × | × | × | 1.03 | 0.57 | ↑ | 1.03 | 0.55 | ↑ |
| ISG15 antiviral mechanism | 1.03 | 0.55 | ↑ | × | × | × | 1.03 | 0.55 | ↑ |
| Defensins | 0.94 | 0.89 | ↑ | 0.94 | 0.90 | ↑ | 0.94 | 0.91 | ↑ |
| Beta defensins | 0.93 | 0.93 | ↑ | 0.92 | 0.95 | ↑ | 0.91 | 0.96 | ↑ |
| Tat-mediated HIV elongation arrest and recovery | 1.06 | 0.50 | ↑ | 1.05 | 0.51 | ↑ | 1.07 | 0.49 | ↑ |
| HIV elongation arrest and recovery | 1.07 | 0.51 | ↑ | 1.06 | 0.50 | ↑ | 1.08 | 0.49 | ↑ |
| Abortive elongation of HIV-1 transcript in the absence of Tat | 1.06 | 0.49 | ↑ | 1.06 | 0.50 | ↑ | × | × | × |
| Pausing and recovery of HIV elongation | 1.05 | 0.49 | ↑ | 1.06 | 0.51 | ↑ | × | × | × |
| Telomere maintenance | 0.98 | 0.69 | ↑ | 0.99 | 0.66 | ↑ | 0.99 | 0.64 | ↑ |
| Packaging of telomere ends | 1.01 | 0.60 | ↑ | 1.02 | 0.58 | ↑ | 1.03 | 0.55 | ↑ |
| Extension of telomeres | 0.96 | 0.83 | ↑ | 0.97 | 0.75 | ↑ | 0.97 | 0.76 | ↑ |
| Telomere C-strand (Lagging Strand) synthesis | 0.97 | 0.74 | ↑ | 0.96 | 0.83 | ↑ | 0.95 | 0.86 | ↑ |
| Interleukin-10 signaling | 1.06 | 0.49 | ↑ | 1.06 | 0.50 | ↑ | 1.07 | 0.49 | ↑ |
| Reactome Pathway | GSEA for Melatonin | GSEA for 6(OH)Mel | GSEA for AFMK | |||
|---|---|---|---|---|---|---|
| NES | Direction | NES | Direction | NES | Direction | |
| Plasma lipoprotein assembly, remodeling, and clearance | 1.08 | ↑ | 1.08 | ↑ | 1.08 | ↑ |
| ABC-family proteins-mediated transport | 1.08 | ↑ | 1.08 | ↑ | × | × |
| Triglyceride catabolism | 1.04 | ↑ | × | × | × | × |
| LDL clearance | 1.09 | ↑ | 1.09 | ↑ | 1.08 | ↑ |
| PPARα activated gene expression | 1.01 | ↑ | × | × | 1.00 | ↑ |
| Assembly of active LPL and LIPC lipase complexes | 1.00 | ↑ | × | × | 1.01 | ↑ |
| Platelet homeostasis | 1.00 | ↑ | 1.00 | ↑ | × | × |
| Plasma lipoprotein clearance | × | × | × | × | 1.07 | ↑ |
| Prostacyclin signaling through prostacyclin receptor | × | × | 1.10 | ↑ | 1.11 | ↑ |
| Platelet calcium homeostasis | × | × | × | × | 0.98 | ↑ |
| Regulation of lipid metabolism by PPARα | × | × | × | × | 0.97 | ↑ |
| Nitric oxide stimulated guanylate cyclase | × | × | × | × | 0.86 | ↑ |
| Vasopressin regulates renal water homeostasis via aquaporins | × | × | 1.04 | ↑ | × | × |
| Regulation of insulin secretion | × | × | 1.04 | ↑ | × | × |
| Triglyceride catabolism | × | × | 1.03 | ↑ | × | × |
| ABC transporters in lipid homeostasis | × | × | 0.89 | ↑ | × | × |
| IPAs | IPA for Melatonin | IPA for AFMK | IPA for 6(OH)Mel | |||
|---|---|---|---|---|---|---|
| Z-Score | Ratio | Z-Score | Ratio | Z-Score | Ratio | |
| PD-1, PD-L1 cancer immunotherapy pathway | 0.33 | 0.11 | 1.51 | 0.15 | × | × |
| Renal cell carcinoma signaling pathway | −2 | 0.06 | −1.34 | 0.07 | × | × |
| Basal cell carcinoma signaling | × | 0.08 | × | × | −2.24 | 0.14 |
| Glioma signaling | −1.63 | 0.06 | −0.70 | 0.08 | × | × |
| Acute myeloid leukemia signaling | −1.34 | 0.08 | −1.41 | 0.11 | −1.34 | 0.08 |
| SPINK1 general cancer pathway | × | × | −0.70 | 0.15 | × | × |
| Estrogen-dependent breast cancer signaling | × | × | −1 | 0.12 | × | × |
| SPINK1 pancreatic cancer pathway | × | × | −0.44 | 0.12 | × | × |
| Small cell lung cancer signaling | × | × | −1 | 0.07 | × | × |
| Endometrial cancer signaling | × | × | −1 | 0.08 | × | × |
| Non-small cell Lung cancer signaling | × | × | −1.34 | 0.09 | × | × |
| Pancreatic adenocarcinoma signaling | × | × | −1.63 | 0.05 | × | × |
| Glioma Invasiveness signaling | × | × | −1.13 | 0.12 | × | × |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefan, J.; Kim, T.-K.; Schedel, F.; Janjetovic, Z.; Crossman, D.K.; Steinbrink, K.; Slominski, R.M.; Zmijewski, J.; Tulic, M.K.; Reiter, R.J.; et al. Differential and Overlapping Effects of Melatonin and Its Metabolites on Keratinocyte Function: Bioinformatics and Metabolic Analyses. Antioxidants 2021, 10, 618. https://doi.org/10.3390/antiox10040618
Stefan J, Kim T-K, Schedel F, Janjetovic Z, Crossman DK, Steinbrink K, Slominski RM, Zmijewski J, Tulic MK, Reiter RJ, et al. Differential and Overlapping Effects of Melatonin and Its Metabolites on Keratinocyte Function: Bioinformatics and Metabolic Analyses. Antioxidants. 2021; 10(4):618. https://doi.org/10.3390/antiox10040618
Chicago/Turabian StyleStefan, Joanna, Tae-Kang Kim, Fiona Schedel, Zorica Janjetovic, David K. Crossman, Kerstin Steinbrink, Radomir M. Slominski, Jaroslaw Zmijewski, Meri K. Tulic, Russel J. Reiter, and et al. 2021. "Differential and Overlapping Effects of Melatonin and Its Metabolites on Keratinocyte Function: Bioinformatics and Metabolic Analyses" Antioxidants 10, no. 4: 618. https://doi.org/10.3390/antiox10040618