
Protective Effects of Lignin-Carbohydrate Complexes from Wheat Stalk against Bisphenol a Neurotoxicity in Zebrafish via Oxidative Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Analysis and Characterization of LCC Preparations
2.3. Zebrafish Husbandry
2.4. Effects of LCCs on the Hatching Rate of Zebrafish
2.5. Effects of LCCs on Zebrafish Behavior
2.6. Antioxidant Properties and Effects of LCCs on Oxidative Stress in Zebrafish
2.7. Neuroprotective Effects of LCCs in Zebrafish
2.8. Real-Time PCR
2.9. Statistical Analysis
3. Results and Discussion
3.1. Composition and Molecular Weight of the LCC Preparations
3.2. Structural Properties of the LCC Preparations
3.3. Effects of LCCs on the Survival Rate and Hatching Rate of Zebrafish
3.4. Effects of LCC-WS-A and LCC-WS-B on the Behavior of BPA-Treated Zebrafish
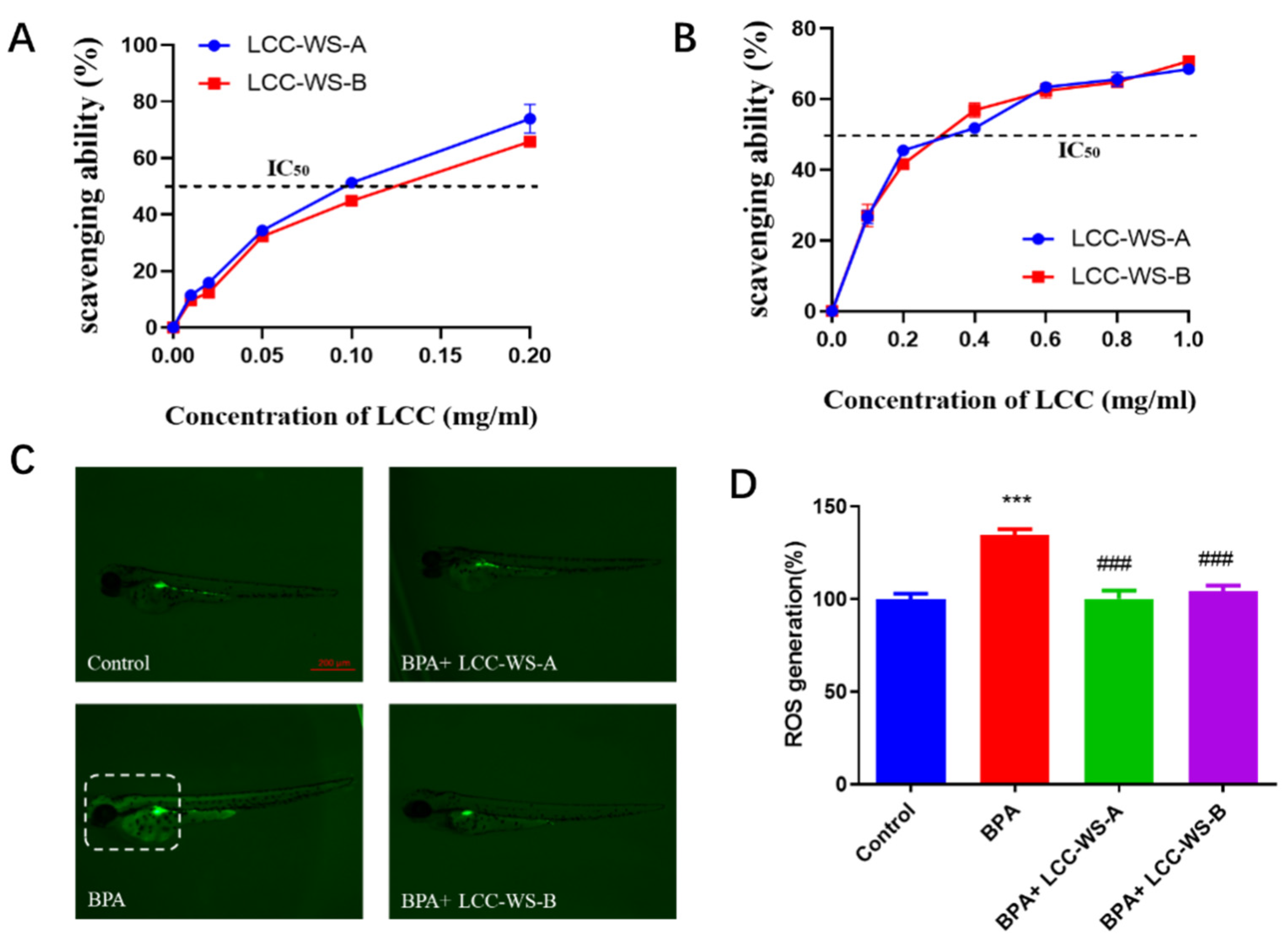
3.5. In Vitro and In Vivo Antioxidant Properties of LCCs and their Protective Effects against BPA-Induced Oxidative Damage in Zebrafish
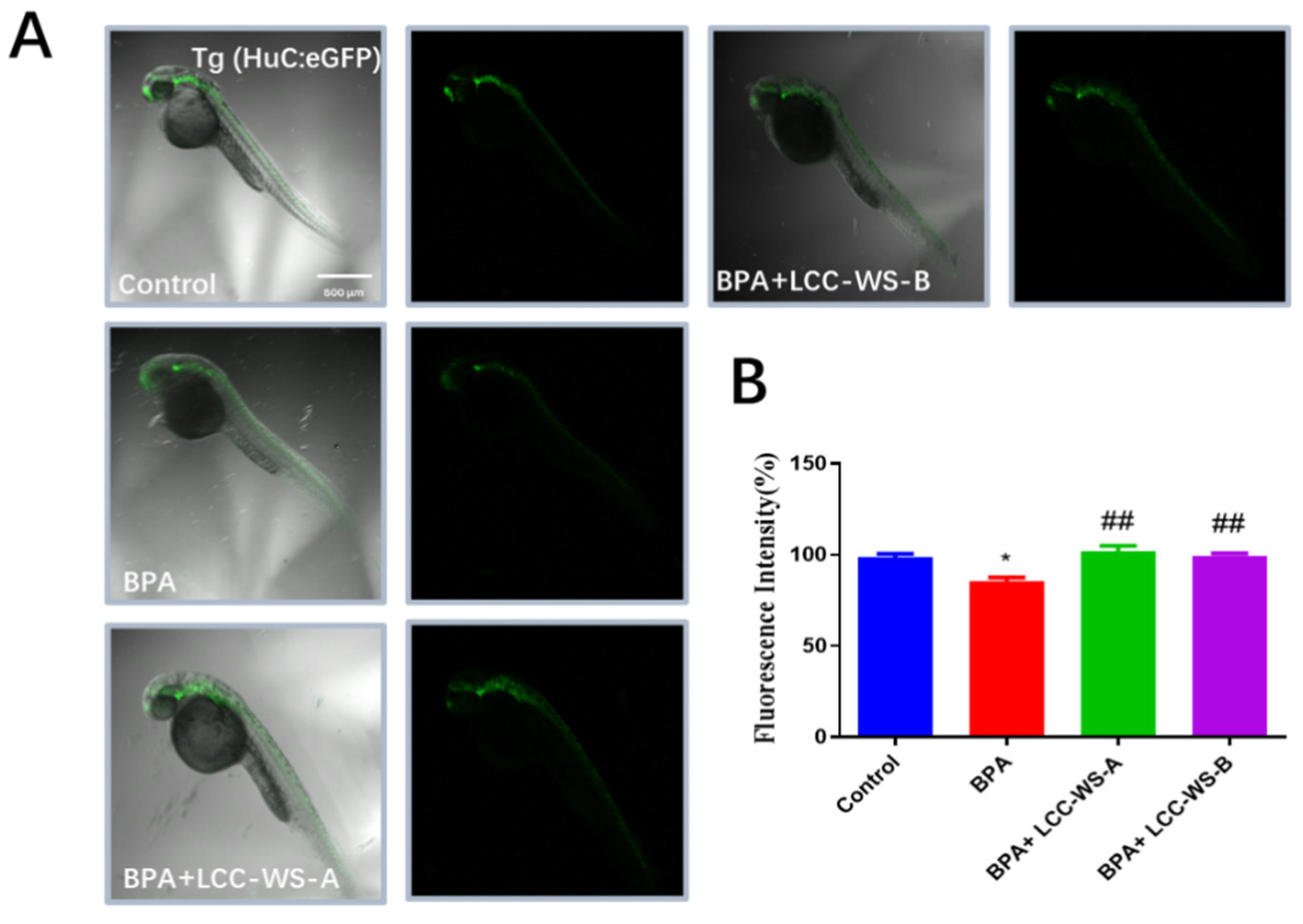
3.6. Effects of LCC-WS-A and LCC-WS-B on Gene Expression in Tg Zebrafish
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fischnaller, M.; Bakry, R.; Bonn, G.K. A simple method for the enrichment of bisphenols using boron nitride. Food Chem. 2016, 194, 149–155. [Google Scholar] [CrossRef]
- Agarwal, S.; Yadav, A.; Tiwari, S.K.; Seth, B.; Chauhan, L.K.; Khare, P.; Ray, R.S.; Chaturvedi, R.K. Dynamin-related protein 1 inhibition mitigates bisphenol a-mediated alterations in mitochondrial dynamics and neural stem cell proliferation and differentiation. J. Biol. Chem. 2016, 291, 15923–15939. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Cao, L.; Wang, F.; Ge, H.; Wu, P.C.; Li, X.W.; Chen, G.H. Accelerated reduction of serum thyroxine and hippocampal histone acetylation links to exacerbation of spatial memory impairment in aged cd-1 mice pubertally exposed to bisphenol-a. Age 2016, 38, 405–418. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, Z.; Zhang, Q.; Song, N.; Cheng, J.; Torres, O.L.; Chen, J.; Zhang, S.; Guo, R. Urinary levels, composition profile and cumulative risk of bisphenols in preschool-aged children from nanjing suburb, china. Ecotoxicol. Environ. Saf. 2019, 172, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Pang, Q.; Li, Y.; Meng, L.; Li, G.; Luo, Z.; Fan, R. Neurotoxicity of bpa, bps, and bpb for the hippocampal cell line (ht-22): An implication for the replacement of bpa in plastics. Chemosphere 2019, 226, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Miyatake, M.; Miyagawa, K.; Mizuo, K.; Narita, M.; Suzuki, T. Dynamic changes in dopaminergic neurotransmission induced by a low concentration of bisphenol-a in neurones and astrocytes. J. Neuroendocrinol. 2006, 18, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Kinch, C.D.; Ibhazehiebo, K.; Jeong, J.H.; Habibi, H.R.; Kurrasch, D.M. Low-dose exposure to bisphenol a and replacement bisphenol s induces precocious hypothalamic neurogenesis in embryonic zebrafish. Proc. Natl. Acad. Sci. USA 2015, 112, 1475–1480. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.K.; Agarwal, S.; Tripathi, A.; Chaturvedi, R.K. Bisphenol-a mediated inhibition of hippocampal neurogenesis attenuated by curcumin via canonical wnt pathway. Mol. Neurobiol. 2016, 53, 3010–3029. [Google Scholar] [CrossRef]
- Khan, J.; Salhotra, S.; Ahmad, S.; Sharma, S.; Abdi, S.A.H.; Banerjee, B.D.; Parvez, S.; Gupta, S.; Raisuddin, S. The protective effect of alpha-lipoic acid against bisphenol a-induced neurobehavioral toxicity. Neurochem. Int. 2018, 118, 166–175. [Google Scholar] [CrossRef]
- Tao, Y.; Wang, T.; Lai, C.; Ling, Z.; Zhou, M.; Yong, Q. The in vitro and in vivo antioxidant and immunomodulatory activity of incomplete degradation products of hemicellulosic polysaccharide (galactomannan) from sesbania cannabina. Front. Bioeng. Biotechnol. 2021, 9, 679558. [Google Scholar] [CrossRef]
- Lin, W.; Yang, J.; Zheng, Y.; Huang, C.; Yong, Q. Understanding the effects of different residual lignin fractions in acid-pretreated bamboo residues on its enzymatic digestibility. Biotechnol. Biofuels 2021, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zheng, L.; Xu, Q.; Xu, L.; Wang, D.; Li, J.; Lu, G.; Huang, C.; Wang, Y. Unveiling the structural properties of water-soluble lignin from gramineous biomass by autohydrolysis and its functionality as a bioactivator (anti-inflammatory and antioxidative). Int. J. Biol. Macromol. 2021, 191, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Lu, G.; Pei, W.; Li, Y.; Zhang, L.; Huang, C.; Jiang, Q. Understanding the relationship between the structural properties of lignin and their biological activities. Int. J. Biol. Macromol. 2021, 190, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Chen, Z.S.; Hu, Y.; Yong, Q. IInsight in the recent application of polyphenols from biomass. Front. Bioeng. Biotechnol. 2021, 9, 753898. [Google Scholar] [CrossRef]
- Dong, H.; Zheng, L.; Yu, P.; Jiang, Q.; Yin, B. Characterization and application of lignin–carbohydrate complexes from lignocellulosic materials as antioxidants for scavenging in vitro and in vivo reactive oxygen species. ACS Sustain. Chem. Eng. 2019, 8, 256–266. [Google Scholar] [CrossRef]
- Hwang, J.H.; Kim, K.J.; Ryu, S.J.; Lee, B.Y. Caffeine prevents lps-induced inflammatory responses in raw264.7 cells and zebrafish. Chem. Biol. Interact. 2016, 248, 1–7. [Google Scholar] [CrossRef]
- Gu, J.; Wang, H.; Zhou, L.; Fan, D.; Shi, L.; Ji, G.; Gu, A. Oxidative stress in bisphenol af-induced cardiotoxicity in zebrafish and the protective role of n-acetyl n-cysteine. Sci. Total Environ. 2020, 731, 139190. [Google Scholar] [CrossRef]
- Wang, L.; Oh, J.Y.; Kim, H.S.; Lee, W.; Cui, Y.; Lee, H.G.; Kim, Y.T.; Ko, J.Y.; Jeon, Y.J. Protective effect of polysaccharides from celluclast-assisted extract of hizikia fusiforme against hydrogen peroxide-induced oxidative stress in vitro in vero cells and in vivo in zebrafish. Int. J. Biol. Macromol. 2018, 112, 483–489. [Google Scholar] [CrossRef]
- Wang, L.; Kim, H.S.; Oh, J.Y.; Je, J.G.; Jeon, Y.J.; Ryu, B. Protective effect of diphlorethohydroxycarmalol isolated from ishige okamurae against uvb-induced damage in vitro in human dermal fibroblasts and in vivo in zebrafish. Food Chem. Toxicol. 2020, 136, 110963. [Google Scholar] [CrossRef]
- Bjorkman, A. Lignin and lignin-carbohydrate complexes. Ind. Eng. Chem. 1957, 49, 1395–1398. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, H.; Fu, Y.; Zhao, Y.; Yuan, Z.; Shao, Z.; Wang, Z.; Qin, M. Short-time hydrothermal treatment of poplar wood for the production of a lignin-derived polyphenol antioxidant. ChemSusChem 2020, 13, 4478–4486. [Google Scholar] [CrossRef]
- Zhou, X.; Xu, Y. Eco-friendly consolidated process for co-production of xylooligosaccharides and fermentable sugars using self-providing xylonic acid as key pretreatment catalyst. Biotechnol. Biofuels 2019, 12, 272. [Google Scholar] [CrossRef]
- Zhang, L.; Gellerstedt, G. Quantitative 2d hsqc nmr determination of polymer structures by selecting suitable internal standard references. Magn. Reson. Chem. 2007, 45, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.; Abril, N.; Morales-Prieto, N.; Monterde, J.; Ayala, N.; Lora, A.; Moyano, R. Hypothalamic-pituitary-ovarian axis perturbation in the basis of bisphenol a (bpa) reproductive toxicity in female zebrafish (danio rerio). Ecotoxicol. Environ. Saf. 2018, 156, 116–124. [Google Scholar] [CrossRef]
- Kim, S.S.; Hwang, K.S.; Yang, J.Y.; Chae, J.S.; Kim, G.R.; Kan, H.; Jung, M.H.; Lee, H.Y.; Song, J.S.; Ahn, S.; et al. Neurochemical and behavioral analysis by acute exposure to bisphenol a in zebrafish larvae model. Chemosphere 2020, 239, 124751. [Google Scholar] [CrossRef]
- Yun, J.; Wei, L.; Li, W.; Gong, D.; Qin, H.; Feng, X.; Li, G.; Ling, Z.; Wang, P.; Yin, B. Isolating high antimicrobial ability lignin from bamboo kraft lignin by organosolv fractionation. Front. Bioeng. Biotechnol. 2021, 9, 683796. [Google Scholar] [CrossRef]
- Bjrkman, A. Studies on finely divided wood. Part 3. Extraction of lignin-carbohydrate complexes with neutral solvents. Sven Papp. 1957, 60, 243–251. [Google Scholar]
- Jiang, B.; Zhang, Y.; Guo, T.; Zhao, H.; Jin, Y. Structural characterization of lignin and lignin-carbohydrate complex (lcc) from ginkgo shells (ginkgo biloba l.) by comprehensive nmr spectroscopy. Polymers 2018, 10, 736. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Wan, G.; Li, M.; Jiang, H.; Wang, S.; Min, D. Impact of bagasse lignin-carbohydrate complexes structural changes on cellulase adsorption behavior. Int. J. Biol. Macromol. 2020, 162, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Altıok, E.; Bayçın, D.; Bayraktar, O.; Ülkü, S. Isolation of polyphenols from the extracts of olive leaves (olea europaea l.) by adsorption on silk fibroin. Sep. Purif. Technol. 2008, 62, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.Y.; Zhang, Q.; Li, Y.F.; Dong, L.L.; Liu, S.L. Optimization of ultrasound extraction of alisma orientalis polysaccharides by response surface methodology and their antioxidant activities. Carbohydr. Polym. 2015, 119, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.K.; El-Sammad, N.M.; Mousa, A.M.; Mohammed, M.H.; Farrag, A.E.R.H.; Hashim, A.N.E.; Werner, V.; Lindequist, U.; Nawwar, E.M. Hypoglycemic and antioxidant activities of caesalpinia ferrea martius leaf extract in streptozotocin-induced diabetic rats. Asian Pac. J. Trop. Biomed. 2015, 5, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Zhang, J.; Chen, Y.; Wang, H.; Guo, M.; Wang, L.; Wang, Z.; Wu, S.; Shi, L.; Gu, A.; et al. Neurobehavioral effects of bisphenol s exposure in early life stages of zebrafish larvae (danio rerio). Chemosphere 2019, 217, 629–635. [Google Scholar] [CrossRef]
- Gu, J.; Wu, J.; Xu, S.; Zhang, L.; Fan, D.; Shi, L.; Wang, J.; Ji, G. Bisphenol f exposure impairs neurodevelopment in zebrafish larvae (danio rerio). Ecotoxicol. Environ. Saf. 2020, 188, 109870. [Google Scholar]
- Shiva, M.; Gautam, A.K.; Verma, Y.; Shivgotra, V.; Doshi, H.; Kumar, S. Association between sperm quality, oxidative stress, and seminal antioxidant activity. Clin. Biochem. 2011, 44, 319–324. [Google Scholar] [CrossRef]
- Brosamle, C.; Halpern, M.E. Characterization of myelination in the developing zebrafish. Glia 2002, 39, 47–57. [Google Scholar] [CrossRef]
- Muller, C.; Bauer, N.M.; Schafer, I.; White, R. Making myelin basic protein -from mrna transport to localized translation. Front. Cell. Neurosci. 2013, 7, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, H.T.; Porton, B.; Czernik, A.J.; Feng, J.; Yiu, G.; Haring, M.; Benfenati, F.; Greengard, P. A third member of the synapsin gene family. Proc. Natl. Acad. Sci. USA 1998, 95, 4667–4672. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preparation | Lignin (%) | Cellulose (%) | Hemicellulose (%) | Molecular Weight | Polyphenols (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Xylan | Arabinan | Galactan | Mannan | Mw (g·mol−1) | Mn (g·mol−1) | PDI | ||||
| LCC-WS-A | 42.5 | 3.4 | 40.5 | 1.9 | 0.2 | 0.1 | 17,600 | 7890 | 2.2 | 3.3 |
| LCC-WS-B | 64.2 | 1.6 | 23.5 | 0.8 | 0 | 0 | 9210 | 4520 | 2.0 | 3.1 |
| Characteristics | LCC-WS-A | LCC-WS-B |
|---|---|---|
| Lignin interunit linkages | ||
| β-O-4 aryl ethers (A) | 21.9 | 14.1 |
| Resinols (B) | 5.5 | - |
| Phenylcoumarans, β-5 (C) | 2.3 | - |
| S:G ratio a | 0.8 | 1.9 |
| LCC linkages | ||
| Benzyl ether (BE) | 2.9 | - |
| Phenyl glycoside (PhGlc) | 13.5 | 4.3 |
| Est b | 6.3 | 5.9 |
| Total c | 22.7 | 10.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, J.; Guo, M.; Zheng, L.; Yin, X.; Zhou, L.; Fan, D.; Shi, L.; Huang, C.; Ji, G. Protective Effects of Lignin-Carbohydrate Complexes from Wheat Stalk against Bisphenol a Neurotoxicity in Zebrafish via Oxidative Stress. Antioxidants 2021, 10, 1640. https://doi.org/10.3390/antiox10101640
Gu J, Guo M, Zheng L, Yin X, Zhou L, Fan D, Shi L, Huang C, Ji G. Protective Effects of Lignin-Carbohydrate Complexes from Wheat Stalk against Bisphenol a Neurotoxicity in Zebrafish via Oxidative Stress. Antioxidants. 2021; 10(10):1640. https://doi.org/10.3390/antiox10101640
Chicago/Turabian StyleGu, Jie, Min Guo, Liping Zheng, Xiaogang Yin, Linjun Zhou, Deling Fan, Lili Shi, Caoxing Huang, and Guixiang Ji. 2021. "Protective Effects of Lignin-Carbohydrate Complexes from Wheat Stalk against Bisphenol a Neurotoxicity in Zebrafish via Oxidative Stress" Antioxidants 10, no. 10: 1640. https://doi.org/10.3390/antiox10101640